

Abstract
Here we report the identification of a new family of helminth neuropeptides with members in both nematodes and flatworms, and include preliminary cell biological and functional characterisation of one of the peptides from the trematode parasite of humans, Schistosoma mansoni. Bioinformatics and Rapid Amplification of cDNA Ends (RACE)-PCR were used to identify the complete S. mansoni neuropeptide precursor gene Sm-npp-1, which encodes three pentapeptides bearing the motif (A/G)FVR(I/L).NH2. Similar peptides were identified in three other flatworm species and in 15 nematode species. Quantitative PCR (qPCR) and immunocytochemical (ICC) analyses showed that Sm-npp-1 is constitutively expressed in larval and adult worms. ICC and confocal microscopy were employed to localise one of the schistosome NPP-1 peptides (GFVRIamide) in adult worms and schistosomules; antibodies labelled a pair of neurones in the cerebral ganglia that extend posteriorly along the main nerve cords. GFVRIamide displayed no detectable co-localisation with FMRFamide-like peptides (FLPs), nor was it detectable in muscle innervation. Exogenously applied peptide had a significant inhibitory effect on the mobility of whole adult worm pairs at 10−5 M (n=9). Finally, we explored Sm-npp-1 function in schistosomules using RNA interference (RNAi); we successfully achieved specific knockdown of the Sm-npp-1 transcript (54.46±10.41 % knockdown, n=3), but did not detect any clear, aberrant mobility or morphological phenotypes. NPP-1-like peptides are a new family of helminth peptides with a cell-specific expression pattern distinct from FLPs and a modulatory effect on schistosome muscular activity.
Keywords: Bioinformatics, Flatworm, Neurobiology, Physiology, RNAi
1. Introduction
Increased availability of genomic and transcriptomic resources have accelerated basic biological research on parasitic helminths. Neurobiology has benefitted from these advances which have expanded our knowledge of helminth neuropeptide diversity; more than 100 neuropeptides have been recently described from 10 flatworm species (McVeigh et al., 2009; Collins et al., 2010) where previously only a handful of neuropeptides had been isolated and characterized (Maule et al., 1991, 1993, 1994; Johnston et al., 1995, 1996). Nevertheless, the newly-described flatworm peptides (encoded on neuropeptide precursor, or npp genes) remain largely uncharacterised, such that their cell-specific expression patterns and biological functions are unknown. Here, we report a first step towards functional characterisation of peptides encoded by one of these novel genes, Sm-npp-1, in the trematode parasite of humans and causative agent of schistosomiasis, Schistosoma mansoni.
Flatworm npp genes encode at least 16 structurally-distinct neuropeptide families, including the L/M/Iamides, PWamides, SGFamides, D5amides, VVamides, amidated tripeptides and unamidated peptides, as well as FMRFamide-like peptide (FLP), neuropeptide F (NPF), cerebral peptide prohormone (CPL), gonadotropin-releasing hormone-like (GRH), insulin-like, myomodulin-like (MPL), pyrokinin-like (PPL), pedal peptide-like (PPP) and secreted peptide (SPP) prohormone families (McVeigh et al., 2009; Collins et al., 2010). FLPs are short, bioactive peptides with RFamide C-termini that are encoded on at least six genes (npp-3, -4, -11, -13, -19 and -23). The biology of FLPs has been reasonably well studied in several flatworm species, with physiology-based assays revealing excitatory effects on whole worms, muscle strips and dispersed muscle fibres. The potent activity of FLPs on dispersed schistosome muscle fibres indicates the occurrence of a muscle-based FLP receptor with appeal as a drug target candidate. Further, this receptor’s candidature as a drug target is enhanced by its promiscuity, in that it is also activated by FLPs of nematode and arthropod origin (Day et al., 1994, 1997; Graham et al., 1997, 2000; Marks et al., 1997; Moneypenny et al., 1997, 2001; Mousley et al., 2004). A FLP-responsive G-protein coupled receptor (GPCR) exists in the turbellarian, Girardia tigrina (Omar et al., 2007), but the site of expression, and thus potential role in muscle contraction, is unknown.
NPFs (36–39 amino-acid peptides with conserved [Y-17]-[Y-10]-GRPRFamide motifs) have been identified from seven species of flatworm (although NPF-immunoreactivity (NPF-IR) has been demonstrated in many more) (Maule et al., 1991; Day and Maule, 1999; Mair et al., 2000; Dougan et al., 2002; Humphries et al., 2004; McVeigh et al., 2009; Collins et al., 2010), and more than one NPF-encoding gene has been identified in three species. While the biological implications of the presence of multiple NPFs in a single species are not understood, we can point to several known effects: flatworm NPF-like peptides display a poorly characterised myoexcitatory effect (Marks et al., 1996; Hrckova et al., 2004), reduce forskolin-stimulated cAMP levels in schistosome homogenates (Humphries et al., 2004), and may regulate turbellarian regeneration (Kreshchenko, 2008; Kreshchenko et al., 2008), as well as development and regeneration of planarian reproductive tissues (Collins et al., 2010).
Exactly how temporal and spatial expression of these novel molecules as well as their effects on target cell types aid progression through the complex schistosome life cycle is not understood. This study focuses on one of the largest and most widespread of the NPP families, the L/M/Iamides. This family is represented by 11 genes (npp-1, -2, -6, -7, -9, -14, -15, -16, -22, -24, and -25 (McVeigh et al., 2009)); here we focus on Sm-npp-1 from S. mansoni. We report cDNA characterisation of the first proposed representatives of a new family of flatworm neuropentapeptides (displaying [G/A]FVR[I/L]. NH2 motifs), and identify these peptides in the schistosome nervous system. Whole worm physiology experiments demonstrate that GFVRIamide clearly reduced the mobility of adult schistosomes, although Sm-npp-1 RNA interference (RNAi) did not elicit a measurable, aberrant motility phenotype. The presence of NPP-1 like peptides in other flatworms and many nematodes illustrates that they represent a large and prominent helminth neuropeptide family, which may signify an appealing source of anthelmintic drug targets.
2. Materials and methods
2.1. Schistosome maintenance
Adults of the NMRI (Puerto Rican) strain of S. mansoni were supplied by Dr. Fred Lewis, Biomedical Research Institute, Rockville, Maryland, USA. Larval stages were maintained at Queen’s University Belfast, (QUB), UK in Biomphalaria glabrata snails. Cercariae shed from infected snails were mechanically transformed to schistosomulae (Lewis, 2001) and cultured at 37°C in a 5 % CO2 atmosphere in Complete RPMI, modified from Krautz-Peterson et al. (2007) (RPMI 1640 containing 20 % FCS, 10 mM HEPES, 2 mM L-Glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin). This culture was supplemented on alternate days with fresh media.
2.2. Bioinformatics
Initial sequence identification was performed through the Basic Local Alignment Search Tool (BLAST) (Altschul et al., 1990) server on the NCBI website (http://www.ncbi.nlm.nih.gov/BLAST). Searches were performed on the GenBank “est_others” dataset limited to the phylum of interest (Platyhelminthes or Nematoda). ‘Expect’ values were set to 10,000,000,000 to reduce the probability of real hits being filtered out by the software. Initial search queries were constructed based on the core structural identities of amidated propeptides, i.e. [KR-Xn-GKR]5 (where Xn represents a variable number of variable amino acids and the subscript 5 denotes that five concatenated repeats of this motif were employed as standard), as part of a larger neuropeptide identification effort (McVeigh et al., 2009). The schistosome I/Lamides reported here were identified by the search string [KRXXXXXGKR]5. All returned hits were translated to amino-acid sequence (www.expasy.ch/tools/dna.html), and all six reading frames were manually analysed for motifs representing putative amidated secretory peptides. All sequences identified were entered into a second wave of BLAST searches in order to identify further similar peptides. Where there was ambiguity, cleavage sites were selected to give the longest possible mature peptide. Secretory signal peptides were identified using the SignalP 3.0 server (www.cbs.dtu.dk/services/SignalP) (Emanuelsson et al., 2007).
2.3. Rapid Amplification of cDNA Ends- (RACE) PCR
mRNA was isolated from 20 mg S. mansoni adult worm pairs with the Dynabeads mRNA Direct Kit (Invitrogen, UK) and reverse transcribed into 5′ and 3′ RACE-ready cDNA with the SMART RACE cDNA Synthesis kit (Clontech, UK), following the manufacturer’s instructions. Schistosoma mansoni Sm-npp-1-specific oligonucleotide primers (designed against expressed sequence tag (EST) accession CD132695; Forward#1, 5′AAAAATAAAACTGTGTATATTCACAATTC; Forward#2, TTTCACCATTACTTTATAGACAATTTACA; Reverse#1, AACAGATCATTTTTTGTTGGTTAAC; Reverse#2, GCTTACCGATCCGAACAAAT) were employed in RACE-PCR methodology following the manufacturer’s instructions. Amplicons were cloned into pCR2.1TOPO, transformed into TOP10F cells (Invitrogen) and the resulting plasmid sequences were analysed by GATC Biotech (Germany).
2.4. Immunocytochemistry (ICC)
The following work involving animals was performed under the auspices of the appropriate UK Home Office licenses, and all aspects of animal care and maintenance guidelines were in accordance with Queen’s University Belfast guidelines.
Polyclonal antisera were raised to GFVRIamide (anti-GFVRIamide) in rabbits (Genosphere Biotechnologies, France) and affinity purified, or raised in guinea-pigs (anti-FMRFamide). Adult S. mansoni pairs were removed from infected Swiss mice, separated, flat-fixed in 4% (w/v) paraformaldehyde (PFA) in 0.1 M PBS, pH 7.4, and stored in antibody diluent (AbD, 0.1 M PBS containing 0.1 % (v/v) Triton X-100, 0.1 % (w/v) BSA and 0.1 % (w/v) NaN3) at 4 °C until use. Schistosomes were triple stained as follows; all steps were performed at 4 °C, washes (in AbD for 24 h with three to four changes of buffer) were performed between each step. Worms were incubated sequentially in: (i) anti-GFVRIamide (dilutions between 1/200 and 1/2000) for 96 h; (ii) goat-anti-rabbit Cy5 (Sigma Aldrich, diluted 1/100 in AbD) for 48 h; (iii) anti-GFVRIamide for 24 h (to ensure occupation of all available binding sites on the anti-rabbit secondary antisera); (iv) anti-FMRFamide (1/400) for 72 h; (v) goat-anti-guinea pig FITC (1/100) for 48 h; (vi) phalloidin-Tetramethyl Rhodamine Isothiocyanate (TRITC) (200 ng/ml) for 72 h, followed by a final 24 h wash. Ten day old schistosomules were fixed and processed as described above, except in a single-labelling protocol. Following fixation, schistosomules were incubated in anti-GFVRIamide (1/500) for 96 h, goat-anti-rabbit FITC (1/100) for 48 h, phalloidin TRITC (200 ng/ml) for 72 h. Worms were mounted in PBS/glycerol (1/9 [v/v]), and viewed on Leica SP2 or SP5 confocal scanning laser microscopes. Control experiments involved preadsorption of antibody with GFVRIamide antigen (1/1500 dilution anti-GFVRIamide preadsorbed with ≥250 ng/ml GFVRIamide antigen) which completely abolished detectable staining.
2.5. Whole worm peptide physiology
Adult S. mansoni pairs were removed from infected Swiss mice up to 8 h before experiments and maintained in RPMI 1640 at 37°C in a 5% CO2 atmosphere. The effects of peptides on worms were assessed by live video recording of worm movement and analysis with Ethovision 3.1 motion-tracking software (Noldus Information Technology, The Netherlands, www.noldus.com). Quantification utilised Ethovision’s “mobility” feature, where the system records multiple images of the worm per second and measures the proportional change in worm area between images – mobility is therefore defined as the percent change in two-dimensional worm area over time. This approach allows for sensitive quantification of gross changes in worm shape and is well suited to analysis of adult schistosomes, which can display considerable movement in one location, without migrating around the field of view.
A single worm pair was placed in the arena (i.e. camera field of view) in 2 ml RPMI 1640 at 37°C in a temperature-controlled dish. Motility was recorded for a pre-treatment period of 5 min to establish a baseline for spontaneous movement, followed by addition of warm peptide and recording of mobility for 5 min. Peptide containing medium was then replaced with fresh medium, and motility recorded for a further 5 min to investigate reversibility (washout) of peptide-induced effects. Data were exported into spreadsheet software and all mobility figures normalised to the pre-treatment baseline mean (equalled 100%). Data were graphed and statistically analysed by one-way ANOVA with Tukey’s post-hoc test with Graphpad Prism software. All peptides were synthesised by EZBiolab (IN, USA).
2.6. RNAi and quantitative PCR (qPCR)
Gene silencing was performed in schistosomules using a protocol modified from published electroporation-induced RNAi methods (Correnti et al., 2005; Krautz-Peterson et al., 2007; Ndegwa et al., 2007; Morales et al., 2008; Zhao et al., 2008; Faghiri and Skelly, 2009). Briefly, a 260 bp region of the Sm-npp-1 open reading frame (ORF) (GU173993) was amplified to produce separate templates containing T7 promoters for sense and antisense strand RNA transcription (forward, 5′-TTTCACCATTACTTTATAGACAATTTACA-3′; reverse, 5′-GCTTACCGATCCGAACAAAT-3′). Single-stranded RNAs were synthesized, annealed to form double-stranded RNA (dsRNA) and purified using the MegaScript RNAi Kit (Ambion, USA). Non-target control dsRNAs included tomato chloroplast-specific ribosomal protein (CSRP from Lycopersicon esculentum; AY568722; primer sequences: forward, 5′-TGGCGACTCTTTCCCTTTC -3′; reverse, 5′-CTATTCCTGACAGGAACA -3′) and S. mansoni alkaline phosphatase (SmAP; EU040139; primer sequences: forward, 5′-TACTGGAGCAAGATATTTGAAAGC -3′; reverse, 5′-GGCGTACATGATAGAAGATTTCG -3′). The integrity of all dsRNAs was validated by non-denaturing agarose gel electrophoresis and quantified using a NanoDrop ND1000 spectrophotometer. Two hundred, 10-day old, in vitro-derived schistosomulae were electroporated in 100 ul electroporation medium (RPMI 1640 containing 10 mM HEPES) containing 10 ug dsRNA using a BTX ECM830 Electro Square Porator (125 V, 20 ms at room temperature in a 4 mm-gap cuvette; two pulses separated by 100 ms). Triplicate transfections were performed for experimental dsRNAs alongside non-target dsRNA control treatments and untreated controls lacking dsRNA. Following electroporation, worms were placed in Complete RPMI medium for 7 days and monitored at least once daily for behavioural, developmental and mobility changes. Fresh medium was added on alternate days.
After 7 days, total RNA was extracted from worms using the RNeasy micro kit (Qiagen), and DNase treated. Two cDNAs were synthesised for each biological replicate using 15 ng total RNA (ABI High-Capacity RNA to cDNA kit, Applied Biosystems), and stored at −20°C.
Knockdown and developmental expression were assessed by qPCR using the Platinum Taq SYBR Green qPCR Supermix (Invitrogen) on a RotorGene Q 5-plex HRM qPCR instrument (Qiagen). PCR reactions were performed in triplicate on each cDNA with the equivalent of 1.5 ng cDNA and 200 nM of each primer per reaction. Primers were designed using Applied Biosystems’ Primer Express 3.0 software and comprised: Sm-npp-1 (GU173993; forward primer: 5′-CACATGGATCCAGTCGTTGT-3′; reverse primer 5′-TGTGATTGTACTTTCAATCTGTCA-3′) as well as alpha-tubulin (Sm-atb, M80214; forward primer: 5′-CATTCATTTGGCGGTGGTACT-3′; reverse primer: 5′-ACACAGCAAACTCCAATTTCGA-3′) and neuronal peptidylglycine-α-hydroxylating monooxygenase (Sm-phm; AY172995; forward primer: 5′-CCAGGCAGTCCAGAACGTTT -3′; reverse primer” 5′-CCATCGCCCAAGAATACACA -3′) reference genes. Primer amplification efficiencies were determined on an individual reaction basis using PCR Miner software (http://miner.ewindup.info/miner/Version2/index.htm) (Zhao and Fernald, 2005). Quantification calculations employed the augmented comparative Ct method (ΔΔCt) (Pfaffl, 2001). In temporal expression analyses, percent-difference figures were calculated relative to the adult stage, which was selected as an arbitrary point of reference. All data were graphed and analysed by one-way ANOVA with Tukey’s post-test, using Graphpad Prism 5 software. Significance was set at P<0.05.
3. Results
We have identified a neuropeptide-encoding cDNA in S. mansoni (Sm-npp-1) which encodes a prepropeptide bearing two novel neuropeptides. Using antisera raised against one of these peptides (GFVRIamide), we demonstrate localisation to the schistosome CNS. Although GFVRIamide lacked measurable activity in isolated muscle cell assays, we showed that exogenously applied GFVRIamide had an inhibitory effect on schistosome mobility. Knockdown of Sm-npp-1 transcript by RNAi was successful, but elicited no changes in mobility or obvious aberrant phenotypes. We identified a large number of similar I/Lamide peptides in both platyhelminth and nematode phyla, suggesting that these peptides represent a novel family of helminth neuropeptides.
3.1. Sm-npp-1 encodes three novel secretory peptides
BLAST scans of platyhelminth ESTs available in the GenBank database identified an EST which encodes a novel S. mansoni prepropeptide of 95 amino acids; the EST sequence was confirmed and the entire cDNA amplified by RACE-PCR. The ORF contains three putative glycine-extended propeptides, each flanked by mono or dibasic cleavage sites, indicating that Sm-npp-1 would produce a single AFVRLamide and two GFVRIamide peptides (GU173993; Fig. 1).
Fig. 1.

Schistosoma mansoni neuropeptide precursor 1 (Sm-npp-1) cDNA sequence. Sm-npp-1 encodes three predicted peptides. Nucleotide sequence is denoted in lowercase and amino-acid translation in uppercase. N-terminal secretory signal peptide sequence is denoted by bold, italicised script. Peptide-encoding region of prepropeptide is highlighted in grey, with predicted peptides in bold, flanking mono/di-basic cleavage residues underlined and putative polyadenylation signal double-underlined.
3.2. NPP-1-like peptides are present in both helminth phyla
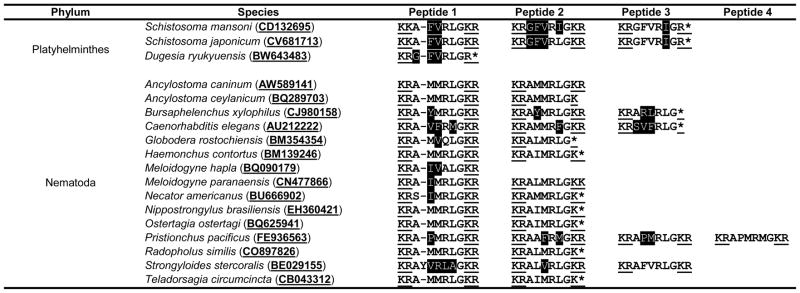
Following discovery of Sm-npp-1, further BLAST searches were performed using GFVRIG and AFVRLG as queries, which identified sequelogs in three other flatworm species (Table 1): Schistosoma japonicum Sj-npp-1 encodes single copies of three similar peptides (AFVRLamide, GFVRLamide, and GFVRIamide); Macrostomum lignano Ml-npp-1 encodes the putative orthologous peptides PSFVRMamide and ASYVRMamide; Schmidtea mediterranea Smed-npp-1 encodes SFVRLamide and three copies of ASFVRLamide (all reported in McVeigh et al., 2009), with Schmidtea peptides confirmed by MS in Collins et al. (2010); while Dugesia ryukyuensis Dr-npp-1 encodes a single copy of GFVRIamide. When BLAST searches were expanded by removing the “Platyhelminth” restriction within the software, we found sequelogs of these peptides in fifteen species of free-living and parasitic nematode (Table 1). Most of the these variants are –RLamides which were not reported amongst the nematode neuropeptide-like proteins (NLPs) (Husson et al., 2005, 2007; Li, 2005; McVeigh et al., 2008). Most notable amongst these is the Caenorhabditis elegans transcript (AU212222), which in fact represents the FMRFamide-like peptide gene flp-20; it encodes two copies of FLP-20 (AMMRFamide) as reported by Li (2005) as well as the previously unreported NLPs AVFRMamide and SFVRLamide. All of the flp-20 transcripts originating from parasitic nematodes encode at least two copies of AXXXLamides but, interestingly, none aside from C. elegans encode FLP-20.
Table 1.
Neuropeptide Precursor-1 (NPP-1)-like peptides in helminths. Conservation between predicted peptides on npp-1-like transcripts is illustrated. Identical residues are denoted by black text on white background, conservative substitutions by white text on black background. Representative accession numbers or references are given for each species.
|
3.3. GFVRIamide is expressed in CNS neurones and does not co-localise with FLPs
In adult worms, a triple-labelling ICC protocol was employed involving an anti-GFVRIamide antiserum in conjunction with anti-FMRFamide serum (serving as a marker for the extensive peptidergic nervous system) to identify the expression patterns of GFVRIamide and examine possible co-localisation of GFVRIamide and FLPs. GFVRIamide-IR was restricted to a small set of neurones within the CNS. In both male and female adults (Fig. 2) IR localized to a single pair of cells with soma in the cerebral ganglia, each of which projected across the cerebral commissure and anteriorly, as well as posteriorly along the main nerve cords (mnc). At no location was any peripheral innervation of organ systems visible (n=25) (e.g. the reproductive system with the egg chamber or ootype, the digestive system), a feature common to FLPs in all classes of flatworm (Arafa et al., 2007; Johnston et al., 1996; Maule et al., 1993; Sebelová et al., 2004). Further, GFVRIamide-IR was not localized within the submuscular nerve plexus, which is richly innervated by both FLP- and NPF-immunopositive nerves. Although both GFVRIamide and FMRFamide IRs localised to the same CNS nerve tracts, their IRs were distinct and did not overlap, i.e. GFVRIamide and FLPs are expressed in separate neurones. A similar pattern of GFVRIamide-IR was observed in schistosomules (Fig. 2E) and cercariae (data not shown), delineating a pair of neuronal cell bodies in the brain which project posteriorly along the main nerve cords, as well as anteriorly towards the oral sucker.
Fig. 2.

Immunocytochemical localisation (ICC) of the Sm-npp-1 neuropeptide GFVRIamide in adult Schistosoma mansoni. (A-D) Blue staining (pseudocoloured Cy5-labelled secondary antibody) indicates GFVRIamide distribution, green (FITC-labelled secondary antibody) represents distribution of FMRFamide-like peptides (FLPs), while red (TRITC; tetrarhodamine isothiocyanate-labelled phalloidin) indicates distribution of filamentous actin; (E) GFVRIamide is denoted by green FITC label. (A) Within the brain of adult S. mansoni, GFVRIamide is expressed in a pair of neurones (arrowheads). GFVRIamide immunoreactivity (IR) projects across the cerebral commissure (CC) and posteriorly along main nerve cords (MNC). (B) One of the bipolar brain neurones expressing GFVRIamide (arrowhead). While GFVRIamide- and FLP-immunopositive nerves occur in close apposition, they never overlap, indicating that they are expressed in separate cells/fibres. (C) FLP-IR is strong in nerves innervating the female ootype (Ot) whereas GFVRIamide-IR appears only in the MNC. (D) Both GFVRIamide- and FLP-IRs are apparent along the entire MNC, running adjacent to the gut (Gt). GFVRIamide-IR is restricted to a single longitudinal nerve tract, while FLP-IR appears in multiple longitudinal nerve tracts in addition to its association with peripheral innervation of somatic muscle layers. (E) In schistosomules, GFVRIamide (stained green) shows a similar expression pattern to adults, localising to neurones associated with each cerebral ganglion (arrowheads) and the longitudinal nerve cords. Ac, acetabulum; OS, oral sucker; Oes, oesophagus.
3.3. Sm-npp-1 is constitutively expressed in cercariae and adults
qPCR analysis of Sm-npp-1 expression relative to Sm-phm (selected as a neuronal reference gene) demonstrated no significant difference between cercariae, schistosomulae and adult worms (n=3; data not shown). These figures support our ICC data, and suggest that Sm-npp-1 may have a constitutive role throughout the schistosome life cycle.
3.4. Exogenously applied GFVRIamide depresses schistosome mobility
The following mobility figures have been normalised relative to a pretreatment mean mobility of 100%, and are expressed as the mean effect over the 5 min treatment period. Exogenous application of GFVRIamide to whole adult worm pairs in vitro elicited an inhibitory effect on worm mobility (10−5 M GFVRIamide: −44.84±4.31 %, P =0.0002, n=9) comparable to YIRFamide (10−5 M YIRFamide mean effect: −51.20±4.72 %, P =0.0014, n=5) (Fig. 3A). The effects of both peptides diminished significantly upon washout, but worm mobility did not recover completely to pre-treatment levels within the assay timeframe (YIRFamide effect post washout: −17.8±9.34 %, P =0.0021 versus treatment; GFVRIamide effect post washout: −28.89±1.55 %, P =0.0371 versus treatment; Fig. 3A).
Fig. 3.

Physiology of the the Sm-npp-1 neuropeptide GFVRIamide in adult Schistosoma mansoni. (A) Effect of GFVRIamide and YIRFamide on mobility of S. mansoni. During a 5 min period of peptide addition (min 5–10, denoted by grey box), both peptides significantly reduce schistosome mobility. Worm mobility recovers following washout (min 10–15), indicating reversibility of effects. (B) Mean effects of GFVRIamide/YIRFamide peptides and analogues on schistosome mobility. Both 10−5 M GFVRIamide and YIRFamide exhibit significant effects compared with vehicle controls (H2O). Neither the unamidated peptides (YIRF-OH, GFVRI-OH) nor an alanine-substituted analogue (GFVRAamide) had significant effects versus vehicle controls. Effects of both YIRFamide and GFVRIamide were significantly different from their relevant control peptides. (C) Alternating sequential addition of GFVRIamide and YIRFamide (first peptide at 10−7 M, second at 10−6 M) induced similar effects to single additions, suggesting that the effects of these two peptides are essentially indistinguishable. All data points are expressed as mean ± SEM. a, amide. *P < 0.05, **P < 0.01, ***P < 0.005.
All controls elicited only small, non-significant effects, and included a water vehicle control (+8.1±3.20 %, P >0.05, n=5) and three peptide analogues each at 10−5 M concentration (GFVRI-OH (non-amidated free acid), −12.8±3.88 %, P >0.05, n=11; GFVRAamide, −23.24±4.18 %, P >0.05, n=10; YIRF-OH, +1.5±2.05 %, P >0.05, n=4) (Fig. 3B).
Sequential addition of GFVRIamide and YIRFamide in alternating order (first peptide at 10−7 M, second peptide at 10−6 M), produced quantitatively and qualitatively similar effects to single additions (Fig. 3C), illustrating that the biological effects of these peptides are essentially indistinguishable in this whole-worm assay. Taken together these experiments show that GFVRIamide and YIRFamide induce similar, inhibitory effects on whole worm mobility. As known for FLPs and NPFs (Day et al., 1994; Humphries et al., 2004), amidation of the C-terminal amino acid (isoleucine) is also a prerequisite for biological function for L/Iamides.
3.5. Sm-npp-1 transcripts are susceptible to RNAi in schistosomules
As a complementary approach to identify the bio-activity of NLP peptides we used an electroporation-based RNAi protocol to induce Sm-npp-1 knockdown in 10 day-old schistosomules in vitro. We transfected a 260 bp dsRNA (100 ng/μl) representing a portion of the Sm-npp-1 ORF, and then maintained worms in culture for 7 days. At this point, transcript knockdown was quantified using qPCR, while post-knockdown phenotype was assessed by manual observation of schistosomule motility and morphology/development. A statistically significant knockdown was achieved (Fig. 4, quantified as ΔΔCt of Sm-npp-1 transcript relative to Sm-atb reference transcript), comprising a 54.46±10.41% (n=3) reduction in Sm-npp-1 dsRNA-treated worms when compared with untreated worms (P <0.001, q=8.190), CSRP dsRNA-treated control worms (16.0±10.76% increase, n=2; P <0.001, q=7.426) and SmAP dsRNA-treated controls (6.72±2.38% reduction, n=2; P <0.01, q=6.209). Manual analysis of post-RNAi worm motility up to day 7 post-electroporation suggested that knockdown did not elicit any measurably aberrant phenotype in either motility (quantified as number of gross worm contractions per minute: untreated, 8±2.27; control dsRNA, 10.25±0.25, Sm-npp-1 dsRNA, 8.83±1.08), or morphology (quantified as distribution of: healthy (translucent appearance, smooth tegument, full movement and regular morphology); aberrant (visible movement, but with aberrant morphology/appearance); or dead (no visible movement, major aberrant morphology), Fig. 4).
Fig. 4.
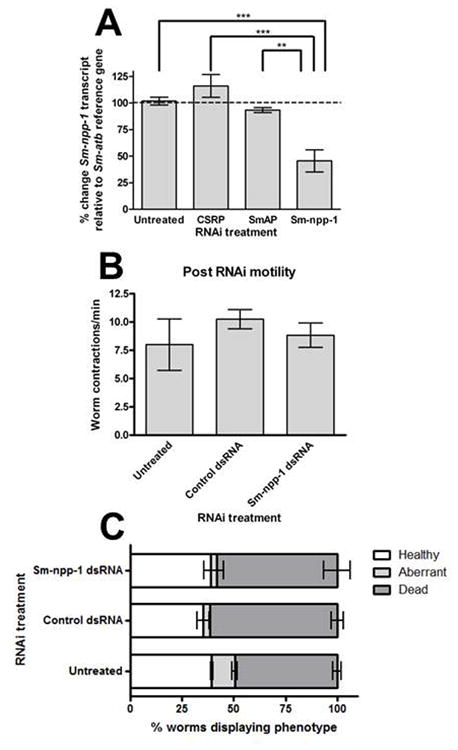
RNA interference (RNAi) analysis of Schistosoma mansoni neuropeptide precursor Sm-npp-1. (A) Quantitative PCR analysis of Sm-npp-1 transcript knockdown. Percent change in Sm-npp-1 transcript levels relative to alpha-tubulin (Sm-atb) reference gene, following treatment with one of the following double-stranded RNAs (dsRNAs): chloroplast-specific ribosomal protein (CSRP) non-target control; S. mansoni alkaline phosphatase (SmAP) non-target control; or target-specific Sm-npp-1. Knockdown of Sm-npp-1 is specific and statistically significant relative to untreated and control-treated worms. (B) Analysis of post-RNAi motility phenotype, quantified as mean number of worm whole-body contractions per minute. (C) Morphological analysis of post-RNAi phenotype. Worms were defined as one of three phenotypes, healthy (normal movement and morphology), aberrant (visible movement, aberrant morphology) or dead (no visible movement, aberrant morphology). Data points are expressed as mean % transcript change ± SEM. **P < 0.01, ***P < 0.005.
4. Discussion
Despite increasing knowledge of the structure and diversity of flatworm neuropeptides, little is known about the cell-specific expression and biological functions of individual peptides. This study has described a new family of helminth neuropeptides and investigated their biology in schistosomes. We have localised the seminal NPP-1-like, I/Lamide peptides in S. mansoni, and demonstrated that GFVRIamide induces a clear reduction in movement of whole, adult schistosomes. Knock-down of Sm-npp-1 transcript elicited no detectable post-RNAi phenotype. In summary, we suggest that GFVRIamides modulate worm motility; however, the explicit functions (direct inhibition of individual muscle fibres or through signalling to muscle innervating nerves) of Sm-NPP-1 peptides in the schistosome nervous system remain to be uncovered.
Bioinformatics searches indicated that peptides with similarity to I/Lamides encoded by Sm-npp-1 also exist in nematodes where transcripts generally encode two peptides with a highly conserved AXXRLamide motif (Table 1). Interestingly, the C. elegans EST actually represents the previously reported flp-20 gene, which encodes two FLP-20 peptides (AMMRFamide) (Li, 2005) but also novel putative neuropeptides (AVFRMamide and SFVRLamide, which would be designated as NLPs under current nematode nomenclature since they are not FLPs (-RFamides) nor insulin-related (INS) peptides). In demonstrating that C. elegans flp-20 encodes both ‘FLP’ and ‘NLP’-like peptides, this phenomenon blurs the distinction between the flp and nlp families of nematode neuropeptide genes, illustrating the futility of casting these putative neuropeptides into separate families based on small differences in amino acid sequence. Although FLP-20 (AMMRFamide) itself has not been reported from any nematode species except C. elegans, strong similarities between the I/Lamides encoded by the nematode transcripts reported here could identify them as flp-20 orthologues, and we propose that the nematode sequences identified here be named as such. While flp-20 is implicated in control of male sexual turning behaviour in C. elegans (Liu et al., 2007), no male-specific behavioural effects were observed in schistosomes during this study, making it difficult to make any meaningful functional comparisons.
While similar peptides (including –RLamides, -SIamides and -FVRIamides) have been described in other invertebrate phyla, including Annelida and Mollusca (Chen et al., 1995; Pedder et al., 2001; El Filali et al., 2006; Jekely et al., 2008; Takahashi et al., 2009), we could find no evidence for the presence of similar peptides in vertebrate host organisms. The NPP-1-like peptides reported here thus represent a group of novel helminth neuropeptides which, if they are absent from host species, and are functionally conserved across helminths, could represent particularly promising avenues for drug development. Provided that their endogenous receptors are ligand selective, drugs targeting helminth-specific receptors should be pathogen-specific and unlikely to exhibit host-toxicity through non-specific interactions with host receptors.
GFVRIamide expression is restricted to a pair of neurones in the schistosome CNS which contrasts strongly with the abundance of FMRFamide-IR in the brain, nerve cords and commissures, as well as innervation of oral and ventral suckers and the female reproductive apparatus (Marks and Maule, 2008). We employed a triple-labelling ICC protocol to investigate possible co-localisation of GFVRIamide and FLPs; GFVRIamide and FMRFamide did not co-localized in any of the specimens examined. These genes are therefore expressed in separate neurones, indicating that GFVRIamide and FLPs perform separate functions in the schistosome nervous system. FMRFamide-IR is clearly visible in muscle-innervating nerves and plexi, consistent with the myoexcitatory properties observed for schistosome FLPs (McVeigh et al., 2005). This spatial expression pattern appears different from that reported for Smed-npp-1, which has been localised, by in situ hybridization, to several neurones in and around the planarian’s pharynx (Collins et al., 2010). This indicates a possible inter-species difference in function for NPP-1-like peptides.
Based on its localisation pattern, GFVRIamide may be involved in interneuronal communication within the schistosome CNS. In order to examine GFVRIamide’s function in greater detail, we performed two further experiments: (i) Exogenous application of GFVRIamide to whole adult schistosomes; (ii) RNAi of Sm-npp-1. In adult worm pairs GFVRIamide reduced motile behaviour at 10−5 M. At the same concentration the positive control peptide, YIRFamide, exhibited a similar magnitude of inhibition, an effect which was surprising given YIRFamide’s potent myoexcitatory effect on individual muscle cells (Day et al., 1997). However, reduced mobility could be a consequence of spastic (excitatory) or flaccid (muscular relaxation) paralyses, both of which were observed during these assays, with a mixture of corkscrewing, curling and waving, but also straightening behaviours (data not shown). It is therefore difficult to ascertain whether the inhibitory effect on schistosome mobility was a consequence of muscular flaccid or spastic paralysis and we recognise this as a shortcoming of this particular assay. Nonetheless, the effects observed appeared receptor-specific since the non-amidated, free-acid analogues GFVRI-OH and YIRF-OH, as well as the alanine substituted analogue GFVRAamide lacked significant activity. These results are consistent with YIRF-OH’s lack of effect on dispersed schistosome muscle fibres (Day et al., 1997) and corroborate the requirement for an amidated C-terminus for bio-activity of many, small peptide transmitters. Additionally, post-peptide washout of the bioactive amidated variants produced a statistically significant return towards pretreatment mobility levels, indicating that these effects are reversible and thus likely receptor-mediated. In summary, it is clear that exogenously applied GFVRIamide has a modulatory effect on the muscular function of adult schistosomes. These points notwithstanding, several GPCRs are present on the schistosome tegument (Chatterjee et al., 2007; El-Shehabi et al., 2009), and some of the effects reported may be due to activation of tegumental receptors. Nevertheless, in an in vivo setting the observed inhibitory effects could have deleterious consequences for the parasite, suggesting that GFVRIamide-mimetics may have potential as candidate drugs.
Electroporation-mediated, dsRNA-induced RNAi of Sm-npp-1 elicited a mean 50% transcript knockdown. Although this level of knockdown is modest when compared with most other experiments reported in the literature (e.g. Krautz-Peterson et al., 2007), it is worth noting that even lower levels of transcript knockdown can successfully ablate a target protein - Taft and Yoshino (2011) report that 30% knockdown of calmodulin mRNA in schistosome primary sporocysts leads to a concurrent 35% reduction in calmodulin protein and a ‘stunted growth’ phenotype. Using qPCR to monitor transcript knockdown over three biological replicates), we observed individual 60.23±7.45%, 59.09±9.38% and 44.06±16.11% reductions in Sm-npp-1 mRNA. In none of these experiments could we discern any aberrant post-RNAi phenotypes, suggesting one or a combination of the following: these levels of transcript knockdown were insufficient to effect a reduction in synaptic levels of Sm-NPP-1 peptides within the assay timeframe (although we attempted to use anti-GFVRIamide serum in a quantitative ICC protocol post-RNAi, schistosomulae did not stain in a sufficiently consistent manner to permit accurate quantification of peptide knockdown); Sm-NPP-1 peptides have a redundant function, making Sm-npp-1 refractory to analysis by RNAi; or our phenotypic assays are inappropriate or too insensitive to identify Sm-npp-1-RNAi phenotypes (e.g. in the flatworm S. mediterrannea, knockdown of neuropeptide-encoding npy transcipts elicits a sexual development phenotype (Collins et al., 2010)). Either way, it is currently impossible to make confident assertions about the specific functions of NPP-1 peptides in the schistosome neuromuscular system. RNAi experiments on a schistosome YIRFamide precursor were precluded by the absence of sequence data.
These points illustrate the difficulty of investigating the functionality of helminth neuropeptides using RNAi technology, although this work, along with another recent study (Stefanic et al., 2010), demonstrate that the schistosome neuromuscular system is not refractory to RNAi-based transcript knockdown. Coupled with the development of appropriately sensitive in vitro or in vivo assays, RNAi should permit functional studies/target validation of neuromuscular genes in parasitic flatworms.
Research Highlights.
We have identified a novel neuropeptide familyin helminths.
Here, we have studied Schistosomamansoni neuropeptide precursor #1(Sm-npp-1).
Sm-NPP-1 peptides localise to the schistosome central nervous system.
Sm-NPP-1 peptides trigger reduced motility in adult schistosomes.
Knockdown of Sm-npp-1 does not elicit measurable motility or morphology phenotypes.
Acknowledgments
The authors wish to thank Alan Wilson (University of York, UK), Padraic Fallon (Trinity College Dublin, Ireland) and John Kusel (University of Glasgow, UK) in particular, for supply of Biomphalaria snails and for assistance with establishing the S. mansoni life cycle at Queen’s University Belfast, UK and to Dr John McGrath (Queen’s University Belfast) for providing access to a quantitative PCR instrument. This work was funded by The National Institutes of Health, USA, grant R01-AI49162.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- Arafa SZ, El-Naggar MM, El-Abbassy SA, Stewart MT, Halton DW. Neuromusculature of Gyrodactylus rysavyi, a monogenean gill and skin parasite of the catfish Clarias gariepinus. Parasitol Int. 2007;56:297–307. doi: 10.1016/j.parint.2007.06.005. [DOI] [PubMed] [Google Scholar]
- Chatterjee S, Op De Beeck J, Rao AV, Desai DV, Vrolix G, Rylant F, Panis T, Bernat A, Van Bergen J, Peeters D, Van Marck E. Prolonged somatostatin therapy may cause down-regulation of SSTR-like GPCRs on Schistosoma mansoni. J Vector Borne Dis. 2007;44:164–180. [PubMed] [Google Scholar]
- Chen ML, Muneoka Y, Walker RJ. Actions of a small cardioactive peptide from mytilus, APNFLAYPRLamide, on central neurones of Helix aspersa. Gen Pharmacol. 1995;26:273–283. doi: 10.1016/0306-3623(94)00190-x. [DOI] [PubMed] [Google Scholar]
- Collins JJ, 3rd, Hou X, Romanova EV, Lambrus BG, Miller CM, Saberi A, Sweedler JV, Newmark PA. Genome-wide analyses reveal a role for peptide hormones in planarian germline development. PLoS Biol. 2010;8:e1000509. doi: 10.1371/journal.pbio.1000509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Correnti JM, Brindley PJ, Pearce EJ. Long-term suppression of cathepsin B levels by RNA interference retards schistosome growth. Mol Biochem Parasitol. 2005;143:209–215. doi: 10.1016/j.molbiopara.2005.06.007. [DOI] [PubMed] [Google Scholar]
- Day TA, Maule AG. Parasitic peptides! The structure and function of neuropeptides in parasitic worms. Peptides. 1999;20:999–1019. doi: 10.1016/s0196-9781(99)00093-5. [DOI] [PubMed] [Google Scholar]
- Day TA, Maule AG, Shaw C, Halton DW, Moore S, Bennett JL, Pax RA. Platyhelminth FMRFamide-related peptides (FaRPs) contract Schistosoma mansoni (trematoda: Digenea) muscle fibres in vitro. Parasitology. 1994;109:455–459. doi: 10.1017/s0031182000080707. [DOI] [PubMed] [Google Scholar]
- Day TA, Maule AG, Shaw C, Pax RA. Structure-activity relationships of FMRFamide-related peptides contracting Schistosoma mansoni muscle. Peptides. 1997;18:917–921. doi: 10.1016/s0196-9781(97)00073-9. [DOI] [PubMed] [Google Scholar]
- Dougan PM, Mair GR, Halton DW, Curry WJ, Day TA, Maule AG. Gene organization and expression of a neuropeptide Y homolog from the land planarian Arthurdendyus triangulatus. J Comp Neurol. 2002;454:58–64. doi: 10.1002/cne.10440. [DOI] [PubMed] [Google Scholar]
- El Filali Z, Van Minnen J, Liu WK, Smit AB, Li KW. Peptidomics analysis of neuropeptides involved in copulatory behavior of the mollusk Lymnaea stagnalis. J Proteome Res. 2006;5:1611–1617. doi: 10.1021/pr060014p. [DOI] [PubMed] [Google Scholar]
- El-Shehabi F, Vermeire JJ, Yoshino TP, Ribeiro P. Developmental expression analysis and immunolocalization of a biogenic amine receptor in Schistosoma mansoni. Exp Parasitol. 2009;122:17–27. doi: 10.1016/j.exppara.2009.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emanuelsson O, Brunak S, von Heijne G, Nielsen H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat Protoc. 2007;2:953–971. doi: 10.1038/nprot.2007.131. [DOI] [PubMed] [Google Scholar]
- Faghiri Z, Skelly PJ. The role of tegumental aquaporin from the human parasitic worm, Schistosoma mansoni, in osmoregulation and drug uptake. FASEB J. 2009;23:2780–2789. doi: 10.1096/fj.09-130757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham MK, Fairweather I, McGeown JG. The effects of FaRPs on the motility of isolated muscle strips from the liver fluke, Fasciola hepatica. Parasitology. 1997;114:455–465. doi: 10.1017/s0031182096008712. [DOI] [PubMed] [Google Scholar]
- Graham MK, Fairweather I, McGeown JG. Second messengers mediating mechanical responses to the FaRP GYIRFamide in the fluke Fasciola hepatica. Am J Physiol Regul Integr Comp Physiol. 2000;279:R2089–94. doi: 10.1152/ajpregu.2000.279.6.R2089. [DOI] [PubMed] [Google Scholar]
- Hrckova G, Velebny S, Halton DW, Day TA, Maule AG. Pharmacological characterisation of neuropeptide F (NPF)-induced effects on the motility of Mesocestoides corti (syn. Mesocestoides vogae) larvae. Int J Parasitol. 2004;34:83–93. doi: 10.1016/j.ijpara.2003.10.007. [DOI] [PubMed] [Google Scholar]
- Humphries JE, Kimber MJ, Barton YW, Hsu W, Marks NJ, Greer B, Harriott P, Maule AG, Day TA. Structure and bioactivity of neuropeptide F from the human parasites Schistosoma mansoni and Schistosoma japonicum. J Biol Chem. 2004;279:39880–39885. doi: 10.1074/jbc.M405624200. [DOI] [PubMed] [Google Scholar]
- Husson SJ, Clynen E, Baggerman G, De Loof A, Schoofs L. Discovering neuropeptides in Caenorhabditis elegans by two dimensional liquid chromatography and mass spectrometry. Biochem Biophys Res Commun. 2005;335:76–86. doi: 10.1016/j.bbrc.2005.07.044. [DOI] [PubMed] [Google Scholar]
- Husson SJ, Mertens I, Janssen T, Lindemans M, Schoofs L. Neuropeptidergic signaling in the nematode Caenorhabditis elegans. Prog Neurobiol. 2007;82:33–55. doi: 10.1016/j.pneurobio.2007.01.006. [DOI] [PubMed] [Google Scholar]
- Jekely G, Colombelli J, Hausen H, Guy K, Stelzer E, Nedelec F, Arendt D. Mechanism of phototaxis in marine zooplankton. Nature. 2008;456:395–399. doi: 10.1038/nature07590. [DOI] [PubMed] [Google Scholar]
- Johnston RN, Shaw C, Halton DW, Verhaert P, Baguna J. GYIRFamide: A novel FMRFamide-related peptide (FaRP) from the triclad turbellarian, Dugesia tigrina. Biochem Biophys Res Commun. 1995;209:689–697. doi: 10.1006/bbrc.1995.1554. [DOI] [PubMed] [Google Scholar]
- Johnston RN, Shaw C, Halton DW, Verhaert P, Blair KL, Brennan GP, Price DA, Anderson PA. Isolation, localization, and bioactivity of the FMRFamide-related neuropeptides GYIRFamide and YIRFamide from the marine turbellarian Bdelloura candida. J Neurochem. 1996;67:814–821. doi: 10.1046/j.1471-4159.1996.67020814.x. [DOI] [PubMed] [Google Scholar]
- Krautz-Peterson G, Radwanska M, Ndegwa D, Shoemaker CB, Skelly PJ. Optimizing gene suppression in schistosomes using RNA interference. Mol Biochem Parasitol. 2007;153:194–202. doi: 10.1016/j.molbiopara.2007.03.006. [DOI] [PubMed] [Google Scholar]
- Kreshchenko ND. Functions of flatworm neuropeptides NPF, GYIRF and FMRF in course of pharyngeal regeneration of anterior body fragments of planarian, Girardia tigrina. Acta Biol Hung. 2008;59(Suppl):199–207. doi: 10.1556/ABiol.59.2008.Suppl.29. [DOI] [PubMed] [Google Scholar]
- Kreshchenko ND, Sedelnikov Z, Sheiman IM, Reuter M, Maule AG, Gustafsson MK. Effects of neuropeptide F on regeneration in Girardia tigrina (platyhelminthes) Cell Tissue Res. 2008;331:739–750. doi: 10.1007/s00441-007-0519-y. [DOI] [PubMed] [Google Scholar]
- Lewis F. Schistosomiasis. In: Coligan JE, editor. Current Protocols in Immunology. Unit 19.1. Chapter 19. New York: Greene Pub. Associates and Wiley-Interscience; 2001. c1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li C. The ever-expanding neuropeptide gene families in the nematode Caenorhabditis elegans. Parasitology. 2005;131(Suppl):S109–27. doi: 10.1017/S0031182005009376. [DOI] [PubMed] [Google Scholar]
- Liu T, Kim K, Li C, Barr MM. FMRFamide-like neuropeptides and mechanosensory touch receptor neurons regulate male sexual touching behaviour in Caenorhabditis elegans. J Neurosci. 2007;27:7174–7182. doi: 10.1523/JNEUROSCI.1405-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mair GR, Halton DW, Shaw C, Maule AG. The neuropeptide F (NPF) encoding gene from the cestode, Moniezia expansa. Parasitology. 2000;120:71–77. doi: 10.1017/s0031182099005296. [DOI] [PubMed] [Google Scholar]
- Marks NJ, Johnson S, Maule AG, Halton DW, Shaw C, Geary TG, Moore S, Thompson DP. Physiological effects of platyhelminth RFamide peptides on muscle-strip preparations of Fasciola hepatica (trematoda: Digenea) Parasitology. 1996;113:393–401. [Google Scholar]
- Marks NJ, Maule AG. Neuropeptides in helminths: occurrence and distribution. Adv Exp Med Biol. 2008;692:49–77. doi: 10.1007/978-1-4419-6902-6_4. [DOI] [PubMed] [Google Scholar]
- Marks NJ, Maule AG, Halton DW, Geary TG, Shaw C, Thompson DP. Pharmacological effects of nematode FMRFamide-related peptides (FaRPs) on muscle contractility of the trematode, Fasciola hepatica. Parasitology. 1997;114:531–539. [PubMed] [Google Scholar]
- Maule AG, Halton DW, Shaw C, Johnston CF. The cholinergic, serotoninergic and peptidergic components of the nervous system of Moniezia expansa (Cestoda, Cyclophillidia) Parasitology. 1993;106:429–440. doi: 10.1017/s0031182000067184. [DOI] [PubMed] [Google Scholar]
- Maule AG, Shaw C, Halton DW, Brennan GP, Johnston CF, Moore S. Neuropeptide F (Moniezia expansa): Localization and characterization using specific antisera. Parasitology. 1991;105:505–512. doi: 10.1017/s0031182000074680. [DOI] [PubMed] [Google Scholar]
- Maule AG, Shaw C, Halton DW, Curry WJ, Thim L. RYIRFamide: A turbellarian FMRFamide-related peptide (FaRP) Regul Pept. 1994;50:37–43. doi: 10.1016/0167-0115(94)90189-9. [DOI] [PubMed] [Google Scholar]
- Maule A, Shaw C, Halton D, Thim L. GNFFRFamide: A novel FMRFamide-immunoreactive peptide isolated from the sheep tapeworm, Moniezia expansa. Biochem Biophys Res Commun. 1993;193:1054–1060. doi: 10.1006/bbrc.1993.1732. [DOI] [PubMed] [Google Scholar]
- Maule AG, Shaw C, Halton DW, Thim L, Johnston CF, Fairweather I, Buchanan KD. Neuropeptide F: A novel parasitic flatworm regulatory peptide from Moniezia expansa (cestoda: Cyclophyllidea) Parasitology. 1991;102:309–316. [Google Scholar]
- McVeigh P, Alexander-Bowman S, Kidd E, Mousley A, Marks NJ, Maule A. Neuropeptide-like protein diversity in phylum nematoda. Int J Parasitol. 2008;38:1493–1503. doi: 10.1016/j.ijpara.2008.05.006. [DOI] [PubMed] [Google Scholar]
- McVeigh P, Kimber MJ, Novozhilova E, Day TA. Neuropeptide signalling systems in flatworms. Parasitology. 2005;131(Suppl):S41–55. doi: 10.1017/S0031182005008851. [DOI] [PubMed] [Google Scholar]
- McVeigh P, Mair GR, Atkinson L, Ladurner P, Zamanian M, Novozhilova E, Marks NJ, Day TA, Maule AG. Discovery of multiple neuropeptide families in the phylum platyhelminthes. Int J Parasitol. 2009;39:1243–1252. doi: 10.1016/j.ijpara.2009.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moneypenny CG, Kreshchenko N, Moffett CL, Halton DW, Day TA, Maule AG. Physiological effects of FMRFamide-related peptides and classical transmitters on dispersed muscle fibres of the turbellarian, Procerodes littoralis. Parasitology. 2001;122:447–455. doi: 10.1017/s0031182001007508. [DOI] [PubMed] [Google Scholar]
- Moneypenny CG, Maule AG, Shaw C, Day TA, Pax RA, Halton DW. Physiological effects of platyhelminth FMRF amide-related peptides (FaRPs) on the motility of the monogenean Diclidophora merlangi. Parasitology. 1997;115:281–288. doi: 10.1017/s0031182097001364. [DOI] [PubMed] [Google Scholar]
- Morales ME, Rinaldi G, Gobert GN, Kines KJ, Tort JF, Brindley PJ. RNA interference of Schistosoma mansoni cathepsin D, the apical enzyme of the hemoglobin proteolysis cascade. Mol Biochem Parasitol. 2008;157:160–168. doi: 10.1016/j.molbiopara.2007.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mousley A, Marks NJ, Halton DW, Geary TG, Thompson DP, Maule AG. Arthropod FMRFamide-related peptides modulate muscle activity in helminths. Int J Parasitol. 2004;34:755–768. doi: 10.1016/j.ijpara.2004.02.005. [DOI] [PubMed] [Google Scholar]
- Ndegwa D, Krautz-Peterson G, Skelly PJ. Protocols for gene silencing in schistosomes. Exp Parasitol. 2007;117:284–291. doi: 10.1016/j.exppara.2007.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omar HH, Humphries JE, Larsen MJ, Kubiak TM, Geary TG, Maule AG, Kimber MJ, Day TA. Identification of a platyhelminth neuropeptide receptor. Int J Parasitol. 2007;37:725–733. doi: 10.1016/j.ijpara.2006.12.019. [DOI] [PubMed] [Google Scholar]
- Pedder SM, Muneoka Y, Walker RJ. Structure-activity and possible mode of action of S-Iamide neuropeptides on identified central neurons of Helix aspersa. Regul Pept. 2001;101:131–140. doi: 10.1016/s0167-0115(01)00279-8. [DOI] [PubMed] [Google Scholar]
- Pereira TC, Pascoal VD, Marchesini RB, Maia IG, Magalhaes LA, Zanotti-Magalhaes EM, Lopes-Cendes I. Schistosoma mansoni: Evaluation of an RNAi-based treatment targeting HGPRTase gene. Exp Parasitol. 2008;118:619–623. doi: 10.1016/j.exppara.2007.11.017. [DOI] [PubMed] [Google Scholar]
- Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29:e45. doi: 10.1093/nar/29.9.e45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sebelová S, Stewart MT, Mousley A, Fried B, Marks NJ, Halton DW. The musculature and associated innervation of adult and intramolluscan stages of Echinostoma caproni (Trematoda) visualised by confocal microscopy. Parasitol Res. 2004;93:196–206. doi: 10.1007/s00436-004-1120-x. [DOI] [PubMed] [Google Scholar]
- Štefanić S, Dvořák J, Horn M, Braschi S, Sojka D, Ruelas DS, Suzuki B, Lim KC, Hopkins SD, McKerrow JH, Caffrey CR. RNA interference in Schistosoma mansoni schistosomula: Selectivity, sensitivity and operation for larger-scale screening. PloS Negl Trop Dis. 2010;4(10):e850. doi: 10.1371/journal.pntd.0000850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi T, McDougall C, Troscianko J, Chen WC, Jayaraman-Nagarajan A, Shimeld SM, Ferrier DE. An EST screen from the annelid Pomatoceros lamarckii reveals patterns of gene loss and gain in animals. BMC Evol Biol. 2009;9:240. doi: 10.1186/1471-2148-9-240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao S, Fernald RD. Comprehensive algorithm for quantitative real-time polymerase chain reaction. J Comput Biol. 2005;12:1047–1064. doi: 10.1089/cmb.2005.12.1047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao ZR, Lei L, Liu M, Zhu SC, Ren CP, Wang XN, Shen JJ. Schistosoma japonicum: Inhibition of mago nashi gene expression by shRNA-mediated RNA interference. Exp Parasitol. 2008;119:379–384. doi: 10.1016/j.exppara.2008.03.015. [DOI] [PubMed] [Google Scholar]