

Abstract
Significant spontaneous recovery occurs after essentially all forms of serious brain injury, although the mechanisms underlying this recovery are unknown. Given that many forms of brain injury such as traumatic brain injury (TBI) induce hippocampal neurogenesis, we investigated whether these newly generated neurons might play a role in recovery. By modeling TBI in transgenic mice, we determined that injury-induced newly generated neurons persisted over time and elaborated extensive dendritic trees that stably incorporated themselves throughout all neuronal layers of the dentate gyrus. When we selectively ablated dividing stem/progenitors at the time of injury with ganciclovir in a nestin–HSV–TK transgenic model, we eliminated injury-induced neurogenesis and subsequently diminished the progenitor pool. Moreover, using hippocampal-specific behavioral tests, we demonstrated that only injured animals with neurogenesis ablated at the time of injury lost the ability to learn spatial memory tasks. These data demonstrate a functional role for adult neurogenesis after brain injury and offer compelling and testable therapeutic options that might enhance recovery.
Introduction
The mechanisms underlying spontaneous recovery from a variety of brain injuries remain unclear, although injury-induced neurogenesis may contribute to some of this observed self-recovery (Eriksson et al., 1998; Chirumamilla et al., 2002; Chen et al., 2003; Richardson et al., 2007). The two most well described populations of neural stem and progenitor cells in mammals are the subventricular zone of the lateral ventricles and the subgranular zone of the dentate gyrus (Alvarez-Buylla et al., 2001; Seaberg and van der Kooy, 2002). Normally, the subventricular zone in adults contributes progenitors to the rostral migratory stream to support ongoing olfactory neurogenesis, whereas the subgranular zone of the dentate gyrus provides new granular neurons throughout life within the hippocampus (Temple and Qian, 1995; Alvarez-Buylla et al., 2001). After many forms of brain injury, progenitor cells proliferate, although it is still unclear whether this activation results in stable and productive neurogenesis (Richardson et al., 2007; Kernie and Parent, 2010).
Evidence for long-lasting hippocampal neurogenesis after traumatic cortical injury has been accumulating since first described almost a decade ago (Dash et al., 2001; Kernie et al., 2001). The dentate gyrus itself develops mostly during the postnatal period, and the reservoir of subgranular zone progenitors persists well into adulthood (Pozniak and Pleasure, 2006). These progenitors develop almost exclusively into granular neurons within the most basal layers of the dentate gyrus, and their functions remain essentially unknown (Duan et al., 2008). After injury, however, even in adult animals, it is clear that progenitors are activated and increased neurogenesis follows whereby new neurons are found in the outer layers of the dentate gyrus, in which normally this would occur only during early development (Dash et al., 2001; Chirumamilla et al., 2002; Chen et al., 2003; Ramaswamy et al., 2005; Richardson et al., 2007; Urrea et al., 2007; Yu et al., 2008). Most of the evidence to support these findings is based on 5-bromo-2-deoxyuridine (BrdU) incorporation into dividing cells that when followed over time express markers, suggesting mature and stable neurogenesis. Although widely used, there are limitations to BrdU-based approaches, particularly after injury (Gould and Gross, 2002).
There remains tremendous speculation regarding the significance of adult neurogenesis, and there are conflicting data suggesting its function in cognitive processes (Kokovay et al., 2008). The purpose of the present study was to determine whether adult hippocampal neurogenesis plays a functional role in the spontaneous recovery that is observed after brain injury. To do this, we used a variety of mouse transgenic models to inducibly and persistently label neural stem and progenitor cells, as well as to ablate their proliferation in response to injury. We demonstrate that injury-induced neurogenesis occurred throughout the dentate gyrus and that these neurons stably incorporated themselves within the hippocampus and elaborated sophisticated dendritic arborizations. Moreover, when ablated at the time of injury using a ganciclovir (Gan)-based system, few newly born neurons were generated, and cognitive recovery was severely impaired.
Materials and Methods
Animals.
Experimental animals were housed and cared for in the Animal Resource Center at University of Texas Southwestern Medical Center (UTSWMC), which is certified by the Association for Assessment and Accreditation of Laboratory Animal Care. All animal experiments were conducted with previous approval of the Institutional Animal Use and Care Committee at UTSWMC in compliance with the highest standard for the humane and compassionate use of animals in biomedical research.
Controlled cortical impact injury.
To perform controlled cortical impact (CCI), the standard protocol and a controlled cortical impact device used to generate brain injuries were used as described previously (Kernie et al., 2001). Eight-week-old male wild-type or transgenic mice were anesthetized with isoflurane. Mice were placed in a stereotactic frame. A midline incision was made, the soft tissues were reflected, and a 5 × 5 mm craniectomy was made between bregma and lambda and 1 mm lateral to the midline. The injury was generated with a 3 mm stainless steel tipped impact device with a deformation of 0.6 mm and constant speed of 4.4 m/s. After injury, the scalp was fastened with staples, and the mice were allowed to recover. After injury, 5-chloro-2-deoxyuridine (CldU) (100 mg/kg; Sigma) was injected intraperitoneally 3 d after injury. Numbers of animals used for each group are listed in the figure legends.
Immunohistochemistry.
All mice were deeply anesthetized with ketamine and xylazine mixture as described above before perfusion. The transcardiac perfusion was performed with 50 ml of 1× PBS, followed by 50 ml of 4% paraformaldehyde (PFA) in PBS. After postfixation in 4% PFA/1× PBS overnight, whole brains were dissected and embedded in 3% agarose/1× PBS. Serial 50 μm sections were cut with a vibratome (VT1000S; Leica). All sections encompassing the hippocampus were collected sequentially in 12-well plates. Free-floating sections were used for immunohistochemistry. For CldU staining, all sections from a single well were washed with 1× PBS for three times and rinsed with water. Sections were washed with 0.3% Triton X-100/1× PBS (wash buffer) for three times and blocked with 5% normal donkey serum (Sigma-Aldrich), which contained wash buffer for 1 h at room temperature. Rat-anti CldU (1:300; AbD Serotec) antibody was used to label CldU overnight at 4°C. The following day, sections were incubated with mouse anti-neuronal-specific nuclear protein (NeuN) (1:500; Millipore Bioscience Research Reagents), washed, and then incubated with secondary antibodies (cyanine 3-conjugated anti-rat antibody and cyanine 2-conjugated anti-mouse; all 1:200; Jackson ImmunoResearch) for 3 h at room temperature. Additional primary antibodies used in this study were rabbit anti-green fluorescent protein (GFP) (1:500; Molecular Biology), goat anti-doublecortin (DCX) (1:100; Santa Cruz Biotechnology), rat anti-BrdU (1:300; Abcam), mouse anti-glutamine synthetase (1:500; Millipore Corporation), and chicken anti-glial fibrillary astrocytic protein (GFAP) (1:500; Aves Labs).
Tamoxifen injections.
Mice were bred to obtain heterozygous nestin–CreERT2 transgenic and heterozygous stop–YFP–ROSA26 to allow for tamoxifen-mediated long-term fate labeling of nestin-expressing progenitors. Twelve double-heterozygous 7-week-old mice were injected with 250 μl (6.7 mg/ml) of tamoxifen (0.6 mg/kg in 9:1 sunflower oil/ethanol) daily for 3 d. After another 3 d, they underwent CCI injury or mock injury and then allowed to recover for 2 months. After this, they were perfused, and brains were processed as outlined above and immunostained for GFP [to detect yellow fluorescent protein (YFP)].
Quantification and confocal microscopy.
To determine the number of YFP-expressing cells 2 months after injury (see Fig. 1) and GFP- and DCX-expressing cells in the dentate gyrus after ganciclovir treatment (see Fig. 2), we used an unbiased stereological approach. A one-in-six series of sections covering the entire hippocampus in its rostrocaudal extension were cut with a vibrating microtome (Leica) at a 50 μm thickness, immunostained, and visualized under light microscopy with the peroxidase/diaminobenzidine method. The cells were counted using a modified optical fractionator and stereological image analysis software (StereoInvestigator; MicroBrightField) operating a computer-driven Olympus microscope regulated in the x-, y-, and z-axes. Areas to be counted were traced with a 20× objective lens, and sample frames (80 × 80 μm) were selected at random by the image analysis software. Cells in the counting frame (40 × 40 μm) were counted under a 40× objective lens. To avoid oversampling, the uppermost and lowermost focal planes were excluded. The total cell number of cells was estimated by multiplying the resulting counts by 6, because every sixth section had been used. Hence, the numbers obtained in this study are absolute numbers per hippocampus (both sides of the dentate gyrus counted for each) and are independent of the volume of the dentate gyrus.
Figure 1.

Controlled cortical injury induces stable dentate gyrus neurogenesis. A–G, Nestin–CreERT2/stop–YFP–ROSA26 mice given 3 d of tamoxifen show YFP-expressing cells that appear to be neurons (A–F, arrowhead) and type 1 stem/progenitors (D, arrow) in the basal granular layer 2 months after tamoxifen and mock injury. B, E, After injury, YFP-expressing neuronal-appearing cells are found throughout the granular layer (arrowheads) on the injured side. C, F, The uninjured side shows a few neurons outside the basal granular layer (arrowhead), but most YFP-expressing cells are type 1 early progenitors in the subgranular zone (F, arrow). G, Unbiased stereological counts of YFP-expressing cells in the granular zone show a threefold increase only in the injured dentate gyrus. Numbers graphed are total numbers of dentate gyrus cells estimated using unbiased stereology (see Materials and Methods). Results shown are from n = 6 in injured animals and n = 4 in controls. Scale bars: A, 50 μm; D, 200 μm. **p < 0.01 using ANOVA with Bonferroni's post hoc correction.
Figure 2.

YFP-expressing cells in the dentate gyrus express neuronal markers. A–H, Nestin–CreERT2/stop–YFP–ROSA26 mice given 3 d of tamoxifen, injured, and killed 2 months later show YFP-expressing cells that colocalize with the neuronal marker NeuN and the astrocyte/progenitor marker GFAP. Control dentate gyrus shows NeuN/YFP-expressing cells restricted to inner granular layer (A–D, arrows), whereas in injured animals, these cells are more numerous and found in outer granular layer (E–H, arrowheads). Cells expressing both YFP and GFAP remain restricted to the subgranular zone and are not found in reactive astrocytes in the hilus or outside granular layer (C,D, G,H). Scale bar: 50 μm.
For cell quantification for double labeling of YFP/NeuN, YFP/GFAP, CldU/NeuN, and GFAP/BrdU-expressing cells, we used a traditional and well established method to account for oversampling as described in detail previously (Miles and Kernie, 2008; Yu et al., 2008). A Carl Zeiss LSM 510 confocal microscope using argon 488, helium 543, and helium 633 lasers and a Carl Zeiss Neofluar 40×/1.3 oil lens was used to determine colocalization of immunohistochemical markers and cell quantification with a pinhole aperture of 3.46 Airy units. Cell quantification was performed from the ipsilateral and contralateral dentate gyrus from injured mice. All brains were embedded in 3% agarose and cut with a vibrating microtome (Leica) at a 50 μm thickness using a sapphire blade in 0.1 m PBS. Free-floating sections through the entire rostral-to-caudal hippocampal axis were collected in 12-well plates with five sections per well. We sampled one well of five sections 50 μm thick, coronally cut, and paraformaldehyde fixed 600 μm apart. Five sections from each well were stained with relevant antibodies as free-floating sections to allow for equal antibody penetration between sections. Sections were mounted using Immunomount with a refractive index of 1.495. Antibody penetration and signal intensity was found to be evenly distributed throughout the entire 50 μm section and was verified on the z-axis in multiple sections from different animals. Each blade of the dentate gyrus was counted separately at 40× under Carl Zeiss immersion oil 518F with 1.5× optical zoom magnification. Nonblinded quantification was performed on cells with fully colocalized antibodies in the granular layer of the dentate gyrus. Every double-positive cell was counted. By scanning serially through the z-axis of each cell, a more precise discrimination of the colocalization of relevant cell markers was permissible.
Ganciclovir administration.
Ganciclovir (200 mg · kg−1 · d−1; Cytovene-IV; Roche Pharmaceuticals) or vehicle (Veh) (dH2O) was delivered via osmotic mini pumps that infuse at a constant rate of 0.5 μl/h for 2 weeks (model 2002; Alzet). The pumps were replaced once after 2 weeks for a total of 4 weeks of treatment. The implantation of pumps was as per the Alzet protocol and is briefly described below: 6-week-old mice were anesthetized with isoflurane. A small incision was made in the skin between the scapulae, and a small pocket was formed using a hemostat to spread the subcutaneous connective tissue apart. The pumps were inserted into the pocket with the opening of pumps pointing away from the incision. The skin incision was closed with staples.
Behavioral overview.
All behavioral testing was conducted by investigators blind to the experimental treatment conditions. Mice were tested in three cohorts, and the total sample size for each experimental group was as follows: ganciclovir-treated group with CCI injury (“Injured + Gan”), n = 12; vehicle-treated group with CCI injury (“Injured + Veh”), n = 10; ganciclovir-treated group with sham injury (“Sham + Gan”), n = 9; and vehicle-treated group with sham injury (“Sham + Veh”), n = 10. All mice were provided with ad libitum food and water and kept on a 12 h light/dark cycle with lights on at 6:00 A.M. Behavioral testing was conducted during the light cycle, and behavioral tests occurred in the following order: Morris water maze, visible water maze, fear conditioning, and rotarod.
Morris water maze.
The Morris water maze task was conducted similar to the method described previously (Powell et al., 2004; Tabuchi et al., 2007; Etherton et al., 2009). A white, circular pool was filled with water that was mixed with gothic white, nontoxic paint to make it opaque, and a circular platform (15 cm in diameter) was submerged 1 cm beneath the surface of the water. The testing room was filled with a number of extramaze cues. Training in the Morris water maze was conducted over 11 consecutive days, and each training day consisted of four individual trials with an intertrial interval of ∼1–1.5 min. Mice were placed in a different starting location for each of the four daily training trials. During training trials, mice were allowed to swim until they found the hidden platform, and if they did not find the platform within 60 s, they were guided to it by the experimenter. Mice remained on the platform for 15 s before being removed. For each training day, data were averaged across the four trials. A probe trial was conducted on day 12; the hidden platform was removed, and mice were placed in the pool and allowed to swim for 60 s.
The visible version of the water maze was conducted during a single day, and there were six trials with an intertrial interval of ∼30 min. A visible cue (a black, foam square) was placed on top of the platform. The platform was placed in a new, random location for each trial, and mice were placed in the same starting location for each trial. All data were collected using Ethovision software (version 2.3.19; Noldus).
Fear conditioning.
For the cued and contextual fear-conditioning task, mice were initially placed in conditioning chambers. They were given a training session that consisted of an initial 2 min habituation period, followed by two presentations of a 30 s auditory tone (white noise) that coterminated with a 2 s, 0.5 mA footshock. There was a 1 min interstimulus interval between tone–shock pairings, and mice remained in the conditioning chambers for 1 min before being returned to their home cage. Freezing behavior (motionless except respirations) was measured live by an observer every 5 s.
A test session to examine contextual fear memory was conducted 24 h after the training session. Mice were placed back into the original training context for 5 min, and freezing behavior (motionless except respirations) was measured by an observer every 5 s. A test session to examine cued fear memory was conducted 3–4 h after the contextual fear memory test. Mice were placed in a novel context (visual, olfactory, auditory, and tactile aspects of the general environment were all changed); they experienced an initial 3 min habituation period, followed by a 3 min presentation of the auditory tone. Freezing behavior (motionless except respirations) was measured by an observer every 5 s, and normalized cue-dependent freezing was calculated by subtracting the percentage of time spent freezing during the habituation period from the percentage of time spent freezing during the tone presentation.
Rotarod.
During the rotarod task, mice were placed on a rotating cylindrical rod that accelerated from 4 to 45 revolutions/min over a period of 5 min. Mice were given four trials per day over 2 d, and the time to either fall off the rod or turn one full revolution was measured.
Statistics.
All statistical analyses of behavioral results were conducted using Statistica software (version 5.5; StatSoft). Data were initially analyzed using two-, three-, or four-way ANOVAs, as appropriate. Sex was initially included as a between-subjects factor; however, if there was no significant interaction or main effect of sex, the data were collapsed across sex. In cases in which there was a significant interaction or main effect of sex, it has been included as a factor in the reported analyses. Other between-subjects factors were injury (CCI injury vs sham) and treatment (vehicle vs ganciclovir), and within-subjects factors were day (for Morris water maze), trial (for visible water maze and rotarod), quadrant (for Morris water maze), or platform (for Morris water maze). Additional analyses were conducted using post hoc Tukey's honestly significant difference (HSD) tests and planned comparisons (contrast analysis and additional ANOVAs). A thorough description of all statistical results can be found in supplemental Tables 1 and 2 (available at www.jneurosci.org as supplemental material).
Statistical analyses of all nonbehavioral results were done using a one-way ANOVA for multiple comparisons with a Newman–Keuls multiple comparison test for post hoc analysis. A Student's t test was performed for paired data. Differences were deemed significant with p < 0.05.
Results
Traumatic brain injury induces long-lasting neurogenesis
We used a previously characterized nestin–Cre–ERT2/stop–YFP–ROSA26 transgenic mouse line that inducibly and specifically labels early stem/progenitors within the mouse brain (Li et al., 2008; Chen et al., 2009). By administering tamoxifen for 3 d to young, adult transgenic (8-week-old) mice, we were able to demonstrate that, 8 weeks later, there were many YFP-labeled cells that reside predominantly in the most superficial layers of the dentate gyrus (Fig. 1A,D). When mice underwent a unilateral, CCI injury 3 d after the last tamoxifen injection, there was triple the number of cells generated when compared with the mock-injured controls (Fig. 1A–G). In addition, the newly generated cells incorporated themselves throughout the dentate gyrus and not just in the most superficial layers (Fig. 1B,E). Notably, these cells expressed long dendritic processes indicative of neurons that have stably integrated themselves within the dentate gyrus (Fig. 1A–F).
Next, to prove that these peripherally incorporated YFP-expressing cells were indeed neurons, we performed triple staining with the mature neuronal marker NeuN as well as the astrocytic and progenitor marker GFAP (Fig. 2). Here, we observed that virtually all YFP-expressing cells in the outer granular layer of the dentate gyrus after injury also expressed NeuN, indicative of stable neurogenesis (Fig. 2E–H). In addition, both in the injured dentate gyrus and in controls, we observed that the YFP–GFAP double-expressing cells were almost exclusively limited to the subgranular zone indicative of early GFAP-expressing progenitors (Fig. 2A–H). We observed 177 ± 1 cells expressing both YFP and NeuN in the ipsilateral injured dentate gyrus compared with 117 ± 10 in mock-injured controls (n = 3 each group, p < 0.01, unpaired t test). Additionally, we observed 61 ± 11 expressing both YFP and GFAP in injured animals and 51 ± 6 in mock-injured controls (n = 3, p = 0.5, unpaired t test). Notably, we did not observe any reactive astrocytes that also express GFAP within the hilus or molecular layer of the dentate gyrus that also expressed YFP, suggesting that YFP-expressing progenitors became either neurons or remained as progenitors when exposed to injury.
Injury-induced neurogenesis is ablated in ganciclovir-treated nestin–HSV–TK transgenic animals
To determine whether these newly generated neurons had any functional relevance, we developed another transgenic mouse line that expresses a modified version of the herpes simplex virus thymidine kinase (HSV–TK) along with GFP controlled by the same nestin promoter and allows for ganciclovir-mediated ablation of dividing neural progenitors. We previously demonstrated that 4 weeks of exposure to ganciclovir is sufficient to specifically ablate >93% of doublecortin-expressing, committed neuroblasts in the adult dentate gyrus (Yu et al., 2008). To ablate the injury-induced neurogenesis associated with traumatic brain injury (TBI), osmotic minipumps either with vehicle or ganciclovir were implanted into the subcutaneous tissue of adult transgenic mice immediately after injury, which exposed the mice to ganciclovir for 4 weeks. In addition, each animal was injected with a single dose of CldU 3 d after injury to determine ganciclovir efficacy in ablating neurogenesis. During pump removal, each cohort of mice underwent behavioral testing, and then their brains were removed and examined histologically.
By immunostaining for the mature neuronal marker NeuN and CldU, we demonstrated that ∼90% of injury-induced neurogenesis was ablated with this technique (Fig. 3A,B). As we determined previously, the administration of ganciclovir only targets dividing progenitors so the relatively quiescent, early stem/progenitor is not susceptible. We therefore also examined the state of nestin/GFP-expressing early progenitors as well as later doublecortin-expressing committed neuroblasts. Here, we observed a modest reduction in each, suggesting that the progenitor population exhibited some recovery 1 month after removal of ganciclovir but remained persistently lower than what was seen in mock-injured controls (Fig. 3C–F).
Figure 3.
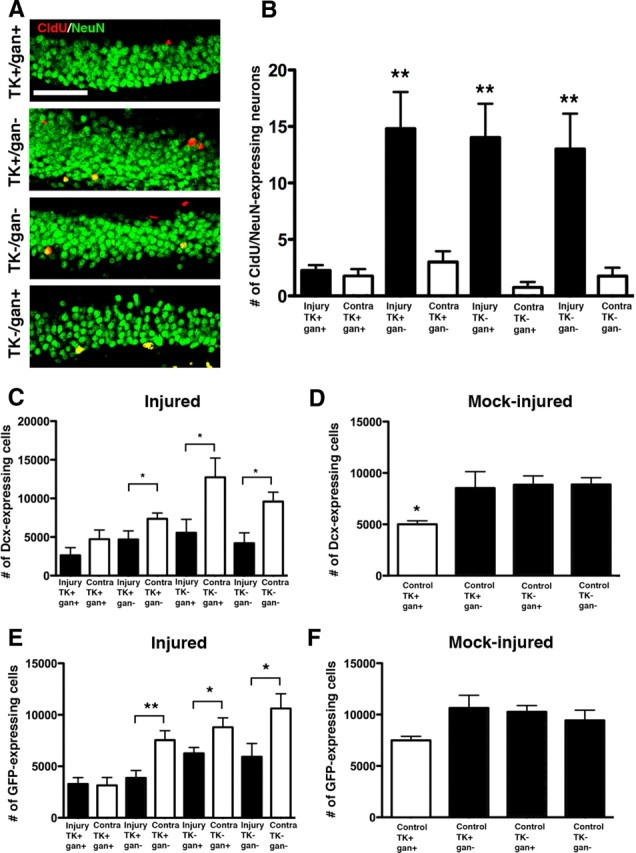
Injury-induced neurogenesis is ablated in ganciclovir-exposed nestin–HSV–TK transgenic mice. A, B, The same injury was performed on nestin–HSV–TK transgenic mice given a single dose of CldU 3 d after injury and treated with ganciclovir for 4 weeks after injury. One month later (2 months after injury), the only mice that demonstrate stable neurogenesis are transgenic mice not given ganciclovir, control mice given ganciclovir, or control mice not given ganciclovir (A). During blinded counting, there is a 90% ablation of neurogenesis only in transgenic animals exposed to ganciclovir (B). C, D, DCX-expressing committed neuroblasts are persistently decreased 2 months after injury and 1 month after ganciclovir treatment in nestin–HSV–TK–GFP transgenic mice. C, In injured transgenic mice given ganciclovir, both the injured and uninjured dentate gyrus have decreased numbers of DCX-expressing cells when unbiased stereological counts are performed 2 months after injury. In control but injured groups, DCX-expressing cells are increased on the uninjured side. D, In uninjured control groups, there are decreased numbers of DCX-expressing cells in the TK mice with ganciclovir. E, F, GFP-expressing early stem/progenitors are persistently decreased 2 months after injury and 1 month after ganciclovir treatment. E, In injured nestin–HSV–TK–GFP transgenic mice, both the injured and uninjured dentate gyrus have equivalent but decreased numbers of GFP-expressing cells when unbiased stereological counts are performed 2 months after injury. In control but injured groups, GFP-expressing progenitors are increased on the uninjured side. F, In uninjured control groups, there is a trend toward decreased numbers of GFP-expressing progenitors in the TK mice with ganciclovir, although this does not reach statistical significance. Numbers graphed are total numbers of dentate gyrus cells estimated using unbiased stereology (see Materials and Methods). *p < 0.05 and **p < 0.01 using t test comparing paired samples in C and E. For B, D, and F, ANOVA with Bonferroni's post hoc analysis was used. n = 5 for Injury+/TK+/gan+, 5 for Injury+/TK+/gan−, 4 for Injury−/TK+/gan+, 4 for Injury−/TK−/gan+, 3 for Control/TK+/gan+, 4 for Control/TK+/gan−, 4 for Control/TK−/gan−, and 3 for Control/TK+/gan−. Scale bar, 50 μm.
Ganciclovir-mediated ablation of progenitors does not impair astrogliosis
Reactive gliosis occurs after essentially all forms of brain injury and likely plays roles in how recovery is mediated (Garcia et al., 2004; Myer et al., 2006). In addition, nestin is expressed in other cell types in addition to early progenitors and includes NG2-expressing oligodendrocytes, reactive astrocytes, and endothelial cells (Ridet et al., 1997; Belachew et al., 2003; Aguirre and Gallo, 2004; Yokoyama et al., 2006). Many transgenic animals have been made that express proteins such as GFP under the control of various aspects of nestin regulatory elements, including the endogenous nestin promoter with all its enhancer elements as well as a minimal viral promoters with the second intron enhancer (Yamaguchi et al., 2000; Kawaguchi et al., 2001; Lagace et al., 2007). Here, we used the full nestin promoter and the second intron progenitor-specific enhancer in both the nestin–CreERT2 transgenic as well as the nestin–HSV–TK mice. Although endogenous nestin is expressed in reactive astrocytes, we demonstrated previously that genes under the control of the neural progenitor-specific form of the nestin promoter used in the transgenic animals in this study are expressed only in progenitors and not in reactive astrocytes in response to both traumatic and hypoxic-ischemic injuries (Miles and Kernie, 2008; Yu et al., 2008). However, because it remains a formal possibility that this nestin–HSV–TK transgenic could ablate both dividing stem/progenitors as well dividing reactive astrocytes, we performed a detailed investigation.
Nestin–HSV–TK mice that were injured and given either ganciclovir or vehicle for 1 month after injury were given BrdU during the peak of astrogliosis (once daily, postinjury days 1–3). One month after injury, the mice were analyzed for reactive gliosis. We observed that there was a similar amount of overall gliosis in both the ganciclovir- and vehicle-treated animals (Fig. 4). Moreover, when quantified, there were similar numbers of proliferative reactive astrocytes in each group as evidenced by coexpression of GFAP and BrdU (101 ± 13.6, control, n = 3; 112 ± 14.4, ganciclovir-treated, n = 4; not significant). Finally, we also observed that the maturation of these reactive astrocytes was similar in each group by performing immunostaining with glutamine synthetase, a marker of mature astrocytes, in which we observed that in each group a subset of reactive astrocytes that expressed BrdU also expressed glutamine synthetase (Fig. 4D,H).
Figure 4.

Ganciclovir does not ablate reactive astrocytosis after injury in nestin–HSV–TK transgenic mice. Transgenic nestin–HSV–TK mice were injured and given ganciclovir via implanted osmotic pumps for 30 d and injected with BrdU on days 1–3 after injury. A, E, There was no difference in overall GFAP-expressing astrogliosis in the hippocampus of TK-expressing mice given either vehicle or ganciclovir. A–C, E–G, The number of dividing cells evidenced by BrdU incorporation at 1–3 d after injury was also similar 30 d later (101 ± 13.6 for control, 112 ± 14.4 for Gan-treated; n = 3 for control and 4 for Gan-treated; not significant). D, H, A subset of reactive astrocytes from each group express BrdU, GFAP, and the mature astrocyte marker glutamine synthetase (GS).
Ablation of neurogenesis does not affect fear conditioning or motor learning
To determine whether the ablation of neurogenesis would modulate the effect of brain injury on nonspatial forms of learning and memory, mice containing the thymidine kinase were trained in contextual and cued fear conditioning as well as a rotarod task after receiving either CCI or a sham injury and treatment with either Gan or Veh. During tests of both cued and contextual fear memory, all mice exhibited comparable levels of freezing (Fig. 5A; cued fear conditioning, two-way ANOVA: injury, p = 0.97; treatment, p = 0.27; injury × treatment, p = 0.85; contextual fear conditioning, two-way ANOVA: injury, p = 0.09; treatment, p = 0.49; injury × treatment, p = 0.26) (for detailed statistical analyses of behavioral tests, see supplemental Table 1, available at www.jneurosci.org as supplemental material). These data suggest not only that ablation of neurogenesis did not modulate the effect of brain injury on fear learning and memory but also that neither ablation of neurogenesis alone nor brain injury alone affected this type of learning and memory. Next, motor coordination and learning were assessed using the rotarod task (Fig. 5B). Although a four-way repeated-measures ANOVA did find significant main effects of brain injury and ganciclovir treatment (sex, p < 0.02; injury, p < 0.02; treatment, p < 0.04; trial, p < 0.000001; no other significant main effects or interactions), additional analysis found that the injured mice with or without ablation of neurogenesis took less time to fall off the rotarod compared with uninjured mice with ablation of neurogenesis (post hoc Tukey's HSD tests: Injured + Gan vs Sham + Gan, p < 0.05; Injured + Veh vs Sham + Gan, p < 0.002). However, there was no difference between injured mice with and without neurogenesis, suggesting that any mild effect of injury was not exacerbated by the ablation of neurogenesis (post hoc Tukey's HSD tests: Injured + Veh vs Injured + Gan, p > 0.10).
Figure 5.
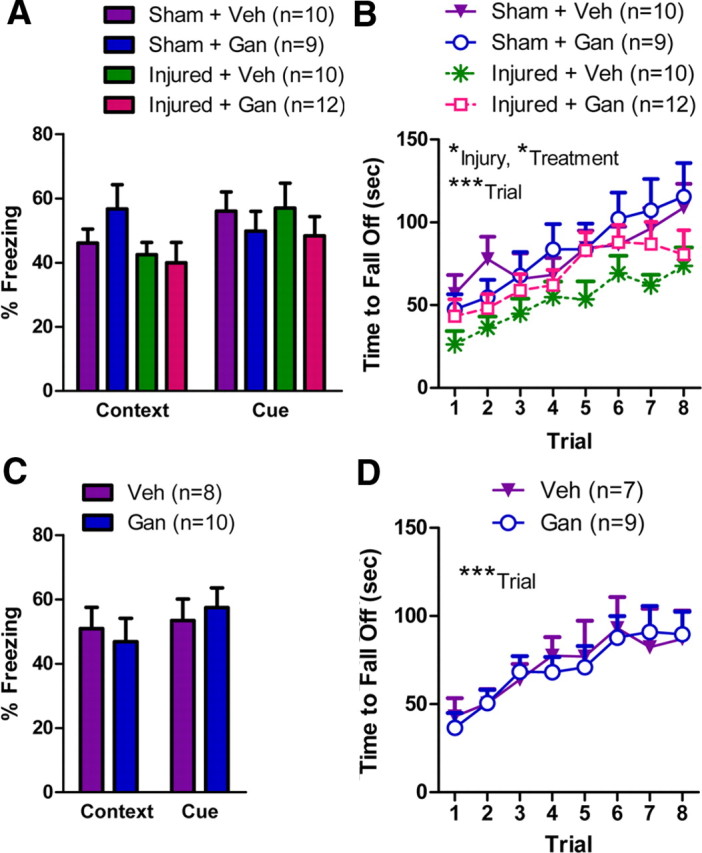
Neither mechanical brain injury nor reduction of neurogenesis affects contextual or cued fear conditioning, but mechanical brain injury results in impaired motor coordination. A, The percentage of time spent freezing during a 5 min test for contextual fear memory (Context) was similar for all mice regardless of injury or treatment. The percentage of cue-dependent freezing (Cue; percentage of time spent freezing during a 3 min presentation of a conditioned auditory stimulus minus the percentage of time spent freezing during an initial 3 min habituation period) during a 6 min test for cued fear memory was also similar for all mice regardless of injury or treatment. For all panels, the data represent means ± SEMs. B, The time for mice to fall off an accelerating rotarod (or turn one full revolution on the rotarod). Trials were conducted across 2 d with four trials per day. Although an initial ANOVA found a significant main effect of both injury and treatment (*p < .05, ***p < .001 for the indicated main effects in a four-way repeated-measures ANOVA), additional analysis found that the two injured groups (Injured + Veh and Injured + Gan) both fell off the rotarod faster than the Sham + Gan group (post hoc Tukey's tests, p < 0.05; no other between-group comparisons were significant). C, In mice lacking the TK transgene, the percentage of time spent freezing during a 5 min test for contextual fear memory (Context) was similar for both vehicle- and ganciclovir-treated mice. The percentage of cue-dependent freezing (Cue), percentage of time spent freezing during a 3 min presentation of a conditioned auditory stimulus minus the percentage of time spent freezing during an initial 3 min habituation period) during a 6 min test for cued fear memory was also similar for vehicle- and ganciclovir-treated mice lacking the TK transgene. D, In mice lacking the TK transgene, the time for mice to fall off an accelerating rotarod (or turn one full revolution on the rotarod) was similar for vehicle- and ganciclovir-treated mice. ***p < .001 for a main effect of trial in a two-way repeated-measures ANOVA.
To determine whether there were any nonspecific effects of ganciclovir on fear conditioning or the rotarod, mice lacking the TK transgene were treated with either vehicle or ganciclovir. Ganciclovir treatment had no effect on cued fear conditioning (Fig. 5C; two-way ANOVA: sex, p < 0.006; treatment, p = 0.84; sex × treatment, p = 0.74), contextual fear conditioning (Fig. 5C; one-way ANOVA: treatment, p = 0.69), or the rotarod task (Fig. 5D; two-way repeated-measures ANOVA: treatment, p = 0.92; trial, p < 0.000001; treatment × trial, p = 0.95), suggesting that there were no nonspecific effects of ganciclovir on these behaviors.
Ablation of neurogenesis exacerbates the cognitive effects of traumatic brain injury
To determine whether ablation of neurogenesis affected injury-induced cognitive deficits, we began behavioral testing 1 month after ablation of neurogenesis and injury, when injury-induced progenitors have had ∼4 weeks to mature and are most likely to functionally integrate into the hippocampus and participate in memory formation (Ge et al., 2006; Kee et al., 2007). Performance in the Morris water maze was used as the primary cognitive outcome to assess recovery. Mice containing the TK transgene were trained in the Morris water maze after receiving either CCI or a sham injury and treatment with either Gan or Veh.
Pharmacogenetic ablation of adult neurogenesis in combination with traumatic brain injury resulted in a modest decrease in initial spatial learning compared with sham controls (Fig. 6) (for detailed statistical analyses of behavioral tests, see supplemental Table 1, available at www.jneurosci.org as supplemental material). Brain-injured mice with ablation of neurogenesis (Injured + Gan) exhibited a longer latency to reach the hidden platform compared with all other groups (Fig. 6A; post hoc Tukey's HSD tests: Injured + Gan vs Injured + Veh, p < 0.04; Injured + Gan vs Sham + Gan, p < 0.006; Injured + Gan vs Sham + Veh, p < 0.0004) (for detailed statistical analyses of behavioral tests, see supplemental Table 1, available at www.jneurosci.org as supplemental material). Although injured mice without neurogenesis (Injured + Gan) did exhibit a mild decrease in average swim speed (Fig. 6B; post hoc Tukey's HSD tests: Injured + Gan vs Sham + Veh, p < 0.004), there was no effect of either brain injury or ganciclovir treatment on swim speed during the first day of training (Fig. 6B; two-way ANOVA: injury, p = 0.12; treatment, p = 0.86; injury × treatment, p = 0.56), suggesting that injured mice without neurogenesis (Injured + Gan) did not have any gross motor impairment but were unable to successfully increase their swim speed as training trials progressed. In another measure of learning that is independent of swim speed, injured mice without neurogenesis (Injured + Gan) traveled longer distances to reach the hidden platform compared with mice that received a sham injury (Fig. 6C; post hoc Tukey's HSD tests: Injured + Gan vs Sham + Gan, p < 0.0008; Injured + Gan vs Sham + Veh, p < 0.0002). Although all groups of mice spent a similar percentage of time swimming along the walls of the maze (thigmotaxis) during the first day of training (two-way ANOVA: injury, p = 0.17; treatment, p = 0.80; injury × treatment, p = 0.49), injured mice without neurogenesis (Injured + Gan) spent a greater percentage of time swimming along the walls of the maze over the rest of training compared with the other groups of mice (Fig. 6D; post hoc Tukey's HSD tests: Injured + Gan vs Injured + Veh, p < 0.03; Injured + Gan vs Sham + Gan, p < 0.0005; Injured + Gan vs Sham + Veh, p < 0.0003). This suggests that, unlike the other groups, the injured mice without neurogenesis (Injured + Gan) did not progress from the initial use of a thigmotaxic search strategy to the use of an efficient spatial strategy to find the hidden platform, and we interpret this result as a learning deficit (Lipp and Wolfer, 1998). These data suggest that ablation of neurogenesis after brain injury results in mild deficits in learning during the Morris water maze.
Figure 6.
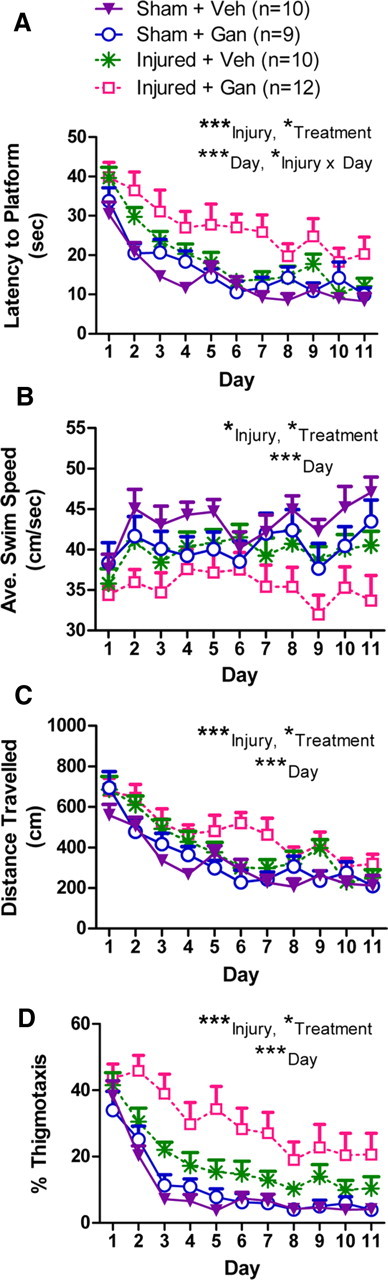
Reduction of neurogenesis in mice with traumatic brain injury results in impairments during training in the Morris water maze. A–D, Morris water maze training trials. *p < 0.05, **p < 0.01, ***p < 0.001 for the indicated main effects in a three-way repeated-measures ANOVA. A, Latency to reach the hidden platform. The Injured + Gan group took significantly longer to find the hidden platform compared with each of the other three groups (post hoc Tukey's tests, p < 0.05; no other between-group comparisons were significant). B, Average swim speed. The Injured + Gan group exhibited a significantly slower swim speed than the Sham + Veh group (post hoc Tukey's tests, p < 0.01; no other between-group comparisons were significant). C, Total distance traveled. The Injured + Gan group traveled a significantly greater distance while searching for the hidden platform compared with both Sham groups (i.e., Sham + Veh and Sham + Gan; post hoc Tukey's tests, p < 0.01; no other between-group comparisons were significant). D, Percentage of time spent swimming near the walls of the maze (i.e., thigmotaxis). The Injured + Gan group spent significantly more time swimming near the wall of the maze compared with each of the other three groups (post hoc Tukey's tests, p < 0.05; no other between-group comparisons were significant).
During the Morris water maze probe trial, brain-injured mice lacking adult neurogenesis (Injured + Gan) exhibited a complete lack of long-term spatial learning and memory. Using three different behavioral measures, injured mice without neurogenesis (Injured + Gan) did not show any discernable preference for the target quadrant or the previous location of the hidden platform (Fig. 7A–C, red bars; planned pairwise comparisons: for A, target vs right, p = 0.69; target vs left, p = 0.61; target vs opposite, p = 0.59; for B, target vs right, p = 0.63; target vs left, p = 0.44; target vs opposite, p = 0.09; for C, target vs right, p = 0.71; target vs left, p = 0.96; target vs opposite, p = 0.82; see paragraph below for similar comparisons within the other groups), suggesting either a lack of spatial memory or greatly impaired spatial learning of the location of the hidden platform. Importantly, the average swim speed during the probe trial was comparable among all groups (Fig. 7D; two-way ANOVA: injury, p = 0.67; treatment, p = 0.45; injury × treatment, p = 0.67), and examination of mice lacking the TK transgene determined that there were no nonspecific effects of ganciclovir on behavior (supplemental Table 2 and supplemental Figs. 1–3, available at www.jneurosci.org as supplemental material).
Figure 7.
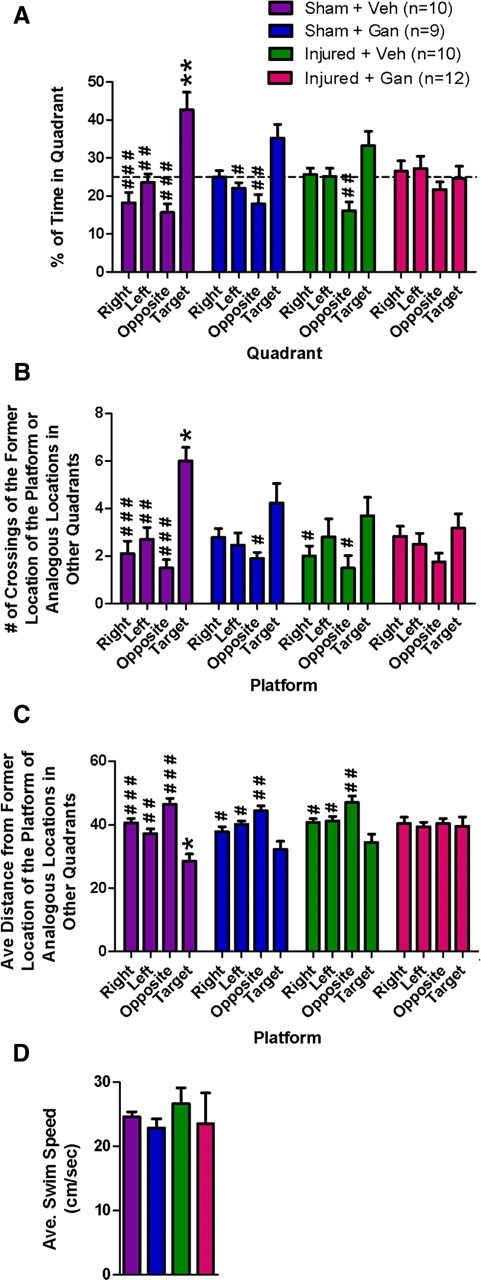
Reduction in neurogenesis results in impaired spatial learning and memory in the Morris water maze and impaired cognitive recovery after traumatic brain injury. A–D, Morris water maze probe trial. *p < 0.05, **p < 0.01 compared with Injured + Gan (post hoc Tukey's tests). #p < 0.05, ##p < 0.01, ###p < 0.001 compared with target quadrant for A or target platform for B and C (planned comparisons within each group). A, Percentage of time spent swimming in each quadrant of the maze. The target quadrant is the quadrant in which the hidden platform was previously located, and the dotted horizontal line represents chance performance (i.e., 25% of time spent in a quadrant). B, Number of times that mice crossed the former location of the hidden platform (i.e., target platform) and other analogous, identically sized locations in the other three quadrants. C, Average distance (averaged across the 1 min probe trial) from the former location of the hidden platform (i.e., target platform) or from other analogous, identically sized locations in the other three quadrants. D, Average swim speed during the 1 min probe trial.
Interestingly, a gradient effect of ablating neurogenesis with or without brain injury is also apparent in probe trial measures of spatial memory. Figure 7, A and B, clearly demonstrates that sham-injured mice with intact neurogenesis (Sham + Veh, purple bars) showed a significant preference for the target quadrant (or previous platform location) over all three other quadrants (or analogous zones in the other three quadrants of similar size and relative location as the hidden platform, termed “analogous platform locations”; planned pairwise comparisons: for Fig. 7A, target vs right, p < 0.00004; target vs left, p < 0.002; target vs opposite, p < 0.00005; for Fig. 7B, target vs right, p < 0.00001; target vs left, p < 0.002; target vs opposite, p < 0.00002). However, mice that received either brain injury alone (Injured + Veh, green bars) or ablation of neurogenesis alone (Sham + Gan, blue bars) significantly preferred the target quadrant (or previous platform location) compared with only one or two of the other quadrants (or analogous platform locations; planned pairwise comparisons: for Fig. 7A, Injured + Veh, target vs right, p = 0.14; target vs left, p = 0.16; target vs opposite, p < 0.006; for Fig. 7A, Sham + Gan, target vs right, p = 0.07; target vs left, p < 0.04; target vs opposite, p < 0.008; for Fig. 7B, Injured + Veh, target vs right, p < 0.04; target vs left, p = 0.34; target vs opposite, p < 0.02; for Fig. 7B, Sham + Gan, target vs right, p = 0.08; target vs left, p = 0.08; target vs opposite, p < 0.02). This suggests that both uninjured mice with ablation of neurogenesis (Sham + Gan) and injured mice with intact neurogenesis (Injured + Veh) exhibit an intermediate level of spatial memory that is slightly less robust than that of fully intact mice (Sham + Veh) but more robust than that of injured mice lacking neurogenesis (Injured + Gan). In total, these data suggest that ablation of adult neurogenesis does indeed exacerbate the cognitive effects of traumatic brain injury.
In the visible version of the water maze, all groups of mice were able to reach a visible platform in <12 s on average (supplemental Fig. 4, available at www.jneurosci.org as supplemental material), suggesting that neither brain injury nor ablation of neurogenesis caused impairments in vision. However, injured mice without neurogenesis (Injured + Gan) did exhibit some impairments in the visible version of the water maze, suggesting that they had mild impairments in either their ability to learn the basic rules of the water maze task or their ability to swim directly to the escape platform (supplemental Fig. 4, available at www.jneurosci.org as supplemental material). Again, these findings suggest that ablation of adult neurogenesis exacerbates the cognitive effects of traumatic brain injury.
Discussion
Because it is reasonably well established that hippocampal progenitors are activated by injury resulting in increased numbers of new neurons within the dentate gyrus, the primary purpose of this study was to establish the relevance of this phenomenon. The possibilities regarding the significance of injury-induced neurogenesis include the fact that the generation of new neurons might be beneficial and contribute to recovery of learning and memory and possibly other functions impaired by brain injury. However, neurogenesis may contribute to TBI-related morbidity such as epilepsy, which is relatively common after moderate and severe TBI. Finally, this reservoir of progenitors may be nothing more than a developmental remnant that is incapable of providing functionally relevant neurons into the sophisticated hippocampal circuitry. Although we demonstrate here that injury-induced neurogenesis occurs in a long-lasting manner, we also note that the progenitor pool itself may become depleted (Fig. 3). Because there is an apparently permanent depletion of GFP-expressing early progenitors after injury, a number of questions remain open. There may either be damage to the ipsilateral dentate gyrus that impairs its ability to sustain baseline neurogenesis or the injury itself may accelerate depletion of the overall progenitor pool. Either possibility suggests that recurrent injuries may not result in such a robust neurogenic response and that this ability to self-repair may be limited.
A number of strategies have emerged to test whether hippocampal neurogenesis has physiologic relevance. Methods to inhibit neurogenesis include systemic or local administration of anti-mitotic agents such as AraC that are known to nonspecifically target all dividing cells, including neural progenitors (Doetsch et al., 1999; Lau et al., 2009). Another is to perform cranial irradiation directed to neurogenic zones such as the subventricular zone and dentate gyrus to more selectively impair neurogenesis by better controlling what cells are exposed (Naylor et al., 2008; Clelland et al., 2009; Hellström et al., 2009; Noonan et al., 2010). Problems with these approaches include both their lack of specificity and potentially toxic side effects that lead to unwanted immune activation that may affect other mediators of recovery or damage not related to neurogenesis. Thus far, the data supporting a role for adult neurogenesis in improving cognitive function in the uninjured state are compelling but rely almost exclusively on these various imperfect techniques.
Recently, genetically engineered mice have become available that can regulate neurogenesis in a more precise and temporally controlled manner. One strategy is to inducibly express diphtheria toxin or its receptor in progenitor cells, which can then be ablated in a temporally controlled manner (Luquet et al., 2005; Durieux et al., 2009). One confounder with this approach is that all progenitors, not just dividing ones, are ablated and therefore the pool becomes depleted and cannot be reactivated. An alternative approach is a transgenic mouse that expresses HSV–TK under the control of the GFAP promoter. This allows for the inducible ablation of dividing cells that express GFAP, which includes early type 1 hippocampal progenitor cells as well as dividing reactive astrocytes (Sofroniew et al., 1999; Morshead et al., 2003; Saxe et al., 2006). Recently, more specific modified versions of HSV–TK have been shown in nestin-expressing progenitor cells to inducibly ablate early progenitors during normal states as well as after injury (Yu et al., 2008; Clelland et al., 2009).
It remains both controversial and unclear whether adult hippocampal neurogenesis contributes to memory formation during normal adult brain maturation. Some of the conflicting results regarding the impact of neurogenesis on memory have been attributed to the heterogeneous manner in which ablation occurs (Deng et al., 2010). However, even with the most recently developed genetic techniques, there remains evidence for hippocampal-based Morris water maze and contextual fear conditioning deficits in some models of ablated neurogenesis but not in others (Dupret et al., 2008; Imayoshi et al., 2008; Zhang et al., 2008; Deng et al., 2009). In our present study, we note no differences in fear conditioning in mock-injured animals with or without neurogenesis intact (Fig. 5) and only very modest differences in Morris water maze-based spatial memory (Figs. 6, 7). Although contextual fear conditioning is generally thought to be a hippocampal-dependent behavior, it is possible that other brain regions required for this task, such as the amygdala (Goosens and Maren, 2001; Onishi and Xavier, 2010), were able to compensate and successfully support the contextual fear learning and memory. For example, even if mice were unable to develop an integrated representation of the context, it is possible that they were able to use a combination of discrete cues in the environment to remember specific aspects of the learning context. In addition, the dentate gyrus has been recently linked to pattern separation within the hippocampus (Deng et al., 2010), and neither contextual fear conditioning nor our Morris water maze task specifically tested pattern separation. Our Morris water maze task might not have been sensitive enough to detect the subtle differences that ablation of neurogenesis impart on hippocampal function, and it is possible that we might detect a stronger effect of neurogenesis ablation with more challenging behavior parameters or a spatial learning task that expressly tests pattern separation. Although the most recent studies do suggest functional roles for adult neurogenesis within some aspects of memory formation, they may become most readily apparent when the experimental animals undergo induced induction of neurogenesis with an acquired injury as demonstrated here (Deng et al., 2010).
Although injury-induced neurogenesis clearly occurs in response to a wide variety of injuries, the mechanisms underlying it need more investigation. The injured brain releases numerous extracellular proteins and ions that may play roles in regulating neurogenesis. Two of the best studied of these, potassium chloride and glutamate, have been implicated in enhancement of proliferation in immature cells while at the same time directing toxicity in more mature cell types (Shi et al., 2007; Mattson, 2008). In addition, the injured brain activates both astrocytes and microglia, which are both known to secrete a variety of growth factors as well as immune modulators that may affect progenitor proliferation and survival (Myer et al., 2006; Bessis et al., 2007). Finally, the progenitor cells themselves make physical contact with the vasculature so circulating factors such as cytokines and growth factors that increase after injury may also direct some of these effects (Mignone et al., 2004). Thus, the mechanisms underlying TBI-induced neurogenesis are likely not straightforward nor easily worked out and therefore remain compelling targets to study, particularly because our data suggest that blockade of such neurogenesis can exacerbate the cognitive deficits results from traumatic brain injury.
Our data show that pharmacogenetic ablation of neurogenesis after brain injury results in impaired learning and memory in the Morris water maze, but ablation of neurogenesis did not modulate the effect of brain injury on motor coordination and learning (as assessed by the rotarod) or fear learning and memory (as assessed by cued and contextual fear conditioning). It is not surprising that the influence of neurogenesis ablation on the behavioral effects of brain injury was limited to spatial learning and memory, given that the most robust neurogenesis within rodents is known to occur within the hippocampus. Although additional study is needed to determine why neither the ablation of neurogenesis nor brain injury alone affected contextual fear conditioning, which is generally thought to be a hippocampal-dependent behavior, it is possible that there was compensation of other brain regions (as discussed above). There is also evidence that different subregions of the hippocampus are required for learning in the contextual fear conditioning and Morris water maze tasks (Richmond et al., 1999), and this could account for the different results in these two tasks. In addition, the modeling demonstrated here affects subventricular zone neurogenesis as well. Although it is less likely that these progenitors contribute to the hippocampal-specific behaviors tested here, it remains formally possible that our observations are confounded by ablation of subventricular zone progenitors.
In multiple measures of learning and memory in the Morris water maze, brain-injured mice lacking neurogenesis (Injured + Gan) exhibited impaired performance compared with all control mice, including mice with either brain injury alone or ablation of neurogenesis alone. As discussed in Results, brain-injured mice lacking neurogenesis (Injured + Gan) also showed increased thigmotaxic behavior during learning in the Morris water maze, suggesting that these mice were unable to progress from the initial use of a thigmotaxic search strategy to the use of an efficient spatial strategy to find the hidden platform (Lipp and Wolfer, 1998; Powell et al., 2004). Although we interpret this as additional evidence for a learning deficit, we acknowledge that we cannot conclusively rule out the possibility that the increased thigmotaxic behavior could be an indication of other possible cognitive deficits, such as increased anxiety-like behavior. Even if that is the case, however, our data still demonstrate that ablation of adult neurogenesis exacerbates the general cognitive deficits resulting from brain injuries and suggest that enhancing hippocampal neurogenesis after injury might be a viable therapeutic strategy.
Footnotes
This work was supported by National Institutes of Health Grant R01 NS048192 (S.G.K.), the Seay Endowed Fund for Research on Brain and Spinal Cord Injuries in Children (S.G.K.), National Institute of Mental Health (C.M.P.), National Institute of Child Health and Human Development (C.M.P.), and The Hartwell Foundation (C.M.P.).
The authors declare no competing financial interests.
References
- Aguirre A, Gallo V. Postnatal neurogenesis and gliogenesis in the olfactory bulb from NG2-expressing progenitors of the subventricular zone. J Neurosci. 2004;24:10530–10541. doi: 10.1523/JNEUROSCI.3572-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez-Buylla A, García-Verdugo JM, Tramontin AD. A unified hypothesis on the lineage of neural stem cells. Nat Rev Neurosci. 2001;2:287–293. doi: 10.1038/35067582. [DOI] [PubMed] [Google Scholar]
- Belachew S, Chittajallu R, Aguirre AA, Yuan X, Kirby M, Anderson S, Gallo V. Postnatal NG2 proteoglycan-expressing progenitor cells are intrinsically multipotent and generate functional neurons. J Cell Biol. 2003;161:169–186. doi: 10.1083/jcb.200210110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bessis A, Béchade C, Bernard D, Roumier A. Microglial control of neuronal death and synaptic properties. Glia. 2007;55:233–238. doi: 10.1002/glia.20459. [DOI] [PubMed] [Google Scholar]
- Chen J, Kwon CH, Lin L, Li Y, Parada LF. Inducible site-specific recombination in neural stem/progenitor cells. Genesis. 2009;47:122–131. doi: 10.1002/dvg.20465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen XH, Iwata A, Nonaka M, Browne KD, Smith DH. Neurogenesis and glial proliferation persist for at least one year in the subventricular zone following brain trauma in rats. J Neurotrauma. 2003;20:623–631. doi: 10.1089/089771503322144545. [DOI] [PubMed] [Google Scholar]
- Chirumamilla S, Sun D, Bullock MR, Colello RJ. Traumatic brain injury induced cell proliferation in the adult mammalian central nervous system. J Neurotrauma. 2002;19:693–703. doi: 10.1089/08977150260139084. [DOI] [PubMed] [Google Scholar]
- Clelland CD, Choi M, Romberg C, Clemenson GD, Jr, Fragniere A, Tyers P, Jessberger S, Saksida LM, Barker RA, Gage FH, Bussey TJ. A functional role for adult hippocampal neurogenesis in spatial pattern separation. Science. 2009;325:210–213. doi: 10.1126/science.1173215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dash PK, Mach SA, Moore AN. Enhanced neurogenesis in the rodent hippocampus following traumatic brain injury. J Neurosci Res. 2001;63:313–319. doi: 10.1002/1097-4547(20010215)63:4<313::AID-JNR1025>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- Deng W, Saxe MD, Gallina IS, Gage FH. Adult-born hippocampal dentate granule cells undergoing maturation modulate learning and memory in the brain. J Neurosci. 2009;29:13532–13542. doi: 10.1523/JNEUROSCI.3362-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng W, Aimone JB, Gage FH. New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nat Rev Neurosci. 2010;11:339–350. doi: 10.1038/nrn2822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doetsch F, García-Verdugo JM, Alvarez-Buylla A. Regeneration of a germinal layer in the adult mammalian brain. Proc Natl Acad Sci U S A. 1999;96:11619–11624. doi: 10.1073/pnas.96.20.11619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan X, Kang E, Liu CY, Ming GL, Song H. Development of neural stem cell in the adult brain. Curr Opin Neurobiol. 2008;18:108–115. doi: 10.1016/j.conb.2008.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupret D, Revest JM, Koehl M, Ichas F, De Giorgi F, Costet P, Abrous DN, Piazza PV. Spatial relational memory requires hippocampal adult neurogenesis. PLoS One. 2008;3:e1959. doi: 10.1371/journal.pone.0001959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durieux PF, Bearzatto B, Guiducci S, Buch T, Waisman A, Zoli M, Schiffmann SN, de Kerchove d'Exaerde A. D2R striatopallidal neurons inhibit both locomotor and drug reward processes. Nat Neurosci. 2009;12:393–395. doi: 10.1038/nn.2286. [DOI] [PubMed] [Google Scholar]
- Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- Etherton MR, Blaiss CA, Powell CM, Südhof TC. Mouse neurexin-1alpha deletion causes correlated electrophysiological and behavioral changes consistent with cognitive impairments. Proc Natl Acad Sci U S A. 2009;106:17998–18003. doi: 10.1073/pnas.0910297106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia AD, Doan NB, Imura T, Bush TG, Sofroniew MV. GFAP-expressing progenitors are the principal source of constitutive neurogenesis in adult mouse forebrain. Nat Neurosci. 2004;7:1233–1241. doi: 10.1038/nn1340. [DOI] [PubMed] [Google Scholar]
- Ge S, Goh EL, Sailor KA, Kitabatake Y, Ming GL, Song H. GABA regulates synaptic integration of newly generated neurons in the adult brain. Nature. 2006;439:589–593. doi: 10.1038/nature04404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goosens KA, Maren S. Contextual and auditory fear conditioning are mediated by the lateral, basal, and central amygdaloid nuclei in rats. Learn Mem. 2001;8:148–155. doi: 10.1101/lm.37601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould E, Gross CG. Neurogenesis in adult mammals: some progress and problems. J Neurosci. 2002;22:619–623. doi: 10.1523/JNEUROSCI.22-03-00619.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellström NA, Björk-Eriksson T, Blomgren K, Kuhn HG. Differential recovery of neural stem cells in the subventricular zone and dentate gyrus after ionizing radiation. Stem Cells. 2009;27:634–641. doi: 10.1634/stemcells.2008-0732. [DOI] [PubMed] [Google Scholar]
- Imayoshi I, Sakamoto M, Ohtsuka T, Takao K, Miyakawa T, Yamaguchi M, Mori K, Ikeda T, Itohara S, Kageyama R. Roles of continuous neurogenesis in the structural and functional integrity of the adult forebrain. Nat Neurosci. 2008;11:1153–1161. doi: 10.1038/nn.2185. [DOI] [PubMed] [Google Scholar]
- Kawaguchi A, Miyata T, Sawamoto K, Takashita N, Murayama A, Akamatsu W, Ogawa M, Okabe M, Tano Y, Goldman SA, Okano H. Nestin-EGFP transgenic mice: visualization of the self-renewal and multipotency of CNS stem cells. Mol Cell Neurosci. 2001;17:259–273. doi: 10.1006/mcne.2000.0925. [DOI] [PubMed] [Google Scholar]
- Kee N, Teixeira CM, Wang AH, Frankland PW. Preferential incorporation of adult-generated granule cells into spatial memory networks in the dentate gyrus. Nat Neurosci. 2007;10:355–362. doi: 10.1038/nn1847. [DOI] [PubMed] [Google Scholar]
- Kernie SG, Parent JM. Forebrain neurogenesis after focal ischemic and traumatic brain injury. Neurobiol Dis. 2010;37:267–274. doi: 10.1016/j.nbd.2009.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kernie SG, Erwin TM, Parada LF. Brain remodeling due to neuronal and astrocytic proliferation after controlled cortical injury in mice. J Neurosci Res. 2001;66:317–326. doi: 10.1002/jnr.10013. [DOI] [PubMed] [Google Scholar]
- Kokovay E, Shen Q, Temple S. The incredible elastic brain: how neural stem cells expand our minds. Neuron. 2008;60:420–429. doi: 10.1016/j.neuron.2008.10.025. [DOI] [PubMed] [Google Scholar]
- Lagace DC, Whitman MC, Noonan MA, Ables JL, DeCarolis NA, Arguello AA, Donovan MH, Fischer SJ, Farnbauch LA, Beech RD, DiLeone RJ, Greer CA, Mandyam CD, Eisch AJ. Dynamic contribution of nestin-expressing stem cells to adult neurogenesis. J Neurosci. 2007;27:12623–12629. doi: 10.1523/JNEUROSCI.3812-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau BW, Yau SY, Lee TM, Ching YP, Tang SW, So KF. Intracerebroventricular infusion of cytosine-arabinoside causes prepulse inhibition disruption. Neuroreport. 2009;20:371–377. doi: 10.1097/WNR.0b013e328324edcd. [DOI] [PubMed] [Google Scholar]
- Li Y, Luikart BW, Birnbaum S, Chen J, Kwon CH, Kernie SG, Bassel-Duby R, Parada LF. TrkB regulates hippocampal neurogenesis and governs sensitivity to antidepressive treatment. Neuron. 2008;59:399–412. doi: 10.1016/j.neuron.2008.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipp HP, Wolfer DP. Genetically modified mice and cognition. Curr Opin Neurobiol. 1998;8:272–280. doi: 10.1016/s0959-4388(98)80151-7. [DOI] [PubMed] [Google Scholar]
- Luquet S, Perez FA, Hnasko TS, Palmiter RD. NPY/AgRP neurons are essential for feeding in adult mice but can be ablated in neonates. Science. 2005;310:683–685. doi: 10.1126/science.1115524. [DOI] [PubMed] [Google Scholar]
- Mattson MP. Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann N Y Acad Sci. 2008;1144:97–112. doi: 10.1196/annals.1418.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mignone JL, Kukekov V, Chiang AS, Steindler D, Enikolopov G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J Comp Neurol. 2004;469:311–324. doi: 10.1002/cne.10964. [DOI] [PubMed] [Google Scholar]
- Miles DK, Kernie SG. Hypoxic-ischemic brain injury activates early hippocampal stem/progenitor cells to replace vulnerable neuroblasts. Hippocampus. 2008;18:793–806. doi: 10.1002/hipo.20439. [DOI] [PubMed] [Google Scholar]
- Morshead CM, Garcia AD, Sofroniew MV, van Der Kooy D. The ablation of glial fibrillary acidic protein-positive cells from the adult central nervous system results in the loss of forebrain neural stem cells but not retinal stem cells. Eur J Neurosci. 2003;18:76–84. doi: 10.1046/j.1460-9568.2003.02727.x. [DOI] [PubMed] [Google Scholar]
- Myer DJ, Gurkoff GG, Lee SM, Hovda DA, Sofroniew MV. Essential protective roles of reactive astrocytes in traumatic brain injury. Brain. 2006;129:2761–2772. doi: 10.1093/brain/awl165. [DOI] [PubMed] [Google Scholar]
- Naylor AS, Bull C, Nilsson MK, Zhu C, Björk-Eriksson T, Eriksson PS, Blomgren K, Kuhn HG. Voluntary running rescues adult hippocampal neurogenesis after irradiation of the young mouse brain. Proc Natl Acad Sci U S A. 2008;105:14632–14637. doi: 10.1073/pnas.0711128105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noonan MA, Bulin SE, Fuller DC, Eisch AJ. Reduction of adult hippocampal neurogenesis confers vulnerability in an animal model of cocaine addiction. J Neurosci. 2010;30:304–315. doi: 10.1523/JNEUROSCI.4256-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Onishi BK, Xavier GF. Contextual, but not auditory, fear conditioning is disrupted by neurotoxic selective lesion of the basal nucleus of amygdala in rats. Neurobiol Learn Mem. 2010;93:165–174. doi: 10.1016/j.nlm.2009.09.007. [DOI] [PubMed] [Google Scholar]
- Powell CM, Schoch S, Monteggia L, Barrot M, Matos MF, Feldmann N, Südhof TC, Nestler EJ. The presynaptic active zone protein RIM1alpha is critical for normal learning and memory. Neuron. 2004;42:143–153. doi: 10.1016/s0896-6273(04)00146-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pozniak CD, Pleasure SJ. Genetic control of hippocampal neurogenesis. Genome Biol. 2006;7:207. doi: 10.1186/gb-2006-7-3-207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramaswamy S, Goings GE, Soderstrom KE, Szele FG, Kozlowski DA. Cellular proliferation and migration following a controlled cortical impact in the mouse. Brain Res. 2005;1053:38–53. doi: 10.1016/j.brainres.2005.06.042. [DOI] [PubMed] [Google Scholar]
- Richardson RM, Sun D, Bullock MR. Neurogenesis after traumatic brain injury. Neurosurg Clin N Am. 2007;18:169–181. xi. doi: 10.1016/j.nec.2006.10.007. [DOI] [PubMed] [Google Scholar]
- Richmond MA, Yee BK, Pouzet B, Veenman L, Rawlins JN, Feldon J, Bannerman DM. Dissociating context and space within the hippocampus: effects of complete, dorsal, and ventral excitotoxic hippocampal lesions on conditioned freezing and spatial learning. Behav Neurosci. 1999;113:1189–1203. doi: 10.1037/0735-7044.113.6.1189. [DOI] [PubMed] [Google Scholar]
- Ridet JL, Malhotra SK, Privat A, Gage FH. Reactive astrocytes: cellular and molecular cues to biological function. Trends Neurosci. 1997;20:570–577. doi: 10.1016/s0166-2236(97)01139-9. [DOI] [PubMed] [Google Scholar]
- Saxe MD, Battaglia F, Wang JW, Malleret G, David DJ, Monckton JE, Garcia AD, Sofroniew MV, Kandel ER, Santarelli L, Hen R, Drew MR. Ablation of hippocampal neurogenesis impairs contextual fear conditioning and synaptic plasticity in the dentate gyrus. Proc Natl Acad Sci U S A. 2006;103:17501–17506. doi: 10.1073/pnas.0607207103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seaberg RM, van der Kooy D. Adult rodent neurogenic regions: the ventricular subependyma contains neural stem cells, but the dentate gyrus contains restricted progenitors. J Neurosci. 2002;22:1784–1793. doi: 10.1523/JNEUROSCI.22-05-01784.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi J, Miles DK, Orr BA, Massa SM, Kernie SG. Injury-induced neurogenesis in Bax-deficient mice: evidence for regulation by voltage-gated potassium channels. Eur J Neurosci. 2007;25:3499–3512. doi: 10.1111/j.1460-9568.2007.05624.x. [DOI] [PubMed] [Google Scholar]
- Sofroniew MV, Bush TG, Blumauer N, Lawrence K, Mucke L, Johnson MH. Genetically-targeted and conditionally-regulated ablation of astroglial cells in the central, enteric and peripheral nervous systems in adult transgenic mice. Brain Res. 1999;835:91–95. doi: 10.1016/s0006-8993(99)01639-x. [DOI] [PubMed] [Google Scholar]
- Tabuchi K, Blundell J, Etherton MR, Hammer RE, Liu X, Powell CM, Südhof TC. A neuroligin-3 mutation implicated in autism increases inhibitory synaptic transmission in mice. Science. 2007;318:71–76. doi: 10.1126/science.1146221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Temple S, Qian X. bFGF, neurotrophins, and the control or cortical neurogenesis. Neuron. 1995;15:249–252. doi: 10.1016/0896-6273(95)90030-6. [DOI] [PubMed] [Google Scholar]
- Urrea C, Castellanos DA, Sagen J, Tsoulfas P, Bramlett HM, Dietrich WD. Widespread cellular proliferation and focal neurogenesis after traumatic brain injury in the rat. Restor Neurol Neurosci. 2007;25:65–76. [PubMed] [Google Scholar]
- Yamaguchi M, Saito H, Suzuki M, Mori K. Visualization of neurogenesis in the central nervous system using nestin promoter-GFP transgenic mice. Neuroreport. 2000;11:1991–1996. doi: 10.1097/00001756-200006260-00037. [DOI] [PubMed] [Google Scholar]
- Yokoyama A, Sakamoto A, Kameda K, Imai Y, Tanaka J. NG2 proteoglycan-expressing microglia as multipotent neural progenitors in normal and pathologic brains. Glia. 2006;53:754–768. doi: 10.1002/glia.20332. [DOI] [PubMed] [Google Scholar]
- Yu TS, Zhang G, Liebl DJ, Kernie SG. Traumatic brain injury-induced hippocampal neurogenesis requires activation of early nestin-expressing progenitors. J Neurosci. 2008;28:12901–12912. doi: 10.1523/JNEUROSCI.4629-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang CL, Zou Y, He W, Gage FH, Evans RM. A role for adult TLX-positive neural stem cells in learning and behaviour. Nature. 2008;451:1004–1007. doi: 10.1038/nature06562. [DOI] [PubMed] [Google Scholar]