

Abstract
As part of the Trypanosoma cruzi Genome Initiative, we have mapped a large portion of the chromosomal bands XVI (2.3 Mb) and XVII (2.6 Mb) containing the highly repetitive and immunodominant antigenic gene families h49 and jl8. Restriction mapping of the isolated chromosomal bands and hybridization with chromosome specific gene probes showed that genes h49 and jl8 are located in a pair of size-polymorphic homologous chromosomes. To construct the integrated map of the chromosomes harboring the h49 and jl8 loci, we used YAC, cosmid, and λ phage overlapping clones, and long range restriction analysis using a variety of probes (i.e., known gene sequences, ESTs, polymorphic repetitive sequences, anonymous sequences, STSs generated from the YAC ends). The total length covered by the YAC contig was approximately 670 kb, and its map agreed and was complementary to the one obtained by long-range restriction fragment analysis. Average genetic marker spacing in a 105 kb region around h49 and jl8 genes was estimated to be 6.2 kb/marker. We have detected some polymorphism in the H49/JL8 antigens-encoding chromosomes, affecting also the coding regions. The physical map of this region, together with the isolation of specific chromosome markers, will contribute in the global effort to sequence the nuclear genome of this parasite.
Trypanosoma cruzi is a parasitic protozoan that causes Chagas disease, an ailment without effective drug treatment affecting 16–18 million people in the American continent (Brener 1973). Classical genetic studies on this parasite have been hampered by the existence of many parasite strains with distinct biological and immunological characteristics, the lack of sexual stages, and a high variability in both number and size of chromosomes (Brener 1973; Solari 1995; Cano et al. 1995; Henriksson et al. 1995, 1996; Santos et al. 1997). For this reason, one of the main goals of the recently started T. cruzi Genome Initiative is to construct a physical map of the parasite's genome as a resource for DNA sequence analysis (Hanke et al. 1996, 1998; Zingales et al. 1997a; Ferrari et al. 1997; Frohme et al. 1998). A physical map of the T. cruzi genome would also facilitate the identification and characterization of genetic loci. Having large chromosome segments that span these loci cloned in YACs will help to identify genes involved in parasite's survival, pathogenicity, and diagnosis.
Of special interest are the regions harboring genes that encode antigens involved in parasite virulence which might be important for the development of immune therapeutics, drugs, and diagnostic reagents. In T. cruzi and other protozoan parasites that infect humans, the occurrence of proteins containing stretches of tandemly arranged amino acid repeats is a commom finding (Frasch et al. 1991). Natural humoral immune responses to many T. cruzi antigens appear to be largely directed to epitopes encoded by the repeat units (Frasch et al. 1991; Levin et al. 1991; Umezawa et al. 1999). It has been proposed that repetitive domains may function as ligands for host proteins or immunomodulators (Buscaglia et al. 1999). The repetitive antigens could help the parasite to evade immune response, presenting to the host a large number of nonprotective epitopes (Kemp et al. 1987). The present work focuses on the characterization of a large chromosomal region harboring genes for the immunodominant repetitive antigens H49 and JL8 as the main markers (Ibanez et al. 1988; Levin et al. 1989; Lafaille et al. 1989; Hoft et al. 1989; Cotrim et al. 1990, 1995). These antigens are high molecular weight proteins located, respectively, along the attachment region between the flagellum and the cell body (Lafaille et al. 1989; Cotrim et al. 1990, 1995) and in the cytoplasm and the peri-nuclear region (Lafaille et al. 1989; Hoft et al. 1995). They are remarkbly immunogenic in man and have been successfully employed in the serological diagnosis of Chagas disease (Levin et al. 1992; Krieger et al. 1992; Carvalho et al. 1993; Paranhos et al. 1994; Pastini et al. 1994; Pereira et al. 1998; Umezawa et al. 1999). The genes h49 and jl8 were mapped on two neighboring chromosomal bands of 2.3 and 2.6 Mb (bands XVI and XVII, respectively, in the nomenclature of Cano et al. 1995) of clone CL Brener, the reference strain of the T. cruzi Genome Initiative. We present here the long-range restriction analysis and YAC contig assembly of a 670 kb region harboring loci h49 and jl8, which covers 26% of the entire chromosome and includes 21 genetic markers. We also present further evidence supporting the disomic nature of of H49/JL8 antigens-encoding chromosomes.
RESULTS AND DISCUSSION
Identification of Markers for Chromosomal Bands XVI and XVII
The chromatin of T. cruzi is poorly condensed during nuclear division and chromosomes can not be visualized by conventional cytogenetic methods (Solari 1995). By using pulsed field gel electrophoresis (PFGE) it is possible to separate the chromosome-sized DNA molecules of this protozoan. The molecular karyotype of clone CL Brener consists of 20 chromosomal bands (0.45–3.5 Mb 12 megabase bands (3.5–1.0 Mb), and 8 intermediate bands (0.45–1.0 Mb) (Cano et al. 1995). The distribution of the ethidium bromide fluorescence is not the same for all chromosomal bands indicating that most of the bands contain two or more comigrating chromosomes which are not necessarily homologous (Cano et al. 1995; Henriksson et al. 1996; Santos et al. 1997). Indeed, the existence of homologous chromosomes which differ substantially in size has been suggested (Henriksson et al. 1995, 1996; Cano et al. 1995). For this reason, we refer to chromosomal bands as those bands separated by PFGE and visible after staining with ethidium bromide.
Genes encoding the immunodominant antigens H49 and JL8 were mapped on two neighboring chromosomal bands of 2.3 Mb and 2.6 Mb (bands XVI and XVII, respectively). Densitometric analysis of the DNA content of bands XVI and XVII indicates the presence of at least two comigrating chromosomes per band which are not necessarily homologous (Cano et al. 1995). To identify additional genetic markers, we have hybridized chromoblots carrying T. cruzi chromosomal bands separated by PFGE with a panel of T. cruzi cloned sequences (6 repetitive sequences, 42 genes encoding structural proteins and RNAs, 120 ESTs, and 9 STSs). Markers mapped on chromosomal bands XVI and XVII are listed in Table 1. Many genetic markers such as C6, SIRE, E13, minisatellite, B11, SRE, STS–SZ23–14, gp90, gp85, gp82, Tc85, cDNA–Sx23 (gp85-like protein), mucin, and 12 ESTs (TEUF0040, TEUF0078, TEUF0548, TEUF0047, TEUF0052, TEUF0059, TEUF0039, TEUF0051, TEUF131, TEUF133, TEUF0115, TEUF0116) were mapped to chromosomal bands XVI and XVII. This hybridization pattern corroborates the hypothesis that bands XVI and XVII carry size-polymorphic homologous chromosomes (Table 1). The existence of two homologs of a diploid chromosome that differ in size had been reported previously in T. cruzi and Trypanosoma brucei (Turner et al. 1997; Frohme et al. 1998; Hanke et al. 1998; Melville et al. 1998).
Table 1.
Summary of Genetic Markers for the Chromosomal Bands XVI and XVII
| Marker | GenBank accession no. | Homology or prominent features | Chromosomal banda | |
|---|---|---|---|---|
| XVI | XVII | |||
| h49 | L09564 | flagellar repetitive antigen | + | + |
| jl8 | AF47956 | cytoplasmic repetitive antigen | + | + |
| Mucin | AF027869 | sialic acid acceptor | + | + |
| gp82 | L14824 | gp82 surface glycoprotein | + | + |
| gp90 | L11287 | gp90 surface glycoprotein | + | + |
| gp85 | J04667 | gp85 surface glycoprotein | + | + |
| Tc-85 | AF085686 | gp85 surface glycoprotein | + | + |
| TEUF0040 | AA441753 | unknown ORF | + | + |
| TEUF0078 | AA426697 | γ-1 elongation factor | + | + |
| TEUF0548 | — | unknown ORF | + | + |
| cDNASX23 | — | amastigote gp85-like protein | + | + |
| B11 | U15615 | retrotransposon | + | + |
| SIRE | X75033 | interspersed repetitive element | + | + |
| C6 | U16295 | interspersed repetitive element | + | + |
| SRE | M63895 | spacer repetitive element | + | + |
| Satellite | K00393 | 196-bp minisatellite sequence | + | + |
| SZ23 | — | anonymous marker | + | + |
| SZ14 | — | anonymous marker | + | + |
| TEUF0047 | AA441756 | unknown ORF | + | + |
| TEUF0052 | AA676166 | histone H3 | + | + |
| TEUF0069 | — | unknown ORF | + | + |
| TEUF0059 | AA441765 | unknown ORF | + | + |
| TEUF0039 | — | unknown ORF | + | + |
| TEUF0051 | — | unknown ORF | + | + |
| TEUF0133 | AA676182 | proteosome 26S subunit | + | + |
| TEUF0131 | AA426657 | D18 protein | + | + |
| TEUF0115 | AA433298 | unknown ORF | + | + |
| TEUF0116 | AA426656 | unknown ORF | + | + |
| Ts cat. | U01098 | trans-sialidase | + | − |
| SL | K02632 | spliced leader sequence | + | − |
| cDNASX42 | — | amastigote gp85-like protein | + | − |
| TEUF0065 | — | unknown ORF | + | − |
| TEUF0101 | AA399705 | elongation factor II | + | − |
| TEUF0054 | AA441761 | unknown ORF | + | − |
| TEUF0143 | AA433312 | unknown ORF | − | + |
| TEUF0137 | AA433310 | unknown ORF | − | + |
| TEUF0132 | AA433308 | unknown ORF | − | + |
| TEUF0095 | AA441779 | unknown ORF | − | + |
| TEUF0049 | AA426691 | reverse transcriptase | − | + |
| TEUF0206 | AA426673 | unknown ORF | − | + |
| TEUF0068 | AA676169 | unknown ORF | − | + |
| TEUF0537 | AA676206 | unknown ORF | − | + |
| TEUF0056 | AA441763 | tyrosine amino transferase | − | + |
| PABP | AF044733 | poly(A) binding protein | − | + |
(+) presence or (−) absence of the marker on chromosomal band.
However, other probes hybridized only with either band XVI or band XVII. For instance, some markers such as the spliced leader sequence, trans-sialidase catalytic domain, cDNA–Sx42, TEUF0065, TEUF0101, and TEUF0054 hybridized only with band XVI whereas probes PABP, TEUF0143, TEUF0137, TEUF0132, TEUF0095, TEUF0049, TEUF0206, TEUF0068, TEUF0537, and TEUF0056 hybridized only with band XVII (Table 1). These results could be explained by the presence of nonhomologous chromosomes in both bands XVI and XVII. To test this hypothesis, the chromosomal bands XVI and XVII were digested with NotI and hybridized with a T. cruzi telomeric sequence (CCCTAA)3 (Chiurillo et al. 1999). In the case where each chromosomal band would contain two or more comigrating heterologous chromosomes, the telomeric probe should hybridize with more than two NotI fragments. Although it is difficult to line up the signals for each band, the telomeric probe hybridized with at least four NotI restriction fragments per band (not shown). Thus, chromosomal bands XVI and XVII contain comigrating heterologous chromosomes, and the two homologs of a diploid chromosome (H49/JL8 antigens-encoding chromosome) that differ in size.
Probes H49 and JL8 Define a Pair of Homologous Chromosomes that Differ in Size
The results above suggest that h49 and jl8 loci are located in pair of size-polymorphic homologs chromosomes. We then investigated whether the overall chromosome structure of these homologs is conserved. The chromosomal bands XVI and XVII were separated by PFGE (Fig. 1A), excised from the preparative gels, and incubated with the restriction enzymes EcoRI or BamHI. After digestion, the restriction fragments were separated by electrophoresis and hybridized to H49 and JL8 probes (Fig. 1B). The restriction patterns of bands XVI and XVII were identical. Probe H49 reacted with two EcoRI fragments (20.0 kb and 9.7 kb) and three BamHI fragments (23.0, 10.0, and 5.5 kb) in both chromosomal bands. Probe JL8 hybridized with a 30.0 kb EcoRI fragment and a 30.0 kb BamHI fragment in bands XVI and XVII.
Figure 1.
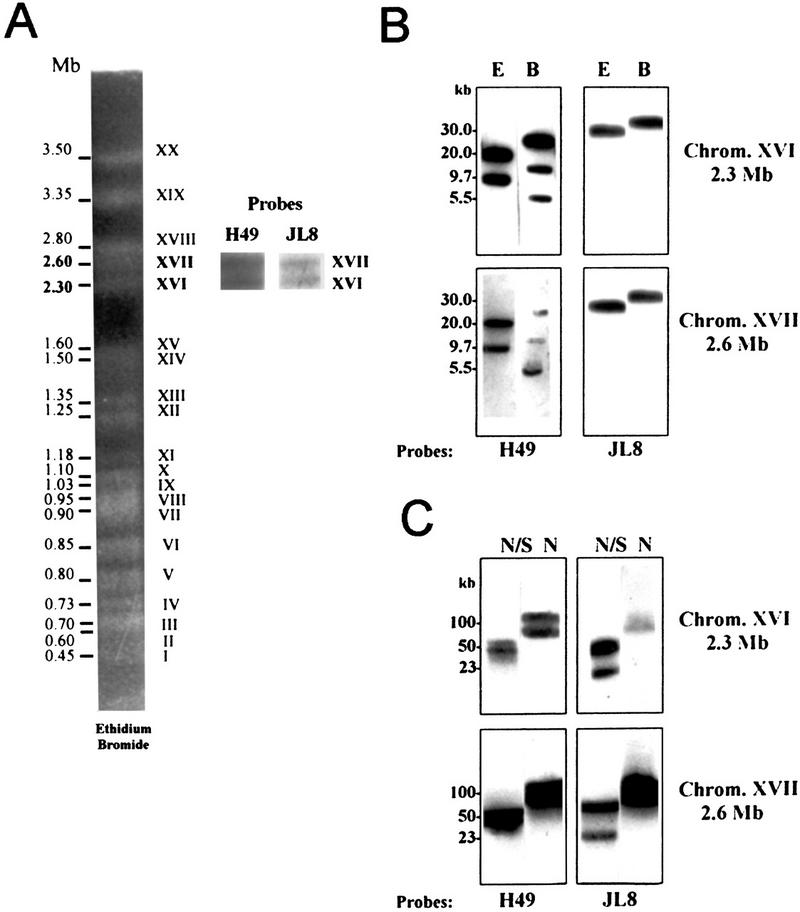
Probes H49 and JL8 define a pair of size-polymorphic homologous chromosomes. (A) Separation of T. cruzi chromosomal bands by PFGE and staining with ethidium bromide. The arabic and roman numerals indicate the sizes and numbers of the chromosomal bands, respectively. The insert shows the hybridization of probes H49 and JL8 with bands XVI and XVII. (B) The chromosomal bands XVI (2.3 Mb) and XVII (2.6 Mb) were isolated from a preparative PFGE gel and digested with EcoRI (E) or BamHI (B) restriction enzymes. The restriction fragments were separated by conventional agarose gel electrophoresis, blotted into a nylon membrane, and hybridized with probes H49 and JL8. (C) The chromosomal bands XVI and XVII were isolated from a preparative PFGE gel and digested with NotI and SfiI (N/S) or NotI (N). Restriction fragments were separated by PFGE (100 V, pulses of 10 sec to 120 sec, 40 hr) and hybridized with probes H49 and JL8.
We then used rare cutting restriction enzymes, such as NotI and SfI, to digest the isolated chromosomal bands (Fig. 1C). Once again, probes H49 and JL8 reacted with the same restriction fragments in both chromosomal bands. Probe H49 hybridized with two NotI restriction fragments of about 100 kb and 90 kb indicating the presence of at least two copies of gene h49. It is noteworthy that probe JL8 hybridized only to the 100 kbNotI restriction fragment, suggesting that gene jl8 is present in only one copy, located within 100 kb of one of the h49 genes. As can be seen in Figure 1, the hybridization intensity with the probes JL8 and H49 seems to be similar for each chromosomal band as confirmed by densitometric analysis (not shown). The restriction maps of the isolated chromosomal bands are co-linear with the genomic restriction map (Fig. 2A) obtained by digestion of total chromosomal DNA. Data indicates that the restriction maps of the regions that flank the h49 and jl8 genes, covering >400 kb, are identical. This information confirms that these genes are located in a pair of size-polymorphic homologous chromosomes.
Figure 2.
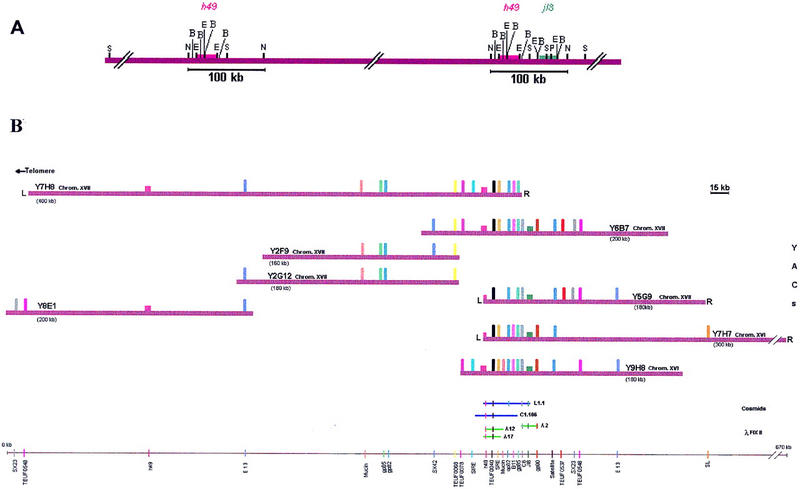
Megarestriction map and YAC contig of the region containing the loci h49 and j18. (A) Megarestriction map of the region spanning the loci h49 and j18. As described in legend of Figure 1, T. cruzi chromosomal DNA and isolated chromosomal bands XVI and XVII were digested with different restriction enzymes, separated by conventional agarose gel electrophoresis or by PFGE, blotted into nylon membrane, and hybridized with probes H49 and JL8. Restriction enzymes: S, SfI; N, NotI; E, EcoRI; B, BamHI; P, PacI. Red and green boxes indicate the location of genes h49 and jl8, respectively. To facilitate comprehension, the multiple HindIII restriction sites within the repetitive region of gene h49, and HaeIII restriction sites within the repetitive region of gene jl8 were not represented. (B) Results of YAC contiging based on marker content of clones. A subset of 8 YACs, 2 cosmids, and 3 λ phages is shown. Marker ascertainment was carried out by PCR and dot blot hybridization. The exact order of markers has not been established, and the intermarker distances are not to scale. There are several discrepancies in the marker content of the map: markers SRE, gp85, gp90, mucin, and B11 are not contained in several YACs. Cosmids and λ phages containing genes h49 and jl8 are shown below the YACs. The YAC ends (L, left end; R, right end) were determined by inverse PCR (Materials and Methods).
Restriction mapping studies indicated that genes h49 and jl8 were located on a same 100 kb NotI restriction fragment and an additional copy of gene h49 was mapped on a 90 kb NotI fragment (Figs. 1, 2A). This additional copy of gene h49 was located ∼370 kb from gene jl8. To investigate whether loci h49 and jl8 were located near a telomeric region, agarose blocks containing whole chromosomal T. cruzi DNA were incubated with increasing amounts of exonuclease Bal31 following digestion with SfiI restriction enzyme (Fig. 3). The loci h49 were located in two SfiI fragments of 350 kb and 450 kb that were sensitive to the treatment with Bal31. On the other hand, JL8 probe reacted with two SfiI fragments (50 and 23 kb, note that there is a SifI restriction site within jl8 gene) which were not affected by the treatment with Bal31 except when large amounts of the enzyme were used. These results suggest that the gene order could be: telomere →h49→h49→jl8. The distance between the telomere and the first copy of gene h49 was estimated to be ≥200 kb.
Figure 3.
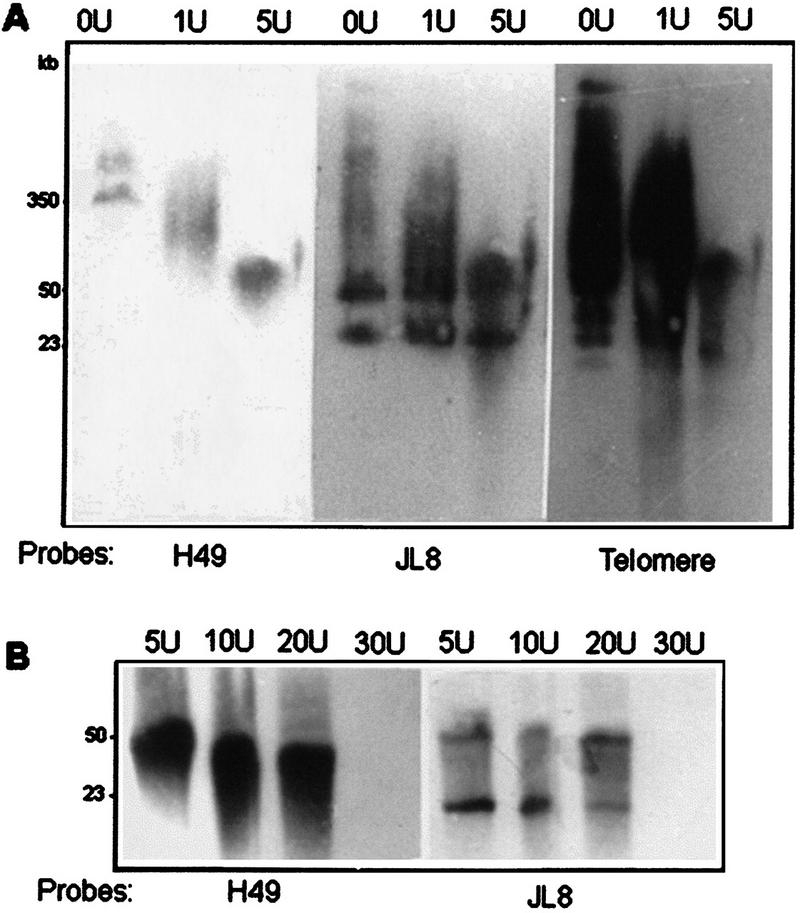
Nuclease Bal31 sensitivity of genes h49 and jl8. Agarose blocks containing whole chromosomal T. cruzi DNA were incubated with increasing amounts of exonuclease Bal31 (1–5 units in panel A, and 5–30 units in panel B) during 6 h at 30°C following digestion with SfiI restriction enzyme. DNA restriction fragments were resolved by PFGE and hybridized with probes H49 and JL8, and a T. cruzi telomere sequence.
The two size-polymorphic homologous chromosomes differ by ∼300 kb. The size changes observed in these chromosomes are not merely due to telomere repeat rearrangment events because the telomere repeat arrays of the T. cruzi chromosomes are short, ranging in size from 0.5–1.0 kb (Chiurillo et al. 1999). Whether other rearrangements are produced by amplification or deletion or by interchromosomal exchange of large DNA segments must still be determined. It is important to note that the size polymorphism of H49/JL8 antigens-encoding chromosomes was also detected in the parental CL strain maintained in different laboratories.
Isolation and Characterization of YAC Clones Harboring Genes h49 and jl8
A T. cruzi YAC library (Ferrari et al. 1997) was screened by PCR and dot blot hybridization using specific oligonucleotides derived from genes h49 and jl8. This strategy identified 6 YAC clones which were sized and characterized for content. Two YACs (insert sizes 200 kb and 400 kb) contained the gene h49 and 4 YACs (insert sizes 180–300 kb) contained the genes h49 and jl8.
To test whether the YAC clones harboring genes h49 and jl8 are mitotically stable in long term cultures, they were grown for 75 generations and examined by Southern blot analysis. No rearrangements were observed suggesting the YAC clones are propagated faithfully during mitosis. This observation was further supported by restriction analysis of h49 and jl8 loci after several rounds of propagation. The hybridization profiles obtained with these probes were identical to those obtained with T. cruzi genomic DNA (see below). These data suggest that the YAC clones carry authentic h49 and jl8 loci and are stable. Finally, to check whether more than one YAC recombinant was present in a single yeast colony, YAC clones were separated by PFGE and hybridized with 32P-labeled pBR322 sequences (present in the YAC cloning vector) or T. cruzi total genomic DNA. This assay confirmed that all colonies tested contained a single YAC recombinant.
Establishment of YAC Contig and Colinearity with the Genomic Restriction Map
To determine whether the physical maps of YACs are colinear with the genomic map, YAC DNAs were digested with EcoRI and hybridized with H49 and JL8 probes (Fig. 4). Clones Y7H8, Y9H8, Y8E1, and Y6B7 contained two EcoRI fragments of 20 kb and 9.7 kb hybridizing with the H49 probe, while clones Y7H7 and Y5G9 presented only the 20 kb EcoRI fragment. Clones Y7H7, Y5G9, Y9H8, and Y6B7 presented an additional EcoRI fragment of 30 kb that hybridized with the JL8 probe. It is interesting to note that the T. cruzi YAC library was constructed with fragments obtained by partial digestion of T. cruzi genomic DNA with EcoRI, and the genes h49 and jl8 are flanked by EcoRI restriction sites (Fig. 2A). This could explain the presence of several clones which contain only the 20 kb EcoRI fragment recognized by the H49 probe.
Figure 4.
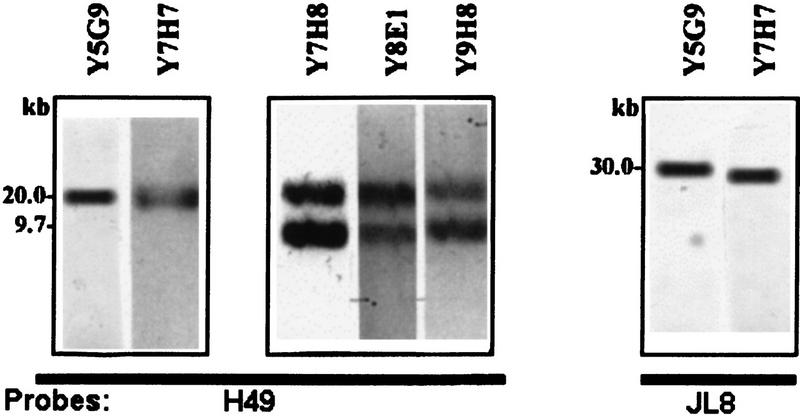
Presence of h49 and jl8 genes on YAC clones. YAC clones containing genes h49 and jl8 (Y5G9 and Y7H7) or gene h49 (Y7H8, Y8E1 and Y9H8) were digested with EcoRI, and the restriction products were resolved by gel electrophoresis, blotted, and hybridized to probes H49 and JL8.
As shown above, genomic restriction mapping indicated that genes h49 and jl8 were located in one 100 kb NotI restriction fragment, and an additional copy of gene h49 was located in a NotI restriction fragment of ∼90 kb. The distance between the copies of the h49 genes should be ∼300 kb and according to the genomic map, YAC clones harboring two copies of the gene h49 must generated two NotI restriction fragments recognized by probe H49. This fact was demonstrated for the clone Y7H8 (data not shown).
To determine the overlapping sections of the YACs and their localization with regard to the genomic restriction map, EST and STS markers, previously assigned to chromosomal bands XVI and XVII, were hybridized with YAC clones in a dot blot hybridization and/or detected by PCR (Table 1). The resulting contig comprises 8 overlapping YAC clones (Fig. 2B) that were aligned based on the presence or absence of 21 DNA markers: 6 repetitive DNA polymorphic sequences (SIRE, C6, SRE, retrotransposon B11, E13, 196-bp minisatellite sequence), 10 genes for known T. cruzi proteins and structural RNA (H49, JL8, gp90, gp82, gp85, trans-sialidase, cDNA Sx23, cDNA Sx42, mucin, spliced leader sequence), and 5 ESTs (TEUF0548, TEUF0068, TEUF0078, TEUF0040, TEUF0537). Subsequent screening of the T. cruzi YAC library with EST TEUF0068, a specific marker for chromosomal band XVII, allowed us to isolate two clones (Y2F9, Y2G12) that covered the region localized between the two copies of gene h49. To further confirm the distal position of clones Y7H8 and Y7H7 at the two extremes of the contig, and the relative positions of YACs, we used YAC left arm primers to amplify by PCR the insert sequences immediately adjacent to the YAC vector.The amplimer of the left arm of Y7H8 hybridized with clone Y8E1 but not with other YACs, while the left amplimer of clone Y7H7 reacted with clones Y5G9, Y9H8, and Y6B7. The contiging of YACs was also confirmed by comparison of the restriction site maps of the recombinant clones with that of T. cruzi genomic DNA (not shown).
As expected, markers belonging to repetitive DNA or multigene families encoding surface glycoproteins mapped at different sites of the YAC contig. For instance, copies of the polymorphic repetitive sequence E13 (10,000 copies/haploid genome, Requena et al. 1996) were found on both sides of the YAC contig. Genes encoding T. cruzi surface glycoproteins (gp90, gp85, gp82, cDNA Sx23, cDNA Sx42, mucin) are part of a large polymorphic group (Colli 1993; Salazar et al. 1996; DiNoia et al. 1998; Freitas et al. 1998) and they were mapped in different YACs. The repertoire of surface glycoprotein related genes is very large, gross calculations placing the figure in hundreds of copies (Peterson et al. 1989; Khan et al. 1991; Franco et al. 1993; Araya et al. 1994; DiNoia et al. 1998; Freitas et al. 1998). Our results are in agreement with previous data showing the presence of many clusters of genes encoding T. cruzi surface glycoproteins spread throughout the genome (Cano et al. 1995; Salazar et al. 1996; Chiurillo et al. 1999).
The described YAC contig represents a genuine reconstruction of a region of the genome, despite missing sequences in several YACs. The hybridization data revealed several minor differences in the gene content of the size polymorphic H49/JL8 antigens-encoding chromosomes. The presence or absence of chromosome specific markers such as the ESTs TEUF0068 and TEUF0537 for band XVII or the spliced leader sequence for band XVI, allowed us to define whether the YAC clone was derived from band XVI or XVII. For instance, clones Y7H8, Y6B7, Y2F9, and Y2G12 were derived from band XVII, whereas clones Y7H7 and Y5G9 were derived from band XVI. This result indicates the existence of some sequence polymorphism in the H49/JL8 antigens-encoding chromosomes. In this case, the chromosome size polymorphism could affect the coding regions and would not be solely confined to repeated sequence, as observed in T. brucei (Melville et al. 1998).
We have also found that some polymorphic repetitive sequences (SRE, SIRE, B11, E13) and genes encoding surface glycoproteins (mucin, gp90, Sx42, Sx23) appear to be absent in several YACs, confirming polymorphism between bands XVI and XVII. T. cruzi reproduces asexually by binary fission and sexual reproduction has never been demonstrated in this parasite (Solari 1995; Tibayrenc et al. 1986). The loss of markers on chromosomal homologs could be common in T. cruzi, possibily as a consequence of a high frequency of mitotic recombination events; in this kind of recombination chromosome pairing may not be very precise.
Interestingly, many genetic markers were concentrated in a region of about 105 kb including loci h49 and jl8 (Fig. 2B). Further confirmation of this arrangement was obtained by analysis of recombinant cosmids (insert sizes ∼30 kb) harboring h49 and/or jl8 genes, and λphage recombinant clones (insert sizes 15–20 kb) containing h49 or jl8 genes (Fig. 2B). Data indicates that the majority of the gene markers are clustered in 1/6 of the whole contig, with an average estimated marker spacing of 1 marker/6.2 kb in the 105 kb region. This might be due either to an unequal gene distribution or to the absence, by chance, of markers for the remainder of the contig. In the latter case, the cloned material present in the YAC contig shown in Figure 2B may prove useful for the isolation of those still unknown genes/markers.
Recently, Andersson et al. (1998) reported the complete sequence of a 93.4 kb contig from T. cruzi chromosome 3. This contig contains a strand-switch region where genes appear to be organized in two long clusters harboring multiple genes on the same strand, with the two clusters oriented in opposite directions. We currently are trying to determine the transcription direction of genes in the region around the h49 and jl8 loci. The mapping of T. cruzi chromosomes indicates the complexity of the genomic DNA in this organism. We have demonstrated the existence of some sequence polymorphism in the H49/JL8 antigens-encoding chromosomes. Thus, the construction of a clone map prior or concomitantly to DNA sequencing is an essential step for the identification of homologous regions and determination of rearrangements.
METHODS
Separation of T. cruzi Chromosomal DNA by PFGE
Epimastigotes from T. cruzi clone CL Brener (Zingales et al. 1997b) were grown to late logarithmic phase (5–10 × 106/ml) and collected by centrifugation. Cells were resuspended in phosphate-buffered saline and mixed with an equal volume of 1% low-melting point agarose. Approximately 1 × 108 cells were used for each gel plug and incubated in a solution containing 0.5 m EDTA (pH 8.0), 1% sodium lauroylsarcosinate, and 1 mg/ml proteinase K at 50°C for 48 hr and stored at 4°C in 0.5 m EDTA (pH 8.0).
Chromosomal bands were separated on agarose gels in a Gene Navigator apparatus (Pharmacia) using a hexagonal electrode array (Cano et al. 1995). PFGE was carried out in 1.2% agarose gels in 0.5 × TBE (45 mm Tris45 mm boric acid, 1 mm EDTA [pH 8.3] at 13°C for 132 hr. Separation was carried out with 5 phases of homogeneous pulses with interpolation at 80 V: phase 1, pulse time 90 sec (run time 30 hr); phase 2200 sec (30 hr); phase 3350 sec (24 hr); phase 4500 sec (24 hr) and phase 5800 sec (24 hr) (Cano et al. 1995). Gels were stained with ethidium bromide (0.5 μg/ml), photographed, and transferred onto nylon filters.
Isolation of Chromosomal Bands XVI and XVII
Plugs containing whole T. cruzi chromosomal DNA were incubated for 1 hr in TE buffer (10 mm Tris-HCl [pH 8.0], 5 mm EDTA) containing 1 mm phenylmethylsufonyl fluoride and washed twice in TE buffer. Chromosomal bands XVI and XVII were separated by PFGE as described above and excised from the preparative agarose gels. Agarose gel slices were washed twice in TE buffer, twice with the restriction endonuclease buffer, and incubated for 1 hr at 4°C with the restriction endonuclease buffer. The restriction enzyme was added (50 units per slice) followed by incubation for 12 hr at the appropriate temperature. In order to check for chromosomal integrity during manipulations, a chromosomal band was manipulated exactly as described above, but without restriction enzyme.
Screening of a T. cruzi YAC Library and Analysis of YAC clones
The YAC library was constructed using DNA of epimastigotes of clone CL Brener in pYAC4 (Ferrari et al. 1997). The library consists of 2,770 individual YACs with a mean insert size of 365 kb, encompassing ∼10 genome equivalents. The library was screened by two different methods. The first involved PCR on DNA pools using primers derived from genes h49 and jl8 and subsequent hybridization of specific pool blots. The second method utilized direct hybridization of DNA probes labeled with 32P to YAC library grid blots as described previously (Ferrari et al. 1997).
Cell plugs (100 μl) were prepared in low melting point agarose from yeasts grown in overnight cultures at 30°C on YPD liquid medium. The plugs were incubated at 30°C for 3 hr in SCEM buffer (1 m sorbitol, 50, mm EDTA, 0.1 m sodium citrate, and 7 mm β-mercaptoethanol) with zymolyase (10 units/plug) at 37°C. They were then washed and treated with 1 ml PKB [0.1 m NaCl, 50 mm EDTA (pH 8.0), 1% N-laurylsarcosine] with 1 mg/ml of proteinase K at 50°C overnight and finally washed several times with 0.5× TBE to remove proteinase K. Yeast chromosomes were separated by electrophoresis on 1% low-melting agarose gels in 0.5 × TBE using a contour-clamped homogenous electric field (CHEF DRIII, BioRad) apparatus with pulse times of 60–90 sec for 18 hr at 12°C, 220 V. After migration, the gels were stained with ethidium bromide and transferred to nylon membranes. Membranes were hybridized overnight with radiolabeled probes at 42°C in 50% formamide, 5× Denhart's solution, 5× SSC (1× SSC = 0.15 m NaCl, 0.015 m sodium citrate), 50 μg/ml yeast tRNA, 100 μg/ml sonicated herring sperm DNA and 0.1% SDS. After hybridization, filters were washed in 0.1× SSC, 0.1% SDS at 55°C for 1 hr.
YAC DNA embedded in agarose was digested with restriction enzymes, separated on agarose gels, and transferred to nylon membranes. Radiolabed probes were used for hybridization as described above. For PCR assays, plugs containing YAC DNA were treated with agarase (Epicentre Technologies, France); the DNA was diluted in water, boiled for 10 min, and stored at − 70°C. PCR reactions were carried out in 50 μl reaction volume mixtures containing 200 μm of each deoxyribonucleotide triphosphate, 1 × Taq polymerase buffer (Perkin Elmer, CA), and 1.25 units of Taq polymerase. The DNA was denaturated at 94°C, and 30 cycles of amplification were performed with denaturation at 94°C for 1 min, annealing temperature specific to each primer for 1 min, and extension at 72°C for 30 sec. A final extension was performed at 72°C for 6 min. The products were separated by electrophoresis on agarose gels and visualized by ethidium bromide staining. DNA fragments were transferred to nylon membranes and hybridized with specific probes.
Vector end PCR was performed as described by Ochman et al. (1988). DNA was digested with HaeIII and digested fragments were ligated. PCR was perfomed on the ligated fragments using primers Y5U and Y5R (Joslyn et al. 1991), specific for the left arm of vector pYAC-4, thus amplifying the YAC end fragments. The PCR products were isolated, radiolabeled, and hybidized against the other YACs.
Screening of T. cruzi Cosmid and Lambda Phage Libraries
A chromosome-specific cosmid library was constructed using DNA fragments from the chromosomal bands XVI and XVII purified from PFGE preparative gels. The chromosomal bands were partially digested with BamHI and inserted into the BamHI cloning site of cosmid Lawrist 4. Ligation, packaging, and transfection were carried out as described by Hanke et al. (1996). A T. cruzi (clone CL Brener) genomic library constructed in lambda phage FixII vector was kindly provided by E. Rondinelli (UFRJ, Rio de Janeiro, Brasil) through the T. cruzi genome network. Cosmid and lambda libraries were screened by hybridization with H49 and JL8 probes.
DNA Probes
ESTs derived from an epimastigote cDNA library (clone CL Brener) were obtained as cDNA fragments by PCR (Brandão et al. 1997). All partially sequenced cDNAs (ESTs) are identified by TEUF number originating in the sequencing laboratory (Table 1). For other cloned T. cruzi DNA sequences (Table 1), the inserts of recombinant plasmids were excised by digestion with appropriate restriction enzymes, purified by gel electrophoresis, and radiolabeled with 32P. Hybridization of the probes with the chromoblots was carried out as described previously (Cano et al. 1995).
Acknowledgments
We thank D. Cohen, D. LePaslier, and J. Dausset (CEPH/Foundation Jean Dausset, Paris, France) for their help and encouragement. This work was supported by grants from FAPESP and Pronex (Brasil); Ibero American Project of Biotechnology, CYTED (Spain); World Bank/UNDP/WHO TDR; CONICIT S1-95000524 (Venezuela); Projet Genome T. cruzi-INGEBI-F. Jean Dausset-CEPH, Ministere d'Affaires Etrangeres (France); Centro Argentino Brasileiro de Biotecnologia (CABBIO); University of Buenos Aires-Project 01/IX-17, 98-2000 (CyT-UBA); CONICET and FONCyT, project BID802/OC-AR PICT 01-00000-01421, Argentina. MRMS is a postdoctoral research fellow of FAPESP and MJL is a John Simon Guggenheim Foundation Fellow (period 98-99).
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact.
Footnotes
E-MAIL franco.dmip@epm.br; FAX 55-11-571-1095.
REFERENCES
- Andersson B, Aslund L, Tammi M, Tran A, Hoheisel JD, Petersson U. Complete sequence of a 93.4kb contig from chromosome 3 of Trypanosoma cruzi containing a strand-switch region. Genome Res. 1998;8:809–816. doi: 10.1101/gr.8.8.809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Araya JE, Cano MI, Yoshida N, Franco da Silveira J. Cloning and characterization of a gene for the stage-specific gp82 kDa surface antigen of metacyclic trypomastigotes of Trypanosoma cruzi. Mol Biochem Parasitol. 1994;65:161–169. doi: 10.1016/0166-6851(94)90124-4. [DOI] [PubMed] [Google Scholar]
- Brandão A, Urmenyi T, Rondinelli E, Gonzalez A, Miranda AB, Degrave W. Identification of transcribed sequences (ESTs) in the Trypanosoma cruzi genome project. Mem Inst Oswaldo Cruz. 1997;92:863–866. doi: 10.1590/s0074-02761997000600024. [DOI] [PubMed] [Google Scholar]
- Brener Z. Biology of Trypanosoma cruzi. Ann Rev Microbiol. 1973;27:345–382. doi: 10.1146/annurev.mi.27.100173.002023. [DOI] [PubMed] [Google Scholar]
- Buscaglia CA, Campetella O, Leguizamon MS, Frasch ACC. The repetitive domain of Trypanosoma cruzi trans-sialidase enhances the immune response against the catalytic domain. J Infect Dis. 1999;77:431–436. doi: 10.1086/514199. [DOI] [PubMed] [Google Scholar]
- Cano MI, Gruber A, Vasquez M, Cortes A, Levin MJ, Gonzalez A, Degrave WM, Rondinelli E, Zingales B, Ramirez JL, et al. Molecular karyotype of clone CL Brener chosen for the Trypanosoma cruzi genome project. Mol Biochem Parasitol. 1995;71:273–278. doi: 10.1016/0166-6851(95)00066-a. [DOI] [PubMed] [Google Scholar]
- Carvalho MR, Krieger MA, Oeleman W, Shikanai-Yassuda MA, Ferreira AW, Pereira JB, Saez-Alquezar A, Dorlhiac-Llacer DF, Chamone DF, Goldenberg S. Chagas' disease diagnosis: Evaluation of several tests in blood bank screening. Transfusion. 1993;33:830–834. doi: 10.1046/j.1537-2995.1993.331094054620.x. [DOI] [PubMed] [Google Scholar]
- Chiurillo MA, Cano MI, Franco da Silveira J, Ramirez JL. Organization of telomeric and sub-telomeric regions of chromosomes from the protozoan parasite Trypanosoma cruzi. Mol Biochem Parasitol. 1999;10:173–182. doi: 10.1016/s0166-6851(99)00047-x. [DOI] [PubMed] [Google Scholar]
- Colli W. Trans-sialidase: A unique enzyme activity discovered in the protozoan Trypanosoma cruzi. FASEB J. 1993;7:1257–1264. doi: 10.1096/fasebj.7.13.8405811. [DOI] [PubMed] [Google Scholar]
- Cotrim PC, Paranhos GP, Mortara RA, Wanderley J, Rassi A, Camargo ME, Franco da Silveira J. Expression in Escherichia coli of a dominant immunogen of Trypanosoma cruzi recognized by human chagasic sera. J Clin Microbiol. 1990;28:519–524. doi: 10.1128/jcm.28.3.519-524.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotrim PC, Paranhos-Baccala G, Santos MR, Mortensen C, Cano MI, Jolivet M, Camargo ME, Mortara RA, Franco da Silveira J. Characterization of a high molecular weight immunodominant antigen associated to the cytoskeleton of Trypanosoma cruzi. Mol Biochem Parasitol. 1995;71:89–98. doi: 10.1016/0166-6851(95)00036-z. [DOI] [PubMed] [Google Scholar]
- Di Noia JM, D'Orso I, Aslund L, Sanchez DO, Frasch ACC. The Trypanosoma cruzi mucin family is transcribed from hundreds of genes having hypervariable regions. J Biol Chem. 1998;273:10843–10850. doi: 10.1074/jbc.273.18.10843. [DOI] [PubMed] [Google Scholar]
- Ferrari I, Brandariz S, Santos MR, Lorenzi H, Requena JM, Schijman A, Vazquez M, Franco da Silveira J, Ben Dov C, Medrano C, et al. Towards the physical map of Trypanosoma cruzi nuclear genome: Construction of YAC and BAC libraries of the reference clone T. cruzi CL Brener. Mem Inst Oswaldo Cruz. 1997;92:843–858. doi: 10.1590/s0074-02761997000600021. [DOI] [PubMed] [Google Scholar]
- Franco FRS, Paranhos-Baccala G, Yamauchi LM, Yoshida N, Franco da Silveira J. Characterization of a cDNA clone encoding the carboxy-terminal domain of 90 kDa surface antigen of Trypanosoma cruzi metacyclic trypomastigotes. Infect Immun. 1993;61:4196–4201. doi: 10.1128/iai.61.10.4196-4201.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frasch ACC, Cazzulo JJ, Aslund L, Pettersson U. Comparison of genes encoding Trypanosoma cruzi antigens. Parasitol Today. 1991;7:148–151. doi: 10.1016/0169-4758(91)90284-u. [DOI] [PubMed] [Google Scholar]
- Freitas-Junior LH, Briones MR, Schenkman S. Two distinct groups of mucin-like genes are differentially expressed in the developmental stages of Trypanosoma cruzi. Mol Biochem Parasitol. 1998;15:101–114. doi: 10.1016/s0166-6851(98)00025-5. [DOI] [PubMed] [Google Scholar]
- Frohme M, Hanke J, Aslund L, Petersson U, Hoheisel JD. Selective generation of chromosomal cosmid libraries within the Trypanosoma cruzi project. Electrophoresis. 1998;19:478–481. doi: 10.1002/elps.1150190404. [DOI] [PubMed] [Google Scholar]
- Hanke J, Sanchez DO, Henriksson J, Aslund L, Petersson U, Frasch ACC, Hoheisel JD. Mapping the Trypanosome cruzi genome: Analyses of representative cosmid libraries. Biotechniques. 1996;21:686–693. doi: 10.2144/96214rr01. [DOI] [PubMed] [Google Scholar]
- Hanke J, Frohme M, Laurent JP, Swindle J, Hoheisel JD. Hybridization mapping of Trypanosome cruzi chromosomes III and IV. Electrophoresis. 1998;19:482–485. doi: 10.1002/elps.1150190405. [DOI] [PubMed] [Google Scholar]
- Henriksson J, Aslund L, Macina RA, Cazullo BMF, Cazzulo JJ, Frasch ACC, Pettersson U. Chromosome specific markers reveal conserved linkage groups in spite of extensive size variation in Trypanosoma cruzi. Mol Biochem Parasitol. 1995;73:64–73. doi: 10.1016/0166-6851(95)00096-j. [DOI] [PubMed] [Google Scholar]
- Henriksson J, Aslund L, Pettersson U. Karyotype variability in Trypanosoma cruzi. Parasitol Today. 1996;12:108–114. doi: 10.1016/0169-4758(96)80670-3. [DOI] [PubMed] [Google Scholar]
- Hoft DF, Kim KS, Otsu K, Moser DR, Yost WJ, Blumin JH, Donelson JE, Kirchhoff LV. T. cruzi expresses diverse repetitive protein antigens. Infect Immun. 1989;57:1957–1967. doi: 10.1128/iai.57.7.1959-1967.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoft DF, Donelson JE, Kirchhoff LV. Repetitive protein antigens of Trypanosoma cruzi have diverse intracellular locations. J Parasitol. 1995;81:549–554. [PubMed] [Google Scholar]
- Ibanez CF, Affranchino JL, Macina RA, Reys MB, Lequizamon S, Camargo ME, Aslund U, Petterson U, Frasch AAC. Multiple Trypanosoma cruzi antigens containing tandemly amino acid sequence motif. Mol Biochem Parasitol. 1988;30:27–34. doi: 10.1016/0166-6851(88)90129-6. [DOI] [PubMed] [Google Scholar]
- Joslyn G, Carlson M, Thliveris A, Albertsen H, Gelbert L, Samowitz W, Groden J, Stevens J, Spirio L, et al. Identification of deletion mutations and three new genes at the familial polyposis locus. Cell. 1991;66:601–613. doi: 10.1016/0092-8674(81)90022-2. [DOI] [PubMed] [Google Scholar]
- Kahn S, Van Voorhis WC, Eisen H. The major 85 kDa surface antigen of the mammalian form of Trypanosoma cruzi is encoded by a large heterogeneous familiy of simultaneously expressed genes. J Exp Med. 1990;172:589–597. doi: 10.1084/jem.172.2.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kemp DJ, Coppel RL, Anders RF. Repetitive proteins and genes of malaria. Ann Rev Microbiol. 1987;41:181–207. doi: 10.1146/annurev.mi.41.100187.001145. [DOI] [PubMed] [Google Scholar]
- Krieger M A, Almeida E, Oelemann W, Lafaille JJ, Pereira JB, Krieger H, Carvalho MR, Goldenberg S. Use of recombinant antigens for the accurate immunodiagnosis of Chagas' disease. Am J Trop Med Hyg. 1992;46:427–434. doi: 10.4269/ajtmh.1992.46.427. [DOI] [PubMed] [Google Scholar]
- Lafaille JJ, Linss J, Krieger MA, Souto-Padron T, Souza W, Goldenberg S. Structure and expression of two Trypanosoma cruzi genes encoding antigenic protein bearing repetitive epitopes. Mol Biochem Parasitol. 1989;35:127–136. doi: 10.1016/0166-6851(89)90115-1. [DOI] [PubMed] [Google Scholar]
- Levin MJ, Mesti EA, Benarous R, Levitus G, Schijman A, Levy-Yeyati P, Ruiz A, Kahan A, Rosembaum MB, Torres HN, Segura EL. Identification of major Trypanosoma cruzi antigenic determinants in chronic Chagas' heart disease. Am J Trop Med Hyg. 1989;41:530–539. doi: 10.4269/ajtmh.1989.41.530. [DOI] [PubMed] [Google Scholar]
- Levin MJ, Franco da Silveira J, Frasch ACC, Camargo ME, Lafon S, Degrave WM, Rangel-Aldao R. Recombinant antigens and Chagas' disease diagnosis: Analysis of a workshop. FEMS Microbiol Immunol. 1991;89:11–20. doi: 10.1111/j.1574-6968.1991.tb04965.x. [DOI] [PubMed] [Google Scholar]
- Melville SE, Leech V, Gerrard CS, Tait A, Blackwell JM. The molecular karyotype of the megabase chromosomes of Trypanosoma brucei and the assignment of chromosome markers. Mol Biochem Parasitol. 1998;94:155–173. doi: 10.1016/s0166-6851(98)00054-1. [DOI] [PubMed] [Google Scholar]
- Ochman H, Gerber AS, Hartl DL. Genetic application of an inverse polymerase chain reaction. Genetics. 1988;120:621–623. doi: 10.1093/genetics/120.3.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paranhos GS, Santos MRM, Cotrim PC, Rassi A, Jolivet M, Camargo ME, Franco da Silveira J. Detection of antibodies in sera from Chagas' disease patients using a T. cruzi immunodominant recombinant antigen. Parasite Immunol. 1994;16:165–169. doi: 10.1111/j.1365-3024.1994.tb00336.x. [DOI] [PubMed] [Google Scholar]
- Pastini AC, Iglesias SR, Carricarte VC, Guerin ME, Sanchez DO, Frasch AAC. Immunoassay with recombinant Trypanosoma cruzi antigens potentially useful for screening donated blood and diagnosing Chagas' disease. Clin Chem. 1994;40:1893–1897. [PubMed] [Google Scholar]
- Pereira CM, Yamauchi LM, Levin MJ, Franco da Silveira J, Castilho BA. Mapping of B cell epitopes in an immunodominant antigen of Trypanosoma cruzi using fusions to Escherichia coli lamB protein. FEMS Microbiol Letters. 1998;164:125–131. doi: 10.1111/j.1574-6968.1998.tb13077.x. [DOI] [PubMed] [Google Scholar]
- Peterson DS, Fouts DL, Manning JE. The 85kd surface antigen gene of Trypanosoma cruzi is telomeric and a member of a multigene family. EMBO J. 1989;8:3911–3916. doi: 10.1002/j.1460-2075.1989.tb08571.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Requena JM, Lopez MC, Alonso C. Genomic repetitive DNA elements of Trypanosoma cruzi. Parasitol Today. 1996;12:279–283. doi: 10.1016/0169-4758(96)10024-7. [DOI] [PubMed] [Google Scholar]
- Salazar NA, Mondragon A, Kelly JM. Mucin-like glycoprotein genes are closely linked to members of the trans-sialidase super-family at multiple sites in the Trypanosoma cruzi genome. Mol Biochem Parasitol. 1996;78:127–136. doi: 10.1016/s0166-6851(96)02617-5. [DOI] [PubMed] [Google Scholar]
- Santos MRS, Cano MI, Schijman A, Vazquez M, Levin MJ, Ramirez JL, Brandão A, Degrave WM, Franco da Silveira J. Trypanosoma cruzi genome project: Nuclear karyotype and gene mapping of clone CL Brener. Mem Inst Oswaldo Cruz. 1997;92:821–828. doi: 10.1590/s0074-02761997000600018. [DOI] [PubMed] [Google Scholar]
- Solari AJ. Mitosis and genome partition in trypanosomes. BioCell. 1995;19:65–84. [PubMed] [Google Scholar]
- Tibayrenc M, Ward P, Moya A, Ayala FJ. Natural populations of Trypanosoma cruzi, the agent of Chagas disease, have a complex multiclonal structure. Proc Natl Acad Sci. 1986;83:115–119. doi: 10.1073/pnas.83.1.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner CMR, Melville SE, Tait A. A proposal for karyotype nomenclature in Trypanosome brucei. Parasitol Today. 1997;13:5–6. doi: 10.1016/s0169-4758(96)20056-0. [DOI] [PubMed] [Google Scholar]
- Umezawa ES, Bastos S, Camargo ME, Yamauchi LM, Santos MR, Gonzalez A, Zingales B, Levin MJ, Sousa O, Rangel-Aldao R, Franco da Silveira J. Evaluation of recombinant antigens for Chagas' disease serodiagnosis in South and Central America. J Clin Microbiol. 1999;37:1554–1560. doi: 10.1128/jcm.37.5.1554-1560.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zingales B, Rondinelli E, Degrave W, Franco da Silveira J, Levin M, LePaslier D, Modabber F, Dobrokhotov B, Swindle J, Kelly JM, et al. The Trypanosoma cruzi genome initiative. Parasitol Today. 1997a;13:16–22. [Google Scholar]
- Zingales B, Pereira MES, Oliveira RP, Almeida KA, Umezawa ES, Souto RP, Vargas N, Cano MI, Franco Da Silveira J, Nehme NS, et al. Trypanosoma cruzi genome project: biological characteristics and molecular typing of clone CL Brener. Acta Tropica. 1997b;68:159–173. doi: 10.1016/s0001-706x(97)00088-0. [DOI] [PubMed] [Google Scholar]