

Abstract
The importance of intraepithelial lymphocytes (IEL) in immunoprotection against orally acquired pathogens is being increasingly recognized. Recent studies have demonstrated that Ag-specific IEL can be generated and can provide an important first line of defense against pathogens acquired via oral route. However, the mechanism involved in priming of IEL remains elusive. Our current study, using a microsporidial model of infection, demonstrates that priming of IEL is dependent on IFN-γ-producing dendritic cells (DC) from mucosal sites. DC from mice lacking the IFN-γ gene are unable to prime IEL, resulting in failure of these cells to proliferate and lyse pathogen-infected targets. Also, treatment of wild-type DC from Peyer’s patches with Ab to IFN-γ abrogates their ability to prime an IEL response against Encephalitozoon cuniculi in vitro. Moreover, when incubated with activated DC from IFN-γ knockout mice, splenic CD8+ T cells are not primed efficiently and exhibit reduced ability to home to the gut compartment. These data strongly suggest that IFN-γ-producing DC from mucosal sites play an important role in the generation of an Ag-specific IEL response in the small intestine. To our knowledge, this report is the first demonstrating a role for IFN-γ-producing DC from Peyer’s patches in the development of Ag-specific IEL population and their trafficking to the gut epithelium.
Cell-mediated immunity involving T cells plays an important role in immune response and survival against infections with obligate intracellular pathogens such as Encephalitozoon cuniculi, a member of the Microsporidia order associated with disseminated infections in mammalian hosts (1–4). Infections acquired via oral route (i.e., a gastrointestinal infection) usually evoke a strong mucosal response. This finding was recently demonstrated in a study conducted in our laboratory where oral infection with E. cuniculi led to an early and rapid intraepithelial lymphocyte (IEL)3 response against this organism (5). IEL from infected mice exhibited a strong Ag-specific immunity and were able to protect naive immunocompromised SCID mice against a lethal challenge with E. cuniculi.
IEL are considered nonconventional lymphocytes located among the epithelial cells in the lumen of the gut and are comprised predominantly of CD3+ T cells (6). Although IEL are known to produce cytokines, their importance in protective immunity against mucosal pathogens has only been recently recognized (7–10). Priming of an Ag-specific IEL response is not yet understood and the mechanism by which IEL provide protection against infectious agents remains somewhat enigmatic. In this study, factors essential for the induction of an Ag-specific IEL response were analyzed, and the mechanism by which they confer immunoprotection against an intracellular infection was evaluated.
Materials and Methods
Mice
Six- to 7-wk-old SCID, perforin knockout (pf−/−), and IFN-γ knockout (IFN-γ−/−) mice on a C57BL/6 genetic background were obtained from the Jackson Laboratory. Age- and sex-matched C57BL/6 controls were obtained from the National Cancer Institute (Frederick, MD). Animals were housed under approved conditions under an approved Institutional Animal Care and Use Committee protocol at the Animal Research Facility of the Louisiana State University Health Sciences Center (New Orleans, LA) and George Washington University Medical Center (Washington, DC).
Parasites and infection
A rabbit isolate of E. cuniculi (genotype II), provided by E. Didier (Tulane Regional Primate Center, Covington, LA), was used throughout the study. The parasites were maintained by continuous passage in rabbit kidney (RK-13) cells obtained from the American Type Culture Collection and in mice infected as previously described (5).
Histopathology
Mice were euthanized using an approved carbon dioxide technique, and liver, spleen, and small intestine were fixed in 10% Formalin and processed for 5-μm histological sections, which were subsequently stained with H&E using standard methods. Sections were also stained with chromotrope 2R (microsporidia) using the Kokoskin et al. (11) with a modification of the 1992 Weber microsporidia stain that uses a 10-min staining time in chromotrope 2R at 50°C. Stained slides were examined on a Nikon Microphot FXA microscope with a Nikon DS-L1 digital photography system.
CTL assay
The CTL assay was performed using the previously described procedure from our laboratory (5). Mouse peritoneal macrophages were harvested 2 days after i.p. injection of 1 ml of thioglycolate (Sigma-Aldrich), washed three times in PBS, and dispensed at a concentration of 5 × 104 cells/well into U-bottom 96-well plates. After an overnight incubation, these macrophages were infected with 2.5 × 105 E. cuniculi spores per well for 48 h and then washed extensively with PBS to remove extracellular parasites. Macrophages were then labeled with 51Cr (0.5 μCi/well) and incubated with IEL at various E:T ratios. Purification of IEL was conducted according to previously described protocol (5). The microtiter plate was then centrifuged at 200 × g for 3 min and incubated at 37°C for 4 h, and samples were removed and assayed for released 51Cr using scintillation counting. The percentage of lysis was calculated as (mean cpm of test sample – mean cpm of spontaneous release)/(mean cpm of maximal release – mean cpm of spontaneous release) × 100.
When indicated, IEL were incubated with 10 nM concanamycin A (CMA; Sigma-Aldrich) for 2 h at 37°C or 10 μg/ml anti-Fas ligand (FasL; eBioscience) for 20 min before their addition to the 51Cr-labeled macrophages (12).
IEL phenotyping for cytotoxic markers
IEL from infected wild-type (WT) animals were isolated at day 7 postinfection (p.i.) and labeled for CD95 ligand (CD95L), perforin, and granzyme B expression. Briefly, the cells were incubated for 4 h with Golgi stop, and labeled with anti-CD8α FITC-conjugated Ab (eBioscience). After permeabilization and fixation with Cytofix kit from BD Pharmingen, according to the manufacturer’s protocol, the cells were incubated with anti-CD95L anti-perforin, or anti-granzyme B PE-conjugated Abs (eBioscience). The cells were analyzed on FACSCalibur (BD Biosciences).
IEL adoptive transfer
IEL from pf−/−, IFN-γ−/−, or WT donors were isolated at day 7 p.i. and transferred to naive SCID animals as previously described (5). Recipients were challenged by oral infection at 48 h post transfer with 2 × 107 E. cuniculi spores. Transferred SCID mice were monitored for morbidity and mortality on a daily basis until the termination of the experiment. For one set of experiments, the CD8 subsets of IEL were purified from naive or infected WT mice before transfer to SCID mice. After IEL purification, the cells were separated in two subsets of CD8β+ and CD8β− cells by magnetic cell sorting with biotin-conjugated anti-CD8β Ab, followed by anti-biotin-coated microbeads (Miltenyi Biotec), according to manufacturer’s instructions (purity ≥94%). The different cell suspensions were adoptively transferred i.p. to SCID recipients. Two days after the transfer, the mice were challenged orally with 2 × 107 E. cuniculi spores and the recipient mice were monitored as described.
For IEL trafficking experiments, SCID mice were adoptively transferred with IEL labeled with CellTrace CFSE proliferation kit, according to the manufacturer’s instructions (Invitrogen Life Technologies). The animals were then challenged as described. Recipient mice were sacrificed 2 days post transfer, and IEL, spleen, and mesenteric lymph node (MLN) lymphocytes were isolated and analyzed for CFSE expression by flow cytometry.
Phenotypic analysis of IEL
WT and IFN-γ−/− mice were orally infected with 2 × 107 E cuniculi spores. At day 7 p.i., purified IEL were labeled for surface markers CD8α, CD8α, TCRαβ, and TCRγδ (eBioscience) as previously described (5). Cells were acquired on a FACSVantage (BD Biosciences).
DC purification and in vitro priming of IEL
DC from Peyer’s patches (PP) were prepared according to an already published protocol (13). Briefly, PP from WT and IFN-γ−/− mice were enzymatically and mechanically disrupted. After a single-cell suspension was prepared, the cells were incubated with anti-CD11c-coated microbeads (Miltenyi Biotec) and separated on MACS separation columns according to the manufacturer’s instructions. Cells were then stained with PE-labeled anti-CD11c and FITC-labeled anti-CD19 Ab (eBioscience). CD11c+/CD19− cells were sorted on a FACSVantage. Purified cells (96–98% pure as determined by flow cytometry) were plated at various concentrations (5–20 × 103) and infected overnight with 20 E. cuniculi spores/DC. After several washes, 104 purified IEL were added to the DC. At 72 h later, 1 μCi [3H]thymidine was added to each well, and proliferative response was determined as described elsewhere (14).
When indicated, different concentrations of anti-IFN-γ Ab (2 and 0.5 μg/ml; R&D Systems) were added to the DC culture overnight at the time of infection. The next day, the cells were washed and the IEL added as previously described.
In vitro IL-12 production by DC
DC were isolated from PP of IFN-γ−/− and WT mice as described. Cells were plated in 96-well plates at concentrations of 5 × 104 cells per well and infected with E. cuniculi (20 spores/DC). After overnight incubation, supernatants were collected and assayed for IL-12 production by cytokine ELISA kit (BioLegend), according to the manufacturer’s instructions.
In vitro priming of splenic CD8αβ T cells by DC from PP
DC from PP of WT and IFN-γ−/− mice were isolated and infected as described. After overnight incubation, magnetically purified, splenic CD8αβ+ T cells (>95% pure) were added to the culture and 72 h later, cells were harvested and labeled with CFSE (25 μM) according to manufacturer’s instructions (Invitrogen Life Technologies). Labeled cells were transferred to naive WT mice (2 × 106 cells/mouse) and 24 h later the recipients were sacrificed. Tissues (spleen, MLN, PP, and IEL) were analyzed for the presence of donor CFSE+ CD3+ cells by FACS analysis.
In another series of experiments, splenic CD8αβ+ T cells were recovered after 3 days of incubation with DC from PP of WT or IFN-γ−/−. The cells were labeled with anti-CD8β allophycocyanin-conjugated and anti-CD8α Pacific blue-conjugated Abs (eBioscience) and surface expression of CCR9 was evaluated by FACS analysis using FITC-conjugated Ab (R&D Systems). IFN-γ production by these cells was analyzed after intracellular staining with PE-conjugated anti-IFN-γ Ab (eBioscience). Data were acquired on FACSAria (BD Biosciences) using Diva software.
Results
IFN-γ−/− and pf−/− mice are susceptible to oral E. cuniculi challenge
Previous data from our laboratory demonstrated that oral E. cuniculi infection induced a strong IEL response in the host (5). IEL from infected animals produced IFN-γ and exhibited ex vivo cytotoxic activity against infected targets. In the present study, gene knockout mice were used to determine the importance of IFN-γ production and cytolytic response in the protective immune mechanism against oral microsporidial infection. Mice lacking IFN-γ or perforin gene were orally infected with 2 × 107 E. cuniculi spores. Knockout mice started to die on day 13 p.i. and all the mice in IFN-γ−/− and pf−/− group succumbed to the infection by day 28 postchallenge. Conversely, as expected, WT mice survived till the termination of the experiment (Fig. 1A). Both IFN-γ−/− and pf−/− mice infected orally with E. cuniculi were subjected to histopathological analysis. IFN-γ−/−, pf−/−, and WT mice were sacrificed on day 10 p.i. (representative tissue sections are shown in Fig. 1B). On necroscopy, the spleen of WT mice was normal in size and appearance (Fig. 1B, i), whereas that of the pf−/− mice was one and a half its normal size and demonstrated mild effacement of the follicles (data not shown) and that of IFN-γ−/− mice was four times its normal size with marked effacement of the follicles and expansion of the red pulp (Fig. 1B, ii and iii). The liver of the WT and pf−/− mice was essentially normal with occasional inflammatory mononuclear cell foci (Fig. 1B, iv), whereas that of the IFN-γ−/− mice demonstrated numerous inflammatory foci (Fig. 1B, v and vi). Intestinal biopsy demonstrated a mild increase in PP size in both pf−/− and IFN-γ−/− animals compared with the control group (Fig. 1B, vii–ix). The increased inflammation in the tissues of knockout mice is suggestive of uncontrolled parasite replication and dissemination from the gastrointestinal tract following oral infection. The findings in the liver and spleen are similar to the pathology seen following i.p. infection of WT animals (1). These results suggest that both IFN-γ production and the cytotoxic response are important for protection (and the prevention of widespread dissemination) following oral infection with E. cuniculi.
FIGURE 1.

IFN-γ−/− and pf−/− mice are susceptible to oral E. cuniculi infection. A, IFN-γ−/−, pf−/−, and WT mice (n = 8 mice/group) were orally infected with 2 × 107 E. cuniculi spores. The animals were monitored daily for survival. The study was performed twice and data are representative of two separate experiments. B, Histopathology of infected mice. Spleen at necroscopy at magnification ×10 (i). WT mouse model demonstrating normal architecture. Spleen of IFN-γ−/− mouse at magnification ×20 (ii) and ×40 (iii), demonstrating effacement of follicles and increase in red pulp. Liver at magnification ×20 (iv). WT mouse model demonstrating normal architecture. Liver IFN-γ−/− mouse at magnification ×20 (v) and ×40 (vi), demonstrating inflammatory foci. Mononuclear cell infiltration into the liver (vi) associated with a slight increase in the number of Kupfer cells in the adjacent areas. PP in the intestinal track of mice are shown (vii–ix). There is some increased activity of lymphoid aggregates. However, no organism was detected. PP from IFN-γ−/− mouse at magnification ×10 (vii) and ×40 (viii). PP at magnification ×20 (ix) from a pf−/− mouse are shown.
Ag-specific cytotoxic activity of IEL is perforin-dependant
Data obtained suggested the importance of perforin-dependent cytotoxic activity in resolving oral E. cuniculi infection. To further assess the molecules involved in cytotoxic pathway, we analyzed IEL for presence of cytotoxic molecules perforin, granzyme B, or CD95L by flow cytometry. IEL from WT mice were isolated at 7 days p.i., and intracellular staining was performed. As shown in Fig. 2A, perforin, granzyme B, and CD95L expression were up-regulated after infection with E. cuniculi compared with IEL isolated from naive mice. To establish that an Ag-specific cytotoxic response exhibited by IEL is exclusively mediated by the perforin/granzyme pathway, blocking assay was conducted. IEL from immune mice were isolated and an inhibition of cytotoxicity assay using CMA or anti-FasL Ab was performed. CMA is a known inhibitor of the perforin/granzyme pathway and anti-FasL Abs inhibit the Fas/FasL pathway (12). IEL were purified at day 7 p.i., incubated with either CMA or anti-FasL, and their cytolytic activity against infected macrophages measured. As expected, naive IEL did not exhibit any detectable lytic activity against the infected target cells (Fig. 2). On the contrary, immune IEL displayed close to 40% lysis of infected targets at an E:T ratio of 20:1, which upon incubation with CMA was reduced by 5-fold (8% vs 40%; p = 0.0049) (Fig. 2B). Treatment with anti-FasL did not affect the CTL response of immune IEL. This result suggests that the cytotoxic mechanism of IEL is mediated mainly by perforin.
FIGURE 2.

Cytotoxic activity of IEL against E. cuniculi-infected target is perforin-dependent. IEL from WT mice were purified at day 7 p.i. (n = 6 mice/group). A, IELs were labeled for cytotoxic markers (perforin, CD95L, and granzyme B). Cells were isolated at day 7 p.i., and expression of perforin, granzyme B, and CD95L were analyzed by intracellular staining. B, Before incubation with 51Cr-labeled uninfected and infected macrophages (E:T ratio of 20:1), IEL were treated with either CMA or anti-FasL Abs. After 4 h of incubation, the cytolytic activity was determined by radioisotope release into the culture supernatant. The experiment was performed twice with similar results. Data are representative of one set of experiments.
IEL from pf−/− and IFN-γ−/− mice are unable to protect SCID recipients against oral challenge with E. cuniculi
To further establish that IFN-γ production and the cytotoxic effect of IEL are important for immune protection against oral E. cuniculi infection, adoptive transfer studies were performed. IEL isolated from pf−/−- or IFN-γ−/−-infected mice were transferred to SCID recipients, which were subsequently challenged with E. cuniculi spores (Fig. 3A). As previously observed, immune IEL from WT mice were able to partially protect SCID mice against a lethal E. cuniculi challenge (5). However, IEL from both pf−/− and IFN-γ−/− mice were unable to protect SCID recipients and all the animals were dead by day 35 p.i. In addition, none of the SCID recipients injected with naive IEL survived (data not shown). To analyze the migrating pattern of IEL in recipient animals, CFSE-labeled IEL isolated from pf−/−, IFN-γ−/−, and WT animals were adoptively transferred to SCID mice. The recipient animals were challenged 1 day after transfer. Two days p.i., the animals were sacrificed, and the spleen, MLN, or IEL compartment was analyzed for the presence of CFSE+ cells. Similar to our previously published data (5), the majority of cells isolated from WT or knockout animals migrated back to the MLN and some of them were recovered from the IEL compartment. A very low number of CFSE+ cells was detected in the spleen (data not shown). These results suggest that elicitation of protective IEL response against oral E. cuniculi infection is dependant on IFN-γ and the cytotoxic function of these cells.
FIGURE 3.

Adoptive transfer of IEL isolated from pf−/− and IFN-γ−/− mice fails to protect SCID mice against E. cuniculi challenge. A, Adoptive transfer of IEL from IFN-γ−/− and pf−/− is unable to protect SCID mice against a lethal challenge with E. cuniculi. IEL from naive and infected pf−/−, IFN-γ−/−, and WT mice (n = 12 mice/group) were isolated at day 7 p.i. A total of 5 × 106 IEL were injected i.p. into SCID mice (n = 6 mice/group). After 48 h, the mice were challenged orally with 2 × 107 spores of E. cuniculi and the survival of the animals was monitored daily until the end of the experiment. B, IEL from WT mice were purified at day 7 p.i. CD8β+ and CD8β− subset were separated and adoptively transferred to SCID recipients (n = 6 mice/group). The recipient animals were orally infected with 2 × 107 E. cuniculi 2 days posttransfer. The survival of the animals was monitored daily until the end of the experiment.
Previous data from our laboratory demonstrated that most of IFN-γ-producing and cytotoxic IEL induced during E. cuniculi infection had a CD8αβ phenotype (5). As both of these properties appear to be critical for host survival against E. cuniculi infection, transfer of CD8αβ IEL alone should ensure the transfer of protection to naive host. To validate this hypothesis, CD8β+ and CD8β− IEL from WT mice were purified and transferred to SCID animals. Twenty-four hours later, the recipients were challenged orally and monitored for morbidity and mortality. As shown in Fig. 3B, CD8β+ donor IEL conferred a high degree of protection to SCID recipients, and 80% of the animals that received these cells survived a lethal E. cuniculi challenge. Conversely, all the SCID mice injected with CD8β− IEL from immune mice succumbed to infection.
Defective induction of IEL response in IFN-γ−/− mice following E. cuniculi infection
These studies demonstrate an important role for IFN-γ in IEL-mediated protection against E. cuniculi. To further understand the role of IFN-γ, a phenotypic analysis of IEL from infected IFN-γ−/− and WT mice was performed at day 7 p.i. Similar to our previous observations, WT mice showed a strong induction of CD8αα, CD8αβ, TCRαβ, and TCRγδ IEL in response to E. cuniculi infection (Fig. 4A). However, a significantly lower increase (p < 0.05) in all the subpopulations of IEL was observed in the infected IFN-γ−/− mice.
FIGURE 4.

IFN-γ−/− mice exhibit a suboptimal IEL response against E. cuniculi infection. IEL from IFN-γ−/− and WT mice (n = 6 mice/group) were isolated 7 days p.i. A, Induction of IEL response in IFN-γ−/− mice following oral E. cuniculi infection. IFN-γ−/− and WT mice orally infected with E. cuniculi were sacrificed at day 7 p.i., and IEL were purified (n = 4 mice/group). Isolated IEL were assayed for CD8αα, CD8αβ, TCRαβ, and TCRγδ expression by FACS analysis. Data represent the mean ± SD of two individual sets of experiments. B, Cytotoxic activity of IEL against E. cuniculi-infected macrophages. IEL from IFN-γ−/− and WT mice were incubated with 51Cr-labeled uninfected and infected macrophages at different E:T ratio (20:1, 40:1). After 4 h of incubation, the cytolytic activity was determined by radioisotope release into culture supernatant. The experiment was performed twice with similar results and data are representative of one experiment.
To establish that the lack of IFN-γ leads to a failure of the Ag-specific IEL response in E. cuniculi-infected animals, functional assay was performed. IEL from WT and IFN-γ−/− mice were isolated at day 7 p.i. and assayed for Ag-specific cytotoxic activity. As shown in Fig. 4B, IEL from IFN-γ−/− mice exhibited a severe defect in induction of Ag-specific cytotoxic response as minimal lytic activity (p < 0.01) against infected targets was observed (Fig. 4B). Predictably, immune IEL from WT mice exhibited a strong cytotoxic activity against infected targets (30 and 45% at E:T ratio of 1:20 and 1:40, respectively). As expected, IEL from naive WT and IFN-γ−/− mice were unable to lyse E. cuniculi-infected macrophages.
In vitro priming of IEL by DC from PP
It has been recently reported that IFN-γ production by DC is very important for their ability to process Ags (15–19). Next, we determined whether the Ag-specific IEL response by DC can be induced and whether IFN-γ is necessary for this process. As PP is a major site for induction of the mucosal immune response (20–22), DC from this site were isolated. Cells from naive IFN-γ−/− and WT mice were plated and infected overnight with E. cuniculi spores. Following extensive washing, IEL from WT mice were added to the culture and after 72 h, the proliferation of IEL was measured by overnight [3H]thymidine incorporation. As demonstrated in Fig. 5A, at a concentration of 5–20 × 103 cells per well, DC from WT mice induced a significant proliferation of IEL (p ≤ 0.015). Conversely, IEL incubated with DC from IFN-γ−/− animals exhibited minimal proliferation, which was not significantly higher than the control wells containing no effector cells (Fig. 5A).
FIGURE 5.
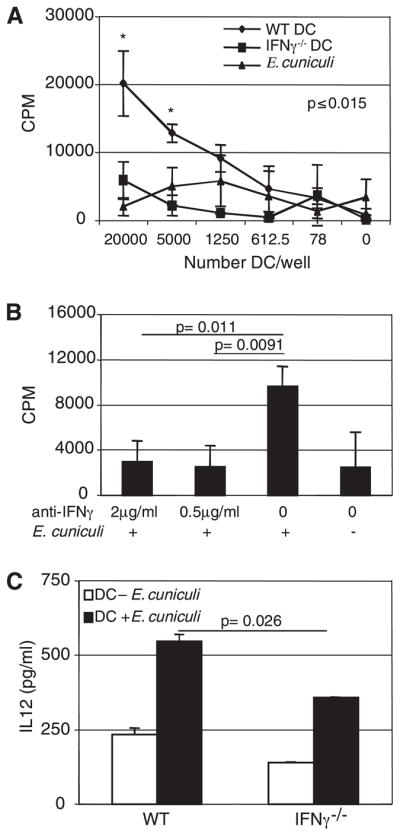
DC from PP of IFN-γ−/− mice are unable to stimulate naive CD8αβ IEL. Immature DC from naive IFN-γ−/− and WT mice (n = 6 mice/group) were isolated, plated, and infected overnight with E. cuniculi. The next day, naive IEL purified from WT mice (n = 6 mice/group) were separated and CD8αβ IEL were added to the culture. A, Presentation was measured as induction of proliferation of CD8αβ IEL. B, DC were treated with anti-IFN-γ or isotype control Ab at the same time that E. cuniculi infection and for the remaining of the experiment. C, IL-12 production in the supernatants of E. cuniculi primed DC from WT and knockout animals, after overnight incubation, was measured by ELISA. Experiments were performed twice with similar results and data are representative of one experiment.
To establish the importance of IFN-γ in Ag-specific priming of IEL, DC from WT mice were incubated overnight with murine anti-IFN-γ Ab. IEL were added 24 h later, and proliferation was measured. As shown in Fig. 5B, anti-IFN-γ treatment of DC inhibited their ability to present E. cuniculi Ag to IEL as Ab treatment led to a 3-fold reduction in the proliferation of IEL (p ≤ 0.02).
Because activated DC are known to release IL-12 in response to various infectious agents (23), in vitro IL-12 production by E. cuniculi-primed DC was measured. Purified DC from naive WT and IFN-γ−/− mice were cultured and pulsed with E. cuniculi spores. After 24 h of incubation, the supernatants were collected and assayed for IL-12 production by ELISA. As shown in Fig. 5C, compared with DC from WT mice, cells isolated from IFN-γ−/− animals release significantly less IL-12 in response to E. cuniculi stimulation (p = 0.026).
These studies demonstrated that IFN-γ produced by DC from PP acts in an autocrine manner and plays an important role in their activation. Recent studies using an OVA Ag model have demonstrated that DC from PP selectively imprint homing receptors on the splenic T cells, priming them and enabling them to migrate preferentially to the small intestine (24). To establish whether IFN-γ production by DC from PP is involved in the selective priming and subsequent migration of T cells to the gut, CD8+ splenocytes from naive animals were purified and incubated with E. cuniculi activated DC from PP of WT or IFN-γ−/− mice. After 72 h of incubation, the cells were collected and labeled with CFSE. A total of 2 × 106 labeled cells were administered to naive WT animals via i.p. route, recipients were sacrificed 24 h later, and tissues (spleen, MLN, PP, IEL) were assayed for the presence of donor CFSE+ CD3+ cells. As shown in Fig. 6, incubation of CD8+ T cells with DC from PP of WT mice led to greater migration to the gut (p ≤ 0.02). Conversely, CD8+ T cells incubated with the DC of IFN-γ−/− animals had significantly reduced capability to traffic to this site, demonstrating the importance of IFN-γ produced by PP DC in the priming process of Ag-specific IEL response. To further assess the level of activation of the splenic CD8β+ before transfer, cells were analyzed for CCR9 expression, a chemokine receptor specific for gut tropism (25), and IFN-γ production. CD8αβ+ T cells from the spleen of naive animals were isolated and primed as described. After 72 h of incubation, the cells were harvested and labeled for surface CCR9 and intracellular IFN-γ expression, as described in Materials and Methods. CD8αβ+ T cells primed by DC from PP of IFN-γ−/− mice exhibited significantly lower level of CCR9 (Fig. 6, C–E) and reduced IFN-γ production (Fig. 6, F–H) when compared with cells primed by DC of WT mice. These studies demonstrate that IFN-γ plays a critical role in the initiation of IEL response during E. cuniculi infection.
FIGURE 6.

Splenic CD8αβ+ T cells primed with DC from IFN-γ−/− mice have impaired ability to traffic to the gut. A and B, Immature DC from PP of naive IFN-γ−/− and WT mice (n = 6 mice/group) were isolated and infected with E. cuniculi. After overnight incubation, purified CD8β+ splenocytes from naive WT mice (n = 4 mice) were added to the culture. Seventy-two hours later, cells were harvested and labeled with CFSE before transfer to WT recipient animals (n = 2 mice/group). Subsequently after 24 h, the recipients were sacrificed and cells from MLN, PP, spleen (A) and IEL (B) recovered and analyzed for the presence of CFSE-labeled CD3+ cells by flow cytometry. C–H, CD8β+ splenocytes isolated from naive mice were primed with PP DC from WT or IFN-γ−/−. After 72 h, CD8β+ cells were labeled for CCR9 expression (C–E) or IFN-γ production by intracellular staining (F–H). Data in dot plots are gated on double positive CD8α+CD8β+ T cells primed with DC from WT (D–G) or IFN-γ−/− (E–H) mice labeled with allophycocyanin-conjugated anti-CD8α and PE-conjugated anti-CCR9 (D and E) or IFN-γ (G and H). Percentage represents surface expression of CCR9 (D and E) and IFN-γ production (G and H) in CD8αβ cells. Experiments were performed twice with similar results and data are representative of one experiment.
Discussion
Previous studies from our laboratory reported a vigorous IEL response during E. cuniculi infection (5). The data presented in this study demonstrate that strong Ag-specific IEL immune response is dependent on IFN-γ-producing mucosal DC. IEL from mice lacking the IFN-γ gene failed to exhibit a robust IEL response and thus were unable to protect naive animals against a lethal challenge. Similarly, treatment of DC from WT mice with anti-IFN-γ Ab neutralized their ability to prime IEL population, and the cells subsequently failed to exhibit Ag-specific proliferation. Our observation also strongly suggest that IFN-γ is involved in the activation of DC in PP in an autocrine manner, which subsequently play an important role in the generation of Ag-specific IEL response in the small intestine. Moreover, IFN-γ production of DC from PP is important both for induction of Ag-specific IEL response and selective migration of primed T cells to gut compartment.
Although the involvement of Ag-specific cytotoxic IEL has been reported in various infectious disease models (10, 26, 27), the mechanism of killing by these IEL has not been described. Recent studies have demonstrated that CTL killing is dependent mostly on two mechanisms. The first mechanism is mediated by Fas-FasL interaction (28) and exocytosis of lytic granules and the second is via perforin, a molecule present in the cytoplasmic granules of the CTLs (29). Perforin release causes the formation of pores on the cellular membrane of target cells permitting the penetration of granzymes into their cytoplasm leading to the apoptosis of the target cell (30, 31). Although increase in all cytotoxic molecules was observed, perforin mediated lysis is apparently the main effector mechanism by which IEL lyse infected cells.
Another important observation made in the current study is the in vitro proliferation of IEL in response to antigenic stimulation. It is generally believed that IEL exhibit phenotypic and functional characteristic of partially activated T cells and these cells do not proliferate as well as other cells (6, 32). Mosley et al. (33) demonstrated that IEL expressing TCRαβ or TCRγδ are largely unresponsive to proliferative signals mediated by the conventional stimulation of the CD3-TCR complex. Our findings are important since this report is the first of the in vitro proliferation studies of IEL in response to Ag-pulsed DC stimulation. It will be very interesting to determine whether a similar finding can be observed with other microbial agents in which IEL are known to play an important role in immune protection.
DC are the most potent professional APC with a unique ability to prime naive T cells (34, 35). They play an important role in the initiation and regulation of the immune response (6, 36). In the PP, DC are separated from the intestinal lumen by only a single layer of cells, the follicle associated epithelium retaining Ag-transporting microfold cells (M cells). In addition, a small number of DC are located directly within the follicle associated epithelium (37–39). Due to their location, these cells are likely to play a crucial role in the induction of immunity to infections acquired via an oral route. Moreover, detection of Listeria and Salmonella in DC from PP per oral infection suggests an important role for these cells in the mucosal protection against pathogens (40, 41). In the present study, we have demonstrated that DC from PP are important for the priming of IEL in vitro. Our data provide strong supportive evidence to the common belief that IEL are primed in the PP (36, 42) and then migrate to the epithelium were they can protect the mucosa against further infections (43). Moreover, recent studies with reovirus have demonstrated that intraduodenal infection in mice led to a rapid increase in precursor CTL population in the PP and IEL compartment (44). Also in the same study, it was demonstrated that adoptive transfer of lymphocytes from PP to SCID mice resulted in the appearance of a large number of CD8+TCRαβ+ IEL. However, there are reports that most IEL populations develop independently of passage through the PP or MLN (45). The variation in these observations can be attributed to different developmental requirements for CD8αα and CD8αβ IEL. Although a recent report suggests that all gut IEL are from thymic origin (46), CD8αα IEL are believed to develop locally in cryptopatches independently of PP (47). Conversely, CD8αβ IEL seems to dependent on priming by DC from PP (24). Similar to these observations, in the current study we demonstrate that gut immunity against E. cuniculi infection is mediated by CD8αβ population of IEL subset and that DC from PP seem to be important for initiating this response.
Priming of IEL by DC appears to be highly dependent on IFN-γ because neutralization of this cytokine abrogates the induction of Ag-specific response. Moreover, increase in IEL levels during E. cuniculi infection in IFN-γ−/− mice was severely compromised and cells from these animals failed to lyse infected target. Recent studies have reported a high level of intracellular IFN-γ expression in IL-12-stimulated DC (16, 17, 48). Similar to these findings, our study suggests that IL-12-dependent IFN-γ production by DC is important for stimulation of IEL response against oral E. cuniculi infection. Moreover, although, the role of IFN-γ in induction of the systemic CTL response has been described in various infectious disease models (14, 49, 50), this study is the first to describe the importance of IFN-γ in the generation of Ag-specific IEL response against an oral pathogen.
Our data demonstrate that DC from PP are important for the trafficking of Ag-primed CD8+ T cells to the gut compartment. When primed with E. cuniculi-pulsed DC from PP, a larger population of adoptively transferred splenic CD8+ T cells ended up in the small intestine (MLN, PP, and IEL compartment) of the donor mice. However, when primed with DC from PP of IFN-γ−/− mice, a reduced population of CD8+ T cells was able to migrate to the gut. Also, CD8+ T cells primed with DC from PP of WT mice exhibit an up-regulation of CCR9, a receptor for chemokine responsible for trafficking of lymphocytes to the gut (25). Conversely, no increase in CCR9 expression was observed on CD8αβ T cells when primed with DC from PP of IFN-γ−/− mice. These findings are supported by a recent study by Mora et al. (24) in which it was reported that DC from PP induce high levels of gut homing molecules on CD8+ T cells enabling them to migrate to the small intestine. The novelty of this report is it is the first to describe the ability of DC from PP to prime and induce a gut-homing ability on CD8+ T cells in an infectious disease model. Moreover, our observations go further in demonstrating the importance of IFN-γ in this process.
Our data presented in this study shed a new light on the induction and function of IEL in general and E. cuniculi infection in particular. Based on these findings, we hypothesize the following: oral E. cuniculi infection leads to an activation of DC in mucosal sites like PP and MLN. The CD8αβ+ T cell population in the gut is primed by these DC, and these T cells subsequently acquire homing receptors specific for mucosal T cells, enabling them to home to the epithelial mucosa. Cells of the intestinal epithelium are the first line of defense against oral pathogen. Ag-specific IEL due to their cytolytic activity keep the infection in control thus preventing it from disseminating to other tissues. The current studies raise several important questions regarding the priming of the IEL response. What is the mechanism by which IFN-γ induces an Ag-specific IEL response against E. cuniculi infection? Is IFN-γ involved in efficient Ag processing and presentation by DC? Are DC exclusive APCs able to present Ag to IEL? On-going experiments in our laboratory should help answer these questions and provide information that will be valuable for targeting these cells for developing therapeutic agents against oral pathogens.
Acknowledgments
We thank Joseph Chaiban for help with the phenotypic study.
Footnotes
This work was supported by National Institutes of Health Grants A143693 (to I.A.K.) and A131788 (to L.M.W.).
Abbreviations used in this paper: IEL, intraepithelial lymphocyte; DC, dendritic cell; WT, wild type; pf, perforin; PP, Peyer’s patch; CMA, concanamycin A; FasL, Fas ligand; CD95L, CD95 ligand; MLN, mesenteric lymph node.
Disclosures
The authors have no financial conflict of interest.
References
- 1.Moretto M, Casciotti L, Durell B, Khan IA. Lack of CD4+ T cells does not affect induction of CD8+ T-cell immunity against Encephalitozoon cuniculi infection. Infect Immun. 2000;68:6223–6232. doi: 10.1128/iai.68.11.6223-6232.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Scott P. Development and regulation of cell-mediated immunity in experimental leishmaniasis. Immunol Res. 2003;27:489–498. doi: 10.1385/IR:27:2-3:489. [DOI] [PubMed] [Google Scholar]
- 3.Combe CL, Curiel TJ, Moretto MM, Khan IA. NK cells help to induce CD8+-T-cell immunity against Toxoplasma gondii in the absence of CD4+ T cells. Infect Immun. 2005;73:4913–4921. doi: 10.1128/IAI.73.8.4913-4921.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pamer EG. Immune responses to Listeria monocytogenes. Nat Rev Immunol. 2004;4:812–823. doi: 10.1038/nri1461. [DOI] [PubMed] [Google Scholar]
- 5.Moretto M, Weiss LM, Khan IA. Induction of a rapid and strong antigen-specific intraepithelial lymphocyte response during oral Encephalitozoon cuniculi infection. J Immunol. 2004;172:4402–4409. doi: 10.4049/jimmunol.172.7.4402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hayday A, Theodoridis E, Ramsburg E, Shires J. Intraepithelial lymphocytes: exploring the Third Way in immunology. Nat Immunol. 2001;2:997–1003. doi: 10.1038/ni1101-997. [DOI] [PubMed] [Google Scholar]
- 7.Mattapallil JJ, Reay E, Dandekar S. An early expansion of CD8αβ T cells, but depletion of resident CD8αα T cells, occurs in the intestinal epithelium during primary simian immunodeficiency virus infection. AIDS. 2000;14:637–646. doi: 10.1097/00002030-200004140-00002. [DOI] [PubMed] [Google Scholar]
- 8.Adjei AA, Jones JT, Enriquez FJ. Differential intra-epithelial lymphocyte phenotypes following Cryptosporidium parvum challenge in susceptible and resistant athymic strains of mice. Parasitol Int. 2000;49:119–129. doi: 10.1016/s1383-5769(00)00030-1. [DOI] [PubMed] [Google Scholar]
- 9.Lepage AC, Buzoni-Gatel D, Bout DT, Kasper LH. Gut-derived intraepithelial lymphocytes induce long term immunity against Toxoplasma gondii. J Immunol. 1998;161:4902–4908. [PubMed] [Google Scholar]
- 10.Müller S, Bühler-Jungo M, Mueller C. Intestinal intraepithelial lymphocytes exert potent protective cytotoxic activity during an acute virus infection. J Immunol. 2000;164:1986–1994. doi: 10.4049/jimmunol.164.4.1986. [DOI] [PubMed] [Google Scholar]
- 11.Kokoskin E, Gyorkos TW, Camus A, Cedilotte L, Purtill T, Ward B. Modified technique for efficient detection of microsporidia. J Clin Microbiol. 1994;32:1074–1075. doi: 10.1128/jcm.32.4.1074-1075.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yanai F, Ishii E, Kojima K, Hasegawa A, Azuma T, Hirose S, Suga N, Mitsudome A, Zaitsu M, Ishida Y, et al. Essential roles of perforin in antigen-specific cytotoxicity mediated by human CD4+ T lymphocytes: analysis using the combination of hereditary perforin-deficient effector cells and Fas-deficient target cells. J Immunol. 2003;170:2205–2213. doi: 10.4049/jimmunol.170.4.2205. [DOI] [PubMed] [Google Scholar]
- 13.Iwasaki A, Kelsall BL. Freshly isolated Peyer’s patch, but not spleen, dendritic cells produce interleukin 10 and induce the differentiation of T helper type 2 cells. J Exp Med. 1999;190:229–239. doi: 10.1084/jem.190.2.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ely KH, Kasper LH, Khan IA. Augmentation of the CD8+ T cell response by IFN-γ in IL-12-deficient mice during Toxoplasma gondii infection. J Immunol. 1999;162:5449–5454. [PubMed] [Google Scholar]
- 15.Hochrein H, Shortman K, Vremec D, Scott B, Hertzog P, O’Keeffe M. Differential production of IL-12, IFN-α, and IFN-γ by mouse dendritic cell subsets. J Immunol. 2001;166:5448–5455. doi: 10.4049/jimmunol.166.9.5448. [DOI] [PubMed] [Google Scholar]
- 16.Stober D, Schirmbeck R, Reimann J. IL-12/IL-18-dependent IFN-γ release by murine dendritic cells. J Immunol. 2001;167:957–965. doi: 10.4049/jimmunol.167.2.957. [DOI] [PubMed] [Google Scholar]
- 17.Ohteki T, Fukao T, Suzue K, Maki C, Ito M, Nakamura M, Koyasu S. Interleukin 12-dependent interferon γ production by CD8α+ lymphoid dendritic cells. J Exp Med. 1999;189:1981–1986. doi: 10.1084/jem.189.12.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fukao T, Matsuda S, Koyasu S. Synergistic effects of IL-4 and IL-18 on IL-12-dependent IFN-γ production by dendritic cells. J Immunol. 2000;164:64–71. doi: 10.4049/jimmunol.164.1.64. [DOI] [PubMed] [Google Scholar]
- 19.Fukao T, Frucht DM, Yap G, Gadina M, O’Shea JJ, Koyasu S. Inducible expression of Stat4 in dendritic cells and macrophages and its critical role in innate and adaptive immune responses. J Immunol. 2001;166:4446–4455. doi: 10.4049/jimmunol.166.7.4446. [DOI] [PubMed] [Google Scholar]
- 20.Iwasaki A, Kelsall BL. Unique functions of CD11b+, CD8α+, and double-negative Peyer’s patch dendritic cells. J Immunol. 2001;166:4884–4890. doi: 10.4049/jimmunol.166.8.4884. [DOI] [PubMed] [Google Scholar]
- 21.Kunkel D, Kirchhoff D, Nishikawa S, Radbruch A, Scheffold A. Visualization of peptide presentation following oral application of antigen in normal and Peyer’s patches-deficient mice. Eur J Immunol. 2003;33:1292–1301. doi: 10.1002/eji.200323383. [DOI] [PubMed] [Google Scholar]
- 22.Fleeton M, Contractor N, Leon F, He J, Wetzel D, Dermody T, Iwasaki A, Kelsall B. Involvement of dendritic cell subsets in the induction of oral tolerance and immunity. Ann NY Acad Sci. 2004;1029:60–65. doi: 10.1196/annals.1309.008. [DOI] [PubMed] [Google Scholar]
- 23.Trinchieri G. Interleukin-12: a proinflammatory cytokine with immunoregulatory functions that bridge innate resistance and antigen-specific adaptive immunity. Annu Rev Immunol. 1995;13:251–276. doi: 10.1146/annurev.iy.13.040195.001343. [DOI] [PubMed] [Google Scholar]
- 24.Mora JR, Bono MR, Manjunath N, Weninger W, Cavanagh LL, Rosemblatt M, Von Andrian UH. Selective imprinting of gut-homing T cells by Peyer’s patch dendritic cells. Nature. 2003;424:88–93. doi: 10.1038/nature01726. [DOI] [PubMed] [Google Scholar]
- 25.Ericsson A, Svensson M, Arya A, Agace WW. CCL25/CCR9 promotes the induction and function of CD103 on intestinal intraepithelial lymphocytes. Eur J Immunol. 2004;34:2720–2729. doi: 10.1002/eji.200425125. [DOI] [PubMed] [Google Scholar]
- 26.Mattapallil JJ, Smit-McBride Z, McChesney M, Dandekar S. Intestinal intraepithelial lymphocytes are primed for γ interferon and MIP-1β expression and display antiviral cytotoxic activity despite severe CD4+ T-cell depletion in primary simian immunodeficiency virus infection. J Virol. 1998;72:6421–6429. doi: 10.1128/jvi.72.8.6421-6429.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Guk SM, Yong TS, Chai JY. Role of murine intestinal intra-epithelial lymphocytes and lamina propria lymphocytes against primary and challenge infections with Cryptosporidium parvum. J Parasitol. 2003;89:270–275. doi: 10.1645/0022-3395(2003)089[0270:ROMIIL]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 28.O’Connell J, O’Sullivan GC, Collins JK, Shanahan F. The Fas counterattack: Fas-mediated T cell killing by colon cancer cells expressing Fas ligand. J Exp Med. 1996;184:1075–1082. doi: 10.1084/jem.184.3.1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Smyth MJ, Kelly JM, Sutton VR, Davis JE, Browne KA, Sayers TJ, Trapani JA. Unlocking the secrets of cytotoxic granule proteins. J Leukocyte Biol. 2001;70:18–29. [PubMed] [Google Scholar]
- 30.Ashton-Rickardt PG. The granule pathway of programmed cell death. Crit Rev Immunol. 2005;25:161–182. doi: 10.1615/critrevimmunol.v25.i3.10. [DOI] [PubMed] [Google Scholar]
- 31.Fan ZS, Zhang QX. Molecular mechanisms of lymphocyte-mediated cytotoxicity. Cell Mol Immunol. 2005;2:259–264. [PubMed] [Google Scholar]
- 32.Sydora BC, Mixter PF, Holcombe HR, Eghtesady P, Williams K, Amaral MC, Nel A, Kronenberg M. Intestinal intraepithelial lymphocytes are activated and cytolytic but do not proliferate as well as other T cells in response to mitogenic signals. J Immunol. 1993;150:2179–2191. [PubMed] [Google Scholar]
- 33.Mosley RL, Whetsell M, Klein JR. Proliferative properties of murine intestinal intraepithelial lymphocytes (IEL): IEL expressing TCR αβ or TCR τδ are largely unresponsive to proliferative signals mediated via conventional stimulation of the CD3-TCR complex. Int Immunol. 1991;3:563–569. doi: 10.1093/intimm/3.6.563. [DOI] [PubMed] [Google Scholar]
- 34.Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. doi: 10.1146/annurev.immunol.18.1.767. [DOI] [PubMed] [Google Scholar]
- 35.Mellman I, Steinman RM. Dendritic cells: specialized and regulated antigen processing machines. Cell. 2001;106:255–258. doi: 10.1016/s0092-8674(01)00449-4. [DOI] [PubMed] [Google Scholar]
- 36.Moser M. Regulation of Th1/Th2 development by antigen-presenting cells in vivo. Immunobiology. 2001;204:551–557. doi: 10.1078/0171-2985-00092. [DOI] [PubMed] [Google Scholar]
- 37.Kelsall BL, Strober W. Distinct populations of dendritic cells are present in the subepithelial dome and T cell regions of the murine Peyer’s patch. J Exp Med. 1996;183:237–247. doi: 10.1084/jem.183.1.237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Johansson C, Kelsall BL. Phenotype and function of intestinal dendritic cells. Semin Immunol. 2005;17:284–294. doi: 10.1016/j.smim.2005.05.010. [DOI] [PubMed] [Google Scholar]
- 39.Iwasaki A, Kelsall BL. Localization of distinct Peyer’s patch dendritic cell subsets and their recruitment by chemokines macrophage inflammatory protein (MIP)-3α, MIP-3β, and secondary lymphoid organ chemokine. J Exp Med. 2000;191:1381–1394. doi: 10.1084/jem.191.8.1381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hopkins SA, Niedergang F, Corthesy-Theulaz IE, Kraehenbuhl JP. A recombinant Salmonella typhimurium vaccine strain is taken up and survives within murine Peyer’s patch dendritic cells. Cell Microbiol. 2000;2:59–68. doi: 10.1046/j.1462-5822.2000.00035.x. [DOI] [PubMed] [Google Scholar]
- 41.Pron B, Boumaila C, Jaubert F, Berche P, Milon G, Geissmann F, Gaillard JL. Dendritic cells are early cellular targets of Listeria monocytogenes after intestinal delivery and are involved in bacterial spread in the host. Cell Microbiol. 2001;3:331–340. doi: 10.1046/j.1462-5822.2001.00120.x. [DOI] [PubMed] [Google Scholar]
- 42.Bharhani MS, Grewal JS, Pilgrim MJ, Enocksen C, Peppler R, London L, London SD. Reovirus serotype 1/strain Lang-stimulated activation of antigen-specific T lymphocytes in Peyer’s patches and distal gut-mucosal sites: activation status and cytotoxic mechanisms. J Immunol. 2005;174:3580–3589. doi: 10.4049/jimmunol.174.6.3580. [DOI] [PubMed] [Google Scholar]
- 43.Arstila T, Arstila TP, Calbo S, Selz F, Malassis-Seris M, Vassalli P, Kourilsky P, Guy-Grand D. Identical T cell clones are located within the mouse gut epithelium and lamina propia and circulate in the thoracic duct lymph. J Exp Med. 2000;191:823–834. doi: 10.1084/jem.191.5.823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cuff CF, Cebra CK, Rubin DH, Cebra JJ. Developmental relationship between cytotoxic α/β T cell receptor-positive intraepithelial lymphocytes and Peyer’s patch lymphocytes. Eur J Immunol. 1993;23:1333–1339. doi: 10.1002/eji.1830230622. [DOI] [PubMed] [Google Scholar]
- 45.Nanno M, Matsumoto S, Koike R, Miyasaka M, Kawaguchi M, Masuda T, Miyawaki S, Cai Z, Shimamura T, Fujiura Y, et al. Development of intestinal intraepithelial T lymphocytes is independent of Peyer’s patches and lymph nodes in aly mutant mice. J Immunol. 1994;153:2014–2020. [PubMed] [Google Scholar]
- 46.Guy-Grand D, Azogui O, Celli S, Darche S, Nussenzweig MC, Kourilsky P, Vassalli P. Extrathymic T cell lymphopoiesis: ontogeny and contribution to gut intraepithelial lymphocytes in athymic and euthymic mice. J Exp Med. 2003;197:333–341. doi: 10.1084/jem.20021639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Suzuki K, Oida T, Hamada H, Hitotsumatsu O, Watanabe M, Hibi T, Yamamoto H, Kubota E, Kaminogawa S, Ishikawa H. Gut crypto-patches: direct evidence of extrathymic anatomical sites for intestinal T lymphopoiesis. Immunity. 2000;13:691–702. doi: 10.1016/s1074-7613(00)00068-6. [DOI] [PubMed] [Google Scholar]
- 48.Munder M, Mallo M, Eichmann K, Modolell M. Direct stimulation of macrophages by IL-12 and IL-18: a bridge built on solid ground. Immunol Lett. 2001;75:159–160. doi: 10.1016/s0165-2478(00)00288-1. [DOI] [PubMed] [Google Scholar]
- 49.Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-γ. Annu Rev Immunol. 1997;15:749–795. doi: 10.1146/annurev.immunol.15.1.749. [DOI] [PubMed] [Google Scholar]
- 50.Moretto M, Durell B, Schwartzman JD, Khan IA. γδ T cell-deficient mice have a down-regulated CD8+ T cell immune response against Encephalitozoon cuniculi infection. J Immunol. 2001;166:7389–7397. doi: 10.4049/jimmunol.166.12.7389. [DOI] [PubMed] [Google Scholar]