

Abstract
BACKGROUND AND PURPOSE
In spite of its widespread clinical application, there is little information on the cellular cardiac effects of the antidiabetic drug rosiglitazone in larger experimental animals. In the present study therefore concentration-dependent effects of rosiglitazone on action potential morphology and the underlying ion currents were studied in dog hearts.
EXPERIMENTAL APPROACH
Standard microelectrode techniques, conventional whole cell patch clamp and action potential voltage clamp techniques were applied in enzymatically dispersed ventricular cells from dog hearts.
KEY RESULTS
At concentrations ≥10 µM rosiglitazone decreased the amplitude of phase-1 repolarization, reduced the maximum velocity of depolarization and caused depression of the plateau potential. These effects developed rapidly and were readily reversible upon washout. Rosiglitazone suppressed several transmembrane ion currents, concentration-dependently, under conventional voltage clamp conditions and altered their kinetic properties. The EC50 value for this inhibition was 25.2 ± 2.7 µM for the transient outward K+ current (Ito), 72.3 ± 9.3 µM for the rapid delayed rectifier K+ current (IKr) and 82.5 ± 9.4 µM for the L-type Ca2+ current (ICa) with Hill coefficients close to unity. The inward rectifier K+ current (IK1) was not affected by rosiglitazone up to concentrations of 100 µM. Suppression of Ito, IKr, and ICa was confirmed also under action potential voltage clamp conditions.
CONCLUSIONS AND IMPLICATIONS
Alterations in the densities and kinetic properties of ion currents may carry serious pro-arrhythmic risk in case of overdose with rosiglitazone, especially in patients having multiple cardiovascular risk factors, like elderly diabetic patients.
LINKED ARTICLE
This article is commented on by Hancox, pp. 496–498 of this issue. To view this commentary visit http://dx.doi.org/10.1111/j.1476-5381.2011.01281.x
Keywords: antidiabetic agents, rosiglitazone, dog cardiomyocytes, action potential, ion currents
Introduction
Rosiglitazone is a thiazolidinedione oral hypoglycemic agent active in both diabetic animal models and patients with type 2 diabetes (Quinn et al., 2008; Doshi et al., 2010). Rosiglitazone, similarly to other thiazolidinediones, like troglitazone or pioglitazone, is a high affinity ligand for the peroxisome proliferator-activated receptor-γ, which is responsible for the insulin-sensitizing action of the compound (Berger et al., 1996; Welters et al., 2004). Beyond improving glycemic control, rosiglitazone and pioglitazone have been shown to exert some cardiovascular benefits (Khandoudi et al., 2002; Cuzzocrea et al., 2003). In contrast, several large clinical studies reported that thiazolidinedione therapy was associated with cardiovascular complications (Krentz, 2009; Kaul et al., 2010). Increased risk of congestive heart failure was observed with the use of both pioglitazone and rosiglitazone (Gerstein et al., 2006; McGuire and Inzucchi, 2008; DREAM Trial Investigators, 2008), while the increased risk of acute myocardial infarction and mortality appeared to be limited to rosiglitazone (Home et al., 2005; Nissen and Wolski, 2007). These latter risks of rosiglitazone therapy have not been confirmed by other clinical studies (Rosen, 2007; Mannucci et al., 2010).
In addition to the known cardiovascular side effects of rosiglitazone, the drug increased the propensity of ventricular fibrillation in pigs, an effect attributed to inhibition of the cardiac ATP-sensitive K+ channel (Lu et al., 2008). However, no direct evidence based on voltage clamp analysis is available to exclude the contribution of other ion currents. Furthermore, rosiglitazone was shown to block a wide variety of non-cardiac ion channels, including neuronal Ca2+ channels, epithelial Na+ channels, ATP-sensitive K+ channels, delayed rectifier K+ channels and L-type Ca2+ channels in aortic smooth muscle cells (Knock et al., 1999; Mishra and Aaronson, 1999; McKay et al., 2000; Pancani et al., 2009; Pavlov et al., 2009). In contrast to the lack of data on cardiac cells with rosiglitazone, another thiazolidinedione derivative troglitazone was shown to effectively block L-type Ca2+ current (ICa) in ventricular myocytes of guinea pigs (Nakajima et al., 1999; Katoh et al., 2000) and rats (Arikawa et al., 2002; 2004;), while in rabbit ventricular cells Na+, Ca2+ and K+ currents were suppressed by the drug (Ikeda and Watanabe, 1998).
In absence of relevant voltage clamp data on the effects of rosiglitazone in mammalian cardiac preparations, we aimed to study the concentration-dependent effects of rosiglitazone on action potential morphology and the underlying ion currents in isolated canine ventricular cardiomyocytes. Canine ventricular cells were chosen because their electrophysiological properties are believed to be very similar to those of human regarding the distribution and kinetic properties of transmembrane ion currents (Szabóet al., 2005; Szentandrássy et al., 2005).
Methods
Isolation of single ventricular myocytes from dog heart
All animal care and experimental procedures conformed to ‘Principles of laboratory animal care’ (NIH publication N° 85–23, revised 1985) and were approved by the local ethical committee. Adult beagle dogs of either sex were anaesthetized with intravenous injections of 10 mg·kg−1 ketamine hydrochloride (Calypsol, Richter Gedeon, Budaspest, Hungary) + 1 mg·kg−1 xylazine hydrochloride (Sedaxylan, Eurovet Animal Health BV, Bladel, The Netherlands). The hearts were quickly removed and placed in Tyrode solution. Single myocytes were obtained by enzymatic dispersion using the segment perfusion technique (Szabóet al., 2005). Briefly, a wedge-shaped section of the ventricular wall supplied by the left anterior descending coronary artery was dissected, cannulated and perfused with oxygenized Tyrode solution containing: NaCl 144, KCl 5.6, CaCl2 2.5, MgCl2 1.2, HEPES 5, and dextrose 11 mM at pH = 7.4. Perfusion was continued until all blood was washed from the coronary system and then switched to a nominally Ca2+-free Joklik solution (Minimum Essential Medium Eagle, Joklik Modification, Sigma-Aldrich Co., St. Louis, MO, USA) for 5 min. This was followed by 30 min perfusion with Joklik solution supplemented with 1 mg·mL−1 collagenase (Type II., Worthington Biochemical Co., Lakewood, NJ, USA) and 0.2% bovine serum albumin (Fraction V., Sigma-Aldrich Co.) containing 50 µM Ca2+. Portions of the left ventricular wall were cut into small pieces and the cell suspension obtained at the end of the procedure, predominantly from the mid myocardial region of the left ventricle, was washed with Joklik solution. Finally the Ca2+ concentration was gradually restored to 2.5 mM. The cells were stored in minimum essential medium until use.
Recording of action potentials
All electrophysiological measurements were performed at 37°C. The rod-shaped viable cells showing clear striations were sedimented in a plexiglass chamber allowing continuous superfusion with oxygenated Tyrode solution. Transmembrane potentials were recorded using 3 M KCl filled sharp glass microelectrodes having tip resistance between 20 and 40 MΩ. These electrodes were connected to the input of an Axoclamp-2B amplifier (Axon Instruments Inc., Foster City, CA, USA). The cells were paced through the recording electrode at a steady cycle length of 1 s using 1 ms wide rectangular current pulses with 120% threshold amplitude. Because the cytosol was not dialysed, time-dependent changes in action potential duration were negligible for at least 60 min under these experimental conditions (Horváth et al., 2006).
Concentration-dependent effects of rosiglitazone were determined in a cumulative manner by applying increasing concentrations of the drug between 1 and 100 µM. Each concentration was superfused for 3 min and the washout usually lasted for 10 min. These incubation and washout periods were sufficient to develop steady-state drug effects and practically full reversal. As the stimulus threshold was gradually increased during the cumulative application of rosiglitazone, the amplitude of stimuli had to be increased accordingly. Action potentials were digitized at 200 kHz using Digidata 1200 A/D card (Axon Instruments Inc.) and stored for later analysis.
Conventional voltage clamp
The cells were superfused with oxygenated Tyrode solution. Suction pipettes, fabricated from borosilicate glass, had a tip resistance of 2 MΩ after filling with pipette solution containing (in mM) K-aspartate, 100; KCl, 45; MgCl2, 1; HEPES, 5; EGTA, 10; K-ATP, 3, or alternatively, KCl, 110; KOH, 40; HEPES, 10; EGTA, 10; TEACl, 20; K-ATP, 3, when measuring potassium or calcium currents respectively (pH = 7.2 in both cases). Membrane currents were recorded with the Axopatch-2B amplifier using the whole cell configuration of the patch clamp technique (Hamill et al., 1981). After establishing a high (1–10 GΩ) resistance seal by gentle suction, the cell membrane beneath the tip of the electrode was disrupted by further suction or by applying 1.5 V electrical pulses for 1 ms. The series resistance was typically 4–8 MΩ before compensation (usually 50–80%). Experiments were discarded when the series resistance was high or substantially increasing during the measurement. Outputs from the clamp amplifier were digitized at 100 kHz under software control (pClamp 6.0, Axon Instruments Inc.). Ion currents were normalized to cell capacitance, determined in each cell using short hyperpolarizing pulses from −10 to −20 mV. Cell capacitance was 131 ± 5 pF in the average of 25 myocytes. The experimental protocol for each measurement is described where pertinent in the Results section. Concentration-dependent effects of rosiglitazone were determined in a cumulative manner by applying increasing concentrations of the drug between 1 and 300 µM.
Action potential voltage clamp
Using the whole cell configuration of the patch clamp technique, action potentials were recorded in current clamp mode from the myocytes superfused with Tyrode solution. The pipette solution was identical to that used for potassium current measurement under conventional voltage clamp conditions. The cells were continuously paced through the recording electrode at a steady stimulation frequency of 1 Hz so a 1–2 ms gap between the stimulus artefact and the upstroke of the action potential could occur. Ten subsequent action potentials were recorded from each cell, which were analysed online. One of these action potentials, having an APD90 value closest to the average of the 10 recorded action potentials, was delivered to the same cell at the identical frequency as command voltage after switching the amplifier to voltage clamp mode. The current trace obtained under these conditions is a horizontal line positioned at the zero level except for the very short segment corresponding to the action potential upstroke (Fischmeister et al., 1984). Rosiglitazone was applied in a cumulative manner using concentrations of 1, 10 and 100 µM. The profile of the ion currents blocked by rosiglitazone was determined by subtracting the post-drug curve from the pre-drug curve. This procedure resulted in composite current profiles containing three distinct current peaks: an early outward peak for the transient outward K+ current (Ito), an inward peak for the ICa and a late outward peak for the rapid delayed rectifier K+ current (IKr) (Bányáz et al., 2007).
Statistics
Results are expressed as mean ± SEM values. Statistical significance of differences was evaluated using one-way anova followed by Student's t-test. Differences were considered significant when P < 0.05. For each type of experiment, both the number of myocytes studied and the number of animals used are specified.
Materials
Rosiglitazone was freshly diluted with Tyrode solution to final concentration on the day of experiment. Rosiglitazone was purchased from Molekula Ltd (Shaftesbury, UK); all other drugs were obtained from Sigma-Aldrich Co. Drug and molecular target nomenclature follows Alexander et al. (2009).
Results
Effect of rosiglitazone on action potential configuration
Rosiglitazone treatment caused complex, concentration-dependent, changes in action potential morphology in canine ventricular myocytes (eight cells from four dogs), paced at a constant frequency of 1 Hz, including reduction of the amplitude of early (phase-1) repolarization, the maximum velocity of depolarization (Vmax), and depression of the plateau potential (Figure 1A). These effects of rosiglitazone were significant at concentrations of 10 µM and higher (Figure 1B and C). Action potential duration was little affected by rosiglitazone; however, APD50 was significantly shortened by 30 µM, while APD90 was lengthened by 100 µM of rosiglitazone (Figure 1D). In spite of the reduction of Vmax, the amplitude of action potential was not decreased by rosiglitazone. In contrast, it was significantly increased in the presence of 100 µM rosiglitazone (Figure 1E). All these effects of rosiglitazone developed rapidly (within 2–3 min) and were readily reversible after superfusion with rosiglitazone-free Tyrode solution for 10 min.
Figure 1.
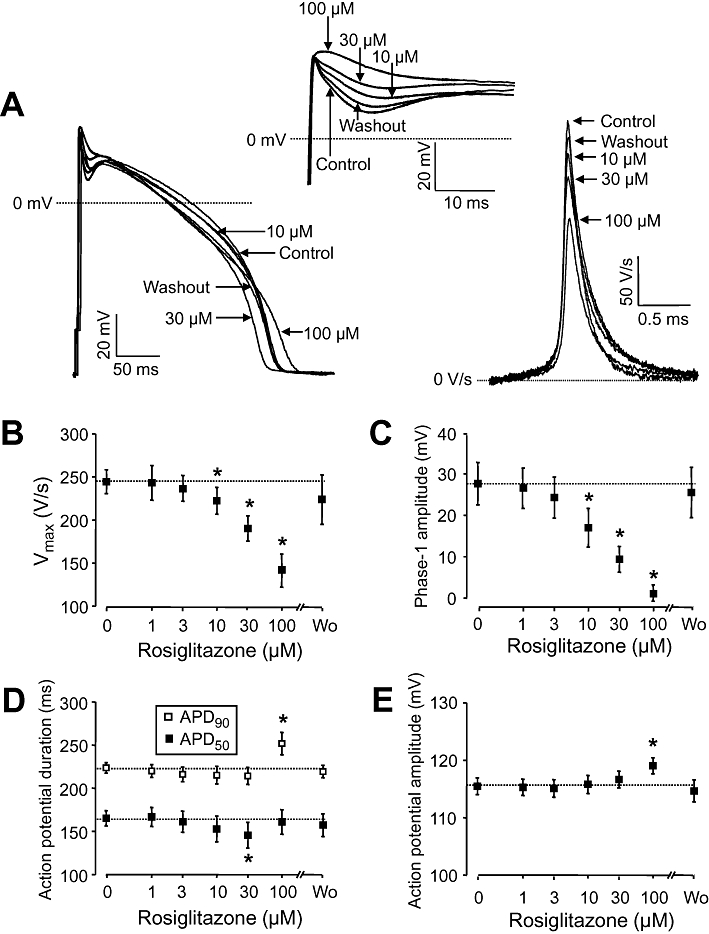
(A) Representative set of superimposed action potentials showing the cumulative concentration-dependent effects of rosiglitazone on action potential configuration. Early events of the action potential are enlarged in the inset, the first time derivatives of action potential upstrokes are depicted on the right. The action potentials were superimposed so as to match their upstrokes. (B–E) Cumulative concentration-dependent effects of rosiglitazone on the maximum rate of depolarization (Vmax), amplitude of phase-1 repolarization, action potential duration measured at 50% (APD50) and 90% (APD90) level of repolarization, and action potential amplitude respectively. Phase-1 amplitude was determined as a difference of overshoot potential and the deepest point of the incisura. Each concentration of rosiglitazone was superfused for 3 min, the washout (Wo) lasted for 10 min. Symbols and bars represent mean ± SEM values of eight myocytes, obtained from four dogs. *P < 0.05, significant changes from pre-drug control values, which are also indicated by dotted lines.
Effect of rosiglitazone on the density and kinetics of cardiac ion currents measured by conventional voltage clamp
In these experiments, performed under conventional voltage clamp conditions, cumulative concentration-dependent drug effects were monitored between 1 and 300 µM, increasing the concentration of rosiglitazone usually in steps of 0.5 log units. Kinetic properties of the channel gating were studied at concentrations, which were close to the half effective blocking concentration of rosiglitazone on the given ion current.
Ito was activated by depolarizations to +50 mV arising from the holding potential of −80 mV and having duration of 200 ms. Before each test pulse, a short (5 ms) depolarization to −40 mV was applied in order to inactivate the fast Na+ current (INa), while ICa and IKr were blocked by 5 µM nifedipine and 1 µM E4031 respectively. Ito was suppressed by rosiglitazone in a concentration-dependent manner in the five myocytes studied (each from a different animal). This effect of rosiglitazone was statistically significant at concentrations ≥10 µM, the EC50 value was 25.2 ± 2.7 µM, and the Hill coefficient 1.27 ± 0.19 (Figure 2A and B). The effect of rosiglitazone developed rapidly (within 2–3 min) and was fully reversible upon washout (Figure 2C). Rosiglitazone altered the gating properties of Ito. The current inactivated as a sum of a faster and a slower component. The amplitude of both components was significantly decreased in the presence of 30 µM rosiglitazone. The fast time constant was not modified, while the slow time constant was doubled by rosiglitazone (Figure 2D). Activation of Ito required larger depolarizations in the presence of rosiglitazone and the activation threshold was shifted from −20 to +20 mV (Figure 2E), while no significant change was observed in the voltage-dependence of inactivation (Figure 2F).
Figure 2.
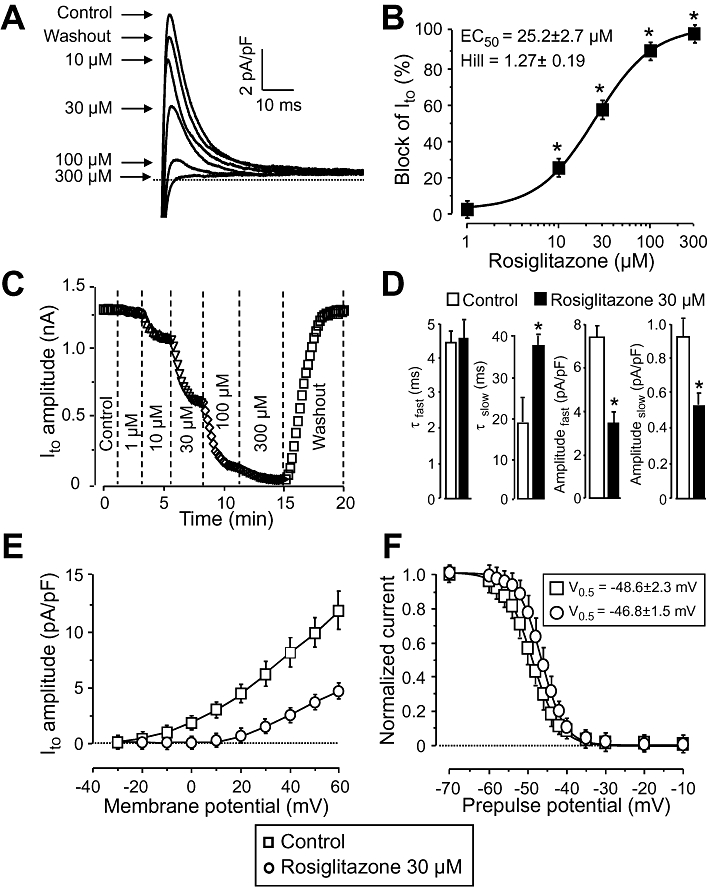
Effect of rosiglitazone on Ito. (A), (B): Cumulative concentration-dependent effects of rosiglitazone on Ito measured under conventional voltage clamp conditions. Representative superimposed Ito current traces (A) recorded before and after superfusion with increasing concentrations of rosiglitazone, and the dose–response curve (B) obtained for Ito blockade in five cells, each from a different animal, including the results of the Hill plot. (C): Time course of development and reversibility of the effect of rosiglitazone on Ito measured in a representative cell. (D)–(F): Effect of 30 µM rosiglitazone on kinetic properties of Ito studied in five myocytes, each from a different dog. (D): Time course of inactivation of Ito. The current decay was fitted as a sum of two (fast and slow) exponential components. (E): Current–voltage relationship obtained for Ito. Peak values of Ito were plotted against the respective test potential shown on abscissa. (F): Effect of rosiglitazone on the voltage-dependence of steady-state inactivation of Ito. Test depolarizations to +50 mV were preceded by a set of prepulses clamped to various voltages between −70 and +10 mV. Peak currents measured after these prepulses were normalized to the peak current measured after the −70 mV prepulse and plotted against the respective prepulse potential. Solid lines were obtained by fitting data to the two-state Boltzmann function. Symbols, columns and bars are means ± SEM. *P < 0.05, significant changes from control.
IKr was activated by 1 s long depolarizing pulses to +50 mV, from the holding potential of −40 mV. IKr was assessed as tail current amplitudes recorded following repolarization to the holding potential. ICa and the slow delayed rectifier K+ current were suppressed by 5 µM nifedipine and 1 µM HMR-1556 respectively. As shown in Figure 3A and B, the amplitudes of the IKr current tails were progressively decreased by increasing concentrations of rosiglitazone. The EC50 value and Hill coefficient were estimated to be 72.3 ± 9.3 µM and 0.91 ± 0.1, respectively, as the mean of five myocytes, each derived from a different animal (Figure 3B). Similarly to results observed with Ito, suppression of the IKr tails developed rapidly and was fully reversible (Figure 3C). Relaxation of IKr current tails (deactivation) followed biexponential kinetics. Although the time constants of both components decreased, this effect failed to reach the level of statistical significance (P > 0.05). In contrast, the amplitudes of both components were significantly reduced by 100 µM rosiglitazone. Voltage-dependence of activation of IKr was moderately shifted towards more negative potentials by 100 µM rosiglitazone: the half-activation voltage (V0.5) of IKr was shifted from −4.8 ± 3.2 to −10.2 ± 1.3 mV (five cells, each from a different dog, P < 0.05). Due to this shift of activation, the block apparently increased with increasing depolarizations to more positive voltages, it was negligible at membrane potentials ≤−5 mV, while it represented close to half of control current amplitude at voltages more positive than +10 mV (Figure 3E). No change in the monoexponential time constant of activation of IKr, determined by using the tail envelope test, was observed in the presence of 100 µM rosiglitazone (Figure 3F).
Figure 3.
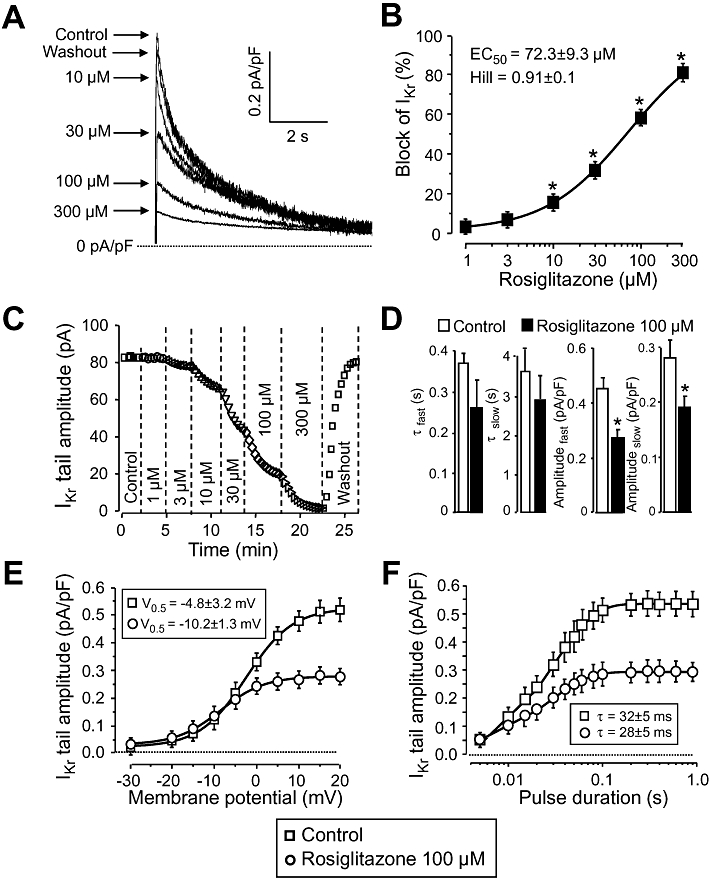
Effect of rosiglitazone on IKr. (A), (B): Concentration-dependent effects of rosiglitazone on IKr measured under conventional voltage clamp conditions. Representative superimposed IKr tail current traces (A) recorded before and after superfusion with increasing concentrations of rosiglitazone, and the dose–response curve (B) obtained for IKr blockade in five cells, each from a different animal, including the results of the Hill plot. (C): Time course of development and reversibility of the effect of rosiglitazone on IKr recorded from a representative cell. (D)–(F): Effect of 100 µM rosiglitazone on kinetic properties of IKr studied in five myocytes, each obtained from a different dog. (D): Time course of deactivation of IKr. Decay of tail currents was fitted as a sum of two (fast and slow) exponential components. (E): Voltage-dependence of activation of IKr was determined by varying the potential for IKr activation as indicated on the abscissa. The results were fitted to the two-state Boltzmann function denoted by solid lines. (F): Time constant of activation was determined by monoexponential fitting of data obtained using the tail envelope test (applying depolarizations to +50 mV and determining tail current amplitudes at −40 mV). Durations of these depolarizing pulses are displayed on the abscissa. Symbols, columns and bars are mean values ± SEM. *P < 0.05, significant changes from control.
ICa was recorded at +5 mV using 400 ms long depolarizations arising from the holding potential of −40 mV. In these experiments, Tyrode solution was supplemented with 3 mM 4-aminopyridine, 1 µM E4031 and 1 µM HMR-1556 in order to block K+ currents. Rosiglitazone blocked ICa in a concentration-dependent manner causing statistically significant block at concentrations ≥10 µM. The EC50 value and Hill coefficient were 82.5 ± 9.4 µM and 0.82 ± 0.08, respectively, in the six myocytes obtained from five dogs (Figure 4A and B). As indicated by Figure 4C, ICa was only partially reversed during the 10 min period of washout following rosiglitazone treatment. Rosiglitazone altered slightly the gating properties of ICa as well. This current inactivated as a sum of two exponential components and no significant changes in the time constants were observed. Although amplitudes of both the fast and slow components were decreased by 100 µM rosiglitazone, suppression of the slow component was more pronounced. (Figure 4D). Rosiglitazone had no effect on the current–voltage relationship obtained for ICa by plotting the amplitudes of the current against the respective test potentials (Figure 4E). In contrast, steady-state inactivation of ICa was enhanced by 100 µM rosiglitazone. The half inactivation voltage (V0.5) was significantly shifted towards more negative potentials (from −12.9 ± 0.3 mV to −15.5 ± 0.3 mV, P < 0.05, five cells, each from a different animal, Figure 4F).
Figure 4.
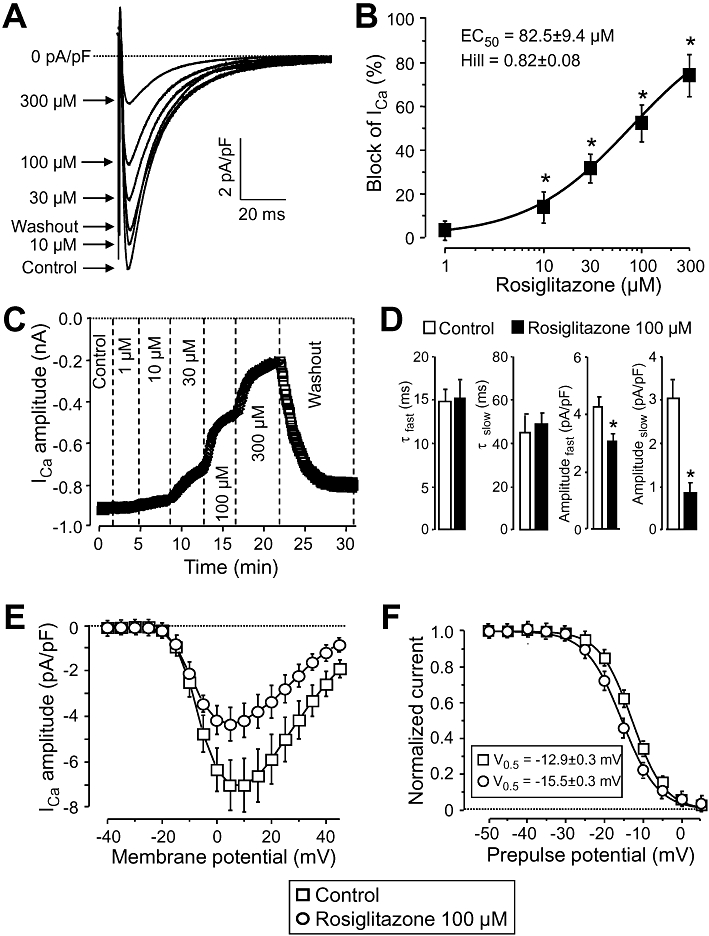
Effect of rosiglitazone on ICa. (A), (B): Cumulative concentration-dependent effects of rosiglitazone on ICa measured under conventional voltage clamp conditions. Representative superimposed ICa current traces (A) recorded before and after superfusion with increasing concentrations of rosiglitazone, and the dose–response curve (B) obtained for ICa blockade in six cells from five dogs, including the results of the Hill plot. (C): Time course of development and reversibility of the effect of rosiglitazone on ICa measured in a representative cell. (D)–(F): Effect of 100 µM rosiglitazone on kinetic properties of ICa studied in five myocytes, derived from five different animals. (D): Time course of inactivation of ICa. The current decay was fitted as a sum of two (fast and slow) exponential components. (E): Current–voltage relationship obtained for ICa. Amplitudes of ICa were plotted against the respective test potential shown on abscissa. (F): Effect of rosiglitazone on the voltage-dependence of steady-state inactivation of ICa. Test depolarizations to +5 mV were preceded by a set of prepulses clamped to various voltages between −50 and +5 mV. Peak currents measured after these prepulses were normalized to the peak current measured after the −50 mV prepulse and plotted against the respective prepulse potential. Solid lines were obtained by fitting data to the two-state Boltzmann function. Symbols, columns and bars are means ± SEM. *P < 0.05, significant changes from control.
The inward rectifier K+ current (IK1) was studied by applying hyperpolarizations to −135 mV from the holding potential of −40 mV. The steady-state current was determined 400 ms after the beginning of the pulse. As it is shown in Figure 5, IK1 was not significantly modified by rosiglitazone up to the concentration of 100 µM in the four cells challenged, each obtained from a different dog. Above this concentration (at 300 µM) a small but fully reversible suppression of IK1 was observed.
Figure 5.
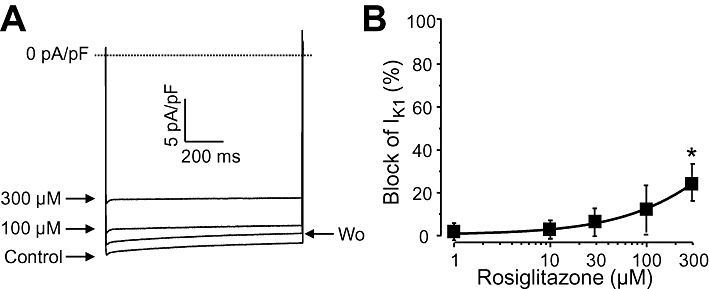
Cumulative concentration-dependent effects of rosiglitazone on IK1 measured under conventional voltage clamp conditions. Representative superimposed IK1 current traces (A) recorded before and after superfusion with increasing concentrations of rosiglitazone, and the dose–response curve (B) obtained for IK1 blockade in four cells, each from a different dog. Symbols and bars are means ± SEM. *P < 0.05, significant changes from control.
Effect of rosiglitazone on ion currents under action potential clamp conditions
The profile of an ion current may be markedly different when comparing under conventional voltage clamp and action potential clamp conditions. An advantage of the action potential clamp technique is that the effect of any drug on the net membrane current can be recorded allowing thus to monitor drug effects simultaneously on more than one ion current. Furthermore, this technique enables us to record true current profiles flowing during an actual cardiac action potential. Of course, in the case of a drug acting on more than one ion current, such as rosiglitazone, a series of peaks can be detected on the current trace, each of them corresponding to the fingerprint of an individual ion current. Accordingly, the early outward current peak, shown in Figure 6, arises when Ito is suppressed, while the inward deflection indicates a blockade of ICa. The late outward current peak, coincident with terminal repolarization of the canine action potential, is a mixture of IKr plus IK1. In our case, however, it is likely caused by pure IKr blockade, because – as we have previously shown – IK1 was not affected by 100 µM rosiglitazone. As demonstrated in Figure 6, rosiglitazone suppressed Ito, IKr and ICa under action potential voltage clamp conditions in a concentration-dependent and largely reversible manner – in line with results of conventional voltage clamp experiments. The amplitudes of the three current peaks (early outward, inward and late outward) are presented in Figure 7 as the mean of five myocytes, each obtained from a different dog.
Figure 6.
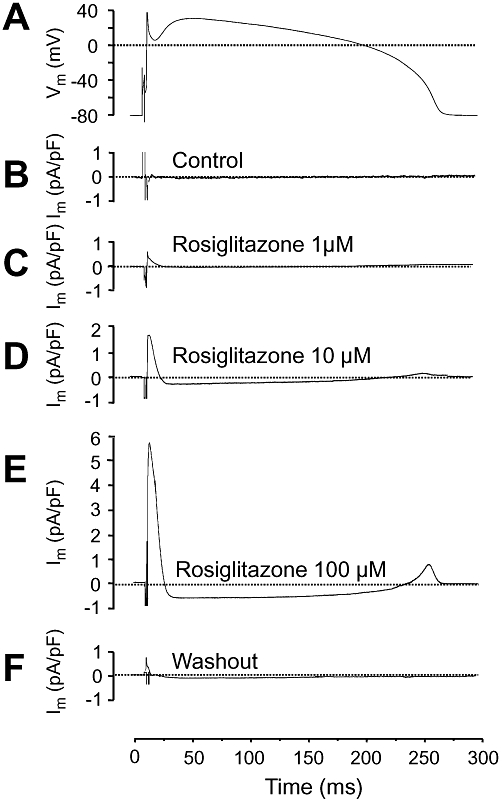
Effects of 1, 10 and 100 µM rosiglitazone on ion currents under action potential voltage clamp conditions. Representative records of a command signal (A), and the underlying current traces obtained before (B), in the presence of (C)–(E), and after washout of rosiglitazone (F). Because ion currents were obtained from the same cell that provided the command action potential, the pre-drug current record was a horizontal line at the zero level. The post-drug difference currents have been inverted in order to present the currents with their conventional polarity. Dotted lines indicate zero current level for each trace except for (A), where it represents zero voltage.
Figure 7.
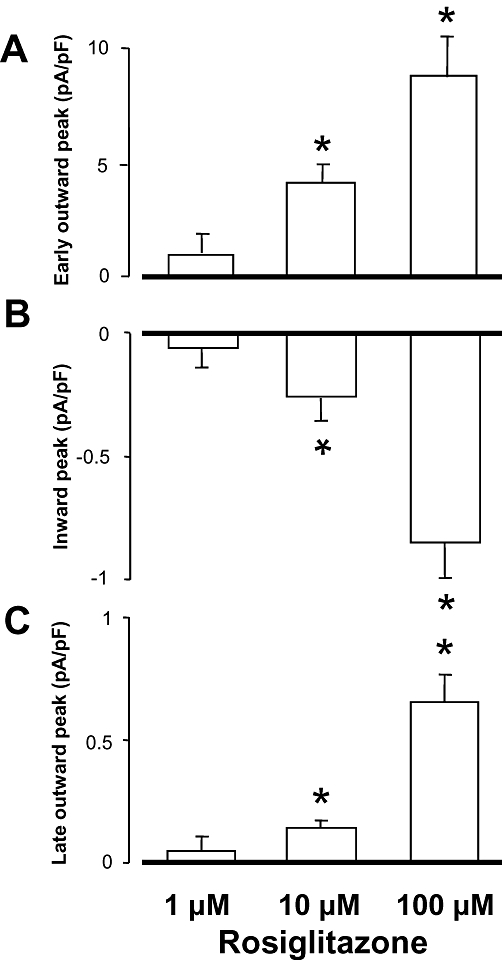
Effects of 1, 10 and 100 µM rosiglitazone on the early outward current peak, indicator of Ito (A), the inward current peak, indicator of ICa (B), and the late outward current peak, indicator predominantly of IKr (C), obtained under action potential voltage clamp conditions in five myocytes, each from a different animal. Columns and bars are means ± SEM. *P < 0.05, significant changes from control.
Discussion
Effects of rosiglitazone on action potential morphology are in line with effects on the underlying ion currents
Effects of rosiglitazone on native cardiac ion currents were first analysed in this study. The results revealed that rosiglitazone suppressed several ion currents, including INa, Ito, IKr and ICa, in a concentration-dependent manner with the concomitant alterations in the configuration of the action potential. These changes can be associated with suppression of various ion currents with concomitant changes in channel gating. For instance, the rosiglitazone-induced decrease in phase-1 repolarization may be due to reduction of Ito. Similarly, the depression of the plateau, observed in the presence of rosiglitazone, may be a consequence of inhibition of Ca2+ and Na+ currents. Concentration-dependent blockade of Ito and ICa have been demonstrated by our present voltage clamp experiments, while the observed suppression of Vmax is believed to be a good indicator of INa blockade (Strichartz and Cohen, 1978).
In spite of the multiple actions of rosiglitazone on cardiac ion channels, action potential duration was little affected by rosiglitazone, except for the moderate reduction of APD50 at 30 µM and lengthening of APD90 at 100 µM concentration. This apparently paradoxical behaviour may be related to differences in blocking potencies on the various ion currents. On the other hand, the lack of marked effects of rosiglitazone on action potential duration suggests that inhibition of inward (window INa and ICa) and outward (IKr and Ito) currents are relatively well compensated. This suggestion is strongly supported by the action potential voltage clamp records shown in Figure 6. Also, suppression of Vmax is usually accompanied with reduction of action potential amplitude. This effect was not observed with rosiglitazone – in contrast – action potential amplitude was significantly increased by the highest rosiglitazone concentration applied (100 µM). This effect is likely to be due to the simultaneous blockade of INa and Ito, with the former effect tending to decrease, and the latter tending to increase, the amplitude of action potentials. Finally, the lack of depolarization is compatible with the inability of rosiglitazone to alter IK1 at concentrations up to 100 µM.
Comparison with troglitazone
Our present results allow some comparison between the cellular cardiac electrophysiological effects of rosiglitazone and troglitazone. Troglitazone blocked ICa with IC50 values close to 10 µM in rat (Arikawa et al., 2002; 2004;), rabbit (Ikeda and Watanabe, 1998) and guinea pig (Nakajima et al., 1999) myocytes. This value is significantly smaller than the IC50 of 92 µM obtained with rosiglitazone in canine ventricular cells (present study). The inhibitory effect of troglitazone on INa is also much stronger than that of rosiglitazone. In contrast to our results, where 100 µM rosiglitazone caused less than 50% reduction of Vmax, 1 µM of troglitazone induced 50% Vmax blockade, while 10 µM of the compound fully eliminated action potentials in rabbit ventricular myocytes (Ikeda and Watanabe, 1998). Thus the difference between the inhibiting potency of rosiglitazone and troglitazone seems to be at least one order of magnitude. In summary, as rosiglitazone is a weaker inhibitor of cardiac ion channels than the other thiazolidinedione derivative troglitazone, less cardiac side effects would be anticipated with rosiglitazone than with troglitazone.
Clinical implications
The lowest concentration of rosiglitazone that caused statistically significant changes in our study was much higher than the peak plasma levels obtained in patients. Peak plasma concentration of 0.8 µg·mL−1 (corresponding to 2 µM) is typical in patients after receiving a single dose of 8 mg rosiglitazone (Park et al., 2004; Wittayalertpanya et al., 2010). Therefore, it is not likely that rosiglitazone, in normal doses, would alter cardiac electrogenesis in healthy individuals. In line with this, no case of sudden death has been reported in association with rosiglitazone therapy. But what might happen with a rosiglitazone overdose? Under such non-normal conditions, rosiglitazone concentrations in the plasma are likely to reach a much higher level, probably several tens of micromoles, where a variety of cardiac ion channels could be suppressed thus favouring the development of cardiac arrhythmias. This may be especially dangerous in patients with multiple cardiovascular risk factors, like elderly diabetic patients with ischemic heart disease. Indeed, rosiglitazone has been shown to increase the propensity for ventricular fibrillation in ischemic pigs (Lu et al., 2008). Thus the pro-arrhythmic action of rosiglitazone may be combined with, and contribute to, the development of other unfavourable cardiac side effects observed in an at-risk group of diabetic patients. This is in line with the consensus statement of the American and European Diabetes Associations, advising clinicians against using rosiglitazone in patients with type 2 diabetes mellitus (Krentz, 2009; Kaul et al., 2010).
Limitations of the study
The present results with rosiglitazone were obtained in healthy canine hearts, while rosiglitazone is usually given to diabetic patients. Diabetes is known to induce marked remodelling in the set of cardiac ion currents in all mammalian species studied, including dogs (Lengyel et al., 2007), rabbits (Lengyel et al., 2008) and rats (Magyar et al., 1992). In addition, differences in the effects of both rosiglitazone and troglitazone were observed when compared between healthy and diabetic rats (Arikawa et al., 2002; Kavak et al., 2008). Our data therefore should be extrapolated to diabetic patients with caution.
Acknowledgments
Financial support for the studies was provided by grants from the Hungarian Scientific Research Fund (OTKA-K68457, OTKA-K73160, CNK-77855). Further support was obtained from the Hungarian Government (TÁMOP-4.2.1/B-09/1/KONV-2010–007), and the Medical and Health Science Center of University of Debrecen (MEC-14/2008).
Glossary
Abbreviations
- APD50
action potential duration measured at 50% level of repolarization
- APD90
action potential duration measured at 90% level of repolarization
- ICa
L-type Ca2+ current
- IK1
inward rectifier K+ current
- IKr
rapid delayed rectifier K+ current
- INa
Na+ current
- Ito
transient outward K+ current
- Vmax
maximum velocity of depolarization
Conflicts of interest
None.
Supporting Information
Teaching Materials; Figs 1–7 as PowerPoint slide.
References
- Alexander SPH, Mathie A, Peters JA. Guide to Receptors and Channels (GRAC), 4th edition. Br J Pharmacol. 2009;158(Suppl. 1):S1–S180. doi: 10.1111/j.1476-5381.2009.00499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arikawa M, Takahashi N, Kira T, Hara M, Saikawa T, Sakata T. Enhanced inhibition of L-type calcium currents by troglitazone in streptozotocin-induced diabetic rat cardiac ventricular myocytes. Br J Pharmacol. 2002;136:803–810. doi: 10.1038/sj.bjp.0704757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arikawa M, Takahashi N, Kira T, Hara M, Yoshimatsu H, Saikawa T. Attenuated inhibition of L-type calcium currents by troglitazone in fructose-fed rat cardiac ventricular myocytes. J Cardiovasc Pharmacol. 2004;44:109–116. doi: 10.1097/00005344-200407000-00015. [DOI] [PubMed] [Google Scholar]
- Bányász T, Magyar J, Szentandrássy N, Horváth B, Birinyi P, Szentmiklósi J, et al. Action potential clamp fingerprints of K+ currents in canine cardiomyocytes: their role in ventricular repolarization. Acta Physiol (Scand) 2007;190:189–198. doi: 10.1111/j.1748-1716.2007.01674.x. [DOI] [PubMed] [Google Scholar]
- Berger J, Bailey P, Biswas C, Cullinan CA, Doebber TW, Hayes NS, et al. Thiazolidinediones produce a conformational change in peroxisomal proliferator-activated receptor-gamma: binding and activation correlate with antidiabetic actions in db/db mice. Endocrinology. 1996;137:4189–41950. doi: 10.1210/endo.137.10.8828476. [DOI] [PubMed] [Google Scholar]
- Cuzzocrea S, Pisano B, Dugo L, Patel NS, Paola RD, Genovese T, et al. Rosiglitazone and 15-deoxy-▵12,14- prostaglandin J2 ligands of the peroxisome proliferator-actvated receptor-γ (PPAR-γ) reduce ischemia/reperfusion injury of the gut. Br J Pharmacol. 2003;140:366–376. doi: 10.1038/sj.bjp.0705419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doshi LS, Brahma MK, Bahirat UA, Dixit AV, Nemmani KV. Discovery and development of selective PPARgamma modulators as safe and effective antidiabetic agents. Expert Opin Investig Drugs. 2010;19:489–512. doi: 10.1517/13543781003640169. [DOI] [PubMed] [Google Scholar]
- DREAM Trial Investigators. Dagenais GR, Gerstein HC, Holman R, Budaj A, Escalante A, Hedner T, et al. Effects of ramipril and rosiglitazone on cardiovascular and renal outcomes in people with impaired glucose tolerance or impaired fasting glucose: results of the Diabetes REduction Assessment with ramipril and rosiglitazone Medication (DREAM) trial. Diabetes Care. 2008;31:1007–1014. doi: 10.2337/dc07-1868. [DOI] [PubMed] [Google Scholar]
- Fischmeister R, DeFelice LJ, Ayer RK, Levi R, DeHaan RL. Channel currents during spontaneous action potentials in embryonic chick heart sells. The action potential patch clamp. Biophys J. 1984;46:267–271. doi: 10.1016/S0006-3495(84)84020-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerstein HC, Yusuf S, Bosch J, Pogue J, Sheridan P, Dinccag N, et al. Effect of rosiglitazone on he frequency of diabetes in patients with impaired glucose tolerance or impaired fasting glucose: a randomised controlled trial DREAM (Diabetes Reduction Assessment with ramipril and rosiglitazone Medication) Lancet. 2006;368:1096–1105. doi: 10.1016/S0140-6736(06)69420-8. [DOI] [PubMed] [Google Scholar]
- Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch. 1981;391:85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- Home PD, Pocock SJ, Beck-Nielsen H, Gomis R, Hanefeld M, Dargie H, et al. Rosiglitazone evaluated for cardiac outcomes and regulation of glycaemia in diabetes (RECORD): study design and protocol. Diabetologia. 2005;48:1726–1735. doi: 10.1007/s00125-005-1869-1. [DOI] [PubMed] [Google Scholar]
- Horváth B, Magyar J, Szentandrássy N, Birinyi P, Nánási PP, Bányász T. Contribution of IKs to ventricular repolarization in canine myocytes. Pflugers Arch. 2006;452:698–706. doi: 10.1007/s00424-006-0077-2. [DOI] [PubMed] [Google Scholar]
- Ikeda S, Watanabe T. Effects of troglitazone and pioglitazone on the action potentials and membrane currents of rabbit ventricular myocytes. Eur J Pharmacol. 1998;357:243–250. doi: 10.1016/s0014-2999(98)00557-3. [DOI] [PubMed] [Google Scholar]
- Katoh Y, Hashimoto S, Kimura J, Watanabe T. Inhibitory action of troglitazone, an insulin-sensitising agent, on the calcium current in cardiac ventricular cells of guinea pig. Jpn J Pharmacol. 2000;82:102–109. doi: 10.1254/jjp.82.102. [DOI] [PubMed] [Google Scholar]
- Kaul S, Bolger AF, Herrington D, Guiliano RP, Eckel RH. Thiazolidinedione drugs and cardiovascular risks. A science advisory from the American Heart Association and American College of Cardiology Foundation. Circulation. 2010;21:1868–1877. doi: 10.1161/CIR.0b013e3181d34114. [DOI] [PubMed] [Google Scholar]
- Kavak S, Emre M, Tetiker T, Kavak T, Kolcu Z, Günay I. Effects of rosiglitazone on altered electrical left ventricular papillart muscle activities of diabetic rat. Naunyn-Schmiedberg's Arch Pharmacol. 2008;376:415–421. doi: 10.1007/s00210-007-0234-y. [DOI] [PubMed] [Google Scholar]
- Khandoudi N, Delerive P, Berrebi-Bertrand I, Buckingham RE, Staels B, Bril A. Rosiglitazone, a peroxisome proliferator-activated receptor-γ, inhibits the Jun NH2-terminal kinase activating protein 1 pathway and protects the heart from inschemic/reperfusion injury. Diabetes. 2002;51:1507–1514. doi: 10.2337/diabetes.51.5.1507. [DOI] [PubMed] [Google Scholar]
- Knock GA, Mishra SK, Aaronson PI. Differential effects of insulin-sensitizers troglitazone and rosiglitazone on ion currents in rat vascular smooth muscle. Eur J Pharmacol. 1999;368:103–109. doi: 10.1016/s0014-2999(99)00020-5. [DOI] [PubMed] [Google Scholar]
- Krentz A. Thiazolidinediones: effects of the development and progression of type 2 diabetes and associated vascular complications. Diabetes Metab Res Rev. 2009;25:112–126. doi: 10.1002/dmrr.937. [DOI] [PubMed] [Google Scholar]
- Lengyel C, Virág L, Bíró T, Jost N, Pacher P, Magyar J, et al. Diabetes mellitus attenuates the repolarization reserve in mammalian heart. Cardiovasc Res. 2007;73:512–520. doi: 10.1016/j.cardiores.2006.11.010. [DOI] [PubMed] [Google Scholar]
- Lengyel C, Virág L, Pacher P, Kocsis E, Nánási PP, Tóth M, et al. Analysis of the electrophysiological effects of the alloxan induced diabetes mellitus in rabbit heart. Acta Physiol Scand. 2008;192:359–368. doi: 10.1111/j.1748-1716.2007.01753.x. [DOI] [PubMed] [Google Scholar]
- Lu L, Reiter MJ, Xu Y, Chicco A, Greyson CR, Schwartz GG. Thiazolidinedione drugs block cardiac KATP channels and may increase propensity for ischemic ventricular fibrillation in pigs. Diabetologia. 2008;51:675–685. doi: 10.1007/s00125-008-0924-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGuire DK, Inzucchi SE. Drugs for the treatment of diabetes mellitus. Part I: Thiazolidinediones and their evolving cardiovascular implications. Circulation. 2008;117:440–449. doi: 10.1161/CIRCULATIONAHA.107.704080. [DOI] [PubMed] [Google Scholar]
- McKay NG, Kinsella JM, Campbell CM, Ashford ML. Sensitivity of Kir6.2-SUR1 currents, in the absence and presence of sodium azide, to the KATP channel inhubitors, ciclazindole and englitazone. Br J Pharmacol. 2000;130:857–866. doi: 10.1038/sj.bjp.0703395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magyar J, Rusznák Z, Szentesi P, Szucs G, Kovács L. Action potentials and potassium currents in rat ventricular muscle during experimental diabetes. J Mol Cell Cardiol. 1992;24:841–853. doi: 10.1016/0022-2828(92)91098-p. [DOI] [PubMed] [Google Scholar]
- Mannucci E, Monami M, Di Bari M, Lamanna C, Gori F, Gensini GF, et al. Cardiac safety profile of rosiglitazone. A comprehensive meta-analysis of randomised clinical trials. Int J Cardiol. 2010;143:135–140. doi: 10.1016/j.ijcard.2009.01.064. [DOI] [PubMed] [Google Scholar]
- Mishra SK, Aaronson PI. Differential block by troglitazone and rosiglitazone of glibenclamide-sensitive K+ current in rat aorta myocytes. Eur J Pharmacol. 1999;368:121–125. doi: 10.1016/s0014-2999(99)00713-x. [DOI] [PubMed] [Google Scholar]
- Nakajima T, Iwasawa K, Oonuma H, Imuta H, Hazama H, Asano M, et al. Troglitazone inhibits voltage-dependent calcium currents in guinea pig cardiac myocytes. Circulation. 1999;99:2942–2950. doi: 10.1161/01.cir.99.22.2942. [DOI] [PubMed] [Google Scholar]
- Nissen SE, Wolski K. Effect of rosiglitazone on the risk of myocardial infarction and death from cardiovascular causes. N Engl J Med. 2007;357:2461–2471. doi: 10.1056/NEJMoa072761. [DOI] [PubMed] [Google Scholar]
- Pancani T, Phelps JT, Searcy JL, Kilgore MW, Chen KC, Porter NM, et al. Distinct modulation of voltage-gated and ligand-gated Ca2+ currents by PPAR-gamma agonists in cultured hipocampal neurons. J Neurochem. 2009;109:1800–1811. doi: 10.1111/j.1471-4159.2009.06107.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park J-Y, Kim K-A, Shin J-G, Lee KY. Effect of ketoconazole on the pharmacokinetics of rosiglitazone in healthy subjects. Br J Clin Pharmacol. 2004;58:397–402. doi: 10.1111/j.1365-2125.2004.02161.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pavlov TS, Levchenko VL, Karpushev AV, Vandewalle A, Staruschenko A. Peroxisome proliferators-activated receptor gamma antagonists decrease Na+ transport via the epithelial Na+ channel. Mol Pharmacol. 2009;76:1333–1340. doi: 10.1124/mol.109.056911. [DOI] [PubMed] [Google Scholar]
- Quinn CE, Hamilton PK, Lockhart CJ, McVeigh GE. Thiazolidinediones: effects on insulin resistance and the cardiovascular system. Br J Pharmacol. 2008;153:636–645. doi: 10.1038/sj.bjp.0707452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosen JC. The rosiglitazone story: lessons from an FDA Advisory Committee meeting. N Engl J Med. 2007;357:844–846. doi: 10.1056/NEJMp078167. [DOI] [PubMed] [Google Scholar]
- Strichartz G, Cohen I. Vmax as a measure of GNa in nerve and cardiac membranes. Biophys J. 1978;23:153–156. doi: 10.1016/S0006-3495(78)85440-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szabó G, Szentandrássy N, Bíró T, Tóth IB, Czifra G, Magyar J, et al. Asymmetrical distribution of ion channels in canine and human left ventricular wall: epicardium versus midmyocardium. Pflugers Arch. 2005;450:307–316. doi: 10.1007/s00424-005-1445-z. [DOI] [PubMed] [Google Scholar]
- Szentandrássy N, Bányász T, Bíró T, Szabó G, Tóth IB, Magyar J, et al. Apico-basal inhomogeneity in distribution of ion channels in canine and human ventricular myocardium. Cardiovasc Res. 2005;65:851–860. doi: 10.1016/j.cardiores.2004.11.022. [DOI] [PubMed] [Google Scholar]
- Welters HJ, McBain SC, Tadayyon M, Scarpello JH, Smith SA, Morgan NG. Expression and functional activity of PPAR-γ in pancreatic beta cells. Br J Pharmacol. 2004;142:1162–1170. doi: 10.1038/sj.bjp.0705844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wittayalertpanya S, Chompootaveep S, Thaworn N, Khemsri W, Intanil N. Oharmacokinetic and bioequivalence study of an oral 8 mg dose of rosiglitazone tablets in Thai healthy volunteers. J Med Assoc Thai. 2010;93:722–728. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.