

Abstract
Microsporidia are obligate intracellular parasites, which can cause complications in immunocompromised individuals. Very little is known about the host immune response generated against these infectious agents. Encephalitozoon cuniculi is the best studied microsporidian and the protective immune response against this parasite is mediated by cytotoxic CD8+ T cells.
1. Introduction
Microsporidia are a ubiquitous group of eukaryotic, intracellular parasites that infect an extremely wide range of hosts in the animal kingdom. They are unique enough to be placed in a separate phylum, Microspora, and are characterized by the polar filament, which injects the sporoplasm into a host cell [1]. The species of microsporidia that infect mammals are unicellular, Gram-positive organisms with mature spores 0.5–2 × 1–4 μm in diameter [2].
Microsporidia are commonly found in laboratory animals such as mice, rabbits and hamsters. Amongst over 140 genera in the phylum Microspora, several different genera have been demonstrated in human disease; in particular, Encephalitozoan spp. were found in many mammals including man [3].
Encephalitozoon cuniculi, which was previously observed in laboratory animals, is considered to be a zoonotic infection [4]. Complications due to E. cuniculi infection have been reported in immunocompromised patients [5]. AIDS patients with peritonitis and hepatitis induced by E. cuniculi infection have been documented. HIV-infected patients with E. cuniculi infection have a wide range of organ involvement including renal failure, pneumonitis, sinusitis and granulomatous liver necrosis [6]. In a recent report, autopsy findings in a patient with AIDS showed disseminated E. cuniculi infection in the brain [7]. In a separate study, a female AIDS patient died from necrotizing microsporidial infection in adrenal glands and kidneys [8]. In this case IFA staining and molecular analysis identified the microsporidian as E. cuniculi. These investigators suggest inclusion of E. cuniculi in the differential diagnosis of disseminated opportunistic infection in AIDS patients. Based on these observations, the National Institute of Allergy and Infectious Diseases has recently classified E. cuniculi as an emerging infectious agent [9].
2. Host immune response to E. cuniculi infection
Little is known regarding host immunity to microsporidia, and in particular E. cuniculi. E. cuniculi was the first mammalian microsporidian successfully grown in vitro [10]. It infects epithelial and endothelial cells, fibroblasts, and macrophages in a variety of mammals, including rabbits, rodents, carnivores, monkeys and humans [4, 11, 12]. In an experimental model, normal mice infected with E. cuniculi usually express few clinical signs of disease [13]. Sensitive and resistant strains of mice exist, as defined by the percentage of macrophages parasitized following intraperitoneal infection, suggesting a genetic basis for innate resistance [13].
Following infection, many mammals have a chronic infection with E. cuniculi, as evidenced by a persistent high antibody titer and ongoing inflammatory process (e.g., persistent encephalitis and kidney lesions in rabbits [14], and congenital disease in carnivores such as foxes [15]. In murine E. cuniculi infection, ascitis develops and then clears in immunocompetent mice. If such mice are then immunosuppressed by corticosteroids they will redevelop ascitis, consistent with a latent persistence of microsporidia in these animals [16]. Other immunodeficient hosts, such as athymic or SCID mice, develop lethal disease after experimental infection [17, 18], usually manifested by ascitis with dissemination of spores. SCID mice are deficient in T and B cells but possess intact natural killer (NK) cell function [19]. Although enhanced NK cell activity in E. cuniculi-infected mice has been reported it does not seem to offer significant in vivo protection [20]. Infected SCID mice reveal numerous microsporidia in visceral and parietal peritoneum as well as in the liver and spleen. Adoptive transfer of sensitized syngeneic T-enriched spleen cells protected athymic or SCID mice against E. cuniculi challenge [21, 22]. Transfer of naive lymphocytes or hyperimmune serum failed to protect or prolong the survival of these mice. Studies by Didier have shown that cytokines released by sensitized T cells activate macrophages to kill E. cuniculi in vitro [23]. These findings suggest that a protective immune response to E. cuniculi is likely dependent on cytokine-producing immune T cells.
2.1. Role of cytokines
Th1 cytokines like IFN-γ and IL-12 are important for protective immunity against a number of intracellular viral, bacterial and parasitic infections [24–26]. Studies with Encephalitozoon intestinalis, a parasite closely related to E. cuniculi, have reported that mice lacking the IFN-γ gene are unable to clear the infection [27]. Based on earlier in vitro observations, it was suggested that IFN-γ also plays an important role in protective immunity against E. cuniculi infection [23, 28] However, the importance of IFN-γ in natural E. cuniculi infection in vivo was not confirmed until recently [29]. In these studies treatment of E. cuniculi-infected mice with neutralizing antibody against IFN-γ or IL-12 resulted in increased mortality for these animals. The use of gene knockout mice further validated the importance of Th1 cytokines in the immune response to E. cuniculi. Both p40−/− mice (which are unable to produce IL-12) and IFN-γ−/− animals succumbed to infection upon E. cuniculi challenge.
Minimal Th2 cytokine production has been observed during E. cuniculi infection [29]. The mRNA for IL-4, a prominent Th2 cytokine, was undetectable in the splenocytes of the infected animals. Similarly, no circulating IL-4 was detected in the sera of infected mice. Nevertheless, an increase in the mRNA for IL-10, another Th2 cytokine, was seen in the splenocytes of such infected animals. As IL-10 has been reported to be involved in the regulation of Th1 immune response in other infectious disease models [30, 31], it is possible that it plays a similar role in E. cuniculi infection.
Cytokine-activated murine peritoneal macrophages can inhibit the replication of E. cuniculi in vitro [32]. This was reported by the studies which demonstrated that inhibition of nitric oxide synthesis prevented parasite growth. However, mice deficient in inducible nitric oxide synthase (NOS2−/−) showed no mortality during E. cuniculi infection [29]. Thus, nitric-oxide-induced killing of E. cuniculi by macrophages or other cells does not appear to be a critical mechanism for control of this infection.
2.2. T-cell subtypes induced in E. cuniculi-infected mice
As stated previously, immune T cells are critical for protection against E. cuniculi infection in the normal host. The role of individual T-cell subtypes during E. cuniculi infection has been recently reported [33]. Phenotypic analysis of the spleen cells from infected animals revealed an increase in the CD8+ T-cell population starting at day 10 postinfection. This rise in the CD8+ T-cell population continued until day 17 p.i. when a > 3-fold increase in the cell type over uninfected controls was observed. Subsequent analysis for activation markers suggested that CD8+ T cells are activated as early as day 3 post-E. cuniculi infection. No significant increase in CD4+ T cells during the course of infection was observed.
To determine whether CD8+ T cells are critical for host survival during E. cuniculi infection gene knockout animals were utilized. All mice deficient in the CD8+ gene succumbed to parasite challenge, showing signs of severe sickness (lethargy, development of ascitis) just before the time of death. Histopathological analysis of the tissues from these animals demonstrated disseminated infection in the spleen and liver. In contrast, E. cuniculi infection did not result in any mortality in CD4−/− mice. Similarly to wild-type controls none of these animals died or demonstrated signs of clinical illness in response to E. cuniculi challenge.
3. Cytotoxic T-cell response to E. cuniculi infection
CD8+ T cells play an important role in the number of intracellular infections [34–36]. The protective effect of CD8+ T cells is mediated by their ability to produce cytokines [37]. Alternatively, CD8+ T cells can reduce the parasite load by killing the infected targets in the host tissue [38]. The major killing mechanism exhibited by CD8+ T cells during E. cuniculi infection is via the perforin pathway [39]. Mice lacking the perforin gene, similar to CD8−/− animals, succumb to E. cuniculi infection. These observations indicate the importance of the cytotoxic T-cell response in E. cuniculi-infected animals. The kinetics of induction of the CD8+ CTL response in E cuniculi-infected mice has been studied [33]. The cytotoxic effect of splenocytes from infected animals was obvious at day 17 p.i. as measured by an in vitro cytotoxicity assay. At this time point effector to target ratio of 20:1 caused 50–60% lysis of E. cuniculi-infected macrophages in vitro. In vitro depletion of CD8+ T cells in the effector cell population resulted in complete loss of cytotoxic activity [33]. These observations suggest that CD8+ T cells mediate the cytolytic effect within the effector immune population. This has been further confirmed by our recent findings in which minimal cytotoxic activity in the splenocytes of CD8−/− mice was detected (Moretto et al., unpublished observations).
3.1. Regulation of CD8+ T-cell immunity in E. cuniculi-infected animals
3.1.1. Role of CD4+ T cells
In a majority of cases, CD8+ T cells are primed via IL-2-producing CD4+ T cells [40]. However, in certain viral infections a normal in vivo CD8+ T-cell response, in the absence of CD4+ T cells, can be induced [41]. As mentioned earlier [33] lack of CD4+ T cells did not affect the outcome of E. cuniculi infection in knockout animals. In-depth analysis of the CD8+ T-cell response to E. cuniculi infection in CD4+ T-cell-deficient animals was performed. Interestingly, a normal antigen-specific CD8+ T-cell response to E. cuniculi infection was observed in CD4−/− mice. The lack of CD4+ T cells did not alter the magnitude of the antigen-specific cytotoxic response and cytokine pattern of CD8+ T cells during E. cuniculi infection. E. cuniculi infection thus offers an example, amongst intracellular parasitic infections, of how CD8+ T cells can be induced in the absence of CD4+ T cells.
3.1.2. Role of γδ T cells
The importance of γδ T cells in response to infectious diseases has been increasingly evident [42] with recent reports indicating that these T cells may be involved in establishing primary immune responses [43]. Studies with Listeria monocytogenes, an intracellular bacteria have demonstrated that an early rise in IFN-γ-producing γδ T cells is followed by an increase in IFN-γ-secreting CD4+ and CD8+ T cells [44]. A many-fold increase in the γδ T-cell population was observed within several days of E. cuniculi infection (Moretto and Khan, unpublished observations). Mice deficient in γδ T cells exhibited susceptibility to E. cuniculi when challenged with very high parasite doses. However, unlike CD8−/− or αβ T-cell-deficient mice, δ−/− animals were able to survive a low-dose infection (Moretto and Khan, unpublished observations). The susceptibility of δ−/− mice can be attributed to downregulation of CD8+ T-cell immunity in these animals, as we observed a significant decrease in the antigen-specific CD8+ T-cell immune response to E. cuniculi infection in these mice (Moretto and Khan, unpublished observations). Based on these observations it appears that γδ T cells play a prominent role in priming of the CD8+ T cell during E. cuniculi infection. The induction of CD8+ T-cell immunity by γδ T cells may be due to their ability to release early IFN-γ necessary from priming the CD8+ T-cell response as reported in other models [43].
4. Humoral immunity in microsporidial infections
Adoptive transfer of immune B lymphocytes into athymic BALB/c (nu/nu) or SCID mice does not protect these animals from death following E. cuniculi infection [45]. Passive transfer of hyperimmune serum into athymic BALB/c (nu/nu) mice does not prevent lethal infection [17]. Nonetheless, during E. cuniculi infection there is a strong antibody response to different antigens of this organism and many of these antibodies are cross-reactive with other microsporidia [17, 46]. In humans, microsporidia antibodies have been found in from 5 to 50% of survey populations, with the highest incidence in those patients from the tropics or who had visited the tropics [47, 48]. In random blood donors the seroprevalence is 5% [49]. Data exist showing that maternal antibodies may protect new-born rabbits from infection with E. cuniculi during the first 2 weeks of life [50]. In addition, in vitro infectivity of microsporidia is reduced by treatment with immune serum and complement [21], monoclonal antibody (mAb 3B6) to spore coat [45] or monoclonal and polyclonal antibodies to polar tube protein (Weiss, unpublished data). Thus, it is likely that antibodies play a role in limiting infection in the host, although they are clearly not sufficient to prevent mortality or to cure infection.
5. Conclusion
Cell-mediated immunity is critical for the survival of E. cuniculi-infected hosts. Over the last several years significant progress has been made in understanding the cellular immune responses to E. cuniculi infection. A working hypothesis of the immune response to E. cuniculi is illustrated in figure 1. Similar to other intracellular infections, E. cuniculi may result in a strong burst of IL-12 production by host macrophages or dendritic cells [51]. Early IL-12 release results in the polarization towards Th1 cytokines manifested by high levels of IFN-γ in the circulation and tissues. γδ T cells, which are increased at early stages of infection, are probably important sources of IFN-γ production. Th2 cytokines such as IL-4, are not detectable throughout the course of infection. The increase in IFN-γ production is known to cause upregulation of class I molecules on the infected cells [52]. This most likely leads to antigen-specific CD8+ T-cell proliferation. CD8+ CTLs are primary effector cells responsible for keeping parasite replication under control [53]. The role of CD4+ T cells in protective immune response to E. cuniculi is somewhat ambiguous. CD4+ T-cell-deficient mice show normal protection against parasite challenge [33] and the CD8+ T-cell immune response in these knockout mice is not compromised (Moretto et al., unpublished observations). Based on these findings, it appears that the CD8+ CTL response during E. cuniculi infection can be launched independently of CD4+ T cells. The presence of γδ T cells appears to be crucial for the induction of optimal CD8+ T-cell immunity.
Figure 1.
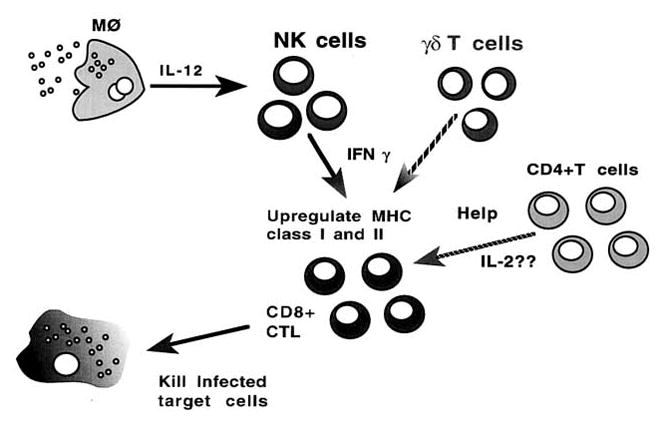
Overall picture of immune response to E. cuniculi infection. CD8+ T cells play a critical role in host defense against E. cuniculi infection. The primary CD8+ CTL response can be induced in the absence of IL-2-secreting CD4+ T cells. IFN-γ-producing NK cells or γδ T cells may play a role in the induction of early CD8+ T-cell immunity.
Based on the information available, the following hypothesis can be put forward: E. cuniculi infection induces a strong CD8+ CTL response, which restricts parasite growth by lysing the infected cells via a perforin-dependent mechanism. The induction of CD8+ CTLs is at least partially regulated by γδ T cells. This regulation may be dependent on the ability of γδ T cells to produce cytokines like IFN-γ. The role of IFN-γ in inducing and maintaining CD8+ T-cell immunity against infectious agents has been clearly demonstrated [54]. There are still, however, important questions which remain to be addressed: is IFN-γ critical for the induction of the CD8+ T response during E. cuniculi infection? What is the mechanism of γδ T-cell induction of the CD8+ CTL response? Ongoing research in our laboratory should provide answers to these critical questions and refine our understanding of the host immune response to these emerging pathogens.
Acknowledgments
This work was supported by National Institute of Health grants AI43963 (IAK) and A131788 (LMW).
References
- 1.Vavra J, Larsson JIR. Structure of the Microsporidia. In: Wittner M, Weiss LM, editors. The Microsporidia and Microsporidiosis. ASM; Washington, D.C: 1999. pp. 7–84. [Google Scholar]
- 2.Weber R, Schwartz DA, Deplazes P. Laboratory diagnosis of microsporidiosis. In: Wittner M, Weiss LM, editors. The Microsporidia and Microsporidiosis. ASM; Washington, D.C: 1999. pp. 315–362. [Google Scholar]
- 3.Sprague V, Becnel JJ. Appendix; checklist of available generic names for microsporidia with type species and hosts. In: Wittner M, Weiss LM, editors. The Microsporidia and Microsporidiosis. ASM; Washington, D.C: 1999. pp. 517–530. [Google Scholar]
- 4.Deplazes P, Mathis A, Baumgartner R, Tanner I, Weber R. Immunologic and molecular characteristics of Encephalitozoon-like microsporidia isolated from humans and rabbits indicate that Encephalitozoon cuniculi is a zoonotic parasite. Clin Infect Dis. 1996;22:557–559. doi: 10.1093/clinids/22.3.557. [DOI] [PubMed] [Google Scholar]
- 5.Shadduck JA, Greeley E. Microsporidia and human infections. Clin Microbiol Rev. 1989;2:158–165. doi: 10.1128/cmr.2.2.158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Didier ES, Didier PJ, Snowden KF, Shadduck JA. Microsporidiosis in Mammals. Microbes Infect. 2000;2:709–720. doi: 10.1016/s1286-4579(00)00354-3. [DOI] [PubMed] [Google Scholar]
- 7.Weber R, Deplazes P, Flepp M, Mathis A, Baumann R, Sauer B, Kuster H, Luthy R. Cerebral microsporidiosis due to Encephalitozoon cuniculi in a patient with human immunodeficiency virus infection. N Engl J Med. 1997;336:474–478. doi: 10.1056/NEJM199702133360704. [DOI] [PubMed] [Google Scholar]
- 8.Mertens RB, Didier ES, Fishbein MC, Bertucci DC, Rogers LB, Orenstein JM. Encephalitozoon cuniculi microsporidiosis: infection of the brain, heart, kidneys, trachea, adrenal glands, and urinary bladder in a patient with AIDS. Mod Pathol. 1997;10:68–77. [PubMed] [Google Scholar]
- 9.Doepel L. Emerging infectious diseases research: NIAID meets the challenge. Dateline: NIAID. 1997;9:1–2. [Google Scholar]
- 10.Shadduck JA. Nosema cuiculi: in vitro isolation. Science. 1969;166:516–517. doi: 10.1126/science.166.3904.516. [DOI] [PubMed] [Google Scholar]
- 11.De Groote MA, Visvesvara G, Wilson ML, Pieniazek NJ, Slemenda SB, daSilva AJ, Leitch GJ, Bryan RT, Reves R. Polymerase chain reaction and culture confirmation of disseminated Encephalitozoon cuniculi in a patient with AIDS: successful therapy with albendazole. J Infect Dis. 1995;171:1375–1378. doi: 10.1093/infdis/171.5.1375. [DOI] [PubMed] [Google Scholar]
- 12.Hollister WS, Canning EU, Colbourn NI, Aarons EJ. Encephalitozoon cuniculi isolated from the urine of an AIDS patient, which differs from canine and murine isolates. J Eukaryot Microbiol. 1995;42:367–372. doi: 10.1111/j.1550-7408.1995.tb01595.x. [DOI] [PubMed] [Google Scholar]
- 13.Niederkorn JY, Shadduck JA, Schmidt EC. Susceptibility of selected inbred strains of mice to Encephalitozoon cuniculi. J Infect Dis. 1981;144:249–253. doi: 10.1093/infdis/144.3.249. [DOI] [PubMed] [Google Scholar]
- 14.Shadduck JA, Pakes SP. Encephalitozoonosis (nosematosis) and toxoplasmosis. Am J Pathol. 1971;64:657–671. [PMC free article] [PubMed] [Google Scholar]
- 15.Botha WS, van Dellen AF, Stewart CG. Canine encephalitozoonosis in South Africa. J S Afr Vet Assoc. 1979;50:135–144. [PubMed] [Google Scholar]
- 16.Innes JR. Parasitic infections of the nervous system of animals. Ann NY Acad Sci. 1970;174:1042–1047. doi: 10.1111/j.1749-6632.1970.tb45626.x. [DOI] [PubMed] [Google Scholar]
- 17.Schmidt EC, Shadduck JA. Murine encephalitozoonosis model for studying the host-parasite relationship of a chronic infection. Infect Immun. 1983;40:936–942. doi: 10.1128/iai.40.3.936-942.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Koudela B, Vitovec J, Kucerova Z, Ditrich O, Travnicek J. The severe combined immunodeficient mouse as a model for Encephalitozoon cuniculi microsporidiosis. Folia Parasitol. 1993;40:279–286. [PubMed] [Google Scholar]
- 19.Hunter CA, Subauste CS, Van Cleave VH, Remington JS. Production of gamma interferon by natural killer cells from Toxoplasma gondii-infected SCID mice: regulation by interleukin-10, interleukin-12, and tumor necrosis factor alpha. Infect Immun. 1994;62:2818–2824. doi: 10.1128/iai.62.7.2818-2824.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Niederkorn JY, Brieland JK, Mayhew E. Enhanced natural killer cell activity in experimental murine encephalitozoonosis. Infect Immun. 1983;41:302–307. doi: 10.1128/iai.41.1.302-307.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schmidt EC, Shadduck JA. Mechanisms of resistance to the intracellular protozoan Encephalitozoon cuniculi in mice. J Immunol. 1984;133:2712–2719. [PubMed] [Google Scholar]
- 22.Hermanek J, Koudela B, Kucerova Z, Ditrich O, Travnicek J. Prophylactic and therapeutic immune reconstitution of SCID mice infected with Encephalitozoon cuniculi. Folia Parasitol. 1993;40:287–291. [PubMed] [Google Scholar]
- 23.Didier ES. Reactive nitrogen intermediates implicated in the inhibition of Encephalitozoon cuniculi (phylum microspora) replication in murine peritoneal macrophages. Parasite Immunol. 1995;17:405–412. doi: 10.1111/j.1365-3024.1995.tb00908.x. [DOI] [PubMed] [Google Scholar]
- 24.Suzuki Y, Conley FK, Remington JS. Importance of endogenous IFN-gamma for prevention of toxoplasmic encephalitis in mice. J Immunol. 1989;143:2045–2050. [PubMed] [Google Scholar]
- 25.Von Herrath MG, Coon B, Oldstone MB. Low-affinity cytotoxic T-lymphocytes require IFN-gamma to clear an acute viral infection. Virology. 1997;229:349–359. doi: 10.1006/viro.1997.8442. [DOI] [PubMed] [Google Scholar]
- 26.Yang J, Mitsuyama M. An essential role for endogenous interferon-gamma in the generation of protective T cells against Mycobacterium bovis BCG in mice. Immunology. 1997;91:529–535. doi: 10.1046/j.1365-2567.1997.00288.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Achbarou A, Ombrouck C, Gneragbe T, Charlotte F, Renia L, Desportes-Livage I, Mazier D. Experimental model for human intestinal microsporidiosis in interferon gamma receptor knockout mice infected by Encephalitozoon intestinalis. Parasite Immunol. 1996;18:387–392. doi: 10.1046/j.1365-3024.1996.d01-128.x. [DOI] [PubMed] [Google Scholar]
- 28.Braunfuchsova P, Kopecky J, Ditrich O, Koudela B. Cytokine response to infection with the microsporidian, Encephalitozoon cuniculi. Folia Parasitol. 1999;46:91–95. [PubMed] [Google Scholar]
- 29.Khan IA, Moretto M. Role of gamma interferon in cellular immune response against murine Encephalitozoon cuniculi infection. Infect Immun. 1999;67:1887–1893. doi: 10.1128/iai.67.4.1887-1893.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gazzinelli RT, Hayashi S, Wysocka M, Carrera L, Kuhn R, Muller W, Roberge F, Trinchieri G, Sher A. Role of IL-12 in the initiation of cell mediated immunity by Toxoplasma gondii and its regulation by IL-10 and nitric oxide. J Eukaryot Microbiol. 1994;41:9S. [PubMed] [Google Scholar]
- 31.Trinchieri G. Cytokines acting on or secreted by macrophages during intracellular infection (IL-10, IL-12, IFN-gamma) Curr Opin Immunol. 1997;9:17–23. doi: 10.1016/s0952-7915(97)80154-9. [DOI] [PubMed] [Google Scholar]
- 32.Didier ES, Varner PW, Didier PJ, Aldras AM, Milli-champ NJ, Murphey-Corb M, Bohm R, Shadduck JA. Experimental microsporidiosis in immunocompetent and immunodeficient mice and monkeys. Folia Parasitol. 1994;41:1–11. [PubMed] [Google Scholar]
- 33.Khan IA, Schwartzman JD, Kasper LH, Moretto M. CD8+ CTLs are essential for protective immunity against Encephalitozoon cuniculi infection. J Immunol. 1999;162:6086–6091. [PubMed] [Google Scholar]
- 34.Harty JT, Bevan MJ. CD8+ T cells specific for a single nonamer epitope of Listeria monocytogenes are protective in vivo. J Exp Med. 1992;175:1531–1538. doi: 10.1084/jem.175.6.1531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Khan IA, Kasper LH. IL-15 augments CD8+ T cell-mediated immunity against Toxoplasma gondii infection in mice. J Immunol. 1996;157:2103–2108. [PubMed] [Google Scholar]
- 36.Tripp RA, Hou S, McMickle A, Houston J, Doherty PC. Recruitment and proliferation of CD8+ T cells in respiratory virus infections. J Immunol. 1995;154:6013–6021. [PubMed] [Google Scholar]
- 37.Ramshaw IA, Ramsay AJ, Karupiah G, Rolph MS, Mahalingam S, Ruby JC. Cytokines and immunity to viral infections. Immunol Rev. 1997;159:119–135. doi: 10.1111/j.1600-065x.1997.tb01011.x. [DOI] [PubMed] [Google Scholar]
- 38.Walsh CM, Matloubian M, Liu CC, Ueda R, Kurahara CG, Christensen JL, Huang MT, Young JD, Ahmed R, Clark WR. Immune function in mice lacking the perforin gene. Proc Natl Acad Sci USA. 1994;91:10854–10858. doi: 10.1073/pnas.91.23.10854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Denkers EY, Yap G, Scharton-Kersten T, Charest H, Butcher BA, Caspar P, Heiny S, Sher A. Perforin-mediated cytolysis plays a limited role in host resistance to Toxoplasma gondii. J Immunol. 1997;159:1903–1908. [PubMed] [Google Scholar]
- 40.Smith KA. Interleukin-2: inception, impact, and implications. Science. 1988;240:1169–1176. doi: 10.1126/science.3131876. [DOI] [PubMed] [Google Scholar]
- 41.Binder D, Kundig TM. Antiviral protection by CD8+ versus CD4+ T cells. CD8+ T cells correlating with cytotoxic activity in vitro are more efficient in antivaccinia virus protection than CD4-dependent IL. J Immunol. 1991;146:4301–4307. [PubMed] [Google Scholar]
- 42.Barnes PF, Grisso CL, Abrams JS, Band H, Rea TH, Modlin RL. Gamma delta T lymphocytes in human tuberculosis. J Infect Dis. 1992;165:506–512. doi: 10.1093/infdis/165.3.506. [DOI] [PubMed] [Google Scholar]
- 43.Ferrick DA, Schrenzel MD, Mulvania T, Hsieh B, Ferlin WG, Lepper H. Differential production of interferon-gamma and interleukin-4 in response to Th1-and Th2-stimulating pathogens by gamma delta T cells in vivo. Nature. 1995;373:255–257. doi: 10.1038/373255a0. [DOI] [PubMed] [Google Scholar]
- 44.Skeen MJ, Ziegler HK. Intercellular interactions and cytokine responsiveness of peritoneal alpha/beta and gamma/delta T cells from Listeria-infected mice: synergistic effects of interleukin 1 and 7 on gamma/delta T cells. J Exp Med. 1993;178:985–996. doi: 10.1084/jem.178.3.985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Enriquez EJ. Microsporidia: immunity and immunodiagnosis, 2nd Workshop Microsporidiosis and Cryptosporidiosis Immunodeficient Patients; Ceske Budejovice, Czeh Republic. 1997. p. 16. [Google Scholar]
- 46.Cox JC, Hamilton RC, Attwood HD. An investigation of the route and progression of Encephalitozoon cuniculi infection in adult rabbits. J Protozool. 1979;26:260–265. doi: 10.1111/j.1550-7408.1979.tb02776.x. [DOI] [PubMed] [Google Scholar]
- 47.Singh M, Kane GJ, Mackinlay L, Quaki I, Yap EH, Ho BC, Ho LC, Lim KC. Detection of antibodies to Nosema cuniculi (Protozoa: Microscoporidia) in human and animal sera by the indirect fluorescent antibody technique. Southeast Asian J Trop Med Public Health. 1982;13:110–113. [PubMed] [Google Scholar]
- 48.Bergquist NR, Stintzing G, Smedman L, Waller T, Andersson T. Diagnosis of encephalitozoonosis in man by serological tests. Br Med J Clin Res. 1984;288:902. doi: 10.1136/bmj.288.6421.902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.van Gool T, Vetter JC, Weinmayr B, Van Dam A, Derouin F, Dankert J. High seroprevalence of Encephalitozoon species in immunocompetent subjects. J Infect Dis. 1997;175:1020–1024. doi: 10.1086/513963. [DOI] [PubMed] [Google Scholar]
- 50.Bywater JE, Kellett BS. Humoral immune response to natural infection with Encephalitozoon cuniculi in rabbits. Lab Anim. 1979;13:293–297. doi: 10.1258/002367779780943305. [DOI] [PubMed] [Google Scholar]
- 51.Sousa CR, Hieny S, Scharton-Kersten T, Jankovic D, Charest H, Germain RN, Sher A. In vivo microbial stimulation induces rapid CD40 ligand-independent production of interleukin 12 by dendritic cells and their redistribution to T cell areas. J Exp Med. 1997;186:1819–1829. doi: 10.1084/jem.186.11.1819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-gamma. Annu Rev Immunol. 1997;15:749–795. doi: 10.1146/annurev.immunol.15.1.749. [DOI] [PubMed] [Google Scholar]
- 53.Pye D, Cox JC. Simple focus assay for Encephalitozoon cuniculi. Lab Anim. 1979;13:193–195. doi: 10.1258/002367779780937807. [DOI] [PubMed] [Google Scholar]
- 54.Ely KH, Kasper LH, Khan IA. Augmentation of the CD8+ T cell response by IFN-gamma in IL-12-deficient mice during Toxoplasma gondii infection. J Immunol. 1999;162:5449–5454. [PubMed] [Google Scholar]