

Abstract
Activated blood platelets mediate the primary response to vascular injury. Although molecular abnormalities of platelet proteins occur infrequently, taken collectively, an inherited platelet defect accounts for a bleeding diathesis in ≈1:20,000 individuals. One rare example of a platelet disorder, Glanzmann thrombasthenia (GT), is characterized by life-long morbidity and mortality due to molecular abnormalities in a major platelet adhesion receptor, integrin αIIbβ3. Transfusion therapy is frequently inadequate because patients often generate antibodies to αIIbβ3, leading to immune-mediated destruction of healthy platelets. In the most severe cases allogeneic bone marrow transplantation has been used, yet because of the risk of the procedure it has been limited to few patients. Thus, hematopoietic stem cell gene transfer was explored as a strategy to improve platelet function within a canine model for GT. Bleeding complications necessitated the use of a mild pretransplant conditioning regimen; therefore, in vivo drug selection was used to improve engraftment of autologously transplanted cells. Approximately 5,000 αIIbβ3 receptors formed on 10% of platelets. These modest levels allowed platelets to adhere to αIIbβ3’s major ligand (fibrinogen), form aggregates, and mediate retraction of a fibrin clot. Remarkably, improved hemostatic function was evident, with ≤135-fold reduced blood loss, and improved buccal bleeding times decreased to 4 min for up to 5 y after transplant. One of four transplanted dogs developed a significant antibody response to αIIbβ3 that was attenuated effectively with transient immune suppression. These results indicate that gene therapy could become a practical approach for treating inherited platelet defects.
Keywords: hematology, recombinant lentivirus-mediated gene transfer, hemostasis
There are several well-characterized inherited genetic defects that affect various aspects of platelet function (activation, adhesion, aggregation, signal transduction, granule storage), which usually manifest themselves clinically as a failure to control bleeding (1). One of the inherited platelet anomalies, Glanzmann thrombasthenia (GT) (thrombasthenia meaning “weak platelets”), was first reported by the Swiss pediatrician Edward Glanzmann in 1918 (2). Although GT is a very rare disorder, occurring in ≈1:1,000,000 individuals worldwide, research efforts have led to the characterization of more than 170 distinct genetic abnormalities within the platelet-specific integrin αIIbβ3 (also known as glycoprotein GPIIb–IIIa complex and INTGA2BB3), resulting in quantitative and qualitative deficiencies of the receptor on the platelet surface (http://sinaicentral.mssm.edu/intranet/research/glanzmann/menu). Because αIIbβ3 is the major receptor for the adhesive ligand fibrinogen, inherited molecular defects of αIIbβ3 prevent platelets from aggregating at the site of a vascular injury to repair damaged blood vessels (3). As a result, the patient experiences severe intermittent mucocutaneous bleeding episodes, beginning at birth and recurring throughout the individual's lifetime. This disorder is particularly life-threatening when patients experience gastrointestinal bleeding or severe bleeding induced by trauma. The current treatment options for GT are transfusion with normal human platelets or use of therapeutic agents (Amicar, Novo7, tranexamic acid) to stop excessive hemorrhage (4). Unfortunately, patients frequently develop antibodies and destroy transfused platelets, and hemostatic agents are usually extremely expensive and their beneficial effects relatively short-lived. Allogeneic bone marrow transplant has been used successfully to restore hemostasis in dogs and humans with GT (5, 6), although leukocyte antigen-compatible human donors are difficult to locate, and donor graft vs. host inconsistencies have produced adverse side effects, which makes this a controversial treatment option. We hypothesized that an autologous transplant of hematopoietic stem cells transduced with genes encoding normal integrin αIIb or β3 may be an ideal approach for correction of GT because de novo synthesis of a biologically normal integrin αIIbβ3 within megakaryocytes should allow trafficking of the entire receptor to the cell surface to allow platelets to participate in wound repair. This is supported by the recent success of using hematopoietic stem cell gene transfer of integrin β2 to generate expression of αLβ2, αMβ2, and αXβ2 on leukocytes for correction of canine leukocyte adhesion deficiency (CLAD) (7).
We previously reported that transduction of G-CSF–mobilized peripheral blood stem cells (G-PBC) with an oncoretrovirus vector encoding integrin β3 generated de novo synthesis of viable integrin αIIbβ3 complexes on megakaryocytes derived from human GT patients (8). Next we showed that platelet function could be corrected within a murine model for GT by transplantation of bone marrow transduced with a lentivirus vector encoding β3 (9). The current investigation expands on this knowledge by demonstrating the development of a clinically relevant strategy for transferring genes into G-PBC to correct platelet function and improve hemostasis within a large-animal model for GT. To accomplish this goal, we used a line of Great Pyrenees affected with GT resulting from an insertion of 14 nucleotides in exon 13 and a splicing defect in intron 13 of the integrin αIIb gene (10). This causes a shift in the reading frame of αIIb and premature truncation of the subunit, leading to a deficiency of αIIbβ3 on the platelet surface and loss of platelet function. Canine GT is essentially identical to human GT in its clinical presentation, characterized by severe intermittent episodes of purpura, epistaxis, and gingival and gastrointestinal hemorrhage. Thus, the outcome of this study should provide a wealth of information toward the potential feasibility for human gene therapy of GT.
Results and Discussion
To investigate whether hematopoietic stem cell gene transfer could alleviate GT in αIIb-deficient (−/−) dogs, cDNA encoding human αIIb was subcloned into a HIV type 1 lentivirus vector (Fig. 1A). The vector is self-inactivating, because an enhancer/promoter (U3) was removed from the 3′ viral LTR (U3/R/U5), and an 889-nt fragment of the human αIIb gene promoter was used to target transcription within megakaryocytes. This promoter binds Ets and GATA factors to instill a high level of megakaryocyte gene transcription and has a repressor region that inhibits expression within other lineages (11). Because expression of αIIb is not known to confer a growth advantage to megakaryocytes, the lentivirus construct has another cDNA cassette encoding the murine stem cell virus gene promoter driving expression of a drug-resistant protein, P140K methylguanine methyltransferase (MGMT), to allow in vivo enrichment of lentivirus-transduced hematopoietic stem cells (12).
Fig. 1.

Lentivirus transduction of GT CD34+ PBC generates integrin αIIbβ3 expression on platelets. (A) Diagram of HIV type 1 lentivirus vector, αIIb-(M)WPT. The 5′ LTR is intact (Left, blue box), whereas the U3 enhancer/promoter of the viral 3′ LTR (Right, blue box) was removed to allow the integrin αIIb gene promoter (large gray arrow) to direct megakaryocyte-specific synthesis of integrin αIIb (small yellow circle) in GT dogs with αIIb deficiency. The αIIb gene promoter binds GATA and Ets transcription factors for high-level gene expression in megakaryocytes, and there is also a repressor region that inhibits gene expression in other lineages. The murine stem cell virus promoter (small gray arrow) was used to drive lineage-nonspecific expression of a drug-selectable, biologically stable form of a DNA repair protein, MGMT (yellow oval) to allow in vivo enrichment of lentivirus-transduced hematopoietic stem cells. WPRE enhanced the efficiency of transgene expression (green diamond). (B) Immunofluorescent flow cytometric histograms of platelets isolated from circulating whole blood of transplant recipients. A representative histogram from dog C at 0.5 y after transplant (Tx, black line) revealed that the MFI (in parentheses) for platelets stained with a fluorochrome-conjugated nonspecific Ig (Upper Left) remained negative and equal to αIIb (−/−, turquoise; +/−, gray; +/+, navy) control animals. Remarkably, the MFI for platelets stained with an Ab specific for integrin αIIb (Upper Right), β3 (Lower Left), and the αIIbβ3 complex (Lower Right) revealed that transplant dog C expressed αIIb, β3, and αIIbβ3 to the surface at intermediate MFI levels compared with the MFI of platelets from αIIb(−/−,+/−,+/+) controls. The MFI of the transplant dogs was ≈6% of normal levels, with the complex specific antibody indicating that ≈5,000 αIIbβ3 receptors were present on the platelet surface. (C) The efficiency of transduction and duration of αIIbβ3 expression was determined by immunofluorescent flow cytometric analysis measuring the percentage of platelets with an MFI over background levels observed in an αIIb(−/−) dog in the presence of saturating Ab specific for αIIb, β3, and αIIbβ3 (used in B) shortly before transplant of dogs A, B, and C until 2, 4, and 5 y after transplant, respectively. GT dogs A, B, and C received an autologous transplant of CD34+ PBC carrying a lentivirus encoding integrin αIIb and a drug-resistant protein (P140K MGMT) and then were treated three times with cytotoxic drugs including O6-BG (100 mg) and carmustine (0.25 mg/kg, 0.40 mg/kg, and 0.45 mg/kg, respectively) (blue arrows). The percentage of platelets that expressed αIIbβ3 stabilized to ≈10% of the total platelet population in dogs A, B, and C.
Canine hematopoietic stem cells were mobilized from the bone marrow into the peripheral blood with cytokines (G-CSF and stem cell factor), and PBC apheresis was performed without adverse incident as described previously with in vitro studies using human GT patients (8). Mononuclear lymphocytes were isolated with Ficoll Paque Plus (GE Healthcare) from the apheresis product, and then canine CD34 antigen-positive cells were purified by immunomagnetic selection (13). Results shown in Table 1 summarize the conditions for autologous transplant of four GT dogs given ≈2.5 × 106 αIIb-transduced PBC per kilogram of body weight at a purity of 88% for canine CD34+ antigen.
Table 1.
Conditions for autologous transplant of αIIb-CD34+ PBC
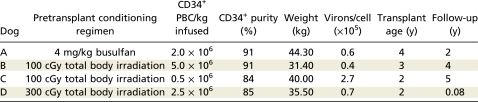 |
A nonmyeloablative pretransplant conditioning regimen was used to help create a niche in the bone marrow for the newly transplanted cells to engraft. The intensity of the conditioning regimen is determined by the level at which the dose becomes toxic to the organs. Earlier studies performed with normal canine models have demonstrated that stable allogeneic mixed donor/host hematopoietic chimerism can be safely established by the administration of a sublethal dose of 200 cGy total body irradiation followed by transient immunosuppression with mycophenolate mofetil and cyclosporine after major histocompatibility complex identical marrow transplantation (14). However, this pretransplant conditioning regimen seemed inappropriate for animals with GT, because one of four dogs died from uncontrolled internal bleeding shortly after allogeneic bone marrow transplant (5). This complication was confirmed when one of the animals enrolled into our study (dog D) died from an intracranial hemorrhage 3 wk after autologous transplant of αIIb-transduced PBC (Table 1). As a result, a new conditioning regimen consisting of a very low dose of 100 cGy total body irradiation was given to two dogs without incident (Table 1). As an alternative approach, one dog was safely administered a mild dose of busulfan at 4 mg/kg (a drug preferentially toxic to hematopoietic stem cells) for pretransplant conditioning, as described in a recent study that demonstrated successful use of busulfan at 10 mg/kg for hematopoietic stem cell gene transfer to correct CLAD (7).
Immunofluorescent analysis with monoclonal antibodies recognizing αIIb, β3, and the αIIbβ3 complex demonstrated that the receptor could be generated on the surface of a subset of peripheral blood platelets after transduction and transplantation of αIIb-transduced PBC into GT dogs (Fig. 1B, black peak). As expected, a low level of normal β3 was detected on the surface of platelets from αIIb-deficient dogs [mean fluorescence intensity (MFI) = 15], because previous studies of GT patients with αIIb defects revealed that β3 can pair with the integrin αv-subunit to form the vitronectin receptor (αvβ3) on platelets (Fig. 1B, Lower Left) (15). The histograms for αIIb (Fig. 1B, Upper Right) and αIIbβ3 complex (Fig. 1B, Lower Right) indicate that a modest level of receptors (≈5,000) formed on the surface of genetically modified platelets, because the MFI was 6% of the level integrin complex detected on normal αIIbβ3 (+/+) canine platelets (80,000 receptors) at saturating antibody concentrations. The data demonstrate successful synthesis of a recombinant normal human αIIb subunit that formed a stable complex with endogenous canine β3 and trafficked to the surface of canine platelets. It is unknown whether the surface receptor level could be improved by the use of canine αIIb for PBC gene transfer. This study used a lentivirus vector encoding human αIIb because it shares a very high similarity of ≥83% with canine αIIb nucleotide and deduced amino acid sequences. The comparison even reaches 92–100% in the β-propeller where the receptor binds to its ligand (16). This is further supported by the demonstration that the chimeric human αIIb–canine β3 receptor has formed a structure that is recognized by a complex-specific antibody (7E3) that has previously been shown to recognize both human and canine αIIbβ3 endogenous receptors, suggesting that the human and canine receptors are very similar in structure (17).
After lentivirus transduction and transplant of PBC, each animal received three courses of a combination of drugs (O6-benzylguanine and carmustine) known to be toxic to untransduced hematopoietic stem cells, to allow in vivo enrichment of PBC genetically modified with the drug-resistant protein P140K MGMT as previously used by others (Fig. 1C, blue arrows) (18). This led to a significant increase in the number of platelets that expressed αIIbβ3 (peaking at 30–40%), which stabilized at ≈10% of the total platelet population in transplant dogs A, B, and C. Integrin αIIbβ3 was detected by immunofluorescence on a subset of platelets for 2, 4, and 5 y, respectively, indicating long-term engraftment of a significant population of progenitor or parental hematopoietic cells within the bone marrow compartment of dogs A, B, and C (Fig. 1C).
One hallmark of GT is a failure of platelets to aggregate upon stimulation with physiological agonists of platelet activation (19). As a test for improved function, peripheral blood platelets were collected from each dog to determine whether the αIIbβ3 integrin complex could become activated, bind fibrinogen, and form platelet aggregates ex vivo. Identical to humans, platelets from GT dogs (−/−) failed to aggregate compared with carrier (+/−) and normal (+/+) animal platelets, which aggregated appreciably when stimulated in vitro with a mixture of platelet activation agonists [ADP, epinephrine, and canine thrombin receptor activation peptide (TRAP)1, -3, and -4] (Fig. 2A). Remarkably, all three GT dogs transplanted with αIIb-transduced PBC contained a subset of platelets (≈10%) that demonstrated a measurable ability to aggregate after activation with the agonist mixture (Fig. 2A). Note that this result is consistent with the percentage of platelets that were detected expressing integrin αIIbβ3 by flow cytometric analysis (Fig. 1C). The outcome from this analysis indicates that intracellular pathways leading to integrin αIIbβ3 inside-out signaling were operational in GT platelets that expressed αIIbβ3 on their surface.
Fig. 2.

Integrin/platelet function was corrected in recipients of αIIb-transduced CD34+ PBC. (A) Aggregation was measured after incubation of washed platelets with fibrinogen and a mixture of activation agonists (ADP, epinephrine, and canine TRAP1, -3, and -4). Platelets isolated from circulating whole blood of αIIb-deficient dogs (−/−) failed to aggregate, whereas the platelets collected from αIIb (+/−, +/+) control animals aggregated to nearly 100% using identical agonist concentrations and reaction conditions. Platelets from αIIb-transduced CD34+ PBC autologous transplant recipients A, B, and C aggregated in direct correlation with the percentage of platelets that expressed αIIbβ3 on their surface (Fig. 1C). Similar results were observed for each transplant dog in at least six separate experiments. (B) An adhesion assay was performed ex vivo to determine whether integrin outside-in signaling initiated by αIIbβ3 interaction with immobilized fibrinogen was corrected in transplant A, B, or C platelets. Washed platelets from αIIb(+/+,−/−) control and transplant dogs A, B, and C were plated onto 100 μg/mL immobilized fibrinogen and allowed to spread for 45 min at 37 °C. αIIb(+/+) platelets bound and spread appreciably onto the fibrinogen, whereas αIIb(−/−) platelets failed to adhere to the fibrinogen. In contrast, transplant dogs A, B, and C platelets bound and spread on fibrinogen-coated plates. This result is representative of three separate experiments. (Magnification, 400×.) (C) Blood was collected from each animal, mixed with human thrombin to allow a fibrin clot to form, and then samples were incubated at 37 °C for up to 24 h. Photographs taken at 3–6 h after addition of thrombin show that normal αIIb(+/+) and carrier αIIb(+/−) samples retracted an appreciable amount of the clot because plasma became readily visible within the test tube. In contrast, the fibrin blood clot remained unchanged within the sample from the αIIb(−/−) GT control dog. In contrast, blood from dogs A, B, and C mediated significant retraction of a fibrin clot similar to the positive controls. This image is representative of the results for dogs A, B, and C from three, seven, and eight separate experiments, respectively, performed for up to 2, 4, and 5 y after PBC transplant, as described in Materials and Methods.
Platelets were placed in a tissue culture plate treated with fibrinogen to determine whether newly expressed integrin αIIbβ3 could adhere to its major ligand, fibrinogen, leading to platelet spreading. As expected, platelets from a normal dog (+/+) adhered and spread on the fibrinogen-coated surface (Fig. 2B). In contrast, GT (−/−) platelets failed to adhere to fibrinogen-coated plates (Fig. 2B). Differently, platelets from transplant dogs A, B, and C (Fig. 2B) showed an improved ability to adhere and spread on immobilized fibrinogen, indicating that outside-in signaling through integrin αIIbβ3 was functional.
Another trademark of human GT is the failure of patient platelets to retract a fibrin clot. Circulating peripheral blood platelets from αIIb-transduced PBC transplant recipient dogs A, B, and C were able to mediate appreciable retraction of a fibrin clot similar to the normal (+/+), carrier (+/−) controls, whereas the negative control (−/−) could not retract the clot. This result demonstrates that platelet function has improved (Fig. 2C).
A buccal mucosa bleeding time assay was performed to determine whether the current level of gene transduction efficiency (10% of the circulating blood platelets expressing 6% of normal αIIbβ3 receptor levels) was sufficient to repair a vascular injury in vivo (Fig. 3A). Identical to human GT patients, the bleeding analysis for GT (−/−) control dogs showed that all of the animals (regardless of age) experienced prolonged bleeding times requiring intervention at the experimental endpoint of 20 min (Fig. 3A, Top). Similar to humans, GT dogs tended to lose a smaller quantity of blood (albeit more than the normal level) as they matured. Fig. 3A shows that GT dogs aged <2.5 y lost ≈2.5 g of blood, whereas GT dogs aged >2.5 y lost ≈1 g of blood in 20 minutes (Fig. 3A, Top). Remarkably, there was a significant reduction in the amount of blood lost (≤135-fold decrease) and a notable improvement in bleeding times beginning at 6 mo after PBC transplant for dogs A, B, and C that was consistent with the presence of platelets expressing αIIbβ3. Specifically, dog A had a reduced bleeding time of 9 min (Fig. 3A, Top), dog B improved to an average of 7 min (Fig. 3A, Middle), and the bleeding time for dog C decreased to an average of 5 min (Fig. 3A, Bottom). Interestingly, this result coincided with the observation of significantly less bruising on the legs of dogs A, B, and C after transplant of αIIb-transduced PBC, demonstrating an obvious sign that hemostatic function had improved in these animals (Fig. 3B). Collectively, our results demonstrate that hematopoietic PBC gene transfer had corrected the GT phenotype.
Fig. 3.

Improved hemostatic function leads to correction of the GT phenotype. (A) BMBT assay was performed to measure the amount of blood lost onto a piece of filter paper every 15 s after making a standardized incision on the upper lip of each dog and recording the duration of the bleed up to the experimental endpoint of 20 min. Normal αIIb(+/+) dogs bleed for ≈3 min and lose a very small amount of blood with this assay, as previously reported (36). Identical to human GT patients, each αIIb(−/−) dog experienced prolonged bleeding until the experimental endpoint of 20 min (Top Left). The young (≤2 y old) GT dogs usually lost ≈2.5 g of blood, whereas the adult dogs (2.5–5.0 y old) tended to lose less blood (≈1.0 g). Remarkably, transplant dogs A (Top Right), B (Middle), and C (Bottom) showed a marked reduction in their bleeding times down to 9, 5, and 4 min, respectively. Likewise, there was a 20-fold (A), 100-fold (B), and 135-fold (C) reduction in blood lost, which correlated with the presence of platelets expressing αIIbβ3. This result was observed for 2, 4, and 5 y after transplant, respectively. (B) Bruising (petechiae, arrow) on the legs decreased significantly for each transplant recipient, as demonstrated by comparing pictures taken before and after transplant for dogs A (Top), B (Middle), and C (Bottom).
To determine whether the PBC transplant recipients developed a humoral antibody response to the newly expressed αIIb, normal αIIb(+/+) platelets were incubated with plasma from each transplant dog and the presence of platelet-bound antibody assessed by flow cytometry using fluorescent anti-canine Ig. Buffer without plasma was used as a negative control, and a monoclonal antibody to the integrin αIIbβ3 complex (7E3) served as a positive control. The histogram in Fig. S1A revealed that (1:10 diluted) plasma from dog B developed a relatively high-affinity antibody that reacted with normal αIIbβ3(+/+) canine and murine platelets. However, plasma from dog B did not react with αIIbβ3(−/−) platelets. As anticipated, dog B plasma also recognized normal αIIbβ3(+/+) human platelets compared with a buffer negative control. This result indicates that the antibody response was specific for an epitope on αIIb or the αIIbβ3 complex that is highly conserved across species. Dog A had a low-affinity antibody for normal canine platelets, whereas plasma from dog C did not react with canine platelets. Further analysis (Fig. S1B) revealed that Ig isolated from dog B plasma had the ability to inhibit aggregation of normal canine platelets in a concentration-dependent manner.
Intravenous Ig (IVIG) has been shown to be effective for treating humans who develop immune-mediated thrombocytopenia (20). Thus, IVIG was administered to dog B to investigate whether it could attenuate the acquired immune response to the αIIbβ3 complex and alleviate destruction of genetically altered platelets. Flow cytometric analysis showed an immediate decrease in the level of plasma Ig that reacted with normal canine platelets after two injections of IVIG (Fig. S1C, brown arrow). As a result, there was also a marked rise in the number of platelets expressing αIIbβ3. The antibody titer remained low and displayed negligible reactivity with platelets expressing αIIbβ3 for 2 y. A complication developed when dog B experienced a dramatic increase in white blood cell counts (Fig. S2A), severe thrombocytopenia (Fig. S2B), and life-threatening hemolytic anemia after administration of an annual vaccination (Figs. S1C and S2 A and B, magenta arrow). This result indicates that this adverse event was not caused by transplant of αIIb-transduced PBC, because destruction was not directed specifically toward transduced platelets. This conclusion is supported by the documentation of the occurrence of severe reactions after vaccination of dogs and humans (21–23). Surprisingly, we observed that administration of the vaccine also correlated directly with a significant increase in antibody titer to αIIbβ3 (Fig. S1C). This presents a potentially serious and previously unrecognized complication for gene transfer protocols that warrants further investigation. Fortunately, hemolytic anemia and thrombocytopenia, as well as the immune response to αIIbβ3, diminished when dog B was given an intermittent treatment regimen of immune-suppressive drugs consisting of prednisone, cyclosporine, and IVIG (Figs. S1C and S2 A and B, brown and green arrows).
The health of the each animal was monitored for several years to determine whether PBC gene transfer was a safe and viable approach for treating GT. Genomic DNA was isolated from peripheral blood leukocytes collected from dogs A, B, and C at 2, 4, and 5 y after transplant, respectively, to investigate whether the αIIb lentivirus caused insertional mutagenesis as has been reported for patients who received retrovirus gene transfer to correct X-linked severe combined immunodeficiency. The presence of the lentivirus vector was confirmed by successful PCR amplification of the woodchuck hepatitis virus postregulatory element (WPRE) from the DNA of dogs A, B, and C (Fig. S3A). This is consistent with detection of the transgene product on the surface of platelets. Analysis to detect specific sites of lentivirus insertion into the genome revealed that eight distinct insertions were identified for dog A, whereas the location of the lentivirus could not be mapped for dog B, and two insertion sites were found for dog C (Fig. S3B and Table S1). The analysis revealed that the lentivirus preferred to integrate in a negative orientation, usually into regions that do not encode genes. Dog A had three lentiviral insertions within early introns of identifiable genes, although none of the genes seem to be oncogenes or trigger neoplastic events. The result of the genomic analysis is consistent with the observation of overall good health for all of the dogs and evaluation of annual peripheral blood smears documenting normal morphology and numbers of circulating hematopoietic cells. Our results indicate that oncogenesis had not occurred in any of the animals at the experimental endpoint. There will always be the potential for insertional mutagenesis to occur because the recombinant lentivirus vector inserted randomly into the genome.
In summary, our results support the feasibility of targeting platelets with genetic therapies, which paves the way for better management of patients with inherited platelet bleeding defects and potentially a wide spectrum of disorders, given that platelets play a role in many cellular processes (24). Overall, this work showed that cytokine-mobilized PBC can be safely collected and undergo genetic manipulation and reinfusion into a large animal with an inherited bleeding disorder. This result led to the successful utilization of a lentivirus vector under transcriptional control of the integrin αIIb gene promoter, which corrected function in a subset of platelets at a level capable of improving hemostasis for at least 5 years. Similar to genetically modified platelets in GT canine transplant recipients, type 2 GT human patients express 5–20% residual levels of αIIbβ3 on the platelet surface. However, most of the reported human cases have also characterized molecular genetic defects within αIIbβ3 that negatively affect integrin structure/function, leading to prolonged bleeding (25). In contrast, we have observed that 10% of platelets from the canine GT transplant recipients could be modified to express a normal form of human αIIb that associated successfully with normal (endogenously expressed) canine β3 to produce a functional integrin receptor complex. This result, in addition to our previous studies showing improved hemostatic function of human GT tissue cultured megakaryocytes engineered to express reduced levels of αIIbβ3 (8), provides a possible explanation of why genetic modification of GT platelets to express a low level of normal αIIbβ3 led to improved hemostatic function within dogs. Of course it remains to be determined whether this level of normal αIIbβ3 would be sufficient to improve hemostasis within human GT patients. In the meantime, we have shown the feasibility of gene therapy for GT in a large animal. We also learned that in vivo drug selection is essential to correct uncontrolled bleeding. It is noteworthy that immune-mediated destruction of GT platelets expressing integrin αIIbβ3 was not detected in two of the dogs. However, an antibody response to the transgene product led to complications in one animal (especially after vaccine administration), which was diminished but not eliminated with transient immune-suppression therapy.
Materials and Methods
Dogs.
Characterization of the molecular defect within αIIb resulting in GT in the Great Pyrenees was previously described (10). Identical to patients, the diagnosis of GT was established by a prolonged bleeding time, failure of platelets to aggregate with physiological agonists of platelet activation, and failure to retract a fibrin clot. Canine platelets contained <5% detectable αIIbβ3, which is consistent with type 1 GT. A licensed, laboratory animal veterinarian routinely examined the health of the dogs. All protocols were approved by the Animal Care and Use Committees of the Medical College of Wisconsin and Auburn University.
Lentivirus Vector.
The αIIb-(M)WPT genetic transfer vector is derived from an HIV type 1 lentivirus backbone (D. Trono, University of Geneva, Switzerland) (26). A cDNA cassette was subcloned into the lentivirus backbone encoding a fragment of the integrin αIIb gene promoter from nucleotide −889 to +35, to direct megakaryocyte-specific transcription (9, 27) of a normal, full-length integrin αIIb subunit . The construct also contains a tissue-nonspecific gene promoter from the murine stem cell virus driving transcription of a drug-selectable DNA repair protein (P140K) MGMT (B. Sorrentino, St. Judes Research Center, Nashville, TN) (28). The correct cDNA cassette gene sequence was confirmed by nucleotide analysis. Recombinant lentivirus was generated from three-plasmid transient cotransfection, as described previously (9). Virus was collected, concentrated 500-fold, and stored at −80 °C until used as described (27). Virus titer was determined by RT-PCR as recently described (29). Replication-competent virions were confirmed absent from stocks with marker rescue assays (8).
Antibodies.
Phycoerythrin (PE)-conjugated Ab to β3 (CD61), FITC-conjugated Ab to αIIb (CD41), and isotype standards (PE-IgG, FITC-IgG) were from BD Biosciences. An FITC–anti-β3 and an isotype control was from Dako. An Ab “7E3” (17) recognizing αIIbβ3 and αvβ3 was a gift from B. Coller (The Rockefeller University, New York, NY). An Ab to canine αIIb “2F9” (30) was from S. Burstein (University of Oklahoma, Oklahoma City, OK). An Ab to canine CD34 “1H6” (13) was purchased from the Fred Hutchinson Cancer Research Center (Seattle, WA).
CD34+ PBC Isolation, Transduction, and Transplant.
Dogs were injected daily with canine recombinant G-CSF (10 μg/kg per day) and canine recombinant stem cell factor (5 μg/kg per day) (gift of Amgen, Thousand Oaks, CA). Cytokine-mobilized PBC collection was performed on the third day using a COBE Spectra Blood Cell Separator. Mononuclear PBC were isolated from the apheresis product with Ficoll-Paque Plus (GE Healthcare). CD34+ PBC were selected with a biotin-conjugated 1H6 Ab and anti-biotin immunomagnetic beads on an Automacs magnetic cell separator (Miltenyi Biotec).
CD34+ PBC were transduced with αIIb-(M)WPT lentivirus similar to a previously described protocol (31). Briefly, 4 × 106 cells per well in a sterile, six-well plate (Falcon-Becton Dickinson) coated with 20 μg/cm2 RetroNectin (Takara Shuzo) were incubated with 1 αIIb-(M)WPT lentivirion per cell in X-Vivo 10 with 20% FCS and rhIL-3, rcaIL-6, rcaSCF, rhTPO, and rhflk2/flt3 ligand. Approximately 2.5 × 106 αIIb-transduced CD34+ PBC/kg and 2.5 × 108 CD34(−) PBC were infused into the cephalic vein of each autologous transplant recipient preconditioned with a nonmyeloablative dose of 4 mg/kg busulfan or 100 cGy total body irradiation using a Pantac 320 orthovoltage X-ray machine, as previously described (5, 32).
In Vivo Enrichment of Lentivirus-Transduced CD34+ PBC.
Dogs were injected three times with 100 mg O6-benzylguanine (Sigma Aldrich) combined with 0.26, 0.41, or 0.45 mg/kg carmustine (Bristol-Myers Squibb) to promote in vivo enrichment of PBC transduced with a drug-resistant form (P140K) of MGMT, as previously described (18).
Blood Collection.
Blood was collected into a vacutube containing 7.5% EDTA anticoagulant as previously described (5). Washed platelets were isolated from blood that was layered onto Fico/Lite for Platelets (Atlanta Biologicals) and used directly for flow cytometry or platelet function analysis, as previously reported (9).
Flow Cytometry.
Immunocytometric analysis was used to determine the number of integrin receptors on the platelet surface and platelet transduction efficiency in the presence of saturating levels of Ab to αIIbβ3, as previously described (9).
Platelet Aggregation.
Aggregation of washed platelets was measured as described previously (9). Briefly, aggregation was initiated by adding 0.15 mg/mL of peak 1 soluble fibrinogen (Enzyme Research Laboratories) (33) and a platelet activation agonist mixture of 100 μM ADP, 20 μM epinephrine (BioData), and 250 μM of each canine TRAP (1, SFFLKN-NH2; 3, TRFGAP-NH2; 4, SFPGQP-NH2) (34) synthesized by our Protein Core Laboratory. Aggregation was monitored on a PAP-4 platelet aggregation profiler and analyzed with data interface software (BioData).
Platelet Spreading on Immobilized Fibrinogen.
Platelet-spreading assay was performed as previously described (35). Briefly, 5 × 106/mL of washed platelets were incubated at 37 °C for 45 min on eight-chamber glass tissue-culture slides (Becton Dickinson) coated with fibrinogen (100 μg/mL). Samples were mounted in Vectashield mounting medium (Vector Laboratories), and images were acquired with a Photometrics SenSys camera from three consecutive fields using a Zeiss Axioscop microscope with a Zeiss 100× oil-immersion lens (1.3 N.A.) and analyzed using Metamorph software (Universal Imaging).
Blood Clot Retraction.
Retraction assays were performed as previously described (5). Briefly, blood (0.5 mL) was drawn directly into a syringe containing 4.5 mL of cold isotonic saline and mixed gently. The mixture (2.0 mL) was added to glass tubes containing 0.1 mL of 10 U/mL human thrombin (Chrono-Log). Tubes were placed at 4 °C for 30 min and then transferred to a 37 °C water bath. Photos were taken with a Nikon DX camera at 3, 4, 5, 6, and 24 h after addition of thrombin. The ability of platelets to retract a clot was directly compared by visualization of the retraction of blood clots from αIIb(−/−,+/−,+/+) controls.
Buccal Mucosa Bleeding Time (BMBT).
The BMBT is defined as the time required for a stream of bleeding to end from a severed lip (1-mm incision) of a dog under sedation (with medetomidine or dexmeditomidine) as previously described (36). Briefly, a single incision was made in the lip of each dog with an automated incision device (Surgicutt; ITC), and blood was collected onto a sterile filter paper (Surgicutt) every 15 s until the bleeding stopped or reached the experimental endpoint of 20 min.
Supplementary Material
Acknowledgments
This work was supported by National Heart, Lung, and Blood Institute Grant R01 HL-68138 (to D.A.W.), American Heart Association Award (Northland Affiliate) 0160441Z (to D.A.W.), generous gifts from the Children's Hospital Foundation (to D.A.W.), the Midwest Athletes Against Childhood Cancer Fund (D.A.W.), John B. and Judith A. Gardetto (D.A.W.), the Glanzmann Research Foundation (D.A.W.), and National Institutes of Health Grant P40 RR024928 to the National Gene Vector Biorepository (T.B.H. and K.C.).
Footnotes
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1016394108/-/DCSupplemental.
References
- 1.Nurden P, Nurden AT. Congenital disorders associated with platelet dysfunctions. Thromb Haemost. 2008;99:253–263. doi: 10.1160/TH07-09-0568. [DOI] [PubMed] [Google Scholar]
- 2.Glanzmann E. Hereditare hamorrhagische thrombasthenie: Ein beitrag zur pathologie dr blut plattchen. J Kinderkr. 1918;88:113. [Google Scholar]
- 3.Coller BS, Shattil SJ. The GPIIb/IIIa (integrin alphaIIbbeta3) odyssey: A technology-driven saga of a receptor with twists, turns, and even a bend. Blood. 2008;112:3011–3025. doi: 10.1182/blood-2008-06-077891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Poon MC, Demers C, Jobin F, Wu JW. Recombinant factor VIIa is effective for bleeding and surgery in patients with Glanzmann thrombasthenia. Blood. 1999;94:3951–3953. [PubMed] [Google Scholar]
- 5.Niemeyer GP, et al. Correction of a large animal model of type I Glanzmann's thrombasthenia by nonmyeloablative bone marrow transplantation. Exp Hematol. 2003;31:1357–1362. doi: 10.1016/j.exphem.2003.09.005. [DOI] [PubMed] [Google Scholar]
- 6.Flood VH, et al. Sustained engraftment post bone marrow transplant despite anti-platelet antibodies in Glanzmann thrombasthenia. Pediatr Blood Cancer. 2005;45:971–975. doi: 10.1002/pbc.20365. [DOI] [PubMed] [Google Scholar]
- 7.Bauer TR, Jr, et al. Successful treatment of canine leukocyte adhesion deficiency by foamy virus vectors. Nat Med. 2008;14:93–97. doi: 10.1038/nm1695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wilcox DA, et al. Megakaryocyte-targeted synthesis of the integrin beta(3)-subunit results in the phenotypic correction of Glanzmann thrombasthenia. Blood. 2000;95:3645–3651. [PubMed] [Google Scholar]
- 9.Fang J, et al. Therapeutic expression of the platelet-specific integrin, alphaIIbbeta3, in a murine model for Glanzmann thrombasthenia. Blood. 2005;106:2671–2679. doi: 10.1182/blood-2004-12-4619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lipscomb DL, Bourne C, Boudreaux MK. Two genetic defects in alphaIIb are associated with type I Glanzmann's thrombasthenia in a Great Pyrenees dog: A 14-base insertion in exon 13 and a splicing defect of intron 13. Vet Pathol. 2000;37:581–588. doi: 10.1354/vp.37-6-581. [DOI] [PubMed] [Google Scholar]
- 11.Prandini MH, Martin F, Thevenon D, Uzan G. The tissue-specific transcriptional regulation of the megakaryocytic glycoprotein IIb gene is controlled by interactions between a repressor and positive cis-acting elements. Blood. 1996;88:2062–2070. [PubMed] [Google Scholar]
- 12.Beard BC, et al. Efficient and stable MGMT-mediated selection of long-term repopulating stem cells in nonhuman primates. J Clin Invest. 2010;120:2345–2354. doi: 10.1172/JCI40767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.McSweeney PA, et al. Characterization of monoclonal antibodies that recognize canine CD34. Blood. 1998;91:1977–1986. [PubMed] [Google Scholar]
- 14.Enssle J, et al. Stable marking and transgene expression without progression to monoclonality in canine long-term hematopoietic repopulating cells transduced with lentiviral vectors. Hum Gene Ther. 2010;21:397–403. doi: 10.1089/hum.2009.076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Newman PJ, Seligsohn U, Lyman S, Coller BS. The molecular genetic basis of Glanzmann thrombasthenia in the Iraqi-Jewish and Arab populations in Israel. Proc Natl Acad Sci USA. 1991;88:3160–3164. doi: 10.1073/pnas.88.8.3160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lipscomb DL, Bourne C, Boudreaux MK. Nucleotide sequence of the canine alphaIIb gene from platelet-derived cDNA. Am J Vet Res. 2001;62:1486–1492. doi: 10.2460/ajvr.2001.62.1486. [DOI] [PubMed] [Google Scholar]
- 17.Artoni A, et al. Integrin beta3 regions controlling binding of murine mAb 7E3: Implications for the mechanism of integrin alphaIIbbeta3 activation. Proc Natl Acad Sci USA. 2004;101:13114–13120. doi: 10.1073/pnas.0404201101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Neff T, et al. Methylguanine methyltransferase-mediated in vivo selection and chemoprotection of allogeneic stem cells in a large-animal model. J Clin Invest. 2003;112:1581–1588. doi: 10.1172/JCI18782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.George JN, Caen JP, Nurden AT. Glanzmann's thrombasthenia: The spectrum of clinical disease. Blood. 1990;75:1383–1395. [PubMed] [Google Scholar]
- 20.Newman PJ, Valentin N. Human platelet alloantigens: Recent findings, new perspectives. Thromb Haemost. 1995;74:234–239. [PubMed] [Google Scholar]
- 21.Jackson ML, Kruth SA. Immune-mediated hemolytic anemia and thrombocytopenia in the dog: A retrospective study of 55 cases diagnosed from 1979 through 1983 at the Western College of Veterinary Medicine. Can Vet J. 1985;26:245–250. [PMC free article] [PubMed] [Google Scholar]
- 22.Duval D, Giger U. Vaccine-associated immune-mediated hemolytic anemia in the dog. J Vet Intern Med. 1996;10:290–295. doi: 10.1111/j.1939-1676.1996.tb02064.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cines DB, Bussel JB, Liebman HA, Luning Prak ET. The ITP syndrome: Pathogenic and clinical diversity. Blood. 2009;113:6511–6521. doi: 10.1182/blood-2009-01-129155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Leslie M. Cell biology. Beyond clotting: The powers of platelets. Science. 2010;328:562–564. doi: 10.1126/science.328.5978.562. [DOI] [PubMed] [Google Scholar]
- 25.Wilcox DA, Paddock CM, Lyman S, Gill JC, Newman PJ. Glanzmann thrombasthenia resulting from a single amino acid substitution between the second and third calcium-binding domains of GPIIb. Role of the GPIIb amino terminus in integrin subunit association. J Clin Invest. 1995;95:1553–1560. doi: 10.1172/JCI117828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wiznerowicz M, Trono D. Conditional suppression of cellular genes: Lentivirus vector-mediated drug-inducible RNA interference. J Virol. 2003;77:8957–8961. doi: 10.1128/JVI.77.16.8957-8961.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wilcox DA, Olsen JC, Ishizawa L, Griffith M, White GC., 2nd Integrin alphaIIb promoter-targeted expression of gene products in megakaryocytes derived from retrovirus-transduced human hematopoietic cells. Proc Natl Acad Sci USA. 1999;96:9654–9659. doi: 10.1073/pnas.96.17.9654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Persons DA, et al. Successful treatment of murine beta-thalassemia using in vivo selection of genetically modified, drug-resistant hematopoietic stem cells. Blood. 2003;102:506–513. doi: 10.1182/blood-2003-03-0677. [DOI] [PubMed] [Google Scholar]
- 29.Lizée G, et al. Real-time quantitative reverse transcriptase-polymerase chain reaction as a method for determining lentiviral vector titers and measuring transgene expression. Hum Gene Ther. 2003;14:497–507. doi: 10.1089/104303403764539387. [DOI] [PubMed] [Google Scholar]
- 30.Burstein SA, Friese P, Downs T, Epstein RE. Canine megakaryocytopoiesis: Analysis utilizing a monoclonal antibody to a 140-kd dog platelet protein. Exp Hematol. 1991;19:47–52. [PubMed] [Google Scholar]
- 31.Horn PA, et al. Efficient lentiviral gene transfer to canine repopulating cells using an overnight transduction protocol. Blood. 2004;103:3710–3716. doi: 10.1182/blood-2003-07-2414. [DOI] [PubMed] [Google Scholar]
- 32.Sokolic RA, et al. Nonmyeloablative conditioning with busulfan before matched littermate bone marrow transplantation results in reversal of the disease phenotype in canine leukocyte adhesion deficiency. Biol Blood Marrow Transplant. 2005;11:755–763. doi: 10.1016/j.bbmt.2005.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mosesson MW, Finlayson JS. Biochemical and chromatographic studies of certain activities associated with human fibrinogen preparations. J Clin Invest. 1963;42:747–755. doi: 10.1172/JCI104767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Boudreaux MK, Catalfamo JL, Klok M, et al. Calcium diacylglycerol guanine nucleotide exchange factor I gene mutations associated with loss of function in canine platelets. Translational Research. 2007;150:81–92. doi: 10.1016/j.trsl.2007.03.006. [DOI] [PubMed] [Google Scholar]
- 35.Boylan B, et al. Identification of FcgammaRIIa as the ITAM-bearing receptor mediating alphaIIbbeta3 outside-in integrin signaling in human platelets. Blood. 2008;112:2780–2786. doi: 10.1182/blood-2008-02-142125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jergens AE, Turrentine MA, Kraus KH, Johnson GS. Buccal mucosa bleeding times of healthy dogs and of dogs in various pathologic states, including thrombocytopenia, uremia, and von Willebrand's disease. Am J Vet Res. 1987;48:1337–1342. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.