

Abstract
Natural scenes contain a rich variety of contours that the visual system extracts to segregrate the retinal image into perceptually coherent regions. Covert spatial attention helps extract contours by enhancing contrast sensitivity for 1st-order, luminance-defined patterns at attended locations, while reducing sensitivity at unattended locations, relative to neutral attention allocation. However, humans are also sensitive to 2nd-order patterns such as spatial variations of texture, which are predominant in natural scenes and cannot be detected by linear mechanisms. We assess whether and how exogenous attention—the involuntary and transient capture of spatial attention—affects the contrast sensitivity of channels sensitive to 2nd-order, texture-defined patterns. Using 2nd-order, texture-defined stimuli, we demonstrate that exogenous attention increases 2nd-order contrast sensitivity at the attended location, while decreasing it at unattended locations, relative to a neutral condition. By manipulating both 1st- and 2nd-order spatial frequency, we find that the effects of attention depend both on 2nd-order spatial frequency of the stimulus and the observer’s 2nd-order spatial resolution at the target location. At parafoveal locations, attention enhances 2nd-order contrast sensitivity to high, but not to low 2nd-order spatial frequencies; at peripheral locations attention also enhances sensitivity to low 2nd-order spatial frequencies. Control experiments rule out the possibility that these effects might be due to an increase in contrast sensitivity at the 1st-order stage of visual processing. Thus, exogenous attention affects 2nd-order contrast sensitivity at both attended and unattended locations.
Keywords: Attention, texture, second-order processing, contrast sensitivity
Introduction
To efficiently guide perception and behavior, the visual system must extract contours and parse the retinal image into perceptually coherent regions. Consider the natural scene presented in Fig. 1A. Many regions in this picture are bounded by luminance-defined contours such as the borders of the two people on the walkway. Much of the early visual system from the retina to primary visual cortex is dedicated to detecting such changes in luminance (DeValois & DeValois, 1988). Linear spatial filters that are selective for spatial frequency and orientation, akin to simple cells in V1, are effective for signaling such contours (Graham, 1989). However, humans are also sensitive to changes in visual attributes other than luminance, which cannot be detected by linear mechanisms. For example, the different sections of the walkway differ in the local orientation of the wooden slats (that is, by textural attributes), but the average luminance is relatively constant across the different sections. As a result, large-scale linear filters tuned to luminance variations are ineffective for segregating the walkway into different sections. Patterns defined by changes in textural attributes (e.g., local orientation, contrast, and spatial frequency) that are not visible to linear filters are commonly referred to as 2nd-order patterns, to distinguish them from 1st-order, luminance-defined patterns. Texture information is vital for segmenting the retinal image into distinct regions as an initial step in object recognition. Successfully segmenting an image, however, is a computationally expensive process.
Figure 1.
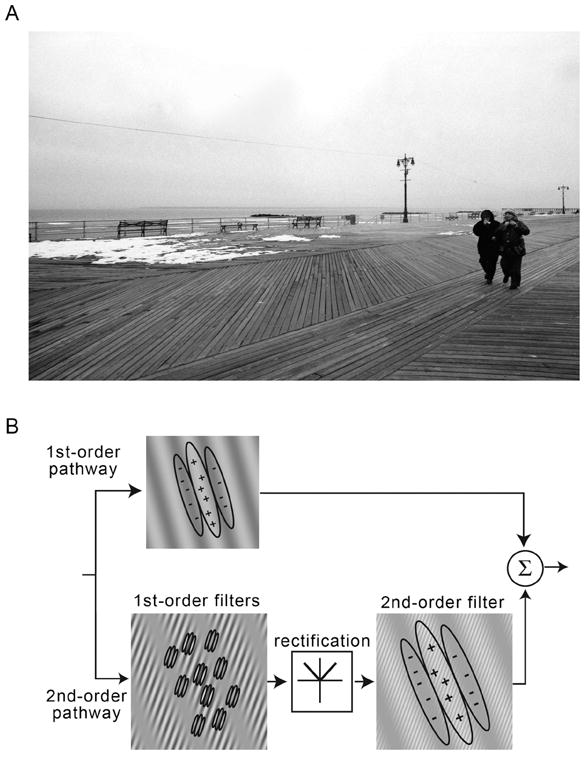
(A) Natural scene containing 1st- and 2nd-order contours. The boundary between the two pedestrians and walkway in the lower right is defined by a change in luminance, hence it is 1st-order. The texture-defined boundaries between the different regions of the walkway are 2nd-order. (B) A typical model of visual processing with parallel pathways for 1st- and 2nd-order stimuli. In the top path, luminance-defined stimuli are signaled by a linear filter. The bottom path is a filter-rectify-filter cascade sensitive to 2nd-order stimuli. Both 1st- and 2nd-order information are combined in a later decision stage (Baker & Mareschal, 2001).
The high metabolic cost of neuronal activity involved in cortical computation renders it impossible to process the overwhelming amount of information arriving at our retinae (Lennie, 2003). Covert spatial attention enables us to manage limited resources by selecting a relevant location or aspect of the visual scene and prioritizing its processing even without directing the eyes to that location (Posner, 1980). Attention affects 1st-order processing, improving behavioral performance in various tasks (see Carrasco, 2006, for a review) and enhancing neural processing of sensory information (see Reynolds & Chelazzi, 2004, for a review). Some of these changes are mediated by an increase in contrast sensitivity of 1st-order linear spatial filters at the attended location and a decrease at unattended locations (Pestilli & Carrasco, 2005; Pestilli, Viera, & Carrasco, 2007). However, little is known about the effects of attention on the processing of 2nd-order texture patterns. Texture perception is usually considered to be pre-attentive (Braun & Sagi, 1990; Julesz, 1981; Schubo & Meinecke, 2007). However, many studies have shown an effect of attention on performance with texture stimuli (Casco, Grieco, Campana, Corvino, & Caputo, 2005; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000, 2008; Yeshurun, Montagna, & Carrasco, 2008).
In this study, we investigate whether and how exogenous attention—the involuntary and transient capture of attention to a location by a peripheral uninformative visual cue—affects contrast sensitivity for 2nd-order patterns at both attended and unattended locations.
Visual perception of 2nd-order texture patterns
Texture patterns—homogenous regions of repeated structures that cannot be detected by linear mechanisms—are predominant in natural scenes (Johnson & Baker, 2004; Schofield, 2000). Detecting a boundary between two texture regions is analogous to detecting a 1st-order, luminance-defined edge (Nothdurft, 1985). Unlike locating a change in luminance (i.e., a typical light/dark edge), finding a 2nd-order texture boundary requires locating changes in local textural properties (e.g., contrast, orientation, or scale) of an underlying fine-scale pattern, which is often referred to as the 1st-order carrier. Because the average luminance may be the same on either side of the texture boundary, 2nd-order edges cannot be detected by a linear mechanism such as those used to detect 1st-order, luminance-defined edges.
Several lines of evidence suggest that 1st- and 2nd-order pattern processing require different neural mechanisms (Ellemberg, Allen, & Hess, 2004; Larsson, Landy, & Heeger, 2006; Morgan, Mason, & Baldassi, 2000; Schofield & Georgeson, 1999; Scott-Samuel & Georgeson, 1999). Like their 1st-order counterparts, 2nd-order channels are tuned for orientation (Arsenault, Wilkinson, & Kingdom, 1999; Dakin, Williams, & Hess, 1999; Graham & Wolfson, 2001) and spatial frequency (Landy & Oruç, 2002; Scott-Samuel & Georgeson, 1999), but have wider bandwidths (Landy & Oruç, 2002). Although the contrast sensitivity function (CSF) for 1st-order patterns is bandpass, peaking for mid-range spatial frequencies (Robson & Graham, 1981), the 2nd-order CSF has been shown to be essentially flat (Landy & Oruç, 2002).
A number of investigators have described edge-based texture segregation models to account for texture segregation performance (Bergen & Adelson, 1988; Graham, 1991; Graham, Beck, & Sutter, 1992; Landy & Bergen, 1991; Malik & Perona, 1990; Sutter, Beck, & Graham, 1989), see Landy and Graham (2004) for a review. A typical model of 2nd-order visual processing (Fig. 1B) comprises two layers of bandpass spatial linear filters separated by a point-wise nonlinearity. The first spatial filter is tuned for orientation and spatial frequency, thus responding strongly to one of the carrier patterns. The output of this first stage is rectified so that spatial regions with large response variability (i.e., regions with large 1st-order linear-filter response, both positive and negative) are mapped to large average response (Bergen & Adelson, 1988). Finally, a second set of larger-scale linear filters performs a more global analysis of the rectified outputs of the 1st-order filters. Appropriately tuned 2nd-order filters will respond robustly to boundaries between regions with different average response strength. Such models of 2nd-order processing are referred to as filter-rectify-filter (FRF), linear-nonlinear-linear (LNL), or “back-pocket” models (Landy & Graham, 2004).
Although originally developed to explain psychophysical data, these models provide an architecture that maps well onto the cortical visual processing cascade. The physiological substrate for the first stage linear filters is likely to correspond to simple cells in area V1 that are selective for spatial frequency and orientation (Graham, 1989). The intermediate point-wise nonlinearity (i.e., rectification) might correspond to the neuronal spiking threshold of 1st-order neurons. Finally, 2nd-order, texture-selective neurons that represent the second stage of filtering are likely to be located in higher extrastriate areas beyond V1 (Baker & Mareschal, 2001; Landy & Graham, 2004). fMRI studies have shown that responses to 2nd-order texture boundaries (Kastner, De Weerd, & Ungerleider, 2000) and 2nd-order orientation-selective adaptation effects (Larsson et al., 2006) are stronger in downstream visual areas, providing support for the notion that 2nd-order processing takes place subsequent to 1st-order filtering.
Effects of covert attention at the 1st-order stage of visual processing
Spatial attention can be allocated overtly, by directing one’s gaze towards a position within the visual scene, or covertly, by attending to an area in the periphery without actually directing gaze towards it, allowing one to selectively process information at a given location in the absence of eye movements (Posner, 1980). Covert spatial attention can be further divided into two types: endogenous and exogenous, which follow different time courses and are triggered by different cues. Endogenous attention is voluntary, conceptually driven, and has a sustained effect, which takes about 300 ms to be deployed and can last several seconds. In contrast, exogenous attention is involuntary, driven by a briefly displayed peripheral cue, and has a transient effect that peaks at about 100 ms and decays shortly thereafter (Cheal & Lyon, 1991; Jonides & Irwin, 1981; Ling & Carrasco, 2006a; Liu, Stevens, & Carrasco, 2007; Muller & Findlay, 1988; Muller & Rabbitt, 1989; Nakayama & Mackeben, 1989; Posner, 1980; Remington, Johnston, & Yantis, 1992). The involuntary transient shift of exogenous attention occurs even when the cues are uninformative (Giordano, McElree, & Carrasco, 2009; Montagna, Pestilli, & Carrasco, 2009; Muller & Rabbitt, 1989; Pestilli & Carrasco, 2005; Pestilli et al., 2007; Posner, 1980) and can impair as well as improve performance (Carrasco, Loula, & Ho, 2006; Talgar & Carrasco, 2002; Yeshurun, 2004; Yeshurun & Carrasco, 1998, 2000; Yeshurun et al., 2008). In this study, we focus on the effects of exogenous attention, the involuntary component of covert spatial attention, on 2nd-order contrast sensitivity.
Many behavioral and neurophysiological studies indicate that exogenous attention improves performance on detection, discrimination, and localization tasks that are primarily limited by stimulus contrast (see Carrasco, 2006; Reynolds & Chelazzi, 2004, for reviews). Compared to a neutral condition, attention enhances contrast sensitivity for 1st-order, luminance-defined patterns at the attended location and decreases sensitivity at unattended locations (Cameron, Tai, & Carrasco, 2002; Carrasco, Penpeci-Talgar, & Eckstein, 2000; Dosher & Lu, 2000; Ling & Carrasco, 2006b; Lu & Dosher, 1998; Pestilli & Carrasco, 2005; Pestilli, Ling, & Carrasco, 2009). These effects of attention on 1st-order processing occur across a wide range of spatial frequencies (Carrasco et al., 2000) and occur early in the visual stream (e.g., Brefczynski & DeYoe, 1999; Gandhi, Heeger, & Boynton, 1999; Herrmann, Montaser-Kouhsari, Carrasco, & Heeger, 2010; Huk & Heeger, 2000; Liu, Pestilli, & Carrasco, 2005; McAdams & Reid, 2005; Reynolds & Chelazzi, 2004; Störmer, McDonald, & Hillyard, 2009). Moreover, many other aspects of 1st-order processing are affected by exogenous attention including spatial resolution (Carrasco & Yeshurun, 2009; Yeshurun & Carrasco, 1998, 1999, 2000), acuity (Carrasco, Williams, & Yeshurun, 2002; Montagna et al., 2009), speed of information accrual (Carrasco, Giordano, & McElree, 2004, 2006; Carrasco & McElree, 2001) and stimulus appearance (Anton-Erxleben, Abrams, & Carrasco, 2010; Anton-Erxleben, Henrich, & Treue, 2007; Carrasco, Ling, & Read, 2004; Fuller & Carrasco, 2006; Gobell & Carrasco, 2005).
Effects of attention at the 2nd-order stage of visual processing
Although selective attention and texture processing have been widely investigated in the past, very few studies have addressed the influence of attention on the processing of 2nd-order patterns. Much of the literature assumes that texture segregation is truly effortless and immediate (Braun & Sagi, 1990; Julesz, 1981; Schubo & Meinecke, 2007). There is a debate regarding whether selective attention is necessary for and whether and how it modifies texture segregation. Consistent with the existence of a pre-attentive texture-segmentation mechanism, fMRI studies have shown that texture segmentation occurs in the absence of perceptual awareness (Kastner et al., 2000; Schira, Fahle, Donner, Kraft, & Brandt, 2004). Although there is some evidence that texture segregation occurs without attention, some textures require substantial time to be processed (Sutter & Graham, 1995), undermining the notion that texture segregation is immediate and effortless. Furthermore, orientation discrimination based on grouping of elements along a texture boundary has been shown to require attention (Casco et al., 2005) and EEG/MEG studies revealed that while texture processing shows some degree of automaticity, it also depends on attentional resources (Appelbaum & Norcia, 2009; Schubo, Meinecke, & Schroger, 2001). These studies, however, manipulated attention by having observers perform tasks on the texture stimuli or not (i.e., passive viewing compared to active processing of texture stimuli), rather than manipulating spatial attention selectively, which makes it difficult to interpret how attention affects texture perception at attended and unattended locations.
Very few studies have systematically manipulated spatial attention to investigate the effects of attention on the processing of 2nd-order patterns. Two studies have shown that endogenous attention affects sensitivity to 2nd-order motion patterns (Allen & Ledgeway, 2003; Lu, Liu, & Dosher, 2000). Detection of 2nd-order motion is based on a filter-rectify-filter mechanism similar to 2nd-order static texture processing (Fig. 1B). However, evidence has shown that these two types of 2nd-order information are detected and represented by separate mechanisms localized in distinct cortical areas (Baker, 1999; Baker & Mareschal, 2001; Larsson et al., 2006). Only one study has directly assessed the effects of exogenous attention on texture segmentation. Using textures composed of narrow-band stimuli to stimulate either 1st- or 2nd-order filters of various scales, Yeshurun and Carrasco (2000) showed that attentional effects on spatial resolution depend on 2nd-order spatial frequency, suggesting that exogenous attention can operate at the second stage of linear filtering. Whether exogenous attention is able to affect the contrast sensitivity of 2nd-order filters is, however, an open question that has not been directly investigated, and is independent of the previous findings concerning spatial resolution.
In the present study, we examine whether and how attention affects contrast sensitivity for 2nd-order, texture-defined patterns at both attended and unattended locations. Observers performed an orientation-discrimination task using 2nd-order, orientation-defined stimuli. We used a classic exogenous cueing paradigm as has been used previously to study the effects of exogenous attention on 1st-order contrast sensitivity (e.g. Carrasco et al., 2000; Pestilli & Carrasco, 2005; Pestilli et al., 2007).
Given that exogenous attention affects spatial resolution depending on the 2nd-order spatial frequency content, regardless of the 1st-order spatial frequency content (Yeshurun & Carrasco, 2000), we measured the effects of exogenous attention on contrast sensitivity for 2nd-order patterns while independently varying both the 1st- and 2nd-order spatial frequency content of the stimuli. Whereas the attentional effects on 2nd-order contrast sensitivity (if any) should not depend on the 1st-order content, they may differ when the 2nd-order spatial frequency is varied.
Experiment 1
Method
Participants
Five graduate students and postdoctoral fellows (3 females) at New York University participated as observers in Experiment 1. Four were experienced psychophysical observers, but all (except one author) were naive as to the purposes of the experiment. All participants had normal or corrected-to-normal vision. The Institutional Review Board at New York University approved the experimental procedures and all participants gave informed consent.
Apparatus
Stimuli were generated using Matlab (MathWorks, Natick, MA) and MGL (http://justingardner.net/mgl) and were displayed on a 21” CRT monitor (1600×1200 at 100 Hz). The display was calibrated using a Photo Research (Chatworth, CA) PR650 SpectraColorimeter to produce linearized lookup tables for the experiment. Eye position was monitored using an infrared video camera system (ISCAN, Burlington, MA) to ensure that all observers were able to maintain steady fixation.
Stimuli
Texture patterns were constructed by spatially modulating two orthogonal gratings (1st-order carriers C1 and C2 oriented ±45°) using a second horizontal or vertical grating with lower spatial frequency (2nd-order modulator M). The resulting stimulus was defined as follows (Fig. 2A):
| (1) |
where L0 is the mean luminance of the display and E is a circular stimulus envelope with raised-cosine edges (4 deg diam., raised-cosine width: 0.2 deg). There was a fixed orientation difference of 45° between the modulator and each of the carriers. For each stimulus, the value of C was set to yield the desired peak luminance contrast for each experiment (70% contrast for Experiments 1 and 2). 2nd-order modulator contrast ranged from 8 to 96%. There were four different texture conditions formed by combining two carrier spatial frequencies (‘low’: 2 cycle/deg; ‘high’: 4 cycle/deg) and two modulator spatial frequencies (‘low’: 0.5 cycle/deg; ‘high’: 1 cycle/deg), designated as LL, HL, LH and HH (Fig. 2B). Note that frequencies that are an octave away activate different channels (Graham, 1989; Landy & Oruç, 2002). Similar patterns have been used in psychophysical and neuroimaging studies of texture segmentation (Landy & Oruç, 2002; Larsson et al., 2006).
Figure 2.
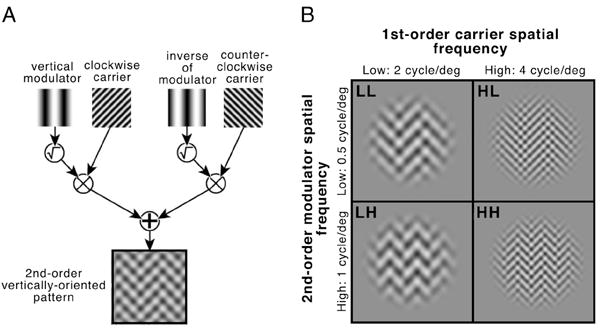
(A) Stimulus construction. Texture patterns were computed by modulating two orthogonal luminance gratings (the carriers, with ±45° orientations) with a third vertical or horizontal modulator grating of lower spatial frequency. A fixed orientation difference of ±45° separated the carriers and modulator. The contrast-modulated carrier patterns were summed, and the result was multiplied by a circular window with raised-cosine smoothed edges. Mean luminance was constant across the stimulus. (B) Stimulus conditions. Two spatial frequencies were used for both the 1st-order carrier (low/L and high/H: 2 and 4 cycle/deg) and 2nd-order modulator (low/L and high/H: 0.5 and 1 cycle/deg), resulting in four texture conditions: LL, HL, LH, and HH.
Procedure
Fig. 3 depicts the trial sequence. Each trial began with a 300 ms fixation cross at the center of the screen. Observers were instructed to fixate the cross throughout each trial. Next, exogenous attention was manipulated via peripheral cues preceding stimulus presentation. The advantage of using an exogenous cue is that it results in a highly controlled manipulation of attention, not dependent on the observer’s strategy (Giordano et al., 2009; Montagna et al., 2009; Nakayama & Mackeben, 1989; Pestilli & Carrasco, 2005; Pestilli et al., 2007). On 2/3 of the trials (cued trials), one black rectangle (width: 0.3 deg, length 1.6 deg, orientation: 45°) was flashed for 60 ms 3.5 deg from one stimulus location (i.e., 1.5 deg border-to-border from the upcoming stimulus). On the remaining 1/3 of the trials (neutral trials), four black rectangles were presented next to the four possible stimulus locations, distributing observers’ exogenous attention across space. After an inter-stimulus-interval (ISI) of 40ms, two 2nd-order texture stimuli were presented simultaneously for 100 ms at two of the four possible locations. Orientation and phase of the 2nd-order modulator were independently randomized across trials for each 2nd-order stimulus in a pair. Goal-directed saccades require about 250 ms (Leigh & Zee, 1991; Mayfrank, Kimmig, & Fischer, 1987). Thus, no eye movements to the stimulus could occur between precue onset and stimulus onset (100 ms). 100 ms after stimulus display, a white response cue was presented near the fixation cross, pointing at one of the two stimuli (the target). Response cues considerably reduce location uncertainty by indicating the exact target location (e.g, Kinchla, Chen, & Evert, 1995; Lu & Dosher, 2004; Luck et al., 1994; Pestilli & Carrasco, 2005; Yeshurun et al., 2008).
Figure 3.
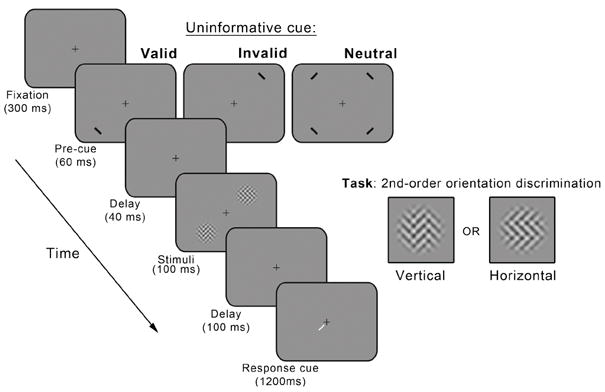
Trial sequence. The trial sequence was the same for Experiments 1 and 2. For Experiment 3, only neutral trials were used.
On cued trials, one stimulus was located at the cued location, and the other was equally likely to be in any of the remaining three locations. Observers were told that the precue was uninformative and that it would not be advantageous to move their eyes to the cued location: the target was equally likely to be the cued (a valid trial) or uncued (an invalid trial) stimulus, and the precue gave no information about target orientation; the precue only indicated stimulus onset, as was the case in the neutral trials (e.g, Lu & Dosher, 2004; Lu et al., 2000; Montagna et al., 2009; Pestilli & Carrasco, 2005; Pestilli et al., 2007). Given that the peripheral cue automatically draws attention to the cued location (Giordano et al., 2009; Jonides & Yantis, 1988; Yantis & Jonides, 1984), even an uninformative cue should affect performance.
The task was a two-alternative, forced-choice, 2nd-order-orientation-discrimination task. Observers were instructed to report the orientation (vertical or horizontal) of the 2nd-order sine wave grating presented at the location indicated by the response cue. Observers were required to respond within 1200 ms after the onset of the response cue by pressing one of two possible keys. Auditory feedback was given. About 2,530 trials per stimulus condition (LL, HL, LH and HH) were collected for each observer in 16 blocks (in 4 experimental sessions)—an equal number of trials for each of 9 2nd-order modulator contrast levels—for a total of ~10,120 experimental trials per observer. The order of the sessions was counterbalanced across stimulus conditions.
The relation between the peripheral precue and the response cue defined the validity of the cue (Downing, 1988; Lu & Dosher, 2004; Luck et al., 1994; Montagna et al., 2009; Pestilli & Carrasco, 2005). The use of a non-predictive precue allows us to isolate the automatic orienting of attention. On valid trials, the location indicated by the precue and the response cue matched, and observers reported the orientation of the cued (‘attended’) stimulus. On invalid trials, the precue and response cue did not match, and observers reported the orientation of the uncued (‘unattended’) stimulus. On neutral trials, all four potential stimulus locations were precued. The spatiotemporal characteristics of the response cue were identical across the three cueing-conditions (valid, invalid, and neutral).
If attention increases 2nd-order contrast sensitivity at the attended location and decreases it at unattended locations, it should improve and worsen performance, respectively, because varying 2nd-order modulator contrast alters performance in this orientation-discrimination task.
Analysis
For each observer, performance was assessed separately for each combination of stimulus condition (LL, LH, HL and HH), 2nd-order modulation contrast and cueing condition (valid, invalid and neutral). We use signal detection theory, treating the vertical 2nd-order stimulus as the signal-present and horizontal 2nd-order stimulus as the signal-absent trials. Performance was evaluated as d′ = z(hit rate) - z(false alarm rate). d′ values were averaged over observers. The data were fit with a Naka-Rushton function:
| (2) |
using a least-squares criterion, where d′(c) represents performance as a function of contrast, d′max is the asymptotic performance at high contrast values, c50 is the contrast at which the observer achieves half the asymptotic performance, and n determines the slope of the psychometric function. For each stimulus condition (e.g., LH), we simultaneously fit the data from the three attention conditions, allowing distinct values of d′max, c50 and n for each attention condition. Confidence intervals and p-values were computed by bootstrapping. Specifically, individual psychophysical trials were randomly resampled with replacement to generate resampled data sets, which were refitted using the same procedure. We repeated this procedure of resampling and refitting 10,000 times to generate bootstrap distributions of the psychometric data and of the fitted parameters. Confidence intervals for the fitted parameters and p-values were extracted from these bootstrap distributions to test if there was a benefit for valid and a cost for invalid, compared to neutral cues either in d′max or c50. Specifically, we used the bootstrap distribution of the differences between the conditions (e.g., valid minus invalid trials) to determine the percentage of the values in the tail of the distribution of the differences greater than zero for changes in d′max, or lower than zero for changes in c50. The use of one-tailed statistical tests is based on previous studies, reporting a benefit for valid and a cost for invalid, compared to neutral cues (Carrasco, 2006; Giordano et al., 2009; Herrmann et al., 2010; Ling & Carrasco, 2006b; Pestilli & Carrasco, 2005; Pestilli et al., 2009; Pestilli et al., 2007).
Results
Fig. 4 shows psychometric functions averaged across observers for each of the four texture conditions. The effects of exogenous attention depended on the spatial frequency of the 2nd-order modulator. When the spatial frequency of the 2nd-order modulator was low (Fig. 4A-B), the three psychometric functions (valid, neutral and invalid) were indistinguishable and no effect of cueing was observed on d′max (pvalid-invalid > 0.5 in both cases), c50 (pvalid-invalid > 0.5 in both cases) or slope (n) (pvalid-invalid > 0.1 in both cases). However, when the spatial frequency of the 2nd-order modulator was high (Fig. 4C-D), performance was consistent with response gain, i.e., attention increased the value of d′max. Compared to the neutral condition, a valid cue to the target enhanced 2nd-order contrast sensitivity, as indicated by an increase in d′max (LH: p < 0.005; HH: p < 0.05), whereas an invalid cue decreased sensitivity, reducing d′max (LH: p < 0.05; HH: p < 0.001). No change of c50 (pvalid-invalid > 0.5 in both cases) or slope (n) (pvalid-invalid > 0.5 in both cases) was observed.
Figure 4.
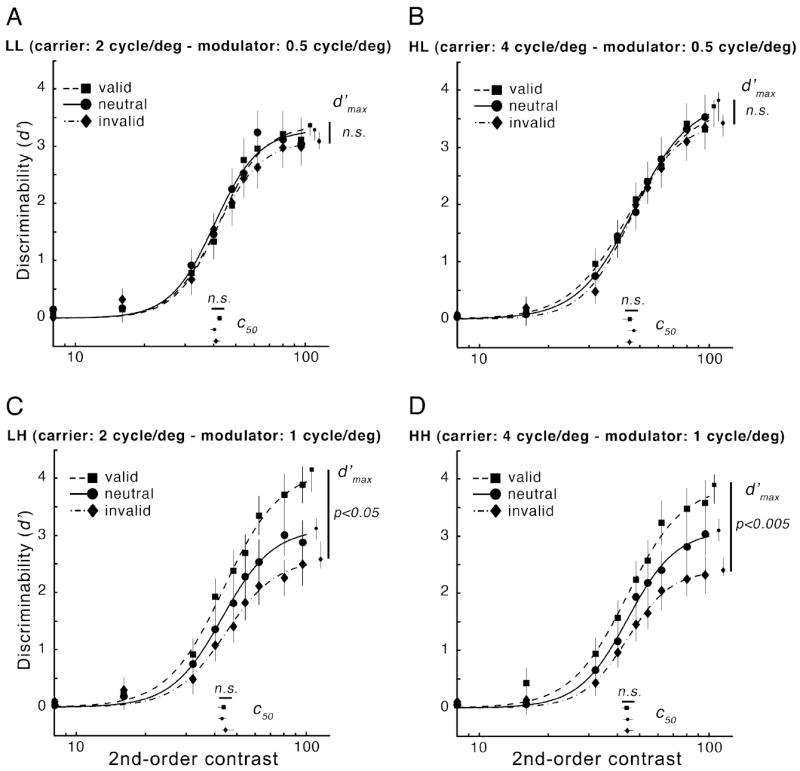
Experiment 1: Effects of exogenous attention on performance (d′) as a function of 2nd-order modulator contrast for the four different texture conditions at 5 deg eccentricity (A: LL; B: HL; C: LH; D: HH; see Fig. 2). Each panel shows psychometric functions and parameter estimates (c50: 2nd-order contrast yielding half-maximum performance; d′max: asymptotic performance) for each cueing condition (valid, neutral and invalid). Each data point represents the mean across observers (n=5). Error bars correspond to ±1 SEM for data points and 68%-confidence intervals obtained by bootstrapping for parameter estimates.
Fig. 5 shows the values of d′max and c50 for individual observers in the valid and invalid attention conditions in which each value is normalized by (i.e., divided by) the corresponding parameter values from the neutral condition. The effect of attention on 2nd-order contrast sensitivity is consistent across observers. For the stimulus conditions with high 2nd-order spatial frequency (Fig. 5C-D), all of the valid d′max values were higher than the neutral d′max values (i.e., the squares fell above the unity line), and all invalid d′max values were lower than the neutral d′max values (i.e., the diamonds fell below the unity line). However, for the two conditions with low 2nd-order spatial frequency conditions there was no clear pattern across valid and invalid conditions, reflecting the absence of attentional modulation.
Figure 5.
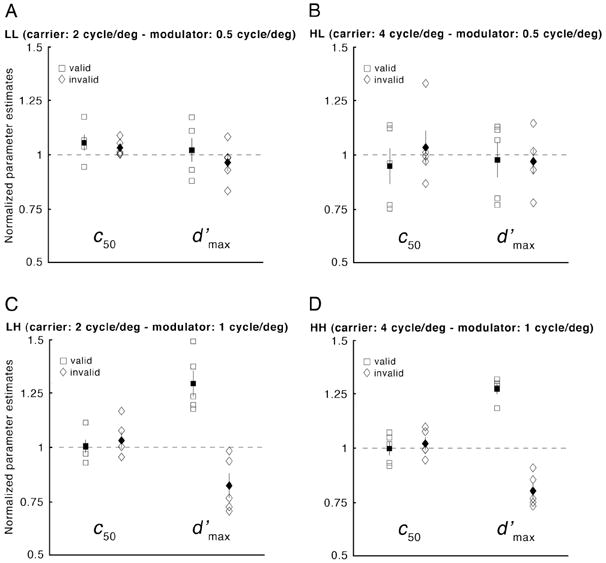
Experiment 1: Effects of exogenous attention on individual observers’ normalized parameter estimates (c50 and d′max; A: LL; B: HL; C: LH; D: HH; see Fig. 2). Each plot displays individual observers’ parameter estimates in the valid (open squares) and invalid (open diamonds) cue conditions normalized by the corresponding values in the neutral-cue condition. Filled symbols indicate mean across observers (±1 SEM).
To characterize how exogenous attention affects 2nd-order contrast sensitivity, we used nested hypothesis tests to compare the full model fit with a pure response gain (c50 parameter constant across attention conditions) and a pure contrast gain (d′max parameter constant across attention conditions). The slope (n) parameter was free to vary across the 3 attention conditions. The full model fits better than one with d′max constrained (for both LH and HH: nested hypothesis test, p < 0.001) but not than one with c50 constrained (for both LH and HH: nested hypothesis test, p > 0.5). This finding supports the conclusion that the effects of exogenous attention on contrast sensitivity for high 2nd-order spatial frequencies are better explained with a response-gain mechanism.
Discussion
The present results indicate that exogenous attention affects contrast sensitivity for 2nd-order, orientation-defined patterns in a manner that depends on 2nd-order spatial frequency. When 2nd-order modulator spatial frequency was high (1 cycle/deg), exogenous attention robustly improved asymptotic performance at high contrasts (d′max), consistent with a response-gain mechanism, with no evidence for contrast gain (i.e., changes in c50). These two types of attentional gain-control mechanisms, contrast gain and response gain, have been considered previously for responses to 1st-order luminance-modulated stimuli (Ling & Carrasco, 2006a; Pestilli et al., 2009). Some studies indicate that exogenous attention affects 1st-order contrast sensitivity mainly via response gain (Ling & Carrasco, 2006a; Pestilli et al., 2009; Pestilli et al., 2007). Our results indicate that the mechanisms of exogenous attention on contrast sensitivity are similar at the 1st- and 2nd-order stages of visual processing.
The absence of attentional effects for patterns with low 2nd-order spatial frequency is intriguing. For 1st-order patterns, exogenous attention has been shown to improve performance at all spatial frequencies tested, with no systematic interaction between the effect of attention and the spatial frequency of the stimulus (e.g., Carrasco et al., 2000). However, our results suggest that exogenous attention affects only the sensitivity of 2nd-order channels tuned to high spatial frequency, without affecting the population tuned to lower 2nd-order spatial frequency. One possible explanation for this is that exogenous attention may only affect the sensitivity of channels tuned to high 2nd-order spatial frequency. This pattern of attentional effects would indicate a difference between the mechanisms of exogenous attention at the 1st- and 2nd-order processing stages.
Alternatively, perhaps the absence of an effect of exogenous attention on contrast sensitivity for low 2nd-order spatial frequencies was specific to the parafoveal location (5 deg eccentricity) used in this experiment. Counter-intuitively, exogenous attention does not benefit performance across all eccentricities in texture segmentation tasks. Attention instead improves performance where resolution is low (i.e., at peripheral locations) and hinders performance where resolution is already high (i.e., at central locations). For instance, Yeshurun and Carrasco (2000) found that in a texture segmentation task, the range of eccentricities in which exogenous attention improved or impaired performance depended on the scale of the 2nd-order pattern as well as the average size of the filters at a given eccentricity. Specifically, performance was impaired for a larger range of eccentricities for textures with low than with high 2nd-order spatial frequency. This finding supports the resolution hypothesis, which postulates that exogenous attention increases spatial resolution at the attended location, even when it impairs performance (Carrasco, Loula et al., 2006; Carrasco & Yeshurun, 2009; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000; Yeshurun et al., 2008). One mechanism by which exogenous attention may enhance spatial resolution is by increasing the sensitivity of the smallest possible filters at the attended area, reweighting the population response in favor of higher-spatial-frequency filters (Balz & Hock, 1997; Carrasco, Loula et al., 2006; Carrasco et al., 2002; Carrasco & Yeshurun, 2009). The absence of attentional effects for the low 2nd-order spatial frequency patterns in Experiment 1 may reflect the fact that exogenous attention enhances contrast sensitivity at the 2nd-order stage of filtering by increasing the sensitivity of the smallest filters at the attended area. As a result, we could expect attentional effects that depend on the scale of the 2nd-order pattern and the average size of the filters at the attended area.
Physiological and behavioral studies indicate that there are more cells tuned to high than low 1st-order spatial frequencies at the fovea (Robson & Graham, 1981) and that the ratio of the number of neurons tuned to high vs. low spatial frequencies decreases with eccentricity (Azzopardi, Jones, & Cowey, 1999; DeValois & DeValois, 1988). The number of 2nd-order spatial frequency channels at the fovea is the same as for 1st-order channels for spatial frequencies up to about 2 cycle/deg, but they are tuned to lower spatial frequencies, with fewer channels tuned to higher spatial frequencies relative to 1st-order (Ellemberg, Allen, & Hess, 2006). Importantly, sensitivity for 1st- and 2nd-order stimuli show a similar spatial frequency-dependent fall-off with eccentricity (Hess, Baker, May, & Wang, 2008).
At the parafoveal location we used, an increase in response of the smallest 2nd-order filters may have resulted in improved contrast sensitivity for patterns with high, but not with low, 2nd-order spatial frequency. Correspondingly, we would expect a benefit of attention on contrast sensitivity for patterns with low 2nd-order spatial frequency located farther in the periphery where the average size of the filters is larger.
Experiment 2
In Experiment 2, we tested the hypothesis that attention had no effect for low second-order frequencies due to the parafoveal location used in Experiment 1. To do so, we measured the effects of exogenous attention on contrast sensitivity for patterns with low 2nd-order spatial frequency at peripheral (10 deg eccentricity) rather than parafoveal (5 deg eccentricity) locations.
Method
Participants, stimuli and procedure were the same as in Experiment 1 except for the following: (1) Only the two spatial-frequency conditions were tested for which no effect of attention on 2nd-order contrast sensitivity was observed in Experiment 1 (LL and HL). (2) Stimulus eccentricity was increased from 5 to 10 deg. (3) To ensure that the precue was still able to efficiently capture observers’ attention, precue size was increased (width .45 deg, length 2.5 deg eccentricity). (4) The distance between the precue and the border of the stimulus was increased from 1.5 to 3 deg to avoid crowding due to the greater eccentricity and larger precue. Each observer completed a total of 5,184 experimental trials in 16 blocks (in 4 experimental sessions), with 2,592 trials per stimulus condition. Data were analyzed using the same procedure as for Experiment 1.
Results and discussion
As shown in Fig. 6, at 10 deg eccentricity, we now find reliable effects of attention for patterns with low 2nd-order spatial frequency. Compared with the neutral condition, the psychometric function for the valid condition is shifted to the left, whereas in the invalid condition it is shifted to the right. Exogenous attention altered performance primarily for intermediate contrasts, leading to robust differences in c50, consistent with a change of contrast gain (LL and HL: pvalid-invalid < 0.05). No significant change in asymptotic performances (d′max) was observed (LL: pvalid-invalid = 0.08; HL: pvalid-invalid > 0.5), which may have resulted from the fact that for some observers, the psychometric functions did not reach asymptote within the available contrast range. The pattern of results for c50 and d′max was consistent across observers (Fig. 7). All (but one for the HL condition) of the valid c50 values (squares) were lower than the neutral c50 values, and all (but two for the LL condition) invalid c50 values (diamonds) were higher than the neutral c50 values. Furthermore, nested hypothesis tests indicated that the full model did not fit better than one with d′max constrained (nested hypothesis tests, p > 0.1 in both cases) nor with c50 constrained (nested hypothesis tests, p > 0.1 in both cases). This finding suggests that the effects of attention in Experiment 2 are explained by either a contrast- or a response-gain mechanism, or a combination of both.
Figure 6.
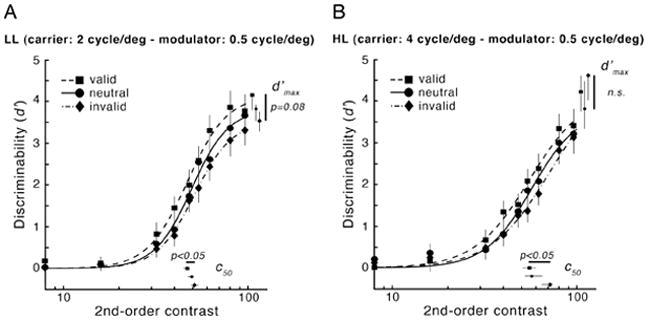
Experiment 2: Effects of exogenous attention on performance (d′) as a function of 2nd-order modulator contrast for the two low 2nd-order modulator spatial frequency texture conditions at 10 deg eccentricity (A: LL; B: HL). Both panels plot psychometric functions and parameter estimates (c50 and d′max) for each cueing condition (valid, neutral and invalid). Plotting conventions as in Fig. 4.
Figure 7.
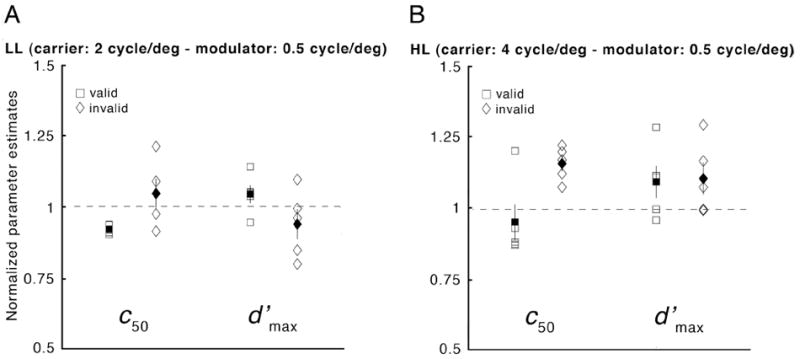
Experiment 2: Effects of exogenous attention on individual observers’ normalized parameter estimates (c50 and d′max; A: LL; B: HL). Each plot displays individual observers’ parameter estimates in the valid (open squares) and invalid (open diamonds) cue conditions normalized by the corresponding values in the neutral-cue condition. Filled symbols indicate mean across observers (±1 SEM).
These results indicate that exogenous attention can affect 2nd-order contrast sensitivity for stimuli with low 2nd-order spatial frequency when the stimuli appear in the periphery (10 deg eccentricity). This finding supports the hypothesis that the absence of attentional effects for patterns with low 2nd-order spatial frequency in Experiment 1 was due to the effects of attention on spatial resolution at parafoveal locations (5 deg eccentricity). In sum, from Experiments 1 and 2 we conclude that exogenous attention affects contrast sensitivity at the 2nd-order stage for both patterns with high and low spatial frequency in a manner that depends on the spatial scale and the eccentricity of the texture.
Experiment 3
Experiments 1 and 2 revealed that exogenous attention improves texture discrimination at the attended location and impairs it at unattended locations. Because we used 2nd-order stimuli that cannot be detected by spatial filters sensitive to 1st-order luminance signals, this finding suggests that exogenous attention affects the sensitivity of linear filters at the 2nd-stage of visual processing directly. However, 2nd-order sensitivity can increase with 1st-order contrast at both central and peripheral locations (Hess et al., 2008), and exogenous attention can increase contrast sensitivity of filters at the 1st-order stage of visual processing (e.g., Dosher & Lu, 2000; Herrmann et al., 2010; Ling & Carrasco, 2006b; Lu & Dosher, 1998; Pestilli & Carrasco, 2005; Pestilli et al., 2009). Thus, it is important to rule out the possibility that our results might reflect the indirect consequence of an increase in sensitivity at the 1st-order stage of visual processing.
In this experiment we examined directly whether an increase in effective 1st-order contrast—similar to an increase in 1st-order contrast sensitivity with attention—could explain the results observed with attention in Experiments 1 and 2. We varied 1st-order contrast without cueing attention to a particular location to determine whether this caused shifts of the 2nd-order contrast sensitivity functions similar to those observed in the previous experiments.
Method
Participants, stimuli and procedure were the same as in Experiments 1 and 2 except for the following: (1) Only neutral cues were used. (2) Five different values of C were used, yielding 1st-order peak luminance contrasts ranging from 60 to 80%. (3) Two stimulus conditions were used: HH at 5 deg eccentricity as in Experiment 1, and LL at 10 deg eccentricity as in Experiment 2. Each observer completed a total of 5,760 experimental trials in 16 blocks (in 4 experimental sessions), with 2,880 trials per stimulus condition.
Results and discussion
Varying the contrast of the 1st-order content had no consistent effect on 2nd-order contrast sensitivity (Fig. 8). One-way ANOVAs revealed no significant effect of 1st-order contrast on either d′max [in both cases F(2,12) < 1] or c50 [LL: F(2,12) = 1.5, p > 0.1; HH: F(2,12) < 1]. This was true for both conditions HH at 5 deg eccentricity (Fig. 8A) and LL at 10 deg eccentricity (Fig. 8B). An effect of 1st-order contrast was found for the HH condition on the slope (n) [F(2,12) = 4.59, p < 0.01], but post-hoc comparisons revealed no clear pattern across 1st-order contrast values. Moreover, no change in slope was observed with exogenous attention in the previous experiments.
Figure 8.
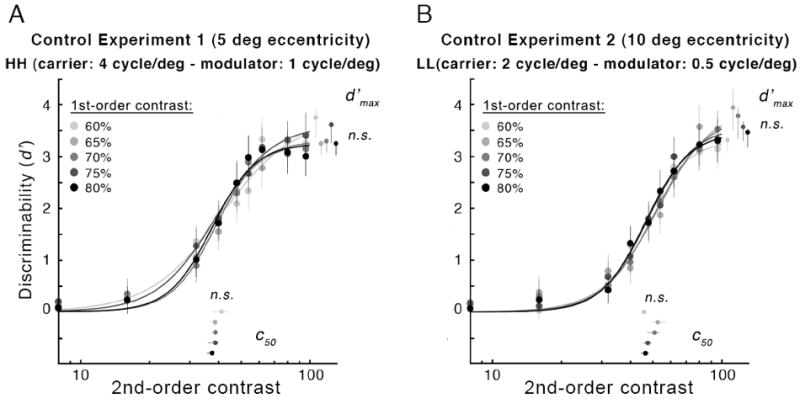
Experiment 3: Effects of 1st-order peak luminance contrast on performance (d′) as a function of 2nd-order modulator contrast (A: HH at 5 deg eccentricity; B: LL at 10 deg eccentricity). Plotting conventions as in Fig. 4, except that the different curves represent different 1st-order peak luminance contrasts in the neutral condition.
It has been shown that 2nd-order contrast thresholds increase with 1st-order contrast (Hess et al., 2008). Our present result does not contradict this previous finding. The changes in 2nd-order sensitivity reported in their study were observed for changes in 1st-order contrast that were larger than the possible consequences of attention. Furthermore, they used contrast-modulated 2nd-order stimuli as opposed to the orientation-modulated 2nd-order stimuli used here.
We conclude that exogenous attention directly affects the sensitivity of 2nd-order channels and that our results from Experiments 1 and 2 are not due to attentional modulation of sensitivity to the 1st-order content of our stimuli.
General discussion
We have shown that exogenous attention increases 2nd-order contrast sensitivity at the attended location and decreases it at unattended locations. The effect of attention was dependent on 2nd-order spatial frequency and eccentricity. Exogenous attention affected contrast sensitivity at parafoveal locations for patterns with high, but not low, 2nd-order spatial frequency (Experiment 1). However, exogenous attention did affect contrast sensitivity for patterns with low 2nd-order spatial frequency when the stimuli were presented at a peripheral location (Experiment 2). We have also shown that these effects on 2nd-order sensitivity were not the indirect consequence of a change in sensitivity at the first stage of visual processing (Experiment 3). The present results provide the first experimental evidence that exogenous attention affects the contrast sensitivity of 2nd-order channels; it increases it at the attended location and decreases it at the unattended location.
Exogenous attention affects 2nd-order contrast sensitivity at both attended and unattended locations
Similar to the effects of attention on 1st-order contrast sensitivity, we showed that exogenous attention increases 2nd-order contrast sensitivity at the attended location and decreases it at unattended locations. The existence of both benefit and cost is consistent with mechanisms of signal enhancement and distracter exclusion that affect contrast sensitivity concurrently (Carrasco, 2006; Carrasco et al., 2000; Dosher & Lu, 2000; Herrmann et al., 2010; Ling & Carrasco, 2006b; Lu et al., 2000; Pestilli & Carrasco, 2005; Pestilli et al., 2009). Whereas the sensory representations of relevant stimuli are boosted at the attended location, consistent with signal enhancement, the strength of stimuli outside the attentional focus is reduced at unattended locations, consistent with distracter exclusion. Exogenous attention affected 2nd-order contrast sensitivity at parafoveal locations via response gain, similar to the effects of exogenous attention on 1st-order contrast sensitivity (Carrasco, 2006; Ling & Carrasco, 2006a; Pestilli et al., 2009; Pestilli et al., 2007). This result may be interpreted as evidence for a common attentional gain-control mechanism at the first and second stages of visual processing. However, in Experiment 2 attention seemed to affect contrast sensitivity at peripheral locations via contrast gain. We do not take this as evidence for different attentional gain-control mechanisms for patterns with high vs. low 2nd-order spatial frequency. Rather, this difference can be explained by the change in eccentricity. First, due to the greater eccentricity used in Experiment 2, contrast sensitivity functions for individual observers did not always reach asymptote, which may have obscured a possible significant change in response gain. Indeed, nested hypothesis tests support the conclusion that in Experiment 1 the effects of exogenous attention on 2nd-order contrast sensitivity are better explained with a response-gain mechanism. However, in Experiment 2 the full model did not fit better than one with either d′max constrained or with c50 constrained, suggesting that the effects of attention are better explained by either a contrast- or a response-gain mechanism or by a combination of both. Second, eccentricity itself may be a factor in how attention affects contrast sensitivity of 2nd-order channels. There has been conflicting evidence regarding the mechanism underlying the effects of attention on contrast sensitivity to 1st-order patterns, with evidence for response gain, contrast gain, or a mixture of both (Cameron et al., 2002; Huang & Dobkins, 2005; Ling & Carrasco, 2006a; Morrone, Denti, & Spinelli, 2004; Pestilli et al., 2009; Pestilli et al., 2007). Recently, the normalization model of attention (Reynolds & Heeger, 2009) has been proposed to reconcile these seemingly conflicting findings. In this model, two critical factors determine how attention affects contrast-response functions: the stimulus size and the attention field size. By changing the relative sizes of these two factors, the model can exhibit response-gain changes, contrast-gain changes, and various combinations of response- and contrast-gain changes. Specifically, when the stimulus is large and the attention field small, attention increases contrast sensitivity via response gain. In contrast, when the stimulus is small and the attention field is large, the model predicts contrast-gain changes. The predictions of this model have been confirmed experimentally by manipulating conjointly the stimulus size and the size of the attention field (Herrmann et al., 2010).
Exogenous attention corresponds to an involuntary capture of attention to a particular location. Three factors in Experiment 2 could have induced a larger attentional window than in Experiment 1: (1) the peripheral precue was at a greater eccentricity; (2) the increased distance between the precue and the stimulus; and (3) although the stimulus size was constant at both eccentricities, they yielded a smaller cortical representation for the peripheral stimuli. According to the normalization model of attention, any or a combination of these three factors could explain a shift from response gain to contrast gain between Experiments 1 and 2.
In sum, our results reveal that exogenous attention affects 2nd-order contrast sensitivity at both attended and unattended locations and that the mechanisms involved at the first and second stages of visual processing may be similar.
Effects of attention on 1st-order contrast sensitivity and early nonlinearities
Could an increase in 1st-order contrast sensitivity with attention (Carrasco, 2006; Pestilli & Carrasco, 2005) affect sensitivity to 2nd-order patterns in Experiments 1 and 2 without a change in the sensitivity of 2nd-order channels? We ruled out this possibility by varying the 1st-order contrast without manipulating attention (Experiment 3). Another issue that must be considered is the possible effects of early nonlinearities. Early luminance nonlinearities, prior to the first stage of linear spatial filtering, can sometimes reveal 2nd-order patterns to the first linear stage. This is particularly true of contrast-modulated stimuli, for which an early nonlinearity can lead to distortion products resulting in Fourier energy (after the nonlinearity) at the frequency of the 2nd-order modulator. However, we used orientation-defined stimuli for which early nonlinearities are ineffective; the distribution of luminance values is identical in the two carrier patterns so that any point-nonlinearity cannot result in differential mean response to the two textures. In addition, if an early nonlinearity were able to demodulate the stimulus, then we would predict an effect of attention in all texture conditions, independent of 2nd-order spatial frequency. Our results were inconsistent with this prediction (Experiment 1). We conclude that exogenous attention directly affects the sensitivity of linear filters at the second stage of visual processing.
Effects of exogenous attention on 2nd-order contrast sensitivity depend on 2nd-order spatial scale
One of our most interesting findings is the dependence of attentional effects on 2nd-order spatial scale. We suggest that the effects of exogenous attention on spatial resolution can explain the absence of attentional modulation for patterns with low 2nd-order spatial frequency at parafoveal locations (Experiment 1).
In a texture segregation task, performance improves as a target texture moves from the fovea into the periphery, with performance peaking at mid-peripheral locations, and then falling at larger eccentricities. The poor performance at central locations is commonly referred to as the central performance drop (CPD). The eccentricity at which performance is best depends on the scale of the texture; enlarging a texture shifts the performance peak to more eccentric locations (Kehrer, 1989). Second-order mechanisms are scale invariant (Kingdom, Keeble, & Moulden, 1995; Landy & Bergen, 1991; Sutter & Graham, 1995), which implies the existence of a link between the scales of the corresponding 1st- and 2nd-order spatial channels (Kingdom & Keeble, 1999), and suggests that the preferred 2nd-order scale depends on eccentricity.
Several studies demonstrate the effects of exogenous attention on acuity and texture segmentation performance. Directing exogenous attention to a target location reduces the differences in performance between foveal and peripheral stimuli in visual search tasks (Carrasco, McLean, Katz, & Frieder, 1998), improves performance in tasks limited by acuity or hyperacuity (Carrasco et al., 2002; Golla, Ignashchenkova, Haarmeier, & Thier, 2004; Montagna et al., 2009; Yeshurun & Carrasco, 1999), and improves or impairs texture segmentation performance depending on both the eccentricity and the scale of the texture target (Carrasco, Loula et al., 2006; Carrasco & Yeshurun, 2009; Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000; Yeshurun et al., 2008). These findings support the hypothesis that exogenous attention enables us to resolve finer details by increasing spatial resolution at the attended location, even when it results in impaired performance.
There are several ways attention could enhance spatial resolution and yet harm central performance (i.e., the CPD). First, attention may narrow the size of 2nd-order receptive fields at the attended area. This hypothesis is compatible with several neurophysiological studies on endogenous attention that indicate receptive fields contract around an attended stimulus (Anton-Erxleben, Stephan, & Treue, 2009; Moran & Desimone, 1985; Reynolds & Desimone, 1999; Womelsdorf, Anton-Erxleben, Pieper, & Treue, 2006).
Alternatively, resolution may be enhanced by increasing the sensitivity of the smallest receptive fields at the attended location (Balz & Hock, 1997). Consequently, overall population sensitivity of 2nd-order spatial filters at the attended area would be shifted toward higher spatial frequencies (Abrams, Barbot, & Carrasco, 2010; Carrasco, Loula et al., 2006). Sensitivity to high spatial frequencies decreases with eccentricity, whereas sensitivity to low spatial frequencies is fairly constant across the visual field (DeValois & DeValois, 1988; Graham, Robson, & Nachmias, 1978; Rovamo, Virsu, & Näsänen, 1978). As a result, the strong responses of filters tuned to high spatial frequencies at central locations may, because of cross-frequency inhibition, weaken responses of lower-spatial-frequency filters that are more useful for the texture segmentation task, leading to the CPD (Carrasco, Loula et al., 2006; Carrasco & Yeshurun, 2009; Morikawa, 2000; Yeshurun & Carrasco, 2000). By increasing the sensitivity of the smaller-scale filters, exogenous attention combined with normalization seems to reduce the sensitivity of larger-scale filters at the attended area, accentuating the CPD (Talgar & Carrasco, 2002; Yeshurun & Carrasco, 1998, 2000). Consistent with this cross-frequency-inhibition hypothesis, removing high spatial frequencies from the texture stimulus by low-pass filtering the display (Morikawa, 2000) or having observers adapt to high spatial frequencies (Carrasco, Loula et al., 2006) eliminates the CPD. Moreover, adapting to high spatial frequencies eliminates the impairment of performance by attention at central locations and diminishes the benefits of attention at peripheral locations (Carrasco, Loula et al., 2006).
Our results strongly support the hypothesis that exogenous attention mediates performance in texture segmentation tasks by increasing the sensitivity of smaller-scale filters that participate in the normalization process. At parafoveal locations (Experiment 1), peripheral cues improved performance for high frequencies, suggesting that attention enhances the sensitivity of small-scale 2nd-order filters. As a result, the sensitivity of 2nd-order filters tuned to patterns with relatively high (1 cycle/deg) 2nd-order spatial frequency was increased, resulting in an enhancement of contrast sensitivity at the attended area. However, the sensitivity of larger-scale filters tuned to patterns with relatively low (0.5 cycle/deg) 2nd-order spatial frequency was not altered, leading to the absence of attentional effects for these stimuli. In contrast, in the periphery, the preferred spatial frequency for filters tuned to the highest spatial frequency at that eccentricity may have been already well matched for the relatively low 2nd-order spatial frequency textures we used. By increasing the sensitivity of the higher-spatial-frequency filters, attention would have therefore increased the contrast sensitivity for the patterns with low 2nd-order spatial frequency. Thus, cross-frequency inhibition can explain the pattern of attentional effects observed in Experiments 1 and 2.
In sum, our results indicate that exogenous attention increases contrast sensitivity of 2nd-order channels whose frequency preference depends on the spatial resolution at the target location. This supports the hypothesis that exogenous attention acts on spatial resolution by increasing the sensitivity of the filters with the highest preferred 2nd-order spatial frequency at the attended area (Carrasco, Loula, et al., 2006; Yeshurun & Carrasco, 2000).
Implications for visual segmentation and object perception
Robust segmentation of the visual scene into distinct perceptually coherent regions is crucial for the reliable detection and identification of objects. By segregating objects from their background contexts, the visual system considerably reduces the complexity of interpreting the retinal input. Object segmentation begins with the detection of discontinuities representing boundaries between adjacent regions, rather than immediate detection of objects per se (Appelbaum, Wade, Pettet, Vildavski, & Norcia, 2008; Li, 2003). Naturally occurring edges are generally defined by spatially coincident changes in both 1st-order luminance and 2nd-order texture information (Johnson & Baker, 2004; Johnson, Kingdom, & Baker, 2005). Access to multiple perceptual cues generally improves performance, and this is the case with 1st- and 2nd-order contours. The presence of both 1st- and 2nd-order cues significantly improves texture segmentation (Smith & Scott-Samuel, 2001), but only when the two cues are correlated in an ecologically valid manner (Johnson, Prins, Kingdom, & Baker, 2007). Perceived edge location is a compromise between the position signaled by texture cues and by other cues such as luminance or motion (Rivest & Cavanagh, 1996). Moreover, when an edge is defined by multiple cues, the cues are combined using a weighted average, with greater weight given to the more reliable cues (Landy & Kojima, 2001).
In natural scenes, 2nd-order texture information may provide a more reliable signal of discontinuities between adjacent surfaces. Indeed, natural scenes contain many luminance edges that do not indicate the presence of an object boundary, generally originating from non-uniform illumination of the surfaces (Kingdom, 2003; Schofield, Hesse, Rock, & Georgeson, 2006; Schofield, Rock, Sun, Jiang, & Georgeson, 2010; Sun & Schofield, 2011). Consequently, by increasing the sensitivity to 2nd-order texture information, exogenous attention can serve to highlight region boundaries in visual scenes, improving object detection and identification.
Conclusion
This study demonstrates that exogenous attention affects contrast sensitivity at the second stage of cortical processing, increasing sensitivity at an attended location and decreasing it at unattended locations. The present findings offer evidence that exogenous attention affects intermediate processes of form vision, thus playing a role in object perception. Together with the effects of covert attention on 1st-order contrast sensitivity (Cameron et al., 2002; Carrasco, 2006; Carrasco et al., 2000; Dosher & Lu, 2000; Herrmann et al., 2010; Ling & Carrasco, 2006b; Lu et al., 2000; Pestilli & Carrasco, 2005; Pestilli et al., 2007), this study suggests that performance tradeoffs between attended and unattended visual attributes may be a general mechanism of selective attention. Our results support the idea that spatial covert attention helps regulate the expenditure of cortical computation at both the first and second stages of visual processing.
Acknowledgments
The research described here was supported by Grants NIH R01-EY016200 to MC and R01-EY16165 to MSL. We would like to thank Katrin Herrmann for analysis code, Romain Barbot for the picture, as well as the members of the Carrasco lab and Toni Saarela for comments on an earlier draft of this paper.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Abrams J, Barbot A, Carrasco M. Voluntary attention increases perceived spatial frequency. Attention, Perception & Psychophysics. 2010;72(6):1510–1521. doi: 10.3758/APP.72.6.1510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Allen HA, Ledgeway T. Attentional modulation of threshold sensitivity to first-order motion and second-order motion patterns. Vision Research. 2003;43(27):2927–2936. doi: 10.1016/j.visres.2003.07.005. [DOI] [PubMed] [Google Scholar]
- Anton-Erxleben K, Abrams J, Carrasco M. Evaluating comparative and equality judgments in contrast perception: attention alters appearance. Journal of Vision. 2010;10(11):6, 1–22. doi: 10.1167/10.11.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anton-Erxleben K, Henrich C, Treue S. Attention changes perceived size of moving visual patterns. Journal of Vision. 2007;7(11):5, 1–9. doi: 10.1167/7.11.5. [DOI] [PubMed] [Google Scholar]
- Anton-Erxleben K, Stephan VM, Treue S. Attention reshapes center-surround receptive field structure in macaque cortical area MT. Cerebral Cortex. 2009;19(10):2466–2478. doi: 10.1093/cercor/bhp002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Appelbaum LG, Norcia AM. Attentive and pre-attentive aspects of figural processing. Journal of Vision. 2009;9(11):18, 1–12. doi: 10.1167/9.11.18. [DOI] [PubMed] [Google Scholar]
- Appelbaum LG, Wade AR, Pettet MW, Vildavski VY, Norcia AM. Figure-ground interaction in the human visual cortex. Journal of Vision. 2008;8(9):8, 1–19. doi: 10.1167/8.9.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arsenault AS, Wilkinson F, Kingdom FA. Modulation frequency and orientation tuning of second-order texture mechanisms. Journal of the Optical Society of America: Optics Image Science and Vision. 1999;16(3):427–435. doi: 10.1364/josaa.16.000427. [DOI] [PubMed] [Google Scholar]
- Azzopardi P, Jones KE, Cowey A. Uneven mapping of magnocellular and parvocellular projections from the lateral geniculate nucleus to the striate cortex in the macaque monkey. Vision Research. 1999;39(13):2179–2189. doi: 10.1016/s0042-6989(98)00319-8. [DOI] [PubMed] [Google Scholar]
- Baker CL., Jr Central neural mechanisms for detecting second-order motion. Current Opinion in Neurobiology. 1999;9(4):461–466. doi: 10.1016/S0959-4388(99)80069-5. [DOI] [PubMed] [Google Scholar]
- Baker CL, Jr, Mareschal I. Processing of second-order stimuli in the visual cortex. Progress in Brain Research. 2001;134:171–191. doi: 10.1016/s0079-6123(01)34013-x. [DOI] [PubMed] [Google Scholar]
- Balz GW, Hock HS. The effect of attentional spread on spatial resolution. Vision Research. 1997;37(11):1499–1510. doi: 10.1016/s0042-6989(96)00296-9. [DOI] [PubMed] [Google Scholar]
- Bergen JR, Adelson EH. Early vision and texture perception. Nature. 1988;333(6171):363–364. doi: 10.1038/333363a0. [DOI] [PubMed] [Google Scholar]
- Braun J, Sagi D. Vision outside the focus of attention. Perception & Psychophysics. 1990;48(1):45–58. doi: 10.3758/bf03205010. [DOI] [PubMed] [Google Scholar]
- Brefczynski JA, DeYoe EA. A physiological correlate of the ‘spotlight’ of visual attention. Nature Neuroscience. 1999;2(4):370–374. doi: 10.1038/7280. [DOI] [PubMed] [Google Scholar]
- Cameron EL, Tai JC, Carrasco M. Covert attention affects the psychometric function of contrast sensitivity. Vision Research. 2002;42(8):949–967. doi: 10.1016/s0042-6989(02)00039-1. [DOI] [PubMed] [Google Scholar]
- Carrasco M. Covert attention increases contrast sensitivity: Psychophysical, neurophysiological and neuroimaging studies. Progress in Brain Research. 2006;154:33–70. doi: 10.1016/S0079-6123(06)54003-8. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Giordano AM, McElree B. Temporal performance fields: visual and attentional factors. Vision Research. 2004;44(12):1351–1365. doi: 10.1016/j.visres.2003.11.026. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Giordano AM, McElree B. Attention speeds processing across eccentricity: feature and conjunction searches. Vision Research. 2006;46(13):2028–2040. doi: 10.1016/j.visres.2005.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Ling S, Read S. Attention alters appearance. Nature Neuroscience. 2004;7(3):308–313. doi: 10.1038/nn1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Loula F, Ho YX. How attention enhances spatial resolution: evidence from selective adaptation to spatial frequency. Perception & Psychophysics. 2006;68(6):1004–1012. doi: 10.3758/bf03193361. [DOI] [PubMed] [Google Scholar]
- Carrasco M, McElree B. Covert attention accelerates the rate of visual information processing. Proceedings of the National Academy of Sciences. 2001;98(9):5363–5367. doi: 10.1073/pnas.081074098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, McLean TL, Katz SM, Frieder KS. Feature asymmetries in visual search: effects of display duration, target eccentricity, orientation and spatial frequency. Vision Research. 1998;38(3):347–374. doi: 10.1016/s0042-6989(97)00152-1. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Penpeci-Talgar C, Eckstein M. Spatial covert attention increases contrast sensitivity across the CSF: support for signal enhancement. Vision Research. 2000;40(10-12):1203–1215. doi: 10.1016/s0042-6989(00)00024-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrasco M, Williams PE, Yeshurun Y. Covert attention increases spatial resolution with or without masks: support for signal enhancement. Journal of Vision. 2002;2(6):4, 467–479. doi: 10.1167/2.6.4. [DOI] [PubMed] [Google Scholar]
- Carrasco M, Yeshurun Y. Covert attention effects on spatial resolution. Progress in Brain Research. 2009;176:65–86. doi: 10.1016/S0079-6123(09)17605-7. [DOI] [PubMed] [Google Scholar]
- Casco C, Grieco A, Campana G, Corvino MP, Caputo G. Attention modulates psychophysical and electrophysiological response to visual texture segmentation in humans. Vision Research. 2005;45(18):2384–2396. doi: 10.1016/j.visres.2005.02.022. [DOI] [PubMed] [Google Scholar]
- Cheal M, Lyon DR. Central and peripheral precuing of forced-choice discrimination. Quarterly Journal of Experimental Psychology. 1991;43(4):859–880. doi: 10.1080/14640749108400960. [DOI] [PubMed] [Google Scholar]
- Dakin SC, Williams CB, Hess RF. The interaction of first- and second-order cues to orientation. Vision Research. 1999;39(17):2867–2884. doi: 10.1016/s0042-6989(98)00307-1. [DOI] [PubMed] [Google Scholar]
- DeValois RL, DeValois KK. Spatial Vision. New York: Oxford University Press; 1988. [Google Scholar]
- Dosher BA, Lu ZL. Mechanisms of perceptual attention in precuing of location. Vision Research. 2000;40(10-12):1269–1292. doi: 10.1016/s0042-6989(00)00019-5. [DOI] [PubMed] [Google Scholar]
- Downing CJ. Expectancy and visual-spatial attention: effects on perceptual quality. Journal of Experimental Psychology: Human Perception and Performance. 1988;14(2):188–202. doi: 10.1037//0096-1523.14.2.188. [DOI] [PubMed] [Google Scholar]
- Ellemberg D, Allen HA, Hess RF. Investigating local network interactions underlying first- and second-order processing. Vision Research. 2004;44(15):1787–1797. doi: 10.1016/j.visres.2004.02.012. [DOI] [PubMed] [Google Scholar]
- Ellemberg D, Allen HA, Hess RF. Second-order spatial frequency and orientation channels in human vision. Vision Research. 2006;46(17):2798–2803. doi: 10.1016/j.visres.2006.01.028. [DOI] [PubMed] [Google Scholar]
- Fuller S, Carrasco M. Exogenous attention and color perception: performance and appearance of saturation and hue. Vision Research. 2006;46(23):4032–4047. doi: 10.1016/j.visres.2006.07.014. [DOI] [PubMed] [Google Scholar]
- Gandhi SP, Heeger DJ, Boynton GM. Spatial attention affects brain activity in human primary visual cortex. Proceedings of the National Academy of Sciences. 1999;96(6):3314–3319. doi: 10.1073/pnas.96.6.3314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giordano AM, McElree B, Carrasco M. On the automaticity and flexibility of covert attention: a speed-accuracy trade-off analysis. Journal of Vision. 2009;9(3):30, 1–10. doi: 10.1167/9.3.30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gobell J, Carrasco M. Attention alters the appearance of spatial frequency and gap size. Psychological Science. 2005;16(8):644–651. doi: 10.1111/j.1467-9280.2005.01588.x. [DOI] [PubMed] [Google Scholar]
- Golla H, Ignashchenkova A, Haarmeier T, Thier P. Improvement of visual acuity by spatial cueing: a comparative study in human and non-human primates. Vision Research. 2004;44(13):1589–1600. doi: 10.1016/j.visres.2004.01.009. [DOI] [PubMed] [Google Scholar]
- Graham N. Visual pattern analyzers. New York: Oxford University Press; 1989. [Google Scholar]
- Graham N. Complex channels, early local nonlinearities, and normalization in texture segregation. In: Landy ML, Movshon JA, editors. Computational models of visual processing. Cambridge, MA: MIT Press; 1991. [Google Scholar]
- Graham N, Beck J, Sutter A. Nonlinear processes in spatial-frequency channel models of perceived texture segregation: effects of sign and amount of contrast. Vision Research. 1992;32(4):719–743. doi: 10.1016/0042-6989(92)90188-o. [DOI] [PubMed] [Google Scholar]
- Graham N, Robson JG, Nachmias J. Grating summation in fovea and periphery. Vision Research. 1978;18(7):815–825. doi: 10.1016/0042-6989(78)90122-0. [DOI] [PubMed] [Google Scholar]
- Graham N, Wolfson SS. A note about preferred orientations at the first and second stages of complex (second-order) texture channels. Journal of the Optical Society of America: Optics Image Science and Vision. 2001;18(9):2273–2281. doi: 10.1364/josaa.18.002273. [DOI] [PubMed] [Google Scholar]
- Herrmann K, Montaser-Kouhsari L, Carrasco M, Heeger DJ. When size matters: attention affects performance by contrast or response gain. Nature Neuroscience. 2010;13(12):1554–1559. doi: 10.1038/nn.2669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hess RF, Baker DH, May KA, Wang J. On the decline of 1st and 2nd order sensitivity with eccentricity. Journal of Vision. 2008;8(1):19, 1–12. doi: 10.1167/8.1.19. [DOI] [PubMed] [Google Scholar]
- Huang L, Dobkins KR. Attentional effects on contrast discrimination in humans: evidence for both contrast gain and response gain. Vision Research. 2005;45(9):1201–1212. doi: 10.1016/j.visres.2004.10.024. [DOI] [PubMed] [Google Scholar]
- Huk AC, Heeger DJ. Task-related modulation of visual cortex. Journal of Neurophysiology. 2000;83(6):3525–3536. doi: 10.1152/jn.2000.83.6.3525. [DOI] [PubMed] [Google Scholar]
- Johnson AP, Baker CL., Jr First- and second-order information in natural images: a filter-based approach to image statistics. Journal of the Optical Society of America: Optics Image Science and Vision. 2004;21(6):913–925. doi: 10.1364/josaa.21.000913. [DOI] [PubMed] [Google Scholar]
- Johnson AP, Kingdom FA, Baker CL., Jr Spatiochromatic statistics of natural scenes: first- and second-order information and their correlational structure. Journal of the Optical Society of America: Optics Image Science and Vision. 2005;22(10):2050–2059. doi: 10.1364/josaa.22.002050. [DOI] [PubMed] [Google Scholar]
- Johnson AP, Prins N, Kingdom FA, Baker CL., Jr Ecologically valid combinations of first- and second-order surface markings facilitate texture discrimination. Vision Research. 2007;47(17):2281–2290. doi: 10.1016/j.visres.2007.05.003. [DOI] [PubMed] [Google Scholar]
- Jonides J, Irwin DE. Capturing attention. Cognition. 1981;10(1-3):145–150. doi: 10.1016/0010-0277(81)90038-x. [DOI] [PubMed] [Google Scholar]
- Jonides J, Yantis S. Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics. 1988;43(4):346–354. doi: 10.3758/bf03208805. [DOI] [PubMed] [Google Scholar]
- Julesz B. Textons, the elements of texture perception, and their interactions. Nature. 1981;290(5802):91–97. doi: 10.1038/290091a0. [DOI] [PubMed] [Google Scholar]
- Kastner S, De Weerd P, Ungerleider LG. Texture segregation in the human visual cortex: A functional MRI study. Journal of Neurophysiology. 2000;83(4):2453–2457. doi: 10.1152/jn.2000.83.4.2453. [DOI] [PubMed] [Google Scholar]
- Kehrer L. Central performance drop on perceptual segregation tasks. Spatial Vision. 1989;4(1):45–62. doi: 10.1163/156856889x00040. [DOI] [PubMed] [Google Scholar]
- Kinchla RA, Chen Z, Evert D. Precue effects in visual search: data or resource limited? Perception & Psychophysics. 1995;57(4):441–450. doi: 10.3758/bf03213070. [DOI] [PubMed] [Google Scholar]
- Kingdom FA. Color brings relief to human vision. Nature Neuroscience. 2003;6(6):641–644. doi: 10.1038/nn1060. [DOI] [PubMed] [Google Scholar]
- Kingdom FA, Keeble D, Moulden B. Sensitivity to orientation modulation in micropattern-based textures. Vision Research. 1995;35(1):79–91. doi: 10.1016/0042-6989(94)e0079-z. [DOI] [PubMed] [Google Scholar]
- Kingdom FA, Keeble DR. On the mechanism for scale invariance in orientation-defined textures. Vision Research. 1999;39(8):1477–1489. doi: 10.1016/s0042-6989(98)00217-x. [DOI] [PubMed] [Google Scholar]
- Landy MS, Bergen JR. Texture segregation and orientation gradient. Vision Research. 1991;31(4):679–691. doi: 10.1016/0042-6989(91)90009-t. [DOI] [PubMed] [Google Scholar]
- Landy MS, Graham N. Visual perception of texture. In: Chalupa LM, Werner JS, editors. The Visual Neurosciences. Cambridge, MA: MIT Press; 2004. pp. 1106–1118. [Google Scholar]
- Landy MS, Kojima H. Ideal cue combination for localizing texture-defined edges. Journal of the Optical Society of America: Optics, Image Science, and Vision. 2001;18(9):2307–2320. doi: 10.1364/josaa.18.002307. [DOI] [PubMed] [Google Scholar]
- Landy MS, Oruç I. Properties of second-order spatial frequency channels. Vision Research. 2002;42(19):2311–2329. doi: 10.1016/s0042-6989(02)00193-1. [DOI] [PubMed] [Google Scholar]
- Larsson J, Landy MS, Heeger DJ. Orientation-selective adaptation to first- and second-order patterns in human visual cortex. Journal of Neurophysiology. 2006;95(2):862–881. doi: 10.1152/jn.00668.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leigh RJ, Zee DS. The neurology of eye movements. Philadelphia, PA: 1991. [Google Scholar]
- Lennie P. The cost of cortical computation. Current Biology. 2003;13(6):493–497. doi: 10.1016/s0960-9822(03)00135-0. [DOI] [PubMed] [Google Scholar]
- Li Z. V1 mechanisms and some figure-ground and border effects. Journal of Physiology - Paris. 2003;97(4-6):503–515. doi: 10.1016/j.jphysparis.2004.01.008. [DOI] [PubMed] [Google Scholar]
- Ling S, Carrasco M. Sustained and transient covert attention enhance the signal via different contrast response functions. Vision Research. 2006a;46(8-9):1210–1220. doi: 10.1016/j.visres.2005.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ling S, Carrasco M. When sustained attention impairs perception. Nature Neuroscience. 2006b;9(10):1243–1245. doi: 10.1038/nn1761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T, Pestilli F, Carrasco M. Transient attention enhances perceptual performance and FMRI response in human visual cortex. Neuron. 2005;45(3):469–477. doi: 10.1016/j.neuron.2004.12.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu T, Stevens ST, Carrasco M. Comparing the time course and efficacy of spatial and feature-based attention. Vision Research. 2007;47(1):108–113. doi: 10.1016/j.visres.2006.09.017. [DOI] [PubMed] [Google Scholar]
- Lu ZL, Dosher BA. External noise distinguishes attention mechanisms. Vision Research. 1998;38(9):1183–1198. doi: 10.1016/s0042-6989(97)00273-3. [DOI] [PubMed] [Google Scholar]
- Lu ZL, Dosher BA. Spatial attention excludes external noise without changing the spatial frequency tuning of the perceptual template. Journal of Vision. 2004;4(10):10, 955–966. doi: 10.1167/4.10.10. [DOI] [PubMed] [Google Scholar]
- Lu ZL, Liu CQ, Dosher BA. Attention mechanisms for multi-location first- and second-order motion perception. Vision Research. 2000;40(2):173–186. doi: 10.1016/s0042-6989(99)00172-8. [DOI] [PubMed] [Google Scholar]
- Luck SJ, Hillyard SA, Mouloua M, Woldorff MG, Clark VP, Hawkins HL. Effects of spatial cuing on luminance detectability: psychophysical and electrophysiological evidence for early selection. Journal of Experimental Psychology: Human Perception and Performance. 1994;20(4):887–904. doi: 10.1037//0096-1523.20.4.887. [DOI] [PubMed] [Google Scholar]
- Malik J, Perona P. Preattentive texture discrimination with early vision mechanisms. Journal of the Optical Society of America. 1990;7(5):923–932. doi: 10.1364/josaa.7.000923. [DOI] [PubMed] [Google Scholar]
- Mayfrank L, Kimmig H, Fischer B. The role of attention in the preparation of visually guided saccadic eye movements in man. In: O’Regan JK, Levy-Schoen A, editors. Eye movements: from physiology to cognition. New York: Elsevier-Science Publishers, North-Holland; 1987. pp. 37–45. [Google Scholar]
- McAdams CJ, Reid RC. Attention modulates the responses of simple cells in monkey primary visual cortex. Journal of Neuroscience. 2005;25(47):11023–11033. doi: 10.1523/JNEUROSCI.2904-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montagna B, Pestilli F, Carrasco M. Attention trades off spatial acuity. Vision Research. 2009;49(7):735–745. doi: 10.1016/j.visres.2009.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moran J, Desimone R. Selective attention gates visual processing in the extrastriate cortex. Science. 1985;229(4715):782–784. doi: 10.1126/science.4023713. [DOI] [PubMed] [Google Scholar]
- Morgan MJ, Mason AJ, Baldassi S. Are there separate first-order and secondorder mechanisms for orientation discrimination? Vision Research. 2000;40(13):1751–1763. doi: 10.1016/s0042-6989(00)00015-8. [DOI] [PubMed] [Google Scholar]
- Morikawa K. Central performance drop in texture segmentation: the role of spatial and temporal factors. Vision Research. 2000;40(25):3517–3526. doi: 10.1016/s0042-6989(00)00170-x. [DOI] [PubMed] [Google Scholar]
- Morrone MC, Denti V, Spinelli D. Different attentional resources modulate the gain mechanisms for color and luminance contrast. Vision Research. 2004;44(12):1389–1401. doi: 10.1016/j.visres.2003.10.014. [DOI] [PubMed] [Google Scholar]
- Muller HJ, Findlay JM. The effect of visual attention on peripheral discrimination thresholds in single and multiple element displays. Acta Psychologica (Amst) 1988;69(2):129–155. doi: 10.1016/0001-6918(88)90003-0. [DOI] [PubMed] [Google Scholar]
- Muller HJ, Rabbitt PM. Reflexive and voluntary orienting of visual attention: time course of activation and resistance to interruption. Journal of Experimental Psychology: Human Perception and Performance. 1989;15(2):315–330. doi: 10.1037//0096-1523.15.2.315. [DOI] [PubMed] [Google Scholar]
- Nakayama K, Mackeben M. Sustained and transient components of focal visual attention. Vision Research. 1989;29(11):1631–1647. doi: 10.1016/0042-6989(89)90144-2. [DOI] [PubMed] [Google Scholar]
- Nothdurft HC. Orientation sensitivity and texture segmentation in patterns with different line orientation. Vision Research. 1985;25(4):551–560. doi: 10.1016/0042-6989(85)90159-2. [DOI] [PubMed] [Google Scholar]
- Pestilli F, Carrasco M. Attention enhances contrast sensitivity at cued and impairs it at uncued locations. Vision Research. 2005;45(14):1867–1875. doi: 10.1016/j.visres.2005.01.019. [DOI] [PubMed] [Google Scholar]
- Pestilli F, Ling S, Carrasco M. A population-coding model of attention’s influence on contrast response: Estimating neural effects from psychophysical data. Vision Research. 2009;49(10):1144–1153. doi: 10.1016/j.visres.2008.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pestilli F, Viera G, Carrasco M. How do attention and adaptation affect contrast sensitivity? Journal of Vision. 2007;7(7):9, 1–12. doi: 10.1167/7.7.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posner MI. Orienting of attention. Quaterly Journal of Experimental Psychology. 1980;32(1):3–25. doi: 10.1080/00335558008248231. [DOI] [PubMed] [Google Scholar]
- Remington RW, Johnston JC, Yantis S. Involuntary attentional capture by abrupt onsets. Perception & Psychophysics. 1992;51(3):279–290. doi: 10.3758/bf03212254. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Chelazzi L. Attentional modulation of visual processing. Annual Review of Neuroscience. 2004;27:611–647. doi: 10.1146/annurev.neuro.26.041002.131039. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Desimone R. The role of neural mechanisms of attention in solving the binding problem. Neuron. 1999;24(1):19–29. 111–125. doi: 10.1016/s0896-6273(00)80819-3. [DOI] [PubMed] [Google Scholar]
- Reynolds JH, Heeger DJ. The normalization model of attention. Neuron. 2009;61(2):168–185. doi: 10.1016/j.neuron.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivest J, Cavanagh P. Localizing contours defined by more than one attribute. Vision Research. 1996;36(1):53–66. doi: 10.1016/0042-6989(95)00056-6. [DOI] [PubMed] [Google Scholar]
- Robson JG, Graham N. Probability summation and regional variation in contrast sensitivity across the visual field. Vision Research. 1981;21(3):409–418. doi: 10.1016/0042-6989(81)90169-3. [DOI] [PubMed] [Google Scholar]
- Rovamo J, Virsu V, Näfsänen R. Cortical magnification factor predicts the photopic contrast sensitivity of peripheral vision. Nature. 1978;271(5640):54–56. doi: 10.1038/271054a0. [DOI] [PubMed] [Google Scholar]
- Schira MM, Fahle M, Donner TH, Kraft A, Brandt SA. Differential contribution of early visual areas to the perceptual process of contour processing. Journal of Neurophysiology. 2004;91(4):1716–1721. doi: 10.1152/jn.00380.2003. [DOI] [PubMed] [Google Scholar]
- Schofield AJ. What does second-order vision see in an image? Perception. 2000;29(9):1071–1086. doi: 10.1068/p2913. [DOI] [PubMed] [Google Scholar]
- Schofield AJ, Georgeson MA. Sensitivity to modulations of luminance and contrast in visual white noise: separate mechanisms with similar behaviour. Vision Research. 1999;39(16):2697–2716. doi: 10.1016/s0042-6989(98)00284-3. [DOI] [PubMed] [Google Scholar]
- Schofield AJ, Hesse G, Rock PB, Georgeson MA. Local luminance amplitude modulates the interpretation of shape-from-shading in textured surfaces. Vision Research. 2006;46(20):3462–3482. doi: 10.1016/j.visres.2006.03.014. [DOI] [PubMed] [Google Scholar]
- Schofield AJ, Rock PB, Sun P, Jiang X, Georgeson MA. What is second-order vision for? Discriminating illumination versus material changes. Journal of Vision. 2010;10(9):2, 2. doi: 10.1167/10.9.2. [DOI] [PubMed] [Google Scholar]
- Schubo A, Meinecke C. Automatic texture segmentation in early vision: evidence from priming experiments. Vision Research. 2007;47(18):2378–2389. doi: 10.1016/j.visres.2007.06.002. [DOI] [PubMed] [Google Scholar]
- Schubo A, Meinecke C, Schroger E. Automaticity and attention: investigating automatic processing in texture segmentation with event-related brain potentials. Brain Research Cognitive Brain Research. 2001;11(3):341–361. doi: 10.1016/s0926-6410(01)00008-8. [DOI] [PubMed] [Google Scholar]
- Scott-Samuel NE, Georgeson MA. Does early non-linearity account for secondorder motion? Vision Research. 1999;39(17):2853–2865. doi: 10.1016/s0042-6989(98)00316-2. [DOI] [PubMed] [Google Scholar]
- Smith AT, Scott-Samuel NE. First-order and second-order signals combine to improve perceptual accuracy. Journal of the Optical Society of America: Optics Image Science and Vision. 2001;18(9):2267–2272. doi: 10.1364/josaa.18.002267. [DOI] [PubMed] [Google Scholar]
- Störmer VS, McDonald JJ, Hillyard SA. Cross-modal cueing of attention alters appearance and early cortical processing of visual stimuli. Proceedings of the National Academy of Sciences. 2009;106(52):22456–22461. doi: 10.1073/pnas.0907573106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun P, Schofield AJ. The efficacy of local luminance amplitude in disambiguating the origin of luminance signals depends on carrier frequency: Further evidence for the active role of second-order vision in layer decomposition. Vision Research. 2011 doi: 10.1016/j.visres.2011.01.008. [DOI] [PubMed] [Google Scholar]
- Sutter A, Beck J, Graham N. Contrast and spatial variables in texture segregation: testing a simple spatial-frequency channels model. Perception & Psychophysics. 1989;46(4):312–332. doi: 10.3758/bf03204985. [DOI] [PubMed] [Google Scholar]
- Sutter A, Graham N. Investigating simple and complex mechanisms in texture segregation using the speed-accuracy tradeoff method. Vision Research. 1995;35(20):2825–2843. doi: 10.1016/0042-6989(95)00045-2. [DOI] [PubMed] [Google Scholar]
- Talgar CP, Carrasco M. Vertical meridian asymmetry in spatial resolution: visual and attentional factors. Psychonomic Bulletin & Review. 2002;9(4):714–722. doi: 10.3758/bf03196326. [DOI] [PubMed] [Google Scholar]
- Womelsdorf T, Anton-Erxleben K, Pieper F, Treue S. Dynamic shifts of visual receptive fields in cortical area MT by spatial attention. Nature Neuroscience. 2006;9(9):1156–1160. doi: 10.1038/nn1748. [DOI] [PubMed] [Google Scholar]
- Yantis S, Jonides J. Abrupt visual onsets and selective attention: evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance. 1984;10(5):601–621. doi: 10.1037//0096-1523.10.5.601. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y. Isoluminant stimuli and red background attenuate the effects of transient spatial attention on temporal resolution. Vision Research. 2004;44(12):1375–1387. doi: 10.1016/j.visres.2003.12.016. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. Attention improves or impairs visual performance by enhancing spatial resolution. Nature. 1998;396(6706):72–75. doi: 10.1038/23936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. Spatial attention improves performance in spatial resolution tasks. Vision Research. 1999;39(2):293–306. doi: 10.1016/s0042-6989(98)00114-x. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. The locus of attentional effects in texture segmentation. Nature Neuroscience. 2000;3(6):622–627. doi: 10.1038/75804. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y, Carrasco M. The effects of transient attention on spatial resolution and the size of the attentional cue. Perception & Psychophysics. 2008;70(1):104–113. doi: 10.3758/pp.70.1.104. [DOI] [PubMed] [Google Scholar]
- Yeshurun Y, Montagna B, Carrasco M. On the flexibility of sustained attention and its effects on a texture segmentation task. Vision Research. 2008;48(1):80–95. doi: 10.1016/j.visres.2007.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]