

Abstract
Ehrlichiae are responsible for important tick-transmitted diseases, including anaplasmosis, the most prevalent tick-borne infection of livestock worldwide, and the emerging human diseases monocytic and granulocytic ehrlichiosis. Antigenic variation of major surface proteins is a key feature of these pathogens that allows persistence in the mammalian host, a requisite for subsequent tick transmission. In Anaplasma marginale pseudogenes for two antigenically variable gene families, msp2 and msp3, appear in concert. These pseudogenes can be recombined into the functional expression site to generate new antigenic variants. Coordinated control of the recombination of these genes would allow these two gene families to act synergistically to evade the host immune response.
Antigenic variation in major surface proteins of tick-borne bacterial pathogens is a primary mechanism for evasion of the host immune response and results in persistent infection. A number of different mechanisms have been reported. For instance, bacteria of the genus Borrelia generate antigenic diversity of the vmp/vls coat proteins through recombination from a tandem array of silent partial pseudogene cassettes into a telomeric expression site on a linear plasmid (1, 2). Ehrlichial genogroup I pathogens, Ehrlichia chaffeensis, E. canis, and Cowdria ruminantium, have recently been shown to contain tandemly repeated copies of the omp1/map1 gene family (3–6). These omp1/map1 tandem repeats are complete gene copies, and multiple copies may be transcriptionally active at a given time, resulting in polymorphic protein expression (3).
In contrast to a tandem array of genes or pseudogenes for antigenically variable proteins, Anaplasma marginale, a member of ehrlichial genogroup II, contains immunodominant major surface protein 2 (msp2) and msp3 families that include 10 or more variable genes widely dispersed throughout the 1.2-Mb genome (7–9). Recently we reported the identification of an operon of four ORFs, containing the msp2 gene at the 3′ terminus (10). The other ORFs of the operon occur as a single copy in the A. marginale genome, and the operon has been demonstrated to be a functional expression site for full-length msp2 transcripts. These msp2 transcripts have highly conserved 5′ and 3′ ends, interrupted by a central hypervariable region characterized by substitutions, insertions, and deletions. The hypervariable region encodes a diverse array of B cell epitopes that result in evasion of the host immune system. Similarly, MSP3 is also structurally and antigenically variable (11). Our data show that aside from the expression site in the operon, msp2 does not occur as a full-length gene, but rather as partial pseudogene cassettes, each containing a different hypervariable region and a portion of the 5′ and 3′ conserved regions. We have identified nine pseudogenes for msp2 and shown that these pseudogenes recombine into the operon expression site to generate a new hypervariable sequence. Partial pseudogene cassettes appear for the msp3 gene family as well, and the pseudogenes for the two gene families often appear close together. The two pseudogene families have the same 5′ sequence, indicating that they use the same mechanism to regulate recombination into the expression site.
Materials and Methods
Southern Analysis.
Genomic DNA of the A. marginale South Idaho strain was digested with the use of KpnI and separated on a 0.7% agarose gel. The blot was initially hybridized with a msp2 5′ end-specific probe corresponding to bp 2–335 and then stripped and rehybridized with an orf2-specific probe corresponding to bp 6–359. Generation and digoxigenin labeling of the probes, hybridization, and detection were as recommended by the manufacturer of the PCR labeling kit (Roche Molecular Biochemicals). High stringency wash conditions were as follows: two washes in 2× SSC, 0.1% SDS (wt/vol) at room temperature, one wash in the same buffer at 65°C, and a final wash in 0.2× SSC, 0.1% SDS (wt/vol) at 65°C (1× SSC = 0.15 M sodium chloride/0.015 M sodium citrate, pH 7). All washes lasted 15 min.
Bacterial Artificial Chromosome (BAC) Library Construction and Manipulation.
Blood was collected from calf 836 during acute A. marginale (St. Maries strain) rickettsemia. Erythrocytes were isolated with the use of Histopaque (Sigma) and embedded in agarose blocks, and cells were lysed within the agarose blocks with the use of proteinase K and SDS (12). A. marginale genomic DNA was partially digested with HindIII, size selected on pulse field gels, ligated into the vector pBELOBAC11, and electroporated into Escherichia coli strain DH10B. A total of 1,536 BAC clones were arrayed into 384 well plates with an average insert size of 110 Kb. BAC 2439 was selected for sequence analysis after probing with a digoxigenin-labeled (Roche Molecular Biochemicals) msp2 probe (bp 375–965). Random shotgun libraries were constructed from partially digested BAC 2439 DNA. The randomly generated fragments were size selected, cloned into pCRScript (Stratagene), and electroporated into E. coli strain XL-1Blue. Insert DNA was sequenced with the use of BigDye terminator chemistry on an ABI 377XL-96 instrument (PE-Applied Biosystems). Data were assembled and analyzed by using sequencher (Gene Codes, Ann Arbor, MI) and PHRED & PHRAP software (Sanger Centre, Cambridge, UK). When required, gene walking or direct BAC sequencing was performed to ensure a minimum of 2× coverage, with an overall average of 3× coverage. The finished sequence contains 44,557 bp (accession number AF305077).
PCR Cloning of Pseudogenes and Operon-Linked msp2 Fragments.
Ticks fed on A. marginale (South Idaho strain)-infected calf 824 subsequently transmitted the infection to calf 828. Blood was taken during acute rickettsemia for both animals, and genomic DNA was isolated with a Puregene DNA extraction kit (Gentra Systems). Total RNA was isolated with the use of TRIzol (GIBCO/BRL). RNA was treated with DNase I, followed by cDNA synthesis with random hexamers, with the use of the Thermoscript reverse transcription–PCR kit from GIBCO/BRL. The primers used to amplify the operon-linked hypervariable regions were orf 2 forward primer TCCTACCAAGCGTCTTTTCCCC and msp2 reverse primer: TTACCACCGATACCAGCACAA. PCR was performed with DNA, RNA (as negative control), or cDNA and fragments cloned with the pMOSBlue cloning system (Amersham Pharmacia). Inserts were sequenced in both directions with the Big Dye kit and an ABI PRISM automated sequencer (PE-Applied Biosystems). Sequences were compiled and analyzed with the vector nti (InforMax, North Bethesda, MD) and gcg (University of Wisconsin) software packages.
Results and Discussion
The msp2 gene is estimated to have 10 or more copies (7), whereas the operon containing the expressed copy of msp2 is reported to be a single copy (10). Importantly, as shown in Fig. 1A, the 5′ end of msp2 also occurs only as a single copy. Southern analysis of KpnI-digested A. marginale DNA with a msp2 5′ end probe (bp 2–335) detects two fragments of 1.2 and 4 kb. These same two bands are detected when the blot is rehybridized with an operon-specific probe (ORF2, bp 6–359) and result from a KpnI polymorphism in the hypervariable region of msp2 (Fig. 1B). Detection of both the 1.2- and 4-kb fragments reflects the oligoclonal nature of the infection in vivo. These data demonstrate that the operon is the only expression site for full-length msp2 transcripts. Therefore the remaining copies of msp2 throughout the genome were hypothesized to be truncated pseudogenes.
Figure 1.

(A) Presence of a single genomic expression site for msp2. (Left) A blot hybridized with a msp2 5′ end-specific probe corresponding to bp 2-335. (Right) The same blot hybridized with an orf2-specific probe corresponding to bp 6-359. Lambda HindIII markers are shown. (B) Schematic representation of the msp2 operon. The hypervariable region of msp2 is stippled. Positions of KpnI sites are shown (K). The KpnI site in the hypervariable region is polymorphic. Arrows indicate the positions of primers used to amplify DNA and cDNA clones.
To identify potential pseudogenes, a BAC library from the St. Maries strain of A. marginale was screened for the presence of msp2. A 45-kb positive clone (BAC 2439) was sequenced, and two pseudogenes for msp2 and two pseudogenes for msp3 (Fig. 2) were found. A third msp2 pseudogene was previously cloned from the Florida strain in conjunction with msp3 (clone 3-11; ref. 8) but remained unrecognized as such until comparison with the sequences in BAC 2439. This third pseudogene is on the opposite strand of DNA, 321 bp downstream from the 3′ end of the msp3-11 coding sequence (Fig. 2). Although these three msp2 pseudogenes do not have uniform start and stop positions, their general structures are similar: they contain a portion of the 5′ conserved region, continue through the hypervariable region, and contain a portion of the 3′ conserved region. Thus, these pseudogenes have a structure capable of recombination into the operon expression site to generate new hypervariable regions of msp2.
Figure 2.

Schematic representation of genomic pseudogenes. BAC 2439 is 45 kb in length. Msp3-11 is 3263 bp in length. It is shown to the same scale as BAC 2439 for comparison and is enlarged to show the positions of primers F1, F2, R1, and R2 used for amplification and cloning of pseudogenes. The sequence of the primers is as follows: F1: GCACCAAAGAATATAGCTGTAAATAC; F2: CCCAGCTTCTGCACAAAC; R1: TGTTGCCCGCCATCC; R2: CTCTAGCACCTTCAGCATC. The complete pseudogene complex is illustrated. Msp2 pseudogenes are striped and msp3 pseudogenes are dark gray. The orientation of pseudogenes is indicated by the direction of the arrow. Ovals indicate the presence of the conserved 600-bp sequence.
The motif of a msp3 pseudogene on one strand and a msp2 pseudogene on the opposite strand was observed both in BAC 2439 and in the msp3-11 clone. The 321 bp separating msp2 from msp3 in each of these clones was identical, even though the respective msp2 and msp3 coding regions themselves were not identical between the two clones. Furthermore, ≈600 bp 5′ to msp2 pseudogenes 1 and 3 and msp3 pseudogenes 1 and 2 was highly conserved (93.8–99.7% identity). This 600-bp 5′ flanking region is indicated in Fig. 2 by ovals. Msp2 pseudogene 2 in BAC 2439 had a 5′ extension that was not seen in any of the other pseudogenes, such that it started at bp 236 relative to pCKR11.2. Southern analysis indicated that this 5′ extension occurred only once in the genome and was an aberration that was specific to the St. Maries strain of A. marginale (data not shown). This pseudogene is incapable of encoding a full-length msp2 gene, and it differs from the other msp2 pseudogenes in that it does not have the conserved 600-bp 5′ flanking region.
The two regions of sequence conservation, the 600-bp 5′ region and the 321-bp 3′ region, flanking the msp2 pseudogenes allowed sets of primers to be designed that could specifically amplify msp2 pseudogenes. Primer positions are shown in Fig. 2 relative to msp2 pseudogene 3. Five distinct DNA fragments were obtained by PCR from A. marginale (South Idaho, calf 824) genomic DNA, with four primer combinations (Fig. 3). These fragments were cloned, and 10 clones from each PCR reaction were sequenced. With this strategy, six additional pseudogenes were identified (Fig. 4). The ORFs for these pseudogenes start uniformly with the sequence PYQGYHSMLTALE, with identity to full-length MSP2 starting with the serine corresponding to amino acid 112 [numbering based on pCKR11.2, a full-length msp2 gene (7)]. The pseudogenes are 186–194 aa in length, with the variation in length due to differences in the hypervariable region. The four pseudogenes generated with primer R1 (Fig. 2) in the 321-bp intergenic region uniformly ended at amino acid 284 (Fig. 4). The remaining two pseudogenes were generated with primer R2 in the 3′ conserved region of msp2 (Figs. 2 and 4).
Figure 3.
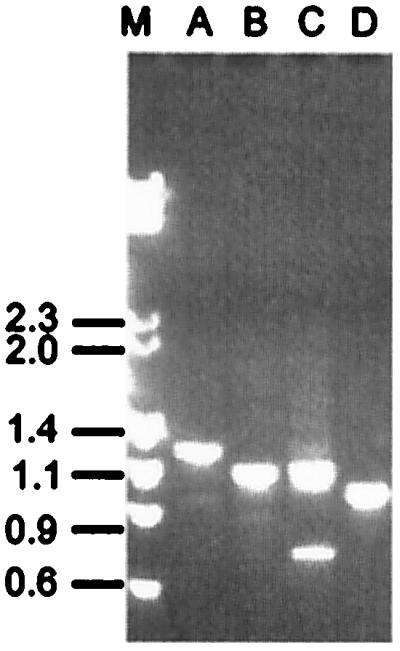
Pseudogene amplicons generated with primers F1, F2, R1, and R2. Amplicons were generated with the use of A. marginale South Idaho DNA from calf 824. The amplicon in Lane A was generated with primers F1 and R1, Lane B used primers F2 and R1, Lane C used primers F1 and R2, and Lane D used primers F2 and R2. Lane M contains the molecular weight standards λHindIII and φχHaeIII.
Figure 4.

(A) Schematic representation of full-length msp2 compared with the msp2 pseudogenes. Numbering of full-length msp2 is indicated in base pairs and amino acids. The hypervariable region is indicated by stippling. (B) The deduced amino acid sequences of the msp2 pseudogenes obtained in this study are shown in comparison to the full-length msp2 sequence. The 5′ and 3′ conserved regions are on a gray background and flank the hypervariable region. The full-length pCKR11.2 continues for 134 aa (not shown). Pseudogenes 1 and 2 are from an A. marginale St. Maries strain, pseudogene 3 is from an A. marginale Florida strain (8), and the remaining pseudogenes are from an A. marginale South Idaho strain and are lettered A-D as denoted in Fig. 3.
The close positioning of msp2 and msp3 pseudogenes in a tail-to-tail arrangement was a recurring motif in the genome. In addition to msp2 pseudogenes 2 and 3 found in this arrangement, four of the six msp2 pseudogenes cloned in this study were amplified with the use of a primer specific for the intergenic region between these two genes, indicating that they, too, were arranged in the same manner. Furthermore, amplification of A. marginale DNA with the use of a single primer, F1, specific for the 5′ flanking region generates a product of ≈4.3 kb, the expected size of the pseudogene complex, corresponding to a 5′ flanking region, a msp3 pseudogene, the 321-bp intergenic region, with a msp2 pseudogene, and a second 5′ flanking region on the opposite strand (Fig. 2).
The pseudogenes are not functional transcription units, as there are no promoter consensus sequences within 350 bp of the first methionine of each pseudogene. Furthermore, polyclonal antibodies that detect a full-length MSP2 (42 kDa) in Western blots do not detect any proteins in the region of 18 kDa, the expected size of the product if the pseudogenes were expressed, indicating that the pseudogenes are not functional coding regions (data not shown). Consequently pseudogene expression would require recombination into the expression site.
To address the question of whether these pseudogenes recombine into the operon expression site of msp2, we examined the expression site at various time points during infection. A. marginale DNA was isolated from two calves infected with the South Idaho strain and amplified with the use of primers spanning the hypervariable region of msp2 and linking these products to the operon, thus establishing that they were derived from the operon expression site (primer positions indicated in Fig. 1). PCR-derived amplicons were cloned, and genomic clones were sequenced. Pseudogene A-3 (AF305503) had a corresponding operon-linked genomic clone (828-45 g), demonstrating that pseudogenes, which reside elsewhere in the South Idaho strain genome, are recombined into the expression site during infection with this strain. To verify that the recombined pseudogene A-3 gave rise to functional transcripts, the experiment was repeated with RNA as the template, and the fragments from the resulting reverse transcription–PCR were cloned. Sequence analysis identified two clones (828-1c and 824-52c) that corresponded to A-3 and 828-45 g. These results demonstrate that the pseudogenes serve as a source of new hypervariable regions for the operon expression site of msp2.
MSP3, like MSP2, is a variable immunodominant surface protein belonging to a multigene family. BAC 2439 sequence data suggest that antigenic diversity of MSP3 is generated by the same mechanism as MSP2. The previously identified msp3-12 clone (8) appears to encode an authentic 5′ end for the gene and is truncated toward the 3′ end, whereas msp3-11 and 3-19 (8) start at aa positions 224 and 187, respectively, relative to msp3-12 (Fig. 5). When these msp3 sequences are spliced in silico they do not encode a fused domain of the appropriate size (86 kDa). The two msp3 genes found in this study (msp3-1 and msp3-2) start at position 130 relative to msp3-12. These two coding sequences start with the sequence PYQGYHSMLTALE and continue for 51 aa with 100% identity to the beginning of the msp2 pseudogenes (Fig. 5). Although these four putative pseudogenes (msp3-1, -2, -11, and -19) are quite divergent, they have two regions of high sequence similarity: a stretch of 93 aa of 100% sequence identity near the amino terminus, following the region of identity to msp2, and a stretch of 61 aa near the carboxy terminus (95–100% identity). Like msp2, these pseudogenes encode a portion of a 5′ conserved region, a divergent region with changes in sequence and length, and a 3′ conserved region. An additional similarity is the 600 bp of the 5′ flanking region that is highly conserved for msp2 pseudogenes 1 and 3 and for msp3 pseudogenes 1 and 2. The same 600-bp region 5′ to the msp2 and msp3 pseudogenes is a substantially sized repeat that likely has a role in the recombination of both msp2 and msp3 pseudogenes. As the 5′ regions of the msp3 and msp2 pseudogenes are the same, specificity for each target expression site must be ensured by the respective 3′ recombinatorial site in the coding region of each gene.
Figure 5.

Alignment of the deduced amino acid sequences of known msp3 (pseudo)genes in comparison with msp2. The gray region highlights a region of sequence similarity between msp2 and msp3. The black background indicates regions of sequence conservation in msp3 flanking the hypervariable region. The hypervariable region of msp3 from alignment position 301–909 is not shown.
Recombination of surface proteins to generate antigenic diversity occurs in several other bacterial pathogens. Tick-borne pathogens of the genus Borrelia undergo recombination of the vmp/vls genes (1, 2). Two distinct features of this system are the telomeric placement of the expression site for each of these genes on a linear plasmid and the tandem array of the pseudogene reservoir. In contrast, the msp2/msp3 system of antigenic diversity differs from Borrelia in that A. marginale contains no extrachromosomal plasmid, and the pseudogenes are distributed throughout the genome. Perhaps more similar to the antigenic variation of A. marginale are the non-tick-transmitted pathogens Neisseria gonorrhoeae (13) and Mycoplasma (14, 15). These bacteria have pseudogenes distributed throughout the genome, with more than one type of pseudogene for a given full-length gene.
The agent of human granulocytic ehrlichiosis is the closest relative to A. marginale in ehrlichial genogroup II. Human granulocytic ehrlichiosis has an ortholog of msp2 called p44 or human granulocytic ehrlichiosis msp2 (blastp value e−100), which is also encoded by a multigene family (16). Several expressed hypervariable regions of p44 were detected by reverse transcription–PCR, cloning, and sequencing (16). Cloning of the genomic counterpart for these expressed hypervariable regions resulted in two genomic clones (p44-15 and 18) that were truncated at both the 5′ and 3′ ends and did not have consensus promoters. Although these two sequences did not start or end in the same positions relative to each other or to a full-length p44, the pattern of pseudogenes distributed throughout the genome appears to be similar to A. marginale msp2. Despite the fact that human granulocytic ehrlichiosis infects host species and cell types different from those infected by A. marginale, immune evasion is likely achieved through the same method of recombination from widely distributed pseudogenes into a functional expression site.
There are two unique features to the A. marginale antigenic variation system described here: the concerted appearance of pseudogenes for two different gene families and the 600 bp of the highly conserved 5′ flanking region. Repeats of this length are highly unlikely to exist in bacteria (17), and that this repeat should be so highly conserved when not encoding a functional gene product indicates that the composition is important. The juxtaposition of this repeat next to the pseudogenes for two gene families that undergo an extremely high rate of recombination is surely important. This arrangement of two pseudogenes in close proximity and the potential for these two gene families to use the same recombinatorial mechanism is a system that allows a greater potential for antigenic variation from a small genome.
Acknowledgments
We thank Lowell Kappmeyer for excellent technical assistance, Ralph Horn for animal management, and Wendy C. Brown and Fred R. Rurangirwa for helpful discussion. BAC library construction and BAC sequencing were provided by Amplicon Express (Pullman, WA). This work was supported by National Institutes of Health Grants RO1 AI44005 and RO1 AI45580 and U.S. Department of Agriculture Grants USDA-ARS-CRIS 5348-32000-012-00D and USDA-SCA 58-5348-8-044.
Abbreviations
- BAC
bacterial artificial chromosome
- msp
major surface protein
Footnotes
References
- 1.Zhang J-R, Hardham J M, Barbour A G, Norris S J. Cell. 1997;89:275–285. doi: 10.1016/s0092-8674(00)80206-8. [DOI] [PubMed] [Google Scholar]
- 2.Barbour A G. Trends Microbiol. 1993;1:236–239. doi: 10.1016/0966-842x(93)90139-i. [DOI] [PubMed] [Google Scholar]
- 3.Yu X-J, McBride J W, Zhang X-F, Walker D H. Gene. 2000;248:59–68. doi: 10.1016/s0378-1119(00)00147-5. [DOI] [PubMed] [Google Scholar]
- 4.Reddy G R, Streck C P. Mol Cell Biol Res Commun. 1999;1:167–175. doi: 10.1006/mcbr.1999.0133. [DOI] [PubMed] [Google Scholar]
- 5.Ohashi N, Unver A, Zhi N, Rikihisa Y. J Clin Microbiol. 1998;36:2671–2680. doi: 10.1128/jcm.36.9.2671-2680.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sulsona C R, Mahan S M, Barbet A F. Biochem Biophys Res Commun. 1999;257:300–305. doi: 10.1006/bbrc.1999.0459. [DOI] [PubMed] [Google Scholar]
- 7.Palmer G H, Eid G, Barbet A F, McGuire T C, McElwain T F. Infect Immun. 1994;62:3808–3816. doi: 10.1128/iai.62.9.3808-3816.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Alleman A R, Palmer G H, McGuire T C, McElwain T F, Perryman L E, Barbet A F. Infect Immun. 1997;65:156–163. doi: 10.1128/iai.65.1.156-163.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Alleman A R, Kamper S M, Viseshakul N, Barbet A F. J Gen Microbiol. 1993;139:2439–2444. doi: 10.1099/00221287-139-10-2439. [DOI] [PubMed] [Google Scholar]
- 10.Barbet A F, Lundgren A, Yi J, Rurangirwa F R, Palmer G H. Infect Immun. 2000;68:6133–6138. doi: 10.1128/iai.68.11.6133-6138.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Alleman A R, Barbet A F. J Clin Microbiol. 1996;34:270–276. doi: 10.1128/jcm.34.2.270-276.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chapter 2. (1997) in Genome Analysis: A Laboratory Manual, eds. Birren, B., Green, E. D., Klapholz, S., Myers, R. M. & Roskams, J. (Cold Spring Harbor Lab. Press, Plainview, NY), pp. 136–138.
- 13.Haas R, Meyer T F. Cell. 1986;44:107–115. doi: 10.1016/0092-8674(86)90489-7. [DOI] [PubMed] [Google Scholar]
- 14.Kenri T, Taniguchi R, Sasaki Y, Okazaki N, Narita M, Izumikawa K, Umetsu M, Sasaki T. Infect Immun. 1999;67:4557–4562. doi: 10.1128/iai.67.9.4557-4562.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Noormohammadi A H, Markham P F, Kanci A, Whithear K G, Browning G F. Mol Microbiol. 2000;35:911–923. doi: 10.1046/j.1365-2958.2000.01766.x. [DOI] [PubMed] [Google Scholar]
- 16.Zhi N, Ohashi N, Rikihisa Y. J Biol Chem. 1999;274:17828–17836. doi: 10.1074/jbc.274.25.17828. [DOI] [PubMed] [Google Scholar]
- 17.Rocha E P C, Danchin A, Viari A. Res Microbiol. 1999;150:725–733. doi: 10.1016/s0923-2508(99)00120-5. [DOI] [PubMed] [Google Scholar]