

Abstract
Aminoethylcysteine ketimine decarboxylated dimer is a natural sulfur-containing compound detected in human plasma and urine, in mammalian brain and in many common edible vegetables. Over the past decade many studies have been undertaken to identify its metabolic role. Attention has been focused on its antioxidant properties and on its reactivity against oxygen and nitrogen reactive species. These properties have been studied in different model systems starting from plasma lipoproteins to specific cellular lines. All these studies report that aminoethylcysteine ketimine decarboxylated dimer is able to interact both with reactive oxygen and nitrogen species (hydrogen peroxide, superoxide anion, hydroxyl radical, peroxynitrite and its derivatives). Its antioxidant activity is similar to that of Vitamin E while higher than other hydrophilic antioxidants, such as trolox and N-acetylcysteine.
Keywords: sulfur-containing antioxidants, aminoethylcysteine ketimine decarboxylated dimer, reactive oxygen species, reactive nitrogen species
1. Introduction
Oxygen is the key molecule which enables aerobic metabolism in living organisms. However, due to its high reactivity, it is also able to damage bio-molecules by producing reactive oxygen species (ROS) [1]. For this reason living organisms have developed a large and complex network of antioxidant molecules and enzymes, able to protect cellular components such as nucleic acids, proteins and lipids from oxidative damage.
According to a general definition, antioxidants can slow down or prevent the oxidation of other molecules by removing free radical intermediates. Cellular antioxidants can mainly act in two ways: (i) preventing these reactive species from being formed, or (ii) inactivating them before they are able to damage cellular components [2–5]. ROS production occurs physiologically during aerobic metabolism and the main role of the antioxidant network present within the cell is to buffer their overproduction, by keeping them at a level where their physiological role can be carried out (i.e., redox signaling) [6]. An imbalance of the antioxidant system may cause severe cellular damage and lead to oxidative stress condition, which is often involved in the pathogenesis of important diseases, such as cancer and atherosclerosis. This imbalance is also implicated in other pathological conditions (such as malaria and rheumatoid arthritis) and could play a role in neurodegenerative diseases and ageing processes [7].
In the last decades, research has been focused on prevention of oxidative damage. The molecular mechanisms involved in the radical scavenging activity of many natural antioxidants and the role they have in human health have been extensively studied. A lot of attention has been focused on dietary antioxidants (α-tocopherol, β-carotene, ascorbic acid), as they may act together with endogenous antioxidant metabolites and enzymes (superoxide dismutase, glutathione peroxidase, catalase) in reducing ROS level. Several epidemiological studies have already highlighted that a high intake of plant products rich in antioxidants is associated with a reduced risk of a number of severe chronic diseases, such as atherosclerosis and cancer. The protection that fruits and vegetables provide against several diseases has been attributed to various antioxidants present in these species, such as vitamin C, vitamin E, α-tocopherol, β-carotene and polyphenolic compounds [8–11].
Related studies have also shown that many of these antioxidant compounds show anti-inflammatory, anti-atherosclerotic, antimutagenic, anticarcinogenic, antibacterial, or antiviral activities to a greater or lesser extent [12–15].
In recent years the work of our research group has been focused on a small group of natural sulfur-containing iminoacids deriving from l-cystathionine, l-lanthionine and (S)-2-aminoethyl-l-cysteine [16]. All of them have been identified in biological tissues and fluids but, until now, their biological function is still unclear [17–24]. Among these compounds, aminoethylcysteine ketimine decarboxylated dimer was also detected in many edible vegetables [25] and, while a physiological role has not yet been described for this molecule, its antioxidant properties have been extensively explored and are described in this review.
2. Sulfur-Containing Antioxidants
Sulfur plays a critical role in biology, it is widely distributed among living organisms and, particularly, is found in many peptides, proteins and low molecular weight compounds [26,27]. Among the sulfur species found in plants, bacteria, fungi and animals, there are many agents with unique chemical and biochemical properties, which are linked to redox processes, metal binding and catalytic reactions. The antibiotic and anticarcinogenic properties of sulfur compounds such as thiols, thiosulfinates, thiosulfonates, isothiocyanates, sulfoxides, sulfones, sulfinates and polysulfides make them interesting from a pharmacological perspective [28]. The full impact of sulfur in biological systems becomes evident when one considers the diversity of sulfur species and the reactions in which sulfur is involved. These reactions are the result of: (i) the range of different oxidation states sulfur can occupy in vivo; (ii) the abundance of different chemotypes possible for each oxidation state; (iii) the various chemical properties that each chemotype exhibits in addition to redox activity, i.e., electrostatic interactions, hydrogen bonding, acid-base properties, nucleophilicity and electrophilicity, metal-binding and catalysis. Among these properties, redox-activity, metal binding and nucleophilic substitution are the mechanism most frequently employed by cells to remove oxidative stressors, adventitious metal ions and toxic substances [26,27].
The best investigated sulfur-containing antioxidant compounds are, cysteine, methionine, taurine, glutathione, lipoic acid, mercaptopropionylglycine and N-acetylcysteine [29].
In recent years, increasing attention has been focused on the discovery of new physiological sulfur compounds that show antioxidant activities. Among these, a very interesting one is aminoethylcysteine ketimine decarboxylated dimer (Figure 1), which is a member of a group of eterocyclic ketimines containing sulfur and nitrogen arising from the metabolism of l-cystathionine, l-lanthionine and (S)-2-aminoethyl-l-cysteine [16]. Many of these ketimines and their derivatives have been found in detectable amounts in mammalian tissues and fluids [17–24].
Figure 1.
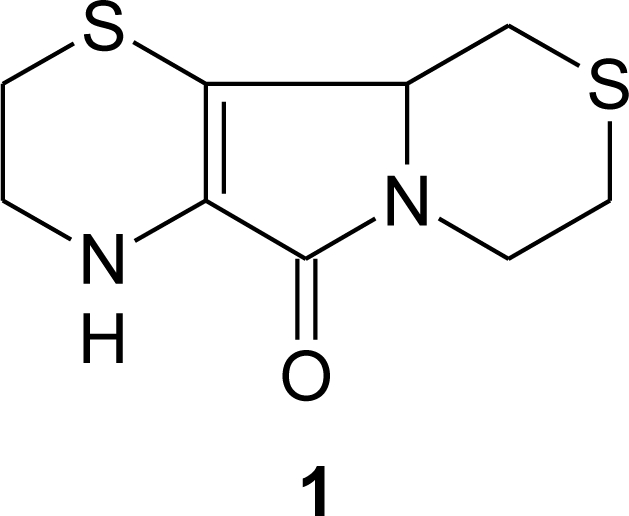
Aminoethylcysteine ketimine decarboxylated dimer.
3. Aminoethylcysteine Ketimine Decarboxylated Dimer
The chemical synthesis of 1 was first achieved by Hermann and coworkers in 1961 in very good yields starting from cysteamine and bromopyruvic acid [30].
Over the past several years, this molecule has been detected in many mammalian tissues and fluids (human plasma, 2–3 μM, [22], human urine [23] and in bovine cerebellum, 0.6–1 nmol/g wet weight [24]). Recently it was also identified in the brain of cysteamine-treated rats [31]. These findings seem particularly important because they allow formulating new hypotheses on the metabolic origin of this molecule. According to Pinto and coworkers [31], 1 cannot be detected in the brain of rats that are not fed a cysteamine-supplemented diet, suggesting that the brain has the capacity to synthesize 1 when supplied with an adequate amount of this aminothiol.
In fact, although a metabolic route leading to in vivo formation of 1 has not been identified yet, one of the current hypotheses is that it can be formed in vivo from the dimerization of aminoethylcysteine ketimine (2), followed by a spontaneous decarboxylation step [20] (Scheme 1).
Scheme 1.

Hypothetical pathway leading to the in vivo synthesis of 1.
Compound 2 can be endogenously synthesized starting from cysteamine and serine in a two-steps biosynthetic path involving cystathionine β-synthase and glutamine transaminase [32].
As 1 has been detected in many common Mediterranean edible vegetables (where it is present at a concentration range of nmol/g plant material), it can be alternatively considered a vitamin-like compound being introduced in the diet.
Besides efforts to detect this molecule in biological samples, over the past decade many studies have been undertaken to identify its metabolic role. Attention has been focused on its antioxidant properties and on its reactivity against oxygen and nitrogen reactive species. These properties have been studied in different model systems starting from plasma lipoproteins to specific cellular lines.
All these studies report that 1 is able to interact with reactive oxygen and nitrogen species (hydrogen peroxide, superoxide anion, hydroxyl radical, peroxynitrite and its derivatives) [33–36] in all the tested models. In addition, in order to understand the molecular mechanism underlying its antioxidant properties, research efforts have been focused on the oxidation products of 1. Until now three different products have been isolated and characterized [37–39], while several others need further analyses to be identified. As expected, sulfur plays a key role in the antioxidant properties of 1. Each sulfur atom can be oxidized to sulfoxide and subsequently to sulfone, leading to the formation of several oxidized species. In addition, a dimeric form and a dehydrogenated product of 1 were also detected, suggesting that the molecular architecture is as important as the presence of sulfur groups.
3.1. Aminoethylcysteine Ketimine Decarboxylated Dimer as Scavenger of Reactive Oxygen Species
Various processes inside the cell produce reactive oxygen species (ROS). Some of the most common ROS are hydrogen peroxide (H2O2), superoxide ion (O2−), hydroxide and hydroxyl ions (OH− and OH- respectively). Compound 1 is able to quench ROS leading to the formation of different oxidative products. In Scheme 2 two oxidation products of 1, which have been isolated and characterized so far, are shown.
Scheme 2.

Oxidation products of aminoethylcysteine ketimine decarboxylated dimer. Reagents and conditions: (i) CuCl2/t-BOOH or 2,2′-azo-bis-2-amidinopropane-HCl, (ii) hydrogen peroxide.
Compound 4, which shows an additional unsaturation with respect to 1, may arise from an oxidative dehydrogenation path that involves the carbons 10 and 10a of the tricyclic structure. This molecule is formed in vitro in the presence of CuCl2/tert-butyl hydroperoxide (t-BOOH) or 2,2′-azo-bis-2-amidinopropane hydrochloride [37]. Compound 5 is produced when 1 reacts with hydrogen peroxide [38].
Several other species have been identified by GC-MS but not yet unambiguously characterized. Among these, the chromatographic analysis shows the presence of at least four different oxidation products, whose mass and fragmentation patterns suggest the formation of 1-sulfone, 4-sulfoxide, 4-sulfone and hydroxyl-1, on the basis of that reported by Pecci et al. [38].
As already mentioned, endogenous ROS are known to play an important role in ethiopathogenesis of some common and severe chronic diseases, such as atherosclerosis.
Besides the in vitro studies carried out to test the quenching activity of 1 towards ROS, ex vivo assays on isolated human low-density lipoproteins (LDL) were performed. It is known that oxidation of LDL is an important event implicated in the pathogenesis of atherosclerosis [40]. Oxidized LDL that are no longer recognized by the LDL receptor of the macrophages, are internalized in these cells, via the scavenger receptor pathway, leading to the formation of foam cells [41,42]. The resistance of LDL to oxidative modification is linked to its fatty acids composition and to circulating levels of antioxidant compounds [43].
Compound 1 has already been found to be associated to plasma lipoproteins in humans [22], while another study demonstrated its efficacy in inhibiting the copper-catalyzed oxidation of human LDL [44]. Cu2+ catalyzes the lipoprotein oxidation in two main ways: (i) at the protein moiety level [45]; (ii) at the lipid moiety level, via decomposion of pre-existing lipid hydroperoxides and generation of peroxyl- and alkoxyl radicals, which initiate lipid peroxidation [46–48]. Since 1 shows scavenging activity against peroxyl (or alkoxyl) radicals, the main propagating species involved in the LDL lipoperoxidation, it seems reasonable to hypothesize that this could be the possible molecular protection mechanism carried out by 1 against LDL oxidation. The biological relevance of this study is that AECK-DD is active at concentrations comparable to those found in human plasma [22]. For this reason AECK-DD antioxidant properties were also tested on a human U937 monocytic cell line in the presence of the oxidant t-BOOH [49].
In fact, it has been recognized that monocyte-endothelium adhesion is a crucial early event in atherogenesis and that plasma antioxidants can prevent or reduce it [50,51]. U937 is a well characterized cell line and the response of this cell line to various inflammatory agents has been well documented [52,53].
The antioxidant activity of 1 is carried out within the U937 cells, as its uptake has been demonstrated by HPLC-ECD and GC-MS analyses [49,54]. A 24 h treatment with 50 and 250 μM AECK-DD, resulted in the incorporation of 0.10 ± 0.01 and 0.47 ± 0.08 ng AECK-DD × 10 [6] cells respectively [49]. In addition, 1 did not display any cytotoxic effect up to the range of mM concentration in the culture medium. Further, at this concentration level, no pro-apoptotic effect has been observed by DNA fragmentation measurements [49]. Compound 1 in concentration range 4–100 μM protects U937 cells from oxidative injury, as revealed by the higher viability maintained with respect to control cells during t-BOOH treatment. It is reported that low concentrations of t-BOOH lethally affect cultured cells by a mechanism that is dependent on the cellular lipids peroxidation [55]. Therefore, an association of 1 to cellular membranes due to its hydrophobicity could explain its high efficiency in protecting cells against t-BOOH-induced oxidative injury. The antioxidant activity of 1 was compared with other known antioxidants with similar results to that of vitamin E, and higher than other hydrophilic antioxidants tested (trolox and N-acetylcysteine). The results obtained by Macone and coworkers [49] indicated that the ability of 1 to protect human monocytic U937 cells from t-BOOH-induced oxidative stress seems to be mediated by its ability to maintain both intracellular glutathione levels and a reducing environment inside the cell, and to slow down the onset of lipid peroxidation.
The protective effect of 1 might be due to a direct quenching of free radicals, produced during t-BOOH treatment, by 1 itself. Moreover, data indicate that 1 is able to significantly reduce the intracellular level of pro-oxidant species in U937 cells in basal conditions. Such studies demonstrated for the first time an antioxidant action of 1 inside the cells, and its ability to modulate cellular response to oxidative stress. As already mentioned, the concentration of 1 in which its antioxidant activity is carried out, is in the range of that measured in human plasma from healthy subjects in fasting conditions [22]. In addition, due to the presence of 1 in human diet, the physiological concentrations of this molecule in non-fasting conditions may be expected to be even higher than those measured in fasting humans. Therefore it can be suggested that, at the concentrations present in human plasma, 1 can really play a significant role in the modulation of oxidative processes in vivo.
3.2. Aminoethylcysteine Ketimine Decarboxylated Dimer as Scavenger of Reactive Nitrogen Species
Reactive nitrogen species (RNS) are a family of molecules derived from nitric oxide (•NO) [56]. In particular, the product of the diffusion-controlled reaction between nitric oxide (•NO) and superoxide anion (O2−) is peroxynitrite (ONOO−/ONOOH), a strong oxidizing and nitrating agent that reacts with several biomolecules [57–63]. Besides its activity as oxidative process modulator, a scavenging activity on peroxynitrite has been described for 1 [36].
Peroxynitrite is an endogenous mediator of various forms of tissue damage in several human pathologies, including neurodegenerative diseases, atherosclerosis, inflammatory and autoimmune diseases. [64,65]. Peroxynitrite is known to mediate oxidation of suitable substrates, either through a direct two-electron mechanism or through an indirect one-electron reaction involving hydroxyl (•OH) and nitrogen dioxide (•NO2) radicals released during peroxynitrite homolysis [59,66–68]. Under physiological conditions, peroxynitrite predominantly reacts with carbon dioxide [69] and the oxidative reactions of peroxynitrite are mediated by (i) the carbonate radical anion (CO3•−); (ii) •NO2 generated by decomposition of the short-lived peroxynitrite-CO2 adduct [70–72]. After reaction with peroxynitrite, 1 undergoes oxidative modifications, yielding a dimeric form of the parent compound (6) (Scheme 3), which has so far been isolated and characterized using 1D and 2D nuclear magnetic resonanceand ion trap mass spectrometry [39]. The proposed mechanism for the formation of the peroxynitrite-oxidation derivative of 1 involves the radical dimerization of 1 [39].
Scheme 3.

Production of a dimeric form of aminoethylcysteine ketimine decarboxylated dimer in the presence of peroxynitrite. Reagents and conditions: (i) peroxynitrite.
Peroxynitrite induces lipid peroxidation [73], oxidizes protein and non-protein thiol groups [61,74] and reacts with tyrosine to yield 3-nitrotyrosine [62]. The occurrence of 3-nitrotyrosine is considered the molecular footprint left by reaction of RNS with biomolecules [75,76], while nitrated tyrosine residues are actually considered as biomarkers in a variety of pathophysiological conditions. In addition, peroxynitrite can oxidize free methionine [77] and methionine residues in proteins, e.g., in the α1-antiproteinase (α1AP), where the oxidation of a critical methionine residue destroys the α1AP activity [78]. Peroxynitrite also plays a role in the oxidation of low density lipoprotein [79]. Its generation in vivo leads to oxidative modification of blood lipoprotein, which is thought to be a critical event in the development of cardiovascular diseases, including atherosclerosis [40–43].
Tyrosine, when exposed to peroxynitrite at neutral pH, undergoes nitration to form 3-nitrotyrosine. Compound 1 decreases the peroxynitrite-mediated tyrosine nitration in a concentration-dependent manner. In the μM range concentration of 1, reduction of nitration was higher than 50%. At higher (100 μM) concentration, the nitration is even completely prevented. Compared with other sulfur compounds with established antioxidant activity, 1 showed a protective effect similar to that of glutathioneand N-acetylcysteine, but higher than that of methionine [36].
As reported above, peroxynitrite can oxidize methionine residues in proteins. Treatment of α1AP with peroxynitrite causes a strong reduction in its elastase inhibitory capacity, as a consequence of the oxidation of a critical methionine [78]. Compound 1 was able to carry out complete protection against α1AP inactivation, even at concentrations much lower than those of peroxynitrite. Compound 1 did not show any protective effect if added to α1AP preincubated with peroxynitrite, indicating that 1 did not reverse but prevent α1AP inactivation by scavenging peroxynitrite and/or its decomposition products [36].
It has been previously reported that peroxynitrite is able to oxidatively modify LDL [79]. LDL preincubation with 1 before peroxynitrite addition clearly demonstrated the effectiveness of 1 in decreasing peroxynitrite-mediated LDL oxidation. Compound 1 was able to prevent the oxidation of LDL almost completely. [36].
These results showed that 1 is a powerful scavenger of peroxynitrite and/or its derived species as it could efficiently protect tyrosine against nitration, α1AP against inactivation and LDL against modification. At present these data do not give indications about the mechanism of protection. Compound 1 could act by directly scavenging peroxynitrite or via a combination with reactive intermediates of peroxynitrite decomposition.
4. Conclusions
Over the past decade many studies have been undertaken to identify the metabolic role of aminoethylcysteine ketimine decarboxylate dimer, a natural sulfur-containing compound detected in human plasma and urine, mammalian brain and in many common edible vegetables. Attention has been focused on its antioxidant properties and on its reactivity against oxygen and nitrogen reactive species. All the reviewed studies report that aminoethylcysteine ketimine decarboxylate dimer is able to interact both with reactive oxygen and nitrogen species (hydrogen peroxide, superoxide anion, hydroxyl radical, peroxynitrite and its derivatives) protecting human low-density lipoprotein and cultured monocytes against oxidative injury. In these model systems, its antioxidant activity produced similar results to that of Vitamin E and higher than other hydrophilic antioxidant, such as trolox and N-acetylcysteine. In addition, 1 is able to quench ROS and RNS leading to the formation of different oxidative products that have been isolated and characterized. The study of several other oxidation products will lead to a better understanding of the molecular mechanism that underlies its powerful action as an antioxidant.
Acknowledgments
Financial support from FIRB, grant no. RBPR05NWWC_006, Sapienza University of Rome, Italy (Funds for selected research topics 2008–2010) and Fondazione Roma (Rome, Italy) are gratefully acknowledged. This work was partially supported by the Istituto Pasteur–Fondazione Cenci Bolognetti, Sapienza University of Roma, Italy.
References
- 1.Davies K. Oxidative stress: The paradox of aerobic life. Biochem. Soc. Symp. 1995;61:1–31. doi: 10.1042/bss0610001. [DOI] [PubMed] [Google Scholar]
- 2.Sies H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997;82:291–295. doi: 10.1113/expphysiol.1997.sp004024. [DOI] [PubMed] [Google Scholar]
- 3.Bjelakovic G, Nikolova D, Gluud LL, Simonetti RG, Gluud C. Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: Systematic review and meta-analysis. JAMA. 2007;297:842–857. doi: 10.1001/jama.297.8.842. [DOI] [PubMed] [Google Scholar]
- 4.Matill HA. Antioxidants. Ann. Rev. Biochem. 1947;16:177–192. doi: 10.1146/annurev.bi.16.070147.001141. [DOI] [PubMed] [Google Scholar]
- 5.Vertuani S, Angusti A, Manfredini S. The antioxidants and pro-antioxidants network: An overview. Curr. Pharm. Des. 2004;10:1677–1694. doi: 10.2174/1381612043384655. [DOI] [PubMed] [Google Scholar]
- 6.Miller RA, Britigan BE. Role of oxidant in microbial pathophysiology. Clin. Microbiol. Rev. 1997;10:1–18. doi: 10.1128/cmr.10.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Valko M, Leibfritz D, Moncol J, Cronin MTD, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007;39:44–84. doi: 10.1016/j.biocel.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 8.Gosslau A, Chen KY. Nutraceuticals, apoptosis, and disease prevention. Nutrition. 2004;20:95–102. doi: 10.1016/j.nut.2003.09.017. [DOI] [PubMed] [Google Scholar]
- 9.Law MR, Morris JK. By how much does fruit and vegetable consumption reduce the risk of ischaemic heart disease? Eur. J. Clin. Nutr. 1998;52:549–556. doi: 10.1038/sj.ejcn.1600603. [DOI] [PubMed] [Google Scholar]
- 10.Gundgaard J, Nielsen JN, Olsen J, Sørensen J. Increased intake of fruit and vegetables: Estimation of impact in terms of life expectancy and healthcare costs. Public Health Nutr. 2003;6:25–30. doi: 10.1079/PHN2002355. [DOI] [PubMed] [Google Scholar]
- 11.Brigelius-Flohé R, Traber MG. Vitamin E: Function and metabolism. FASEB J. 1999;13:1145–1155. [PubMed] [Google Scholar]
- 12.Migliore L, Coppedè F. Environmental-induced oxidative stress in neurodegenerative disorders and aging. Mutat. Res. 2009;674:73–84. doi: 10.1016/j.mrgentox.2008.09.013. [DOI] [PubMed] [Google Scholar]
- 13.Mitscher LA, Telikepalli H, McGhee E, Shankel DM. Natural antimutagenic agents. Mutat. Res. 1996;350:142–143. doi: 10.1016/0027-5107(95)00099-2. [DOI] [PubMed] [Google Scholar]
- 14.Owen RW, Giacosa A, Hull WE, Haubner R, Spiegelhalder B, Bartsch H. The antioxidant/anticancer potential of phenolic compounds isolated from olive oil. Eur. J. Canc. 2000;36:1235–1247. doi: 10.1016/s0959-8049(00)00103-9. [DOI] [PubMed] [Google Scholar]
- 15.Sala A, Recio MD, Giner RM, Manez S, Tournier H, Schinella G, Rios JL. Anti-inflammatory and antioxidant properties of Helichrysum italicum. J. Pharm. Pharmacol. 2002;54:365–371. doi: 10.1211/0022357021778600. [DOI] [PubMed] [Google Scholar]
- 16.Cavallini D, Ricci G, Duprè S, Pecci L, Costa M, Matarese RM, Pensa B, Antonucci A, Solinas SP, Fontana M. Sulfur-containing cyclic ketimines and imino acids. A novel family of endogenous products in the search for a role. Eur. J. Biochem. 1991;202:217–23. doi: 10.1111/j.1432-1033.1991.tb16365.x. [DOI] [PubMed] [Google Scholar]
- 17.Nardini M, Matarese RM, Pecci L, Antonucci A, Ricci G, Cavallini D. Detection of 2H-1,4-thiazine-5,6-dihydro-3-carboxylic acid(aminoethylcysteine ketimine) in the bovine brain. Biochem. Biophys. Res. Commun. 1990;166:1251–1256. doi: 10.1016/0006-291x(90)91000-i. [DOI] [PubMed] [Google Scholar]
- 18.Ricci G, Vesci L, Nardini M, Arduini A, Storto S, Rosato N, Cavallini D. Detection of 2H-1,4-thiazine-5,6-dihydro-3,5-dicarboxylic acid (lanthionine ketimine) in the bovine brain by a fluorimetric assay. Biochim. Biophys. Acta. 1989;990:211–215. doi: 10.1016/s0304-4165(89)80036-4. [DOI] [PubMed] [Google Scholar]
- 19.Cavallini D, Pecci L, Matarese RM, Ricci G, Achilli M. Gas-chromatographic mass-spectrometric detection of 1,4-hexahydrothiazepi±ne-3,5-dicarboxylic acid (cyclothionine) in bovine brain. J. Biol. Chem. 1985;29:15577–15579. [PubMed] [Google Scholar]
- 20.Matarese RM, Pecci L, Ricci G, Nardini M, Antonucci A, Cavallini D. Hexahydro-1,4-thiazepine-3,5-dicarboxylic acid and thiomorpholine-3,5-dicarboxylic acid are present in normal human urine. Proc. Natl. Acad. Sci. USA. 1987;84:5111–5114. doi: 10.1073/pnas.84.15.5111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yu S, Sugahara K, Zhang J, Ageta T, Kodama H, Fontana M, Duprè S. Simultaneous determination of urinary cystathionine, lanthionine, (S)-(2-aminoethyl)-l-cysteine and their cyclic compounds using liquid chromatography-mass spectrometry with atmospheric pressure chemical ionization. J. Chrom. B. 1997;698:301–307. doi: 10.1016/s0378-4347(97)00295-8. [DOI] [PubMed] [Google Scholar]
- 22.Matarese RM, Macone A, Antonini R, Maggio A, Antonucci A. Identification of aminoethylcysteine ketimine decarboxylated dimer in human plasma. J. Chromatogr. B. 1999;732:137–144. doi: 10.1016/s0378-4347(99)00272-8. [DOI] [PubMed] [Google Scholar]
- 23.Matarese RM, Macone A, Maggio A, Cavallini D. Aminoethylcysteine ketimine decarboxylated dimer detected in normal human urine by gas-liquid chromatography, selected ion monitoring, and mass spectrometry. J. Chromatogr. B. 1996;683:269–272. doi: 10.1016/0378-4347(96)00105-3. [DOI] [PubMed] [Google Scholar]
- 24.Matarese RM, Macone A, Crescentini G, Duprè S, Cavallini D. Detection of a decarboxylated dimer of aminoethylcysteine ketimine in bovine cerebellum. Neurochem. Int. 1998;32:365–368. doi: 10.1016/s0197-0186(97)00094-6. [DOI] [PubMed] [Google Scholar]
- 25.Macone A, Nardini M, Antonucci A, Maggio A, Matarese RM. Identification of aminoethylcysteine ketimine decarboxylated dimer, a natural antioxidant, in dietary vegetables. J. Agric. Food Chem. 2002;50:2169–2172. doi: 10.1021/jf0113934. [DOI] [PubMed] [Google Scholar]
- 26.Jacob C, Giles GI, Giles NM, Sies H. Sulfur and selenium: The role of oxidation state in protein structure and function. Angew. Chem. Int. Ed. 2003;42:4742–4258. doi: 10.1002/anie.200300573. [DOI] [PubMed] [Google Scholar]
- 27.Giles NM, Watts AB, Giles GI, Fry FH, Littlechild JA, Jacob C. metal and redox modulation of cysteine protein function. Chem. Biol. 2003;10:677–693. doi: 10.1016/s1074-5521(03)00174-1. [DOI] [PubMed] [Google Scholar]
- 28.Jacob C. A scent of therapy: Pharmacological implications of natural products containing redox-active sulfur atoms. Nat. Prod. Rep. 2006;23:851–863. doi: 10.1039/b609523m. [DOI] [PubMed] [Google Scholar]
- 29.Atmaca G. Antioxidant effects of sulfur-containing amino acids. Yonsei Med. J. 2004;45:776–788. doi: 10.3349/ymj.2004.45.5.776. [DOI] [PubMed] [Google Scholar]
- 30.Hermann P. Zur reaktion von halogenbrenztrabensaure mit thiolaminen. Chem. Ber. 1961;94:442–445. [Google Scholar]
- 31.Pinto JT, Khomenko T, Szabo S, McLaren GD, Denton TT, Krasnikov BF, Jeitner TM, Cooper AJL. Measurement of sulfur-containing compounds involved in the metabolism and transport of cysteamine and cystamine. Regional differences in cerebral metabolism. J. Chromatogr. B. 2009;877:3434–3441. doi: 10.1016/j.jchromb.2009.05.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Greenberg DM. In: Metabolic Pathways. Greenberg DM, editor. Academic press; New York, NY, USA: 1975. pp. 505–520. [Google Scholar]
- 33.Pecci L, Montefoschi G, Antonucci A, Cavallini D. Antioxidant properties of the decarboxylated dimer of aminoethylcysteine ketimine. Physiol. Chem. Phys. Med. NMR. 1995;27:223–229. [PubMed] [Google Scholar]
- 34.Pecci L, Fontana M, Montefoschi G, Cavallini D. Aminoethylcysteine ketimine decarboxylated dimer protects submitochondrial particles from lipid peroxidation at a concentration not inhibitory of electron transport. Biochem. Biophys. Res. Commun. 1994;205:264–268. doi: 10.1006/bbrc.1994.2659. [DOI] [PubMed] [Google Scholar]
- 35.Antonucci A, Pecci L, Coccia R, Fontana M, Cavallini D. The oxidation of aminoethylcysteine ketimine dimer by oxygen reactive species. Amino Acids. 1994;7:83–88. doi: 10.1007/BF00808449. [DOI] [PubMed] [Google Scholar]
- 36.Fontana M, Pecci L, Macone A, Cavallini D. Antioxidant properties of the decarboxylated dimer of aminoethylcysteine ketimine: Assessment of its ability to scavenge peroxynitrite. Free Radic. Res. 1998;29:435–440. doi: 10.1080/10715769800300481. [DOI] [PubMed] [Google Scholar]
- 37.Macone A, Caiazzo A, Antonucci A, Fochi I, Nardini M, Duprè S, Matarese RM. Synthesis and characterization of a dehydrogenation product arising from the oxidation of aminoethylcysteine ketimine decarboxylated dimer. J. Nat. Prod. 2007;70:1046–1048. doi: 10.1021/np060641t. [DOI] [PubMed] [Google Scholar]
- 38.Pecci L, Antonucci A, Pinnen F, Cavallini D. Identification of an oxidation product of aminoethylcysteine ketimine dimer. Amino Acids. 2000;18:61–67. doi: 10.1007/s007260050005. [DOI] [PubMed] [Google Scholar]
- 39.Mannina L, Viel S, Duprè S, Pecci L, Fontana M, Pinnen F, Antonucci A, Segre L. Structural elucidation of the oxidation product of aminoethylcysteine ketimine decarboxylated dimer by peroxynitrite. Tetrahedron. 2004;60:4151–4157. [Google Scholar]
- 40.Steinberg D, Parthasarathy S, Carew TE, Khoo JC, Witztum JL. Beyond cholesterol. modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989;320:915–924. doi: 10.1056/NEJM198904063201407. [DOI] [PubMed] [Google Scholar]
- 41.Witztum JL, Steinberg D. Role of oxidized low-density lipoprotein in atherogenesis. J. Clin. Invest. 1991;88:1785–1792. doi: 10.1172/JCI115499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Steinbrecher UP, Zhang HF, Lougheed M. Role of oxidatively modified LDL in atherosclerosis. Free Radic. Biol. Med. 1990;9:155–168. doi: 10.1016/0891-5849(90)90119-4. [DOI] [PubMed] [Google Scholar]
- 43.Esterbauer H, Gebicki J, Puhl H, Jürgens G. The role of lipid peroxidation and antioxidants in oxidative modification of LDL. Free Radic. Biol. Med. 1992;13:341–390. doi: 10.1016/0891-5849(92)90181-f. [DOI] [PubMed] [Google Scholar]
- 44.Matarese RM, Macone A, Fontana M, Duprè S, Cavallini D. Antioxidant activity of aminoethylcysteine ketimine decarboxylated dimer on copper induced LDL oxidation. Biochem. Mol. Biol. Int. 1998;46:829–837. doi: 10.1080/15216549800204372. [DOI] [PubMed] [Google Scholar]
- 45.Giessauf A, Steiner E, Esterbauer H. Early destruction of tryptophan residues of apolipoprotein B is a vitamin E-independent process during copper-mediated oxidation of LDL. Biochim. Biophys. Acta. 1995;1256:221–232. doi: 10.1016/0005-2760(95)00024-7. [DOI] [PubMed] [Google Scholar]
- 46.Noguchi N, Gotoh N, Niki E. Dynamics of the oxidation of low density lipoprotein induced by free radicals. Biochim. Biophys. Acta. 1993;1168:348–357. [PubMed] [Google Scholar]
- 47.Halliwell B, Gutteridge JM. Role of free radicals and catalytic metal ions in human disease: An overview. Methods Enzymol. 1990;186:1–85. doi: 10.1016/0076-6879(90)86093-b. [DOI] [PubMed] [Google Scholar]
- 48.Smith C, Mitchinson MJ, Aruoma OI, Halliwell B. Stimulation of lipid peroxidation and hydroxyl-radical generation by the contents of human atherosclerotic lesions. Biochem. J. 1992;286:901–905. doi: 10.1042/bj2860901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Macone A, Matarese RM, Gentili V, Antonucci A, Duprè S, Nardini M. Effect of aminoethylcysteine ketimine decarboxylated dimer, a natural sulfur compound present in human plasma, on tert-butyl hydroperoxide-induced oxidative stress in human monocytic U937 cells. Free Radic. Res. 2004;38:705–714. doi: 10.1080/10715760410001705159. [DOI] [PubMed] [Google Scholar]
- 50.Devaraj S, Li D, Jialal I. The effects of alpha tocopherol supplementation on monocyte function. decreased lipid oxidation, interleukin 1 beta secretion, and monocyte adhesion to endothelium. J. Clin. Invest. 1996;98:756–763. doi: 10.1172/JCI118848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Weber C, Erl W. Modulation of vascular cell activation, function, and apoptosis: Role of antioxidants and nuclear factor-kappa B. Curr. Top. Cell. Regul. 2000;36:217–235. doi: 10.1016/s0070-2137(01)80010-8. [DOI] [PubMed] [Google Scholar]
- 52.Jarvis WD, Kolesnick RN, Fornari FA, Traylor RS, Gewirtz DA, Grant S. Induction of apoptotic DNA damage and cell death by activation of the sphingomyelin pathway. Proc. Natl. Acad. Sci. USA. 1994;91:73–77. doi: 10.1073/pnas.91.1.73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Quillet-Mary A, Jaffrézou JP, Mansat V, Bordier C, Naval J, Laurent G. Implication of mitochondrial hydrogen peroxide generation in ceramide-induced apoptosis. J. Biol. Chem. 1997;272:21388–21395. doi: 10.1074/jbc.272.34.21388. [DOI] [PubMed] [Google Scholar]
- 54.Nardini M, Macone A, Matarese RM. Determination of aminoethylcysteine ketimine decarboxylated dimer in human plasma and cultured cells by high performance liquid chromatography with electrochemical detection. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003;795:319–327. doi: 10.1016/s1570-0232(03)00597-x. [DOI] [PubMed] [Google Scholar]
- 55.Masaki N, Kyle ME, Farber JL. Tert-butyl hydroperoxide kills cultured hepatocytes by peroxidizing membrane lipids. Arch. Biochem. Biophys. 1989;269:390–399. doi: 10.1016/0003-9861(89)90122-7. [DOI] [PubMed] [Google Scholar]
- 56.Radi R. Nitric oxide, oxidants and protein nitration. Proc. Natl. Acad. Sci. USA. 2004;101:4003–4008. doi: 10.1073/pnas.0307446101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Koppenol WH, Moreno JJ, Prior WA, Ischiropoulos H, Beckman JS. Peroxynitrite, a cloaked oxidant formed by nitric oxide and superoxide. Chem. Res. Toxicol. 1992;5:834–842. doi: 10.1021/tx00030a017. [DOI] [PubMed] [Google Scholar]
- 58.Huie RE, Padmaja S. The reaction of NO with superoxide. Free Radic. Res. Commun. 1993;18:195–199. doi: 10.3109/10715769309145868. [DOI] [PubMed] [Google Scholar]
- 59.Pryor WA, Squadrito GL. The chemistry of peroxynitrite: A product from the reaction of nitric oxide with superoxide. Am J Physiol. 1995;268:L699–L722. doi: 10.1152/ajplung.1995.268.5.L699. [DOI] [PubMed] [Google Scholar]
- 60.Szabo C, Ohshima H. DNA damage induced by peroxynitrite: Subsequent biological effects. Nitric Oxide. 1997;1:373–385. doi: 10.1006/niox.1997.0143. [DOI] [PubMed] [Google Scholar]
- 61.Trujillo M, Radi R. Peroxynitrite reaction with the reduced and the oxidized forms of lipoic acid: New insight into the reaction of peroxynitrite with thiols. Arch. Biochem. Biophys. 2002;397:91–98. doi: 10.1006/abbi.2001.2619. [DOI] [PubMed] [Google Scholar]
- 62.Alvarez B, Radi R. Peroxynitrite reactivity with amino acids and proteins. Amino Acids. 2003;25:295–311. doi: 10.1007/s00726-003-0018-8. [DOI] [PubMed] [Google Scholar]
- 63.Kurz CR, Kissner R, Nauser T, Perrin D, Koppenol WH. Rapid scavenging of peroxynitrous acid by monohydroascorbate. Free Radic. Biol. Med. 2003;35:1529–1537. doi: 10.1016/j.freeradbiomed.2003.08.012. [DOI] [PubMed] [Google Scholar]
- 64.Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol. Rev. 2007;87:315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Szabo C, Ischiropoulos H, Radi R. Peroxynitrite: Biochemistry, pathophysiology and development of therapeutics. Nat. Rev. Drug Discov. 2007;6:662–680. doi: 10.1038/nrd2222. [DOI] [PubMed] [Google Scholar]
- 66.Goldstein S, Czapski G. Direct and indirect oxidations by peroxynitrite. Inorg. Chem. 1995;34:4041–4048. doi: 10.1016/s0891-5849(96)00280-8. [DOI] [PubMed] [Google Scholar]
- 67.Richeson CE, Mulder P, Bowry VW, Ingold KU. The complex chemistry of peroxynitrite decomposition: New insights. J. Am. Chem. Soc. 1998;12:7211–7219. [Google Scholar]
- 68.Merenyi G, Lind J. Free radical formation in the peroxynitrous (ONOOH)/peroxynitrite (ONOO−) system. Chem. Res. Toxicol. 1998;11:243–246. doi: 10.1021/tx980026s. [DOI] [PubMed] [Google Scholar]
- 69.Lymar SV, Hurst JK. Rapid reaction between peroxynitrite ion and carbon dioxide: Implications for biological activity. J. Am. Chem. Soc. 1995;117:8867–8868. [Google Scholar]
- 70.Denicola A, Freeman BA, Trujillo M, Radi R. Peroxynitrite reaction with carbon dioxide/bicarbonate: Kinetics and influence on peroxynitrite-mediated oxidations. Arch. Biochem. Biophys. 1996;333:49–58. doi: 10.1006/abbi.1996.0363. [DOI] [PubMed] [Google Scholar]
- 71.Goldstein S, Czapski G. The effect of bicarbonate on oxidation by peroxynitrite: Implication for its biological activity. Inorg. Chem. 1997;36:5113–5117. [Google Scholar]
- 72.Augusto O, Bonini MG, Amanso AM, Linares E, Santos CCX, De Menezes SL. Nitrogen dioxide and carbonate radical anion: Two emerging radicals in biology. Free Radic. Biol. Med. 2002;32:841–859. doi: 10.1016/s0891-5849(02)00786-4. [DOI] [PubMed] [Google Scholar]
- 73.Radi R, Beckman JS, Bush KM, Freeman BA. Peroxynitrite-induced membrane lipid peroxidation: The cytotoxic potential of superoxide and nitric oxide. Arch. Biochem. Biophys. 1991;288:481–487. doi: 10.1016/0003-9861(91)90224-7. [DOI] [PubMed] [Google Scholar]
- 74.Radi R, Beckman JS, Bush KM, Freeman BA. Peroxynitrite oxidation of sulfhydryls. The cytotoxic potential of superoxide and nitric oxide. J. Biol. Chem. 1991;266:4244–4250. [PubMed] [Google Scholar]
- 75.Ischiropoulos H. Biological tyrosine nitration: A pathophysiological function of nitric oxide and reactive oxygen species. Arch. Biochem. Biophys. 1998;356:1–11. doi: 10.1006/abbi.1998.0755. [DOI] [PubMed] [Google Scholar]
- 76.Greenacre SA, Ischiropoulos H. Tyrosine nitration: Localization, quantification, consequences for protein function and signal transduction. Free Radic. Res. 2001;34:541–581. doi: 10.1080/10715760100300471. [DOI] [PubMed] [Google Scholar]
- 77.Pryor WA, Jin X, Squadrito GL. One- and two-electron oxidation of methionine by peroxynitrite. Proc. Natl. Acad. Sci. USA. 1994;91:11173–11177. doi: 10.1073/pnas.91.23.11173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Moreno JJ, Pryor WA. Inactivation of α1-antiproteinase inhibitor by peroxynitrite. Chem. Res. Toxicol. 1992;5:425–431. doi: 10.1021/tx00027a017. [DOI] [PubMed] [Google Scholar]
- 79.Graham A, Hogg N, Kalyanaraman B, O′Leary V, Darley-Usmar V, Moncada S. Peroxynitrite modification of low-density lipoprotein leads to recognition by the macrophage scavenger receptor. FEBS Lett. 1993;330:181–185. doi: 10.1016/0014-5793(93)80269-z. [DOI] [PubMed] [Google Scholar]