

Abstract
Background
In the current study, we have investigated whether low density lipoprotein receptor knockout mice (LDLR-KO), moderate oxidative stress model and cholesteremia burden display gastroparesis and if so whether nitrergic system is involved in this setting. In addition, we have investigated if sepiapterin (SEP) supplementation attenuated impaired nitrergic system and delayed gastric emptying.
Methods
Gastric emptying and nitrergic relaxation were measured in overnight fasting mice. nNOSα dimerization, anti-oxidant markers such as Nrf2, GCLM, GCLC, HO-1, catalase (CAT) and superoxide dismutase (SOD1) were measured using standard methods. Biopterin levels and intestinal transit time were measured using HPLC and dye migration assay, respectively. Wild type (WT) and LDLR-KO were supplemented with SEP.
Key Results
In LDLR null stomachs, 1) significant reduction in rate of gastric emptying, gastric pyloric and fundus nitrergic relaxation & nNOSα dimerization, 2) elevated oxidized biopterins and reduced ratio of BH4/BH2+B, 3) reduced Nrf2 and GCLC protein expression & no change in GCLM, HO-1, Cat, Sod1 and 4) accelerated small intestinal motility were noticed. Supplementation of SEP restored delayed gastric emptying, impaired pyloric and fundus nitrergic relaxation with restoration of nNOS dimerization and nNOS expression.
Conclusions and Inferences
This novel data suggests that hyperlipidemia and/or suppression of selective antioxidants may be a potential cause of developing gastroparesis in diabetic patients.
Keywords: Gastroparesis, LDLR-KO, nitrergic relaxation, nNOS dimerization, Nrf2, sepiapterin
INTRODUCTION
Low-density lipoprotein (LDL) is a type of lipoprotein that transports cholesterol and triglycerides from liver to peripheral tissues1. The LDL receptor (LDLR) is a cell-surface receptor that mediates the endocytosis of cholesterol-rich intermediate density lipoproteins (IDL) and LDL from plasma.2 Deficiency of LDLR display elevated plasma cholesterol level, IDL and LDL without a significant change in high density lipoproteins and triglyceride levels.3, 4 which increases lipid storage in the body system1–3 that may eventually predisposes to oxidative stress and diabetes.5
Gastroparesis is a clinical condition which is associated with abnormal gastric motility and is affecting predominantly young women.6 Gastric motility is regulated in large part by the enteric nervous system (ENS) mainly, excitatory (acetylcholine) and inhibitory system [nitric oxide (NO) and vasoactive intestinal peptide].7 NO produced by neuronal nitric oxide synthase (nNOS) and its importance in gastric function has been established by the findings of pyloric hypertrophy and gastric dilation in nNOS−/− mice.8, 9 The catalytic activity of NOS depends on dimerization with help of (6R)-tetrahydrobiopterin (BH4).10, 11 Enzymatic uncoupling of NOS or reduced NOS dimerization due to lack of BH4 may in part account for lower NO production and increased oxidative stress factors such as superoxide.12 We have recently reported that BH4 synthesis is reduced in diabetic female and leading to impaired nNOS activity and delayed gastric emptying for solids which has been restored by supplementation of BH4.13, 14
Nrf2 (NF-E2-related factor 2) is a transcription factor that protects the cells from oxidative stress by activating the phase II antioxidant enzymes (Glutamate-cysteine ligase catalytic, (GCLC) and modifier (GCLM) subunits and HO1) through ARE (antioxidative responsive element.15 Recent work suggests that the Nrf2 is critical for protecting GI tract against disease by regulating a multifaceted cellular antioxidant defense.16, 17 Studies also have shown that NO donors activates the transcription up-regulation of phase II enzymes, through Nrf2 via ARE in vascular endothelial cells.18
Recently it has been reported that VLDLR expression in skeletal muscle and heart is reduced in streptozotocin (STZ) induced diabetic rats while there was no change in LDL-receptor expression, as assessed in liver.19 However, mice with STZ induced diabetes and LDL receptor defects often develop severe hyperlipidemia.20, 21 It has also been reported that the severe elevation of serum cholesterol and TG concentrations in STZ-induced diabetic rats.19 And it is a known fact that exaggerated postprandial hypertriglyceridemia is common in diabetic patients and this may also contribute for the development of gastroparesis.5
Treatment of diabetic vascular endothelial cells with sepiapterin (SEP) (the BH4 precursor in the salvage pathway) significantly improves NO synthesis and cholinergic vascular relaxation in diabetic mice.22 Recently, we have reported that, sepiapterin reverses the changes in gastric nNOS dimerization and function in diabetic gastroparesis23. However, there have been no studies on the effects of SEP in LDLR-KO mice associated with gastroparesis. In the present study, we investigated whether deficiency of LDLR is involved in the pathogenesis of gastroparesis through nNOS uncoupling due to altered biopterin levels associated with antioxidant(s) suppression and if so supplementation of SEP attenuated impaired gastric nitrergic relaxation and delayed gastric emptying.
MATERIALS AND METHODS
Experimental mice
Adult female C57BL LDLR-KO homozygous (−/−) (12 wk old) and their wild type (WT) (+/+) were procured from Jackson laboratories (Sacramento, CA) and maintained in the institutional animal care facility under controlled temperature, humidity and light-dark cycle (12:12 h), with free access to rodent chow and water. The protocols for using these mice were approved by the Institutional Animal Care and Use Committees at the Meharry Medical College, Nashville, Tennessee, in accordance with the recommendations of National Institutes of Health, Guide for the Care and Use of Laboratory Animals. WT and LDLR-KO animals were provided with SEP tablets (20 mg/Kg body wt/day) for 11 days. Sepiapterin tablets were given daily to the animal(s) those were placed individually in a separate cage (1 gram tablet made in the diet with chocolate flavor). These tablets were given to each animal every day in the morning hours (between 9.00 AM to 10 AM) prior to providing their regular rat chow. On the last day of SEP (Schircks Laboratories, Switzerland) supplementation, gastric emptying and gastric fundus nitrergic relaxation studies were performed. Animals were sacrificed by cervical dislocation to collect gastric fundus and antrum muscular tissue for future analysis. Tissue samples collected from animals were snap frozen in liquid nitrogen and stored at −80°C until analyzed.
Solid gastric emptying studies
Solid gastric emptying studies were performed as reported earlier.13, 14 Briefly, after fasting for overnight known amount of normal diet was fed to the animals for 3 h. At the end of 3 h, the remaining food was collected and calculated the amount of food intake. Then animals were fasted again for 2 h without food and water. At the end of fasting, animals were sacrificed and gastric tissues were isolated and measured the weight of the whole stomach. Then food contents were removed by opening the stomach and measured the empty stomach weight. The rate of gastric emptying was calculated according to the following equation: gastric emptying (% in 2h) = (1 - gastric content food intake−1) × 100. Following this protocol, we have noticed that 4 out of 5 animals displayed delayed gastric emptying.
Measurement of glucose
Drop of blood was collected from the tail vein fasted (2 h, 12 h) LDLR-KO and WT mice without anesthesia for determination of plasma glucose levels using Alphatrack™ Blood monitoring pack.
Measurement of stool frequency and weight
Small intestinal motility was measured according to the method of Li et al.24 Briefly, each mice was placed in a clear plastic cage for 1 h. Stools were collected immediately after expulsion and placed in sealed tubes. The total stools were weighed to provide a wet weight, then dried overnight at 65°C and weighed again to provide a dry weight.
Small intestine transit
Following a 12 h fast, each animal was administered 0.1 mL of methylene blue labeled 10 % glucose by oral gavage. After sacrifice (0–30 min after gavage) the animals, stomachs were clamped with a string tied both at the lower esophageal sphincter and the pylorus to prevent dye leakage. The small intestine was removed and intestine transit was assessed by measuring the distance the dye front had traveled from the pylorus.25, 26
Measurement of biopterin levels
Determination of Biopterins in Gastric Antrum Muscular Tissue
Biopterin levels were determined in antrum homogenates by HPLC followed by electrochemical and fluorescent detection, as described previously.27 Briefly, samples were homogenized in phosphate-buffered saline (50 mM L−1), pH 7.4, containing dithioerythritol (1 mM L−1) and EDTA (100 μM L−1). Following centrifugation (15 min at 13,000 rpm and 4 °C), the samples were transferred to new, cooled micro tubes and precipitated with cold phosphoric acid (1 M L−1), trichloroacetic acid (2 M L−1), and dithioerythritol (1 mM L−1). The samples were vigorously mixed and then centrifuged for 15 min at 13,000 rpm and 4 °C. The samples were injected onto an isocratic HPLC system and quantified using sequential electrochemical (Coulochem III, ESA Inc.) and fluorescence (Jasco) detection. HPLC separation was performed using a 250 mm, ACE C-18 column (Hichrom) and mobile phase comprising of sodium acetate (50 mM L−1), citric acid (5 mM L−1), EDTA (48 μM L −1), and dithioerythritol (160 μM L−1) (pH 5.2) (all ultrapure electrochemical HPLC grade) at a flow rate of 1.3 mL/min. Background currents of +500 μA and −50 μA were used for the detection of BH4 on electrochemical cells E1 and E2, respectively. 7,8-BH2 and biopterin were measured using a Jasco FP2020 fluorescence detector. Quantification of BH4, BH2, and biopterin was done by comparison with authentic external standards and normalized to sample protein content.
Organ Bath Studies
Electric field stimulation (EFS)-induced NANC relaxation was studied in circular gastric fundus and pylorus strips as reported earlier.13, 14 Strips were tied with silk thread at both ends and were mounted in 10 ml water-jacketed organ baths containing Krebs buffer at 37°C and continuously (Danish Myo Technology, USA). Tension for each muscle strip bubbled with 95% O2 - 5% CO2 was monitored with an isometric force transducer and analyzed by a data acquisition system (Power Lab, ADI, USA). A passive tension equal to 2 g was applied on each strip in the 1h equilibration period through an incremental increase (0.5 g, four times, at 15 min interval). Strips were exposed to atropine, phentolamine and propranolol (10 μM each) in bath solution for 1 h to block cholinergic and adrenergic responses. 5-hydroxytryptamine (100 μM) pre-contracted strips were exposed to EFS (90V, 2Hz, 1-ms pulse for duration of 1 min) to elicit NANC relaxation by low frequency (2 Hz) The NO dependence of nitrergic relaxations was confirmed by preincubation with NG-nitro-L-arginine methyl ester (L-NAME, 100 μM; 30 min). At the end of each experiment, the muscle strip was blotted dry with filter paper and weighed. Comparisons between groups were performed by measuring the area under the curve (AUC mg tissue−1) of the EFS-induced relaxation (AUCR) for 1 min and the baseline for 1 min (AUCB) according to the formula (AUCR – AUCB) weight of tissue (mg)−1 = AUC mg of tissue−1.
Western blot analysis
nNOSα, GTPCH-1, DHFR, Nrf2, GCLC, GCLM and HO-1 protein was quantified in gastric antrum homogenates from the 2 groups using standard western blot analysis, as described in our previous study.13, 14 Proteins were measured by Bio-Rad protein assay (Bio-Rad, Hercules, CA) and 30 μg protein was separated by SDS polyacrylamide gel electrophoresis (SDS-PAGE). The membrane was immunoblotted with polyclonal nNOSα (Zymed Laboratories Inc., CA), Nrf2, GTPCH-1, DHFR, GCLC, and GCLM primary antibodies (Santacruz Biotechnology, CA) and anti-rabbit IgG conjugated with horseradish peroxidase (Sigma Chemical, St. Louis, MO) as secondary antibody. Binding of antibodies to the blots was detected with enhanced chemiluminescence system (ECL, Amersham Pharmacia Biotech, Piscataway, NJ) following manufacturer’s instructions. Stripped blots were re-probed with γ-tubulin specific polyclonal antibodies (Sigma Chemical, St. Louis, MO) to enable normalization of signals between samples. Band intensities were analyzed using Bio-Rad Gel Doc (Bio-Rad, Hercules, CA).
nNOSα dimerization in mice gastric antrum
Levels of nNOSα monomer and dimer were quantified by western blotting via Low temperature (LT)-PAGE in gastric antrum homogenates as described previously.6 LT-SDS-PAGE was performed on ice. 30 μg of protein in standard Laemmli buffer was incubated at 0°C for 30 min and then separated by using a 6% separating gel. All gels and buffers were pre-equilibrated to 4°C prior to electrophoresis and the buffer tank placed in an ice-bath during electrophoresis to maintain the gel temperature below 15°C. A polyclonal antibody specific to nNOSα (Zymed Laboratories) and anti-rabbit IgG conjugated with horseradish peroxidase (Sigma Chemical, St. Louis, MO) were used as the primary and secondary antibodies, respectively.
Activity of antioxidant enzymes in LDLR-KO mice
Stomach antrum was homogenized in cold 50 mM potassium phosphate, pH 7.0. The homogenate was centrifuged at 10,000 × g and 4°C for 15 min and the supernatant was separated to measure total protein and the activities of catalase (CAT). Cat activity in was assayed by a commercially available kit from Cayman Chemical Company (Ann Arbor, MI) and the data were expressed as nmol min−1 mg protein−1. For total superoxide dismutase (SOD1) activity, tissue was homogenized in 20 mM HEPES buffer, pH 7.2. The homogenate was centrifuged at 1,500 × g for 5 min at 4°C and the supernatant was separated to measure total protein and the activities of Sod1. Sod1 activity in stomach antrum was assayed by a commercially available kit from Cayman Chemical Company (Ann Arbor, MI) and the results were expressed as U mg protein−1.
Statistics
Data were presented as mean ± standard error (SE). Statistical comparisons between groups were determined by Student’s t-test using Graph pad prism Version 5.0 (Graph pad software, San Diego, CA). A p value of less than 0.05 was considered statistically significant.
RESULTS
Body weights and glucose levels in LDLR-KO mice
We first investigated the body weights and glucose levels difference among the female LDLR-KO and age-matched WT mice. As shown in Table 1, no significant difference in body weight was observed in LDLR-KO mice compared to wild type mice. Compared to WT mice, LDLR-KO mice showed slightly higher plasma glucose levels under fasted conditions (2 h and 12 h), and suggesting impaired glucose tolerance in LDLR-KO mice (Table 1).
Table 1.
Blood glucose level and body weight in the wild type and LDLR-KO female mice
| WT | KO | |
|---|---|---|
| Body Weight, g | 18.5 ± 1.0 | 21.0 ± 0.7 |
| Blood Glucose, mg/dl | ||
| 2 h | 187.2 ± 11.1 | 225.5 ± 5.0* |
| 12 h | 97.4 ± 3.7 | 125.0 ± 11.9* |
Wild type (WT); Knock out (KO). Results are expressed as mean ± SEM (N = 5).
p < 0.05 compared to WT.
Delayed gastric emptying in LDLR-KO mice
Fig. 1 shows the solid gastric emptying (% in 2 hours) in female LDLR-KO mice. A significant delay (58 ± 6.11) in the solid gastric emptying was observed in LDLR-KO mice compared to WT (78 ± 0.88) animals. Our results show that supplementation of sepiapterin (79.30 ± 2.97) restored delayed gastric emptying in LDLR-KO mice whereas no significant change in sepiapterin treated WT mice (75 ± 0.33).
Fig.1.

Solid gastric emptying in female wild type (WT), LDLR-KO, sepiapterin (SEP) supplemented WT mice and sepiapterin (SEP) supplemented LDLR-KO mice. The values are mean ± SE for four animals in each group. Statistical significance was determined by Student t-test. Values are mean ± SE (n = 4). *p<0.05 compared with WT animals and #p<0.05 compared with LDLR-KO animals.
Transient increase in small intestinal motility in LDLR-KO mice
The LDLR-KO mice didn’t show any change in stool frequency in one hour after 12 h fasting than WT animals (Fig. 2A). Similarly, the stool weight also not much differ among LDLR-KO and WT mice (Fig. 2B). Whereas the intestine transit time was significantly increased in LDLR-KO mice when compared with WT mice at the time point of 5 min, interestingly it didn’t show much difference at 30 min interval (Fig. 2C).
Fig. 2.



(A). Stool frequency was monitored in LDLR-KO mice and wild type (WT) mice for one hour period. Since the small intestinal functions to remove water, this confirms the utility of 1 h stool frequency as a measure of colon transit time. (B) The stool weight was collected during one hour period and it expressed weight in gm in LDLR-KO and WT mice. (C) The intestine transit time was monitored by measurement of dye migration in the colon. Statistical significance was determined by Student t-test. Values are mean ± SE (n = 5). *p < 0.05 compared with WT animals.
Elevated oxidized biopterin levels in LDLR-KO mice
Fig. 3A shows the levels of total biopterins (BH4, BH2 and B) in WT and LDLR-KO mice. No significant change was observed in BH4 level, whereas a significant increase in BH2 as well as B levels was observed in LDLR-KO mice compared to WT mice. The ratio of BH4 and total biopterin was significantly reduced in LDLR-KO mice compared to WT animals. No significant change in the expression of GTPCH-1 and DHFR was observed in LDLR-KO mice (Figs. 3B & C)
Fig. 3.
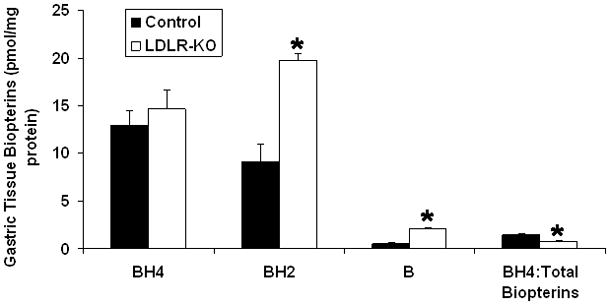
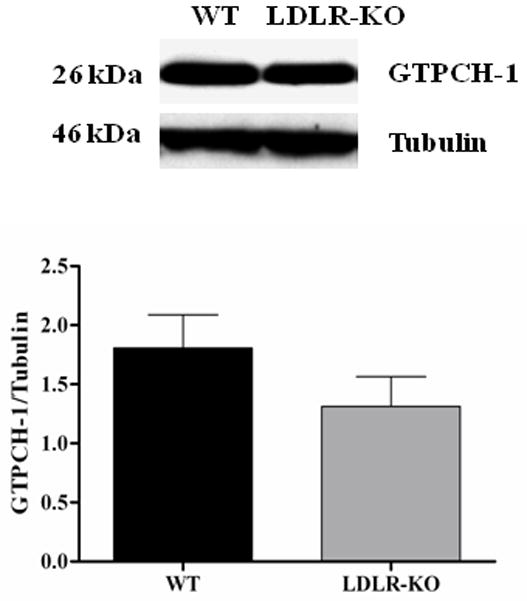
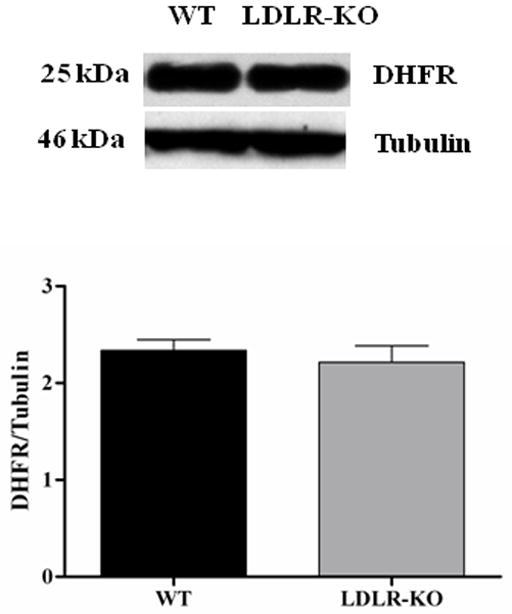
(A) Levels of total biopterins in LDLR-KO female mice gastric tissue. Biopterin levels were measured in wild type (WT) and LDLR-KO female mice gastric antrum tissue using HPLC. (B) Representative immunoblot and densitometric analysis data for protein expression of BH4 synthesizing enzymes GTPCH-1 and (C) DHFR. Statistical significance was determined by Student t-test. Values are mean ± SE (n = 4). *p < 0.05 compared with WT animals.
Impairment of gastric fundus and pyloric nitrergic relaxation in LDLR-KO mice
Fig. 4A shows the nitrergic relaxation in fundus muscular tissue following EFS (2 Hz) in WT, SEP supplemented WT mice, LDLR-KO mice and sepiapterin (SEP) supplemented LDLR-KO mice. The gastric fundus nitrergic relaxation was impaired in female LDLR-KO mice compared to WT mice (−0.28 ± 0.01 vs. −0.12 ± 0.02). Whereas, supplementation of SEP resulted in complete reversal (−0.23 ± 0.02) of impaired nitrergic relaxation in female LDLR-KO mice, but no effect on wild animals (−0.25 ± 0.02). Fig. 4B shows the nitrergic relaxation in pyloric muscular tissue following EFS (2 Hz) in WT, SEP supplemented WT mice, LDLR-KO mice and sepiapterin (SEP) supplemented LDLR-KO mice. The gastric pyloric nitrergic relaxation was impaired in female LDLR-KO mice compared to WT mice (−0.02 ± 0.02 vs. −0.20 ± 0.07). Whereas, supplementation of SEP (−0.13 ± 0.07) significantly (p<0.05) reversed of impaired nitrergic relaxation in female LDLR-KO mice, but no effect on wild animals (−0.19 ± 0.07).
Fig. 4.
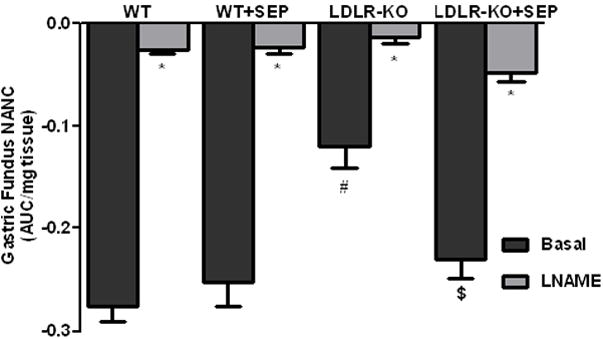
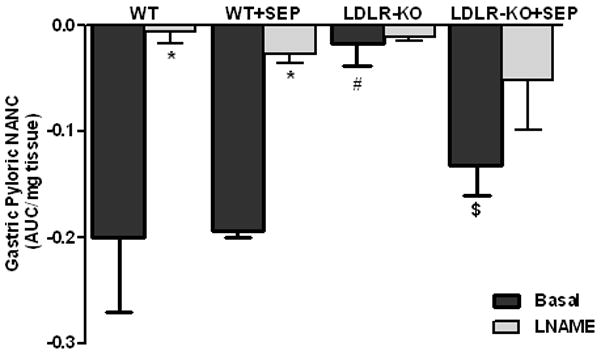
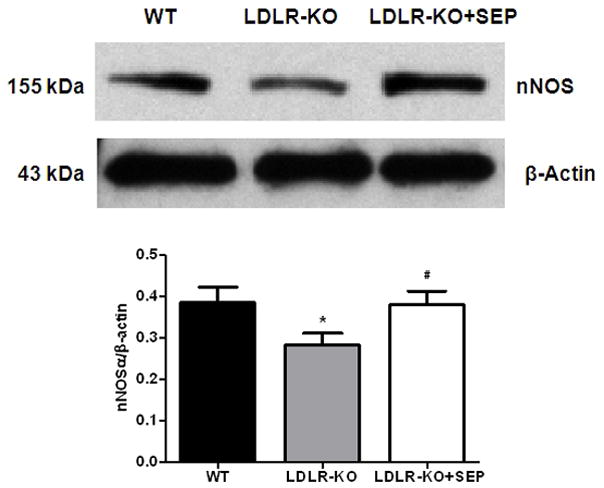
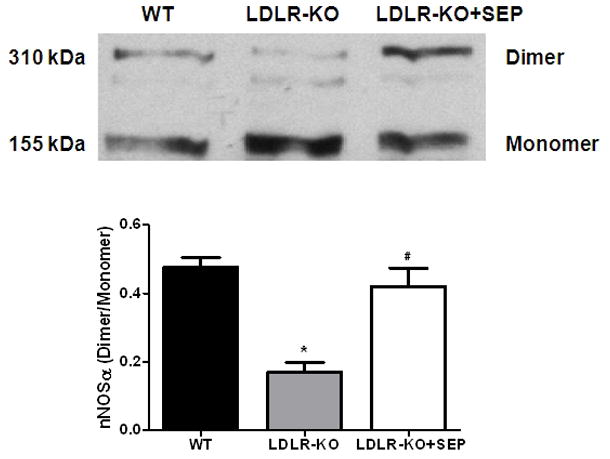
(A) Impairment of nitrergic relaxation in female wild type (WT), SEP supplemented WT mice, LDLR-KO and sepiapterin (SEP) supplemented LDLR-KO mice gastric fundus muscular tissues. (B) Impairment of nitrergic relaxation in female wild type (WT), SEP supplemented WT mice, LDLR-KO and sepiapterin (SEP) supplemented LDLR-KO mice gastric pylorus muscular tissues. The nitric oxide (NO) dependence of the NANC relaxations in all groups was confirmed by preincubation (30 min) with the NO inhibitor nitro-L-arginine methyl ester (L-NAME; 100 μM). The values are mean ± SE of four samples in each group. Statistical significance was determined by Student t-test. Values are mean ± SE (n = 4), * p < 0.05 compared with the response in the absence of L-NAME, # p < 0.05 compared with WT animals and $ p < 0.05 compared with LDLR-KO animals. nNOSα protein expression and dimerization in LDLR-KO female mice gastric antrum tissues (C) Representative immunoblot and densitometric analysis data for nNOSα protein expression. (D) Representative immunoblot and densitometric analysis data for nNOSα protein dimerization. Statistical significance was determined by Student t-test. Values are mean ± SE (n = 4), *p < 0.05 compared with WT animals and # p < 0.05 compared with LDLR-KO animals.
Decreased gastric antrum nNOSα protein expression and dimerization in LDLR-KO mice
According to Fig. 4C the protein level of nNOSα, the only functional isoform of nNOS in gastric antrum muscular tissue, was significantly (p<0.05) decreased (0.28 ± 0.03 vs 0.39 ± 0.04) in LDLR-KO mice when compared to WT whereas, supplementation of SEP resulted in reversal of nNOS expression (0.38 ± 0.03) in female LDLR-KO mice. To measure whether decrease in the nNOSα was the result of altered nNOS dimer levels in LDLR-KO mice, we performed the dimerization study by LT-PAGE gel. As depicted in Fig. 4D, a significant (p<0.05) decrease in the dimer/monomer ratio was seen in LDLR-KO mice compared to WT mice (0.17 ± 0.03 vs 0.48 ± 0.03) whereas, its restored in SEP supplemented LDLR-KO (0.42 ± 0.05) animals.
Reduced gastric antrum expression of Nrf2 in LDLR-KO mice
Fig. 5A demonstrates the expression of Nrf2 in LDLR-KO mice. The protein level of Nrf2 was reduced significantly in LDLR-KO mice (0.24 ± 0.04 vs. 0.62 ± 0.01) when compared to WT mice.
Fig. 5.
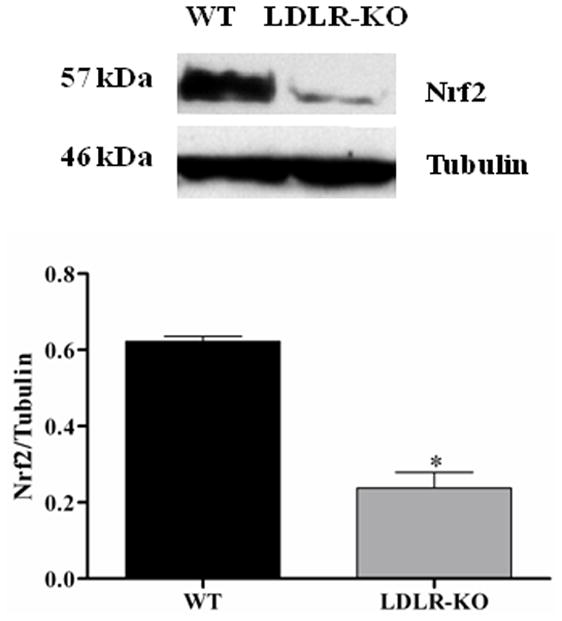
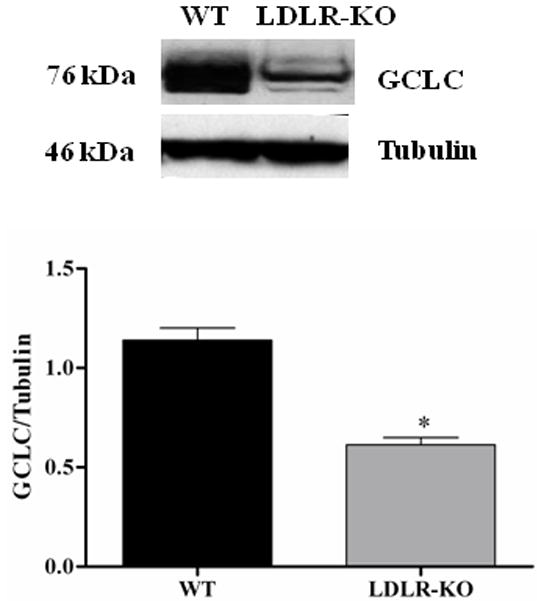
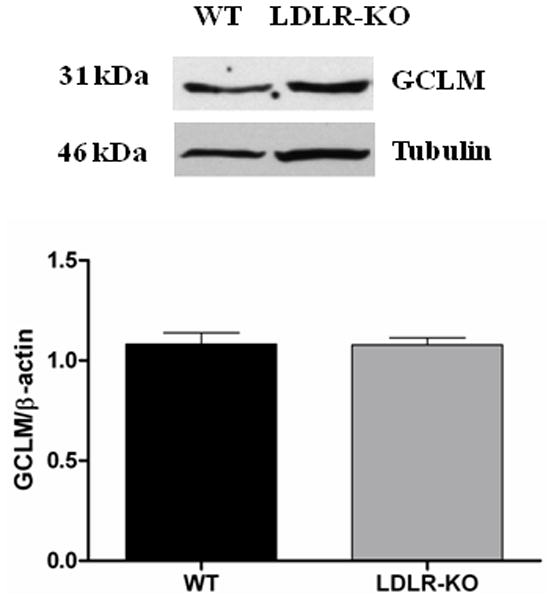
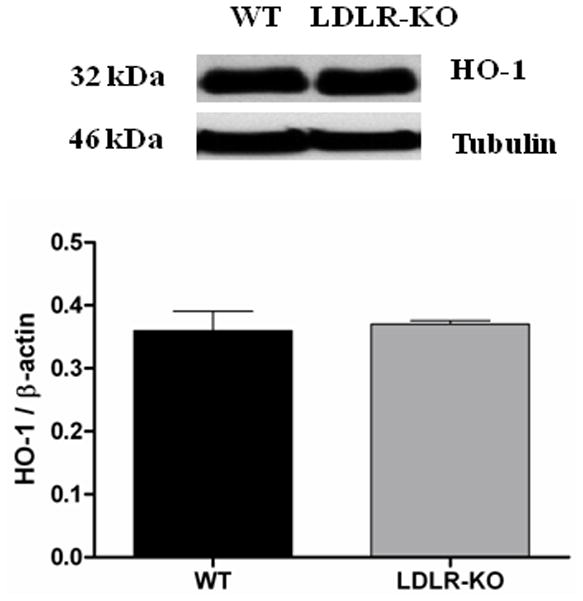
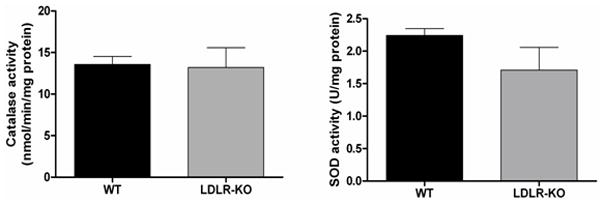
Nrf2 and GCLC protein expression in LDLR-KO female mice gastric antrum tissues. (A) Representative immunoblot and densitometric analysis data for Nrf2 protein expression. (B-D) Representative immunoblot and densitometric analysis data for GCLC, GCLM and HO-1 protein expression. The values are mean ± SE of three to four samples in each group. (E) CAT activity, and SOD1 activity in LDLR-KO female mice gastric antrum tissues. Values are mean ± SE (n = 4). Statistical significance was determined by student t-test. *p < 0.05 compared with wild type (WT) animals.
Reduced gastric antrum expression of GCLC in LDLR-KO mice
The protein levels of GCLC in WT and LDLR-KO mice were shown in Fig 5B. The protein level of GCLC was significantly reduced in LDLR-KO mice (0.6 ± 0.03 vs. 1.14 ± 0.06) when compared to WT, a phase II enzyme whose expression is regulated by Nrf2. However, no significant change in the protein expression of GCLM and HO-1 was observed in LDLR-KO mice (Figs 5C & 5D).
Activity of antioxidant enzymes (CAT & SOD1) in LDLR-KO female mice
SOD1 is a family of ubiquitous antioxidant enzymes that catalyze the conversion of superoxide anion radicals to hydrogen peroxide (H2O2) and molecular oxygen, whereas, CAT is a ubiquitous antioxidant enzyme that is involved in the conversion of H2O2 to molecular oxygen and water. No significant change was observed in both the activities of CAT as well as SOD1 in LDLR-KO mice compared with age matched WT animals (Fig. 5E).
DISCUSSION
Gastroparesis is a disorder of delayed gastric emptying, with the most common known cause being diabetes. Recently using streptozotocin-induced diabetic model (Type 1 diabetes in humans), we have reported that impairment in nitrergic relaxation and reduction in nNOS dimerization (critical of enzyme activity and subsequent nitric oxide production), enzyme activity and altered biopterin levels are responsible for developing gastroparesis in female rats.13, 14 Hyperlipidemia as well as oxidative stress may be some of the detrimental causes for observed impaired gastric motility dysfunctions associated with diabetes and it has not been addressed previously. Oxidative stress which not only interplays with NO ability to activate sGC (soluble guanylate cyclase) but also NO bioavailability because superoxide anion quenches NO, transforming it into peroxynitrite.28
This is the first comprehensive report of gastrointestinal dysmotility in LDLR-KO mice, an animal model of hyperlipidemia. The elevation of hyperlipidemia status and its associated molecules in the GI tract perhaps may leads to consistently depletion of nitrergic function. The accelerated small intestinal motility observed in LDLR-KO mice (Fig. 2) is consistent with the loss of nitrergic neuronal function in the GI tract.13, 14 However; additional studies are warranted to investigate the role of nitrergic neurons in this setting.
Although, elevated hyperlipidemia is one of the detrimental factor for diabetes induced complications; to date there are no literature available if this impairs biopterin and nitrergic system and thus leading to gastroparesis. In the current study, we have first investigated whether LDLR-KO mice display gastroparesis and if so nitrergic system is involved in gastric emptying. The present study demonstrates that gastric emptying is delayed in female LDLR-KO mice compared to WT mice and it’s restored by supplementation with SEP (Fig. 1). In addition, our data demonstrates that impaired nitrergic relaxation observed in LDLR-KO mice was restored by SEP supplementation (Fig. 4A & B). Since the predominant form of nNOS in the enteric nerves of the stomach is nNOSα, we next investigated if the expression and dimerization of nNOS is altered in LDLR-KO mice.13 As shown in Fig 4, both nNOSα expression (Fig. 4C) and nNOSα dimerization (Fig. 4D) was impaired in LDLR-KO mice; supplementation of SEP restored this. The above data for the first time suggest that, hyperlipidemia causes impairment in nitrergic mediated gastric motility and leads to gastroparesis; SEP treatment may be beneficial to regulate the gastric motility by modulating the lipid metabolism. Further studies are warranted to address this in detail.
BH4, an essential co-factor for NOS is intracellularly produced from GTP via GTP-cyclohydrolase I (GTPCH-1) or, alternatively, from SEP via salvage pathway and acts as a redox switch in the oxygenase domain of NOS.29 Reduced levels of BH4 impair the production of NO, and leads to increased superoxide radical production, due to nNOS uncoupling. The superoxide radical then reacts with NO resulting into the production of peroxynitrite. This further reduces biological availability of NO.30 BH4 deficiency has been associated with diabetic complications including gastroparesis.14 This information provides a rational basis for the use of supplemental sepiatperin (SEP; a precursor for BH4 biosynthesis via salvage pathway) in diabetic conditions. In this study, we demonstrate that supplementation of SEP normalizes both delayed gastric emptying (Fig. 1) and impaired nitrergic relaxation in female LDLR-KO mice (Fig. 4). This is in good agreement with our previous study, where we showed that supplementation of BH4 or SEP restored the delayed gastric emptying associated with diabetes in female rats.13, 14, 23 Though BH4 levels have not been altered in LDLR-KO mice, the finding that significant increase in the oxidized biopterin levels followed by a decrease BH4:BH2+B level (Fig. 3A) may ultimately reduce the BH4 bioavailability. Several studies support the notion that a decrease in the ratio of BH4:BH2 without changing in absolute BH4 levels are associated with increased vascular diseases.22, 31 Elevated oxidized biopterins (BH2, B) not only reduces the bioavailability of NOS cofactor BH4, but also competes with BH4 to bind to NOS domain that may eventually down regulates the dimerization of the enzyme and function.14 In addition, GTPCH1 and DHFR gene knock out studies demonstrated that either major reductions in de novo BH4 biosynthesis or an altered BH4:BH2 ratio is sufficient to induce eNOS uncoupling.32 The data reported in the current studies also demonstrate that a reduction in the ratio of BH4 vs BH2+B may be responsible for decreased nNOS dimerization, nitrergic relaxation and thus leading to delayed gastric emptying.
To investigate the factors involved in reduced BH4 bioavailability we checked the expression of GTPCH (GTP cyclohydrolase, which converts GTP to 7,8-dihydroneopterintriphosphate) and DHFR (dihydro folate reductase which converts 7,8-dihydrobiopterin to BH4) in both control, LDLR-KO mice tissues. As shown in Figs. 3B & C, we did not observe any change in the expression of GTPCH, DHFR in LDLR-KO mice. Apart from these observations, our data indicate that in LDLR-KO mice gastric oxidized biopterin levels (BH2, B) but not BH4 have been elevated (Fig. 3A). Collectively, these results suggests that elevated hyperlipidemia status leads to increased gastric oxidized biopterins and nNOS uncoupling which results in impaired nitrergic relaxation and thus gastroparesis.
We also noticed a significant decrease in the expression levels of both Nrf2 as well as phase II enzymes, GCLC which is essential for the synthesis of glutathione (GSH) in LDLR-KO mice (Figs. 5A & B). Nrf2 is a transcriptional factor that protects the cells from oxidative stress by activating the antioxidant enzymes including GSH synthesis enzymes (GCLC, GCLM) which is critical for the protection of neuronal cells. Genes such as heme oxygenase-1 (HO-1), SOD1, CAT, and Gpx1 are representative of canonical antioxidant genes regulated by Nrf2. Recent work suggests that the Nrf2 is critical for protecting GI tract against disease by regulating a cellular antioxidant defense.16, 17 The above findings may suggest that reduced levels of phase II enzymes elevate oxidative stress and thus impairs both nitrergic system and gastric motility functions. We can also conclude that chronic depletion of LDLR may be one of the detrimental factors in the pathogenesis of gastroparesis due to increased hyperlipidemia. This is in good agreement with previous results which states that Nrf2 inhibits lipid accumulation and oxidative stress in mouse liver after feeding a high fat diet, probably by interfering with lipogenic and cholesterologenic pathways.33 Recent studies have shown that HO-1 reverses gastroparesis in Type 1 diabetic mice by protecting against oxidative stress.34, 35 However, data from LDLR-KO stomachs show that no change in the expression of antioxidant enzymes HO-1 (Fig. 5D) and activity of CAT & SOD1 (Fig. 5E) may explain that this is not a severe oxidative stress model; instead probably it is a moderate stress model. Previous studies have demonstrated that high fat fed LDLR-KO displays hyperglycemia, severe hypertriglyceridemia and elevated oxidative stress.3 The molecular mechanisms responsible for decreased nNOS dimerization and/or how LDL influences gastric motility functions are not currently understood. Recent studies have demonstrated that more than 50% of patients with idiopathic gastroparesis are obese or overweight.36 These studies further report that women had more gastroparesis symptoms than in men; however, the etiological connection is completely unknown.36 In addition, loss of ICC and/or reduced nNOS containing neurons that are noticed in diabetic rodents may also account for the impairment of stomach motility and delay in gastric emptying in LDLR null mice.37–38 Additional experiments are warranted to address in detail with regard to mechanisms of nNOS regulation as well as the potential role for NO on gastric smooth muscle function in both normal and high fat fed LDLR-KO mice. The results obtain by using LDLR-KO animal model may provide an opportunity to understand the molecular mechanisms that are involved in idiopathic gastroparesis.
Acknowledgments
We thank to Freeman Michael, PhD Konjeti, Rajasekhar, PhD (Vanderbilt University Medical Center) and Farrugia Gianrico, M.D (Mayo Clinic) for scientific discussions. Finally, we thank Shanthi Srinivasan, M.D and Anitha Mallappa (Emory University) for providing us the intestinal transit protocol as well as guidance on completing the transit experiments.
Financial Support for the original research was provided by the NIH-NIDDK R21DKO76704 (PG), P60DK020593 pilot project funds (PG) and from RCMI G12RR03032 (PG), as start-up funds at Meharry Medical College, Nashville, TN, USA.
Footnotes
DISCLOSURES
There are no conflict of interest to disclose for the authors except Pandu R. Gangula (the University of Texas Medical Branch, Galveston, TX has filed a patent application in his name). Pandu R Gangula is involved in the design of experiment, and the interpretation of results as well as writing the manuscript. Kalpana Ravella, Vijayakumar Chinnathambi and Sutapa Mukhopadhyay were involved in conducting some of the experiments as well as helping in writing the manuscript. Ashley Hale and Keith Channon were involved in conducting biopterin analysis using HPLC system.
References
- 1.Schreyer SA, Vick C, Lystig TC, Mystkowski P, Leboeuf REC. LDL receptor but not apolipoprotein E deficiency increases diet-induced obesity and diabetes in mice. Am J Physiol Endocrinol Metab. 2002;282:E207–E214. doi: 10.1152/ajpendo.2002.282.1.E207. [DOI] [PubMed] [Google Scholar]
- 2.Brown MS, Goldstein JL. A receptor-mediated pathway for cholesterol homeostasig. Science (Wash DC) 1986;232:34–47. doi: 10.1126/science.3513311. [DOI] [PubMed] [Google Scholar]
- 3.Ishibashi S, Brown MS, Goldstein JL, Gerard RD, Hammer RE, Herz J. Hypercholesterolemia in low density lipoprotein receptor knockout mice and its reversal by adenovirus-mediated gene delivery. J Clin Invest. 1993;92:883–893. doi: 10.1172/JCI116663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Knowles JW, Maeda N. Genetic modifiers of atherosclerosis in mice. Arterioscler Thromb Vasc Biol. 2000;20:2336–2345. doi: 10.1161/01.atv.20.11.2336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Xiea W, Xingb D, Zhaob Y, Sub H, Mengb Z, Chenb Y, Dub LT. A new tactic to treat postprandial hyperlipidemia in diabetic rats with gastroparesis by improving gastrointestinal transit. European J Pharmacol. 2005;510:113–120. doi: 10.1016/j.ejphar.2005.01.019. [DOI] [PubMed] [Google Scholar]
- 6.Soykan I, Sivri B, Sarosiek I, Kiernan B, McCallum RW. Demography, clinical characteristics, psychological and abuse profiles, treatment, and long-term follow-up of patients with gastroparesis. Dig Dis Sci. 1998;43:2398–2404. doi: 10.1023/a:1026665728213. [DOI] [PubMed] [Google Scholar]
- 7.Chandrasekharan B, Srinivasan S. Diabetes and the enteric nervous system. Neurogastroenterol Motil. 2007;19:951–60. doi: 10.1111/j.1365-2982.2007.01023.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Huang PL, Dawson TM, Bredt DS, Snyder SH, Fishman MC. Targeted disruption of the neuronal nitric oxide synthase gene. Cell. 1993;75:1273–1286. doi: 10.1016/0092-8674(93)90615-w. [DOI] [PubMed] [Google Scholar]
- 9.Mashimo H, Kjellin A, Goyal RK. Gastric stasis in neuronal nitric oxide synthase-deficient knockout mice. Gastroenterology. 2000;119:766–773. doi: 10.1053/gast.2000.16509. [DOI] [PubMed] [Google Scholar]
- 10.Klatt P, Pfeiffer S, List BM, Lehner D, Glatter O, Bachinger HP, Werner ER, Schmidt K, Mayer B. Characterization of heme-deficient neuronal nitric-oxide synthase reveals a role for heme in subunit dimerization and binding of the amino acid substrate and tetrahydrobiopterin. J Biol Chem. 1996;271:7336–7342. doi: 10.1074/jbc.271.13.7336. [DOI] [PubMed] [Google Scholar]
- 11.Crane BR, Arvai AS, Ghosh DK, Wu C, Getzoff ED, Stuehr DJ, Tainer JA. Structure of nitric oxide synthase oxygenase dimer with pterin and substrate. Science. 1998;279:2121–2126. doi: 10.1126/science.279.5359.2121. [DOI] [PubMed] [Google Scholar]
- 12.Vasquez-Vivar J, Hogg N, Martasek P, Karoui H, Pritchard KA, Jr, Kalyanaraman B. Tetrahydrobiopterin-dependent inhibition of superoxide generation from neuronal nitric oxide synthase. J Biol Chem. 1999;274:26736–26742. doi: 10.1074/jbc.274.38.26736. [DOI] [PubMed] [Google Scholar]
- 13.Gangula PR, Maner WL, Micci MA, Garfield RE, Pasricha PJ. Diabetes induces sex-dependent changes in neuronal nitric oxide synthase dimerization and function in the rat gastric antrum. Am J Physiol Gastrointest Liver Physiol. 2007;292:G725–G733. doi: 10.1152/ajpgi.00406.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gangula PR, Mukhopadhyay S, Ravella K, Cai S, Channon KM, Garfield RE, Pasricha PJ. Tetrahydrobiopterin (BH4), a cofactor for nNOS, restores gastric emptying and nNOS expression in female diabetic rats. Am J Physiol Gastrointest Liver Physiol. 2010;298:G692–G699. doi: 10.1152/ajpgi.00450.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lee JM, Li J, Johnson DA, Stein TD, Kraft AD, Calkins MJ, Jakel RJ, Johnson JA. Nrf2, a multi-organ protector? FASEB J. 2005;19:1061–1066. doi: 10.1096/fj.04-2591hyp. [DOI] [PubMed] [Google Scholar]
- 16.Aleksunes LM, Manautou JE. Emerging Role of Nrf2 in Protecting Against Hepatic and Gastrointestinal Disease. Toxicologic Pathology. 2007;35:459–473. doi: 10.1080/01926230701311344. [DOI] [PubMed] [Google Scholar]
- 17.Kalthoff S, Ehmer U, Freiberg N, Manns MP, Strassburg CP. Interaction between oxidative stress sensor Nrf2 and xenobiotic-activated aryl hydrocarbon receptor in the regulation of the human phase II detoxifying UDP-glucuronosyltransferase 1A10. J Biol Chem. 2010;285:5993–6002. doi: 10.1074/jbc.M109.075770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shibuya A, Onda K, Kawahara H, Uchiyama Y, Nakayama H, Omi T, Nagaoka M, Matsui H, Hirano T. Sofalcone, a gastric mucosa protective agent, increases vascular endothelial growth factor via the Nrf2-heme-oxygenase-1 dependent pathway in gastric epithelial cells. Biochem Biophys Res Commun. 2010;398:581–4. doi: 10.1016/j.bbrc.2010.06.124. [DOI] [PubMed] [Google Scholar]
- 19.Iwasaki T, Takahashi S, Takahashi M, Zenimaru Y, Kujiraoka T, Ishihara M, Nagano M, Suzuki J, Miyamori I, Naiki H, Sakai J, Fujino T, Miller NE, Yamamoto TY, Hattori H. Deficiency of the Very Low-Density Lipoprotein (VLDL) Receptors in Streptozotocin-Induced Diabetic Rats: Insulin Dependency of the VLDL Receptor. Endocrinology. 2005;146:3286–3294. doi: 10.1210/en.2005-0043. [DOI] [PubMed] [Google Scholar]
- 20.Keren P, George J, Shaish A, Levkovitz H, Janakovic Z, Afek A, Goldberg I, Kopolovic J, Keren G, Harats D. Effect of hyperglycemia and hyperlipidemia on atherosclerosis in LDL receptor-deficient mice: establishment of a combined model and association with heat shock protein 65 immunity. Diabetes. 2000;49:1064–1069. doi: 10.2337/diabetes.49.6.1064. [DOI] [PubMed] [Google Scholar]
- 21.Vikramadithyan RK, Hu Y, Noh HL, Liang CP, Hallam K, Tall AR, Ramasamy R, Goldberg IJ. Human aldose reductase expression accelerates diabetic atherosclerosis in transgenic mice. J Clin Invest. 2005;115:2434–2443. doi: 10.1172/JCI24819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pannirselvam M, Simon V, Verma S, Anderson T, Triggle CR. Chronic oral supplementation with sepiapterin prevents endothelial dysfunction and oxidative stress in small mesenteric arteries from diabetic (db/db) mice. Br J Pharmacol. 2003;140:701–706. doi: 10.1038/sj.bjp.0705476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gangula PR, Mukhopadhyay S, Pasricha PJ, Ravella K. Sepiapterin reverse the changes in gastric nNOS dimerization and function in diabetic gastroparesis. Neurogastroenterol Motil. 2010;22:1325–31. doi: 10.1111/j.1365-2982.2010.01588.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li ZS, Schmauss C, Cuenca A, Ratcliffe E, Gershon MD. Physiological modulation of intestinal motility by enteric dopaminergic neurons and the D2 receptor: analysis of dopamine receptor expression, location, development, and function in wild-type and knock-out mice. J Neurosci. 2006;26:2798–2807. doi: 10.1523/JNEUROSCI.4720-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Anitha M, Chandrasekharan B, Salgado JR, Grouzmann E, Mwangi S, Sitaraman SV, Srinivasan S. Glial-derived neurotrophic factor modulates enteric neuronal survival and proliferation through neuropeptide Y. Gastroenterology. 2006a;131:1164–1178. doi: 10.1053/j.gastro.2006.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Anitha M, Gondha C, Sutliff R, Parsadanian A, Mwangi S, Sitaraman SV, Srinivasan S. GDNF rescues hyperglycemia-induced diabetic enteric neuropathy through activation of the PI3K/Akt pathway. J Clin Invest. 2006b;116:344–356. doi: 10.1172/JCI26295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Crabtree MJ, Tatham AL, Al-Wakeel Y, Warrick N, Hale AB, Cai S, Channon KM, Alp NJ. Quantitative regulation of intracellular endothelial nitric-oxide synthase (eNOS) coupling by both tetrahydrobiopterin-eNOS stoichiometry and biopterin redox status: insights from cells with tet-regulated GTP cyclohydrolase I expression. J Biol Chem. 2009;284:1136–1144. doi: 10.1074/jbc.M805403200. [DOI] [PubMed] [Google Scholar]
- 28.Pacher P, Beckman JS, Liaudet L. Nitric o xide and peroxynitrite in health and disease. Physiol Rev. 2007;87:315–424. doi: 10.1152/physrev.00029.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tayeh MA, Marletta MA. Macrophage oxidation of l-arginine to nitric oxide, nitrite, and nitrate: tetrahydrobiopterin is required as a cofactor. J Biol Chem. 1989;264:19654–19658. [PubMed] [Google Scholar]
- 30.Pall ML. Nitric oxide synthase partial uncoupling as a key switching mechanism for the NO/ONOO- cycle. Med Hypotheses. 2007;69:821–825. doi: 10.1016/j.mehy.2007.01.070. [DOI] [PubMed] [Google Scholar]
- 31.Takeda M, Yamashita T, Shinohara M, Sasaki N, Takaya T, Nakajima K, Inoue N, Masano T, Tawa H, Satomi-Kobayashi S, Toh R, Sugiyama D, Nishimura K, Yokoyama M, Hirata K, Kawashima S. Plasma tetrahydrobiopterin/dihydrobiopterin ratio: a possible marker of endothelial dysfunction. Circ J. 2009;73:955–62. doi: 10.1253/circj.cj-08-0850. [DOI] [PubMed] [Google Scholar]
- 32.Crabtree MJ, Tatham AL, Hale AB, Alp NJ, Channon KM. Critical role for tetrahydrobiopterin recycling by dihydrofolate reductase in regulation of endothelial nitric-oxide synthase coupling: relative importance of the de novo biopterin synthesis versus salvage pathways. J Biol Chem. 2009;284:28128–36. doi: 10.1074/jbc.M109.041483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Tanaka Y, Aleksunes LM, Yeager RL, Gyamfi MA, Esterly N, Guo GL, Klaassen CD. NF-E2-Related Factor 2 Inhibits Lipid Accumulation and Oxidative Stress in Mice Fed a High-Fat Diet. J Pharmacol Exp Ther. 2008;325:655–664. doi: 10.1124/jpet.107.135822. [DOI] [PubMed] [Google Scholar]
- 34.Choi KM, Gibbons SJ, Nguyen TV, Stoltz GJ, Lurken MS, Ordog T, Szurszewski JH, Farrugia G. Heme oxygenase-1 protects interstitial cells of Cajal from oxidative stress and reverses diabetic gastroparesis. Gastroenterology. 2008;135:2055–2064. doi: 10.1053/j.gastro.2008.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Choi KM, Kashyap PC, Dutta N, Stoltz GJ, Ordog T, Shea Donohue T, Bauer AJ, Linden DR, Szurszewski JH, Gibbons SJ, Farrugia G. CD206-positive M2 macrophages that express heme oxygenase-1 protect against diabetic gastroparesis in mice. Gastroenterology. 2010;138:2399–2409. doi: 10.1053/j.gastro.2010.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Parkman HP, Yates K, Hasler WL, Nguyen L, Pasricha PJ, Snape WJ, Farrugia G, Koch KL, Abell TL, McCallum RW, Lee L, Unalp-Arida A, Tonascia J, Hamilton F National Institute of Diabetes and Digestive and Kidney Diseases Gastroparesis Clinical Research Consortium. Clinical features of idiopathic gastroparesis vary with sex, body mass, symptom onset, delay in gastroparesis, and gastroparesis severity. Gastroenterol. 2011;140:101–115. doi: 10.1053/j.gastro.2010.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Farrugia G. Interstitial cells of Cajal in health and disease. Neurogastroenterol Motil. 2008;20:54–63. doi: 10.1111/j.1365-2982.2008.01109.x. [DOI] [PubMed] [Google Scholar]
- 38.Kashyap P, Farrugia G. Oxidative stress: key player in gastrointestinal complications of diabetes. Neurogastroenterol Motil. 2011;23:111–114. doi: 10.1111/j.1365-2982.2010.01659.x. [DOI] [PMC free article] [PubMed] [Google Scholar]