

Abstract
In the mid-1950s, Levine and his colleagues reported that brief intermittent exposure to early life stress diminished indications of subsequent emotionality in rats. Here we review ongoing studies of a similar process in squirrel monkeys. Results from these animal models suggest that brief intermittent exposure to stress promotes the development of arousal regulation and resilience. Implications for programs designed to enhance resilience in human development are discussed.
Keywords: emotion regulation, cognitive control, prefrontal plasticity, HPA axis, curiosity, anxiety, cortisol, squirrel monkey
INTRODUCTION
The negative consequences of early life stress are well recognized in contemporary perspectives on human mental health. Chronic exposure to severe forms of stress has been linked to the development of mood, anger, anxiety, and substance abuse disorders (Anda et al., 2006; Heim, Newport, Mletzko, Miller, & Nemeroff, 2008; Repetti, Taylor, & Seeman, 2002; Turner & Lloyd, 2003). Far less researched, but of equal importance, are indications that early intermittent exposure to stress does not result in vulnerability but instead enhances arousal regulation and resilience. Variously described in studies of humans as inoculating, steeling, or toughening (Dienstbier, 1989; Masten, 2001; Mortimer & Staff, 2004; Rutter, 2006), the notion that intermittent exposure to early life stress induces resilience is rooted in observations that Levine and his colleagues first described in the mid-1950s.
Based on Freud’s theory that early life stress contributes to the development of subsequent emotional instability, Levine set out to investigate this hypothesis as described elsewhere in detail (Levine, 1962; Levine, Chevalier, & Korchin, 1956). Levine and his colleagues expected to find that infant rats exposed to intermittent foot shocks sufficient to elicit evasive movements would show signs of subsequent emotional instability in adolescence and adulthood. Results of their experiments were not supportive of Freudian theory and Levine concluded that early intermittent stress “results in the capacity for the organism to respond more effectively when confronted with novel situations or, in other words, to exhibit a diminished emotional response” (Levine, 1962).
Thirty years later, in the Laboratory of Developmental Psychobiology that Levine founded at Stanford University, we began to investigate the long-term effects of brief intermittent mother–infant separations in squirrel monkeys. Squirrel monkeys live in groups comprised of males and females of all ages. In natural and semi-natural conditions, mothers and other group members intermittently leave newly weaned offspring to forage for food beginning at 3–6 months of age (Boinski & Fragaszy, 1989; Lyons, Kim, Schatzberg, & Levine, 1998). Initially, brief intermittent separations studied in controlled experimental conditions (see below) elicit distress peep-calls and increase plasma levels of cortisol with partial habituation of these measures of arousal observed over repeated intermittent social separations (Coe, Glass, Wiener, & Levine, 1983; Hennessy, 1986). Working on the same misapprehension that beset earlier investigations of postnatal stress in rats, we expected to find that intermittent separations would increase emotional instability. Instead, we found that monkeys exposed to early intermittent separations showed fewer behavioral indications of anxiety, diminished stress-levels of cortisol, and increased sensitivity to glucocorticoid feedback regulation of the hypothalamic–pituitary–adrenal (HPA) axis compared to age-matched monkeys not exposed to intermittent separations (Levine & Mody, 2003; Lyons, Martel, Levine, Risch, & Schatzberg, 1999; Lyons, Yang, Mobley, Nickerson, & Schatzberg, 2000).
Based on these findings, follow-up studies were designed to prospectively test the hypothesis that intermittent separations represent a form of stress inoculation that enhances arousal regulation and resilience. Socially housed squirrel monkeys were randomized to either brief intermittent separations or a nonseparated control condition at 17 weeks of age. For each of 10 total separation sessions, 1 monkey was removed from the natal group for a 1-hr session once a week. In the nonseparated control condition, age-matched monkeys remained undisturbed in their natal groups. After completion of the postnatal protocols at 27 weeks of age, all of the monkeys were maintained in identical conditions. Behavioral and hormonal measures of anxiety were assessed in the presence of each monkey’s mother at 9 months of age (Parker, Buckmaster, Schatzberg, & Lyons, 2004). Cognitive control of behavior was examined at 1.5 years of age (Parker, Buckmaster, Justus, Schatzberg, & Lyons, 2005) and exploratory behavior was assessed at 2.5 years of age (Parker et al., 2007). Noninvasive neuroimaging was performed to investigate prefrontal brain development at 3.3 years of age (Katz et al., 2009). Puberty occurs at 2–3 years of age and the average maximum life span is ~21 years. All procedures were conducted in accordance with the NIH Guide and were IACUC approved.
AROUSAL REGULATION, COGNITIVE CONTROL, CURIOSITY, AND RESILIENCE
At 9 months of age, each monkey was tested along with its mother for 30 min on five consecutive days in a novel environment that contained an assortment of foods and various toy-like objects (Parker et al., 2004). Similar test conditions have been used to demonstrate that severe forms of early life stress increase subsequent indications of anxiety in marmoset monkeys and macaques (Andrews & Rosenblum, 1993; Dettling, Feldon, & Pryce, 2002; Hinde & Spencer-Booth, 1971). Here the novel environment test was used determine whether intermittent separations enhance the development of arousal regulation and resilience.
In the novel environment stress test, the previously separated and nonseparated monkeys were initially similar but differences emerged over repeated test sessions. Gradually, the previously separated monkeys showed fewer signs of anxiety inferred from decreased maternal clinging and increased exploration compared to monkeys not exposed to intermittent separations. Plasma levels of pituitary–adrenal stress hormones were likewise diminished in monkeys previously exposed to intermittent separations compared to nonseparated controls (Fig. 1). As described elsewhere in greater detail (Parker et al., 2004), these results support the suggestion that intermittent separations represent a form of stress inoculation that enhances arousal regulation and resilience in novel situations.
FIGURE 1.
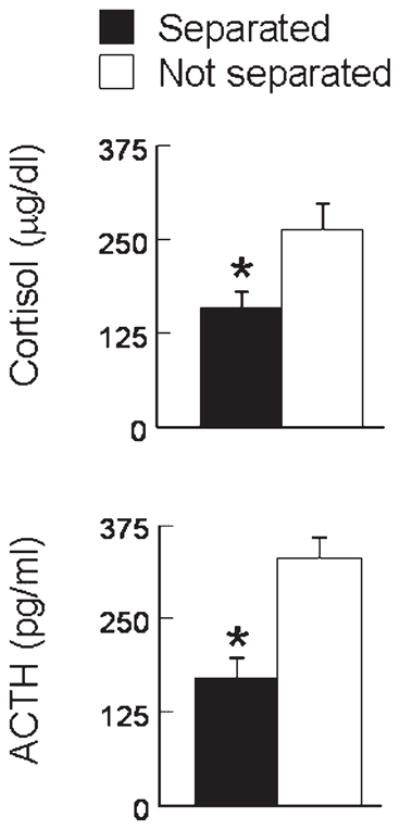
Early intermittent separations diminish subsequent pituitary–adrenal responses to a novel test environment. Posttest plasma levels of cortisol (top panel) and adrenocortico-tropic hormone (bottom panel) are presented for previously separated and nonseparated monkeys (n = 9–10 monkeys per condition; mean ± SEM). Asterisks signify a significant (p <.05) separation effect (adapted from data presented in Parker et al., 2004).
At 1.5 years of age, all monkeys were administered a behavioral test of cognitive control that specifically focused on response inhibition (Parker et al., 2005). Monkeys were required to inhibit a previously rewarded reaching response in order to efficiently retrieve food treats from a clear plastic box. In marmoset monkeys and macaques, performance on this test is impaired by lesions of prefrontal cortex (Diamond, 1990; Dias, Robbins, & Roberts, 1996; Wallis, Dias, Robbins, & Roberts, 2001) but not lesions of the hippocampus (Diamond, Zola-Morgan, & Squire, 1989). Test performance is likewise impaired in a marmoset model of parental neglect (Pryce, Dettling, Spengler, Spaete, & Feldon, 2004) and in squirrel monkeys treated with cortisol according to a protocol that simulates a chronic neuroendocrine stress response (Lyons, Lopez, Yang, & Schatzberg, 2000). Here this test was used to assess whether intermittent separations affect cognitive control of behavior.
The ability to inhibit a previously rewarded behavioral response gradually improved over repeated test trials. At peak levels of performance, however, all of the previously separated monkeys successfully completed all test trials whereas fewer than half of the monkeys not exposed to intermittent separations achieved similar levels of success (Parker et al., 2005). These and related findings suggest that early intermittent separations enhance cognitive control of behavior.
At 2.5 years of age, all monkeys were administered a test to examine curiosity in a stress-free condition (Parker et al., 2007). Each monkey was provided free access to explore a familiar or a novel toy-like object secured to the wall of a compartment affixed to the monkey’s home cage. Similar test conditions have been used to demonstrate that severe forms of early life stress diminish subsequent exploratory tendencies in macaques (Hinde & Spencer-Booth, 1971; Roder, Timmermans, & Vossen, 1989; Sackett, 1972). Here exploration of novelty was used to determine whether intermittent separations induce subsequent differences in curiosity and exploratory behavior.
All previously separated and all but one of the nonseparated monkeys approached the compartment and subsequently peered inside. More of the previously separated monkeys left the home cage to enter the compartment, and they entered faster, with greater frequency, and spent more time inside the compartment compared to nonseparated monkeys (Parker et al., 2007). Inside the compartment, more previously separated than nonseparated monkeys explored both objects, and the previously separated monkeys showed a significant preference for the novel object compared to the familiar object (Fig. 2).
FIGURE 2.
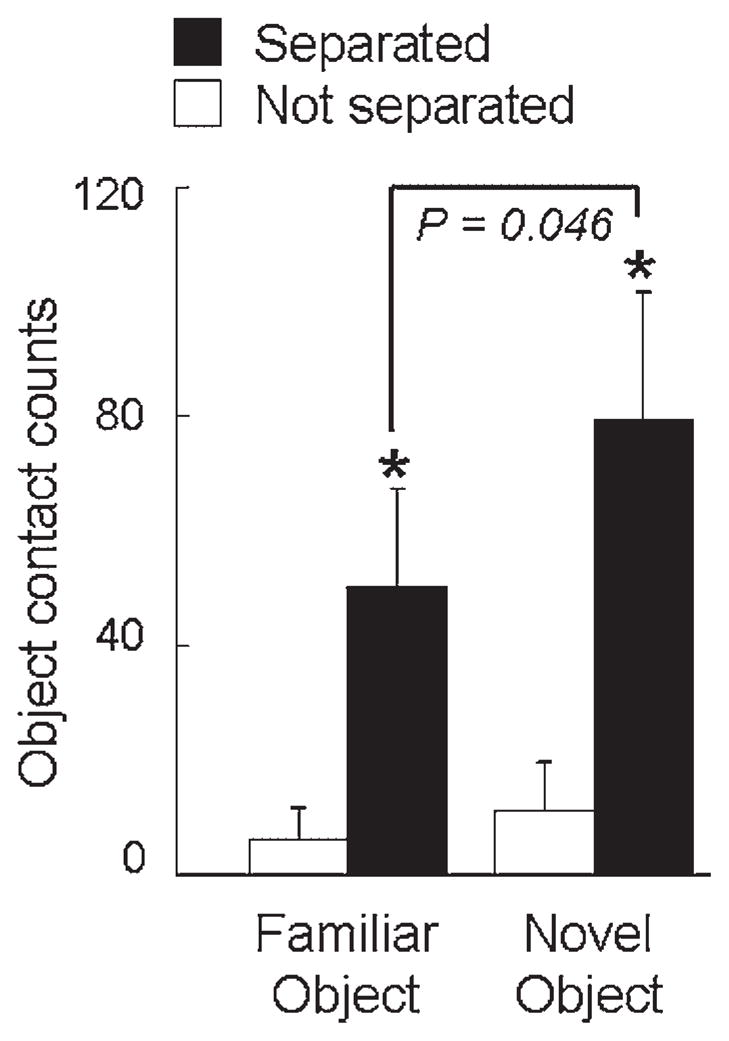
Early intermittent separations increase subsequent novelty-seeking behavior. Measures of familiar versus novel object exploration are presented for previously separated and nonseparated monkeys (n = 9–10 monkeys per condition; mean ± SEM). Asterisks signify separation effects (p <.05) and the bracket shows that previously separated monkeys preferred to interact with the novel toy-like object (adapted from data presented in Parker et al., 2007).
In the earlier novel environment test conducted at 9 months of age, significantly more object exploration was also observed in the previously separated compared to nonseparated monkeys (Parker et al., 2004). Object exploration in the earlier study was assessed, however, in stressful conditions that involved involuntary removal of monkeys from the home cage. Both the previously separated and nonseparated monkeys in the earlier study had higher plasma levels of pituitary–adrenal hormones after the novel environment test compared to baseline levels measured in undisturbed home cage conditions (Parker et al., 2004). Moreover, posttest plasma levels of pituitary–adrenal hormones were significantly higher in the nonseparated compared to separated monkeys (Fig. 1). Differences in pituitary–adrenal activity were therefore confounded with differences in exploratory behavior assessed at 9 months of age.
In the study conducted at 2.5 years of age, exploratory behavior was examined by providing free access to a compartment attached to each monkey’s home cage (Parker et al., 2007). To further reduce the potentially stressful nature of the test situation, each monkey was acclimated to all test procedures prior to the start of the study. Posttest cortisol levels at 2.5 years of age were not significantly different from baseline levels measured in undisturbed home cage conditions. Moreover, neither baseline nor posttest cortisol levels at 2.5 years of age correlated with exploratory behavior in the stress-free test condition. Individual differences previously observed in object exploration at 9 months of age were, however, correlated with differences in test compartment entries and total time spent inside the compartment at 2.5 years of age (Lyons & Parker, 2007). Individual differences previously observed in cognitive control at 1.5 years of age were also correlated with test compartment entries and total time spent inside the compartment at 2.5 years of age. Taken together, these findings suggest that early intermittent separations induce broad and enduring trait-like transformation in cognitive, motivational, and emotional aspects of arousal regulation and resilience (Lyons, Parker, Katz, & Schatzberg, 2009).
MATERNAL MEDIATION
Stress inoculation-induced resilience in monkeys resembles the effects of early intermittent “postnatal handling” that Levine and his colleagues identified in studies of rats. Neonatal rats removed from the nest and briefly separated from nest-related cues respond with behavioral and physiological indications of arousal (Hofer, 1996; Lyons & Banks, 1982). When studied as adults, these rats then display diminished emotionality, increased exploration, improved learning, and diminished HPA axis activation compared to rats raised in standard undisturbed laboratory conditions (Fernandez-Teruel et al., 2002; Levine, 1957, 2000; Levine, Alpert, & Lewis, 1957; Meaney et al., 1996).
In rats, these outcomes appear to partially reflect the effects of maternal behavior directed toward pups that are briefly separated and then returned to the nest (Denenberg, 1999; Smotherman & Bell, 1980). Brief intermittent separations increase arched-back nursing after reunions (Pryce, Bettschen, & Feldon, 2001) and in some studies high levels of maternal care persist throughout early development (Lee & Williams, 1974). Importantly, the neuroendocrine indications of resilience observed in rats exposed to intermittent separations can be replicated in nonmanipulated offspring that naturally receive high levels of maternal care (Liu et al., 1997). That increased maternal care is sufficient to induce resilience in rats is corroborated by evidence that the amount of maternal stimulation received by pups during development is negatively correlated with later stress-induced activation of the HPA axis (Liu et al., 1997; Rosenberg, Denenberg, & Zarrow, 1970). Maternal mediation might therefore explain Levine’s observation that pups removed intermittently from the nest and exposed to brief foot shock show subsequent signs of diminished emotionality as adults (Levine, 1962; Levine et al., 1956).
To determine whether resilience in monkeys exposed to early intermittent separations is maternally mediated, we examined maternal behavior and subsequent neuroendocrine measures of resilience in monkeys that were randomized to three postnatal conditions (Parker, Buckmaster, Sundlass, Schatzberg, & Lyons, 2006). In one condition, each monkey was separated from its mother and the natal group for 10 weekly 1-hr sessions. In the second condition, each monkey and its mother were removed together as a pair and separated from the natal group for 10 weekly 1-hr sessions. Both of these social separation conditions elicit locomotor agitation, distress vocalizations, and HPA axis activation with baseline measures of arousal restored soon after reunion with members of the natal group (Coe et al., 1983; Jordan, Hennessy, Gonzalez, & Levine, 1985). In the third treatment condition, nonseparated monkeys remained undisturbed in their natal groups.
Neither separation condition induced long-lasting changes in maternal care (Parker et al., 2006). Moreover, the transient changes observed in maternal behavior did not correspond with differences in the development of arousal regulation and resilience. Monkeys exposed along with their mother to intermittent separations received less maternal care in the home cage as mothers direct their attention toward re-integration with the group. Yet both intermittent separation conditions induced neuroendocrine indications of resilience compared to control monkeys not exposed to intermittent separations (Fig. 3). Specifically, monkeys separated alone or separated along with their mother exhibited diminished cortisol responses to a subsequent novel environment stress test compared to monkeys not exposed to either separation condition (Parker et al., 2006). These and related findings suggest that arousal regulation and resilience correspond more closely to prior stress exposure than to separation-induced changes in maternal care. Similar conclusions have been reported in recent investigations of developmental plasticity and early life stress in rats (Macri & Wurbel, 2006; Tang, Akers, Reeb, Romeo, & McEwen, 2006).
FIGURE 3.

Early intermittent separation-induced changes in neuroendocrine indications of resilience are not maternally mediated. The percentage of contact attempts initiated by mothers in the home cage (top panel) and plasma cortisol levels after exposure to a novel environment stress test (bottom panel) are presented for 9–11 monkeys in each treatment condition (mean ± SEM). Treatment conditions without shared superscript letters differ significantly from one another (p <.05) and conditions with shared letters do not (adapted from data presented in Parker et al., 2006).
PREFRONTAL NEUROPLASTICITY
Based on our findings that early intermittent separations enhance arousal regulation and improve prefrontal-dependent cognitive control of behavior, noninvasive neuroimaging was used to examine separation effects on prefrontal brain development. Monkeys randomized to brief intermittent separations or a nonseparated condition were initially studied as described above and then neuroimaging was performed at 3.3 years of age (Katz et al., 2009). In keeping with previous evidence (Lyons, Afarian, Schatzberg, Sawyer-Glover, & Moseley, 2002) we found that monkeys exposed to early intermittent separations had larger ventromedial but not larger dorsolateral prefrontal cortical volumes compared to monkeys not exposed to intermittent separations (Fig. 4). Larger volumes do not reflect increased cortical thickness but instead represent surface area expansion of ventromedial prefrontal cortex (Katz et al., 2009).
FIGURE 4.
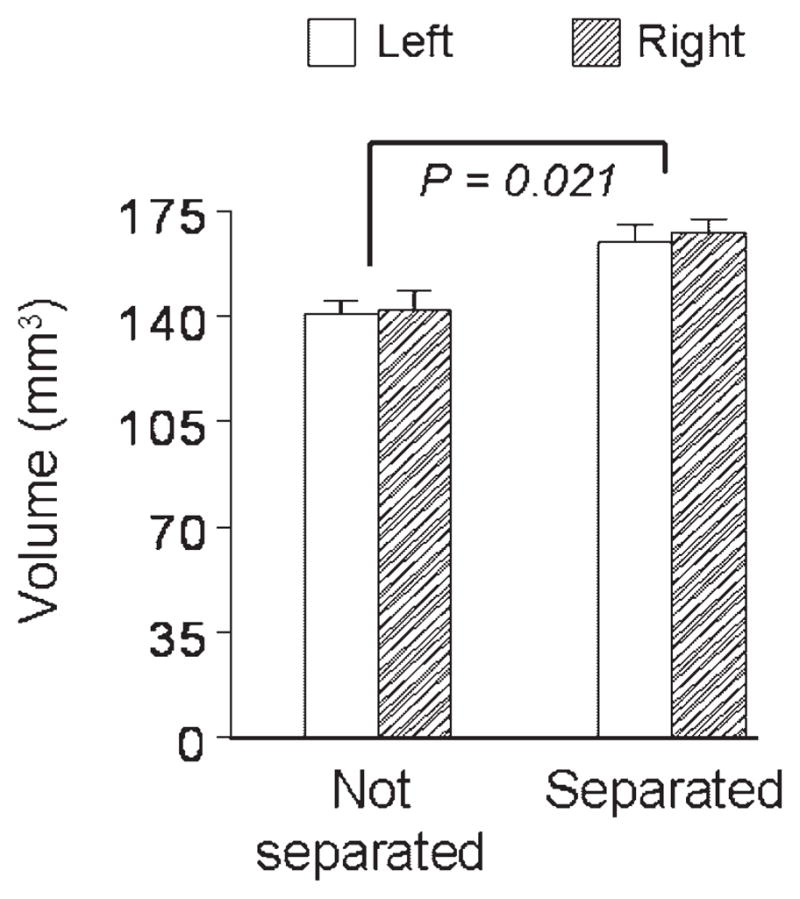
Early intermittent separation-induced adaptations in prefrontal cortex. Ventromedial prefrontal cortical volumes are larger in monkeys exposed to early intermittent separations compared to nonseparated controls (n = 9–10 monkeys per condition; mean ± SEM). Separation effects were not discerned in dorsolateral prefrontal cortex (data not shown) and the separation-by-brain side interactions were not statistically significant (adapted from data presented in Katz et al., 2009).
These findings are of interest because increased ventromedial prefrontal size in humans predicts diminished impulsivity (Matsuo et al., 2009), lower harm avoidance (Yamasue et al., 2008), and greater retention of learned extinction of fear (Milad et al., 2005). Recent neuroimaging studies of humans support results from animal research confirming that learned extinction of fear is mediated by prefrontal downregulation of arousal via inhibitory connections that diminish neural output from the amygdala (Delgado, Nearing, Ledoux, & Phelps, 2008; Quirk & Mueller, 2008). Additional evidence likewise suggests that differences in the balance between top-down prefrontal regulation and arousal inducing amygdala activation may account for trait-like differences in coping with intermittent stress (Bishop, 2007; Drabant, McRae, Manuck, Hariri, & Gross, 2009).
Despite evidence that coping with stress depends on myelinated prefrontal cortical and subcortical (e.g., amygdala) interconnections, myelination is seldom considered in discussions of neural plasticity as a mechanism for experience-dependent resilience. In the study described above, magnetic resonance diffusion tensor imaging was used to examine prefrontal myelination in terms of fractional anisotropy (Katz et al., 2009). Fractional anisotropy represents water proton diffusion directionality on a scale from zero to one. A value of zero indicates that diffusion is unrestricted in all directions, whereas a value of one indicates that diffusion is completely restricted along a single direction. High values of fractional anisotropy occur when tissue microstructure restricts diffusion directionality as exemplified by myelination of axons in white matter. Myelination increases nerve conduction velocities and facilitates synchronous firing of neurons by reducing travel distance effects in distributed networks (Fields, 2008). Coordination of firing inputs to maximize temporal summation at postsynaptic neurons is the foundation for a key concept in neuroplasticity and development—neurons that fire together, wire together. In children, myelination of prefrontal connections determined by fractional anisotropy increases with age (Barnea-Goraly et al., 2005) and maturation of prefrontal-dependent functions (Casey et al., 2007).
In monkeys, exposure to early intermittent separations increases ventromedial but not dorsolateral prefrontal white matter measures of fractional anisotropy (Fig. 5). This difference coincides with the difference in ventromedial and not dorsolateral prefrontal cortical volumes described above. White matter measures of fractional anisotropy in ventromedial but not dorsolateral regions correlate with their respective volumes of prefrontal cortex (Katz et al., 2009). These findings suggest that the learning-like process of coping with early intermittent stress increases prefrontal myelination and expands a region of cortex that broadly controls arousal regulation and resilience.
FIGURE 5.
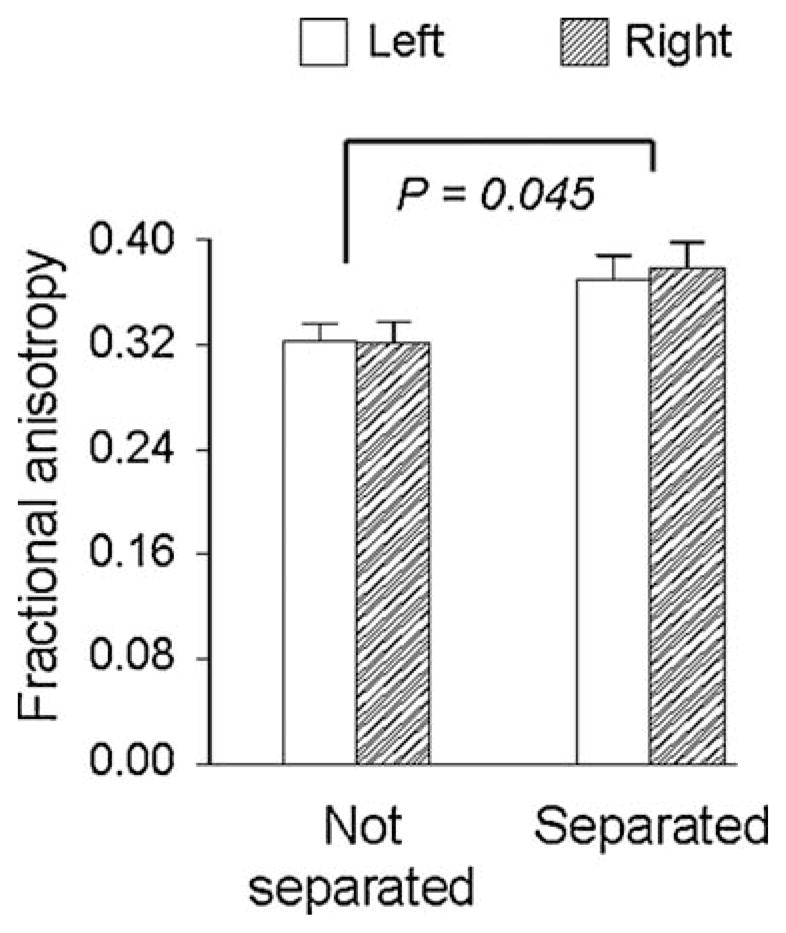
Early intermittent separation-induced adaptations in prefrontal white matter. Ventromedial prefrontal white matter fractional anisotropy is greater in monkeys previously exposed to early intermittent separations compared to nonseparated controls (n = 9–10 monkeys per condition; mean ± SEM). Separation effects were not discerned in the white matter region underlying dorsolateral prefrontal cortex (data not shown) and the separation-by-brain side interactions were not statistically significant (adapted from data presented in Katz et al., 2009).
The effect size for these neuroadaptations is similar to that observed in studies of environmental enrichment in marmoset monkeys (Kozorovitskiy et al., 2005) and rats (Rosenzweig, 2003). Enrichment entails repeated exposure to novel inanimate and social stimulation (Fox, Merali, & Harrison, 2006) that elicits neuroendocrine indications of stress coping in rats (Moncek, Duncko, Johansson, & Jezova, 2004). After exposure to enrichment, however, rats show diminished indications of anxiety, increased exploration, and enhanced prefrontal-dependent cognitive control compared to nonenriched rats (Fox et al., 2006; Schrijver, Pallier, Brown, & Wurbel, 2004). These findings combined with our studies suggest that enrichment effects may be mediated, in part, by the process of coping with stress.
ENHANCING HUMAN RESILIENCE
Controlled exposure to stress-related cues is a key feature of resiliency training for people that work in conditions where performance in the face of adversity is required, for example, medical and military personnel, aviators, police, firefighters, and rescue workers (Meichenbaum, 2007; Stetz et al., 2007). A similar process also occurs during cognitive behavior exposure therapies for stress-induced psychopathologies. Intermittent exposure to relevant stressors is thought to induce a learning-like process that modifies erroneous psychobiological conditions that impair mental health (De Raedt, 2006; Foa & Kozak, 1986).
Although resiliency training and exposure psychotherapies are generally administered by psychologists and psychiatrists, these interventions build on a process that appears to spontaneously occur in most humans without professional training or formal guidance (Epstein, 1983). Adult humans, for example, tend to cope better with spousal loss, illness, and accidents if they have previously experienced and coped with stress in childhood (Khoshaba & Maddi, 1999). Work-related stress likewise has fewer deleterious mental health effects in adults previously exposed to work stress in adolescence (Mortimer & Staff, 2004). These and related investigations of infants (Tronick, 2006) and young children (Ellis, Essex, & Boyce, 2005; Gunnar, Frenn, Wewerka, & Van Ryzin, 2008) suggest that a spontaneous learning-like stress coping process promotes arousal regulation and resilience.
The public health impact of understanding how spontaneous stress coping promotes resilience is potentially quite high. According to the World Health Organization, stress-related health disorders will be the second leading cause of disabilities by the year 2020. The American Institute of Stress has determined that 75–90% of all medical visits are stress-related and cost the nation 42 billion dollars each year. Levine’s seminal observation that early intermittent exposure to stress does not increase vulnerability, but instead promotes the development of resilience warrants renewed attention in programs designed to improve human health.
Acknowledgments
This study was supported by Public Health Service grants MH47573, MH66537, MH77884, and DA16902.
References
- Anda RF, Felitti VJ, Bremner JD, Walker JD, Whitfield C, Perry BD, et al. The enduring effects of abuse and related adverse experiences in childhood. A convergence of evidence from neurobiology and epidemiology. European Archives of Psychiatry and Clinical Neuroscience. 2006;256(3):174–186. doi: 10.1007/s00406-005-0624-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrews MW, Rosenblum LA. Assessment of attachment in differentially reared infant monkeys (Macaca radiata): Response to separation and a novel environment. Journal of Comparative Psychology. 1993;107(1):84–90. doi: 10.1037/0735-7036.107.1.84. [DOI] [PubMed] [Google Scholar]
- Barnea-Goraly N, Menon V, Eckert M, Tamm L, Bammer R, Karchemskiy A, et al. White matter development during childhood and adolescence: A cross-sectional diffusion tensor imaging study. Cerebral Cortex. 2005;15(12):1848–1854. doi: 10.1093/cercor/bhi062. [DOI] [PubMed] [Google Scholar]
- Bishop SJ. Neurocognitive mechanisms of anxiety: An integrative account. Trends in Cognitive Science. 2007;11(7):307–316. doi: 10.1016/j.tics.2007.05.008. [DOI] [PubMed] [Google Scholar]
- Boinski S, Fragaszy DM. The ontogeny of foraging in squirrel monkeys, Saimiri oerstedi. Animal Behaviour. 1989;37:415–428. [Google Scholar]
- Casey BJ, Epstein JN, Buhle J, Liston C, Davidson MC, Tonev ST, et al. Frontostriatal connectivity and its role in cognitive control in parent-child dyads with ADHD. American Journal of Psychiatry. 2007;164(11):1729–1736. doi: 10.1176/appi.ajp.2007.06101754. [DOI] [PubMed] [Google Scholar]
- Coe CL, Glass JC, Wiener SG, Levine S. Behavioral, but not physiological, adaptation to repeated separation in mother and infant primates. Psychoneuroendocrinology. 1983;8(4):401–409. doi: 10.1016/0306-4530(83)90019-7. [DOI] [PubMed] [Google Scholar]
- De Raedt R. Does neuroscience hold promise for the further development of behavior therapy? The case of emotional change after exposure in anxiety and depression. Scandinavian Journal of Psychology. 2006;47(3):225–236. doi: 10.1111/j.1467-9450.2006.00511.x. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Nearing KI, Ledoux JE, Phelps EA. Neural circuitry underlying the regulation of conditioned fear and its relation to extinction. Neuron. 2008;59(5):829–838. doi: 10.1016/j.neuron.2008.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denenberg VH. Commentary: Is maternal stimulation the mediator of the handling effect in infancy? Developmental Psychobiology. 1999;34(1):1–3. doi: 10.1002/(sici)1098-2302(199901)34:1<1::aid-dev2>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- Dettling AC, Feldon J, Pryce CR. Repeated parental deprivation in the infant common marmoset (Callithrix jacchus, primates) and analysis of its effects on early development. Biological Psychiatry. 2002;52(11):1037–1046. doi: 10.1016/s0006-3223(02)01460-9. [DOI] [PubMed] [Google Scholar]
- Diamond A. Developmental time course in human infants and infant monkeys, and the neural bases of, inhibitory control of reaching. Annals of the New York Academy of Sciences. 1990;608:637–676. doi: 10.1111/j.1749-6632.1990.tb48913.x. [DOI] [PubMed] [Google Scholar]
- Diamond A, Zola-Morgan S, Squire LR. Successful performance by monkeys with lesions of the hippocampal formation on AB and object retrieval, two tasks that mark developmental changes in human infants. Behavioral Neuroscience. 1989;103(3):526–537. doi: 10.1037//0735-7044.103.3.526. [DOI] [PubMed] [Google Scholar]
- Dias R, Robbins TW, Roberts AC. Primate analogue of the Wisconsin Card Sorting Test: Effects of excitotoxic lesions of the prefrontal cortex in the marmoset. Behavioral Neuroscience. 1996;110(5):872–886. doi: 10.1037//0735-7044.110.5.872. [DOI] [PubMed] [Google Scholar]
- Dienstbier RA. Arousal and physiological toughness: Implications for mental and physical health. Psychological Review. 1989;96(1):84–100. doi: 10.1037/0033-295x.96.1.84. [DOI] [PubMed] [Google Scholar]
- Drabant EM, McRae K, Manuck SB, Hariri AR, Gross JJ. Individual differences in typical reappraisal use predict amygdala and prefrontal responses. Biological Psychiatry. 2009;65(5):367–373. doi: 10.1016/j.biopsych.2008.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis BJ, Essex MJ, Boyce WT. Biological sensitivity to context: II. Empirical explorations of an evolutionary-developmental theory. Development and Psychopathology. 2005;17(2):303–328. doi: 10.1017/s0954579405050157. [DOI] [PubMed] [Google Scholar]
- Epstein S. Natural healing processes of the mind: Graded stress inoculation as an inherent coping mechanism. In: Meichenbaum D, Jaremko ME, editors. Stress reduction and prevention. New York: Plenum Press; 1983. pp. 39–66. [Google Scholar]
- Fernandez-Teruel A, Gimenez-Llort L, Escorihuela RM, Gil L, Aguilar R, Steimer T, et al. Early-life handling stimulation and environmental enrichment. Are some of their effects mediated by similar neural mechanisms? Pharmacology Biochemistry and Behavior. 2002;73(1):233–245. doi: 10.1016/s0091-3057(02)00787-6. [DOI] [PubMed] [Google Scholar]
- Fields RD. White matter in learning, cognition and psychiatric disorders. Trends in Neuroscience. 2008;31(7):361–370. doi: 10.1016/j.tins.2008.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foa EB, Kozak MJ. Emotional processing of fear: Exposure to corrective information. Psychological Bulletin. 1986;99(1):20–35. [PubMed] [Google Scholar]
- Fox C, Merali Z, Harrison C. Therapeutic and protective effect of environmental enrichment against psychogenic and neurogenic stress. Behavioral Brain Research. 2006;175(1):1–8. doi: 10.1016/j.bbr.2006.08.016. [DOI] [PubMed] [Google Scholar]
- Gunnar MR, Frenn K, Wewerka SS, Van Ryzin MJ. Moderate versus severe early life stress: Associations with stress reactivity and regulation in 10–12-year-old children. Psychoneuroendocrinology. 2008;34(1):62–75. doi: 10.1016/j.psyneuen.2008.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heim C, Newport DJ, Mletzko T, Miller AH, Nemeroff CB. The link between childhood trauma and depression: Insights from HPA axis studies in humans. Psychoneuroendocrinology. 2008;33(6):693–710. doi: 10.1016/j.psyneuen.2008.03.008. [DOI] [PubMed] [Google Scholar]
- Hennessy MB. Multiple, brief maternal separations in the squirrel monkey: Changes in hormonal and behavioral responsiveness. Physiology and Behavior. 1986;36(2):245–250. doi: 10.1016/0031-9384(86)90011-9. [DOI] [PubMed] [Google Scholar]
- Hinde RA, Spencer-Booth Y. Effects of brief separation from mother on rhesus monkeys. Science. 1971;173(992):111–118. doi: 10.1126/science.173.3992.111. [DOI] [PubMed] [Google Scholar]
- Hofer MA. Multiple regulators of ultrasonic vocalization in the infant rat. Psychoneuroendocrinology. 1996;21(2):203–217. doi: 10.1016/0306-4530(95)00042-9. [DOI] [PubMed] [Google Scholar]
- Jordan TC, Hennessy MB, Gonzalez CA, Levine S. Social and environmental factors influencing mother-infant separation-reunion in squirrel monkeys. Physiology and Behavior. 1985;34(4):489–493. doi: 10.1016/0031-9384(85)90038-1. [DOI] [PubMed] [Google Scholar]
- Katz M, Liu C, Schaer M, Parker KJ, Ottet M, Epps A, et al. Prefrontal plasticity and stress inoculation-induced resilience. Developmental Neuroscience. 2009;31(4):293–299. doi: 10.1159/000216540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khoshaba DM, Maddi SR. Early experiences in hardiness development. Consulting Psychology Journal: Practice and Research. 1999;51(2):106–116. [Google Scholar]
- Kozorovitskiy Y, Gross CG, Kopil C, Battaglia L, McBreen M, Stranahan AM, et al. Experience induces structural and biochemical changes in the adult primate brain. Proceeds of the National Academy of Sciences USA. 2005;102(48):17478–17482. doi: 10.1073/pnas.0508817102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee MH, Williams DI. Changes in licking behaviour of rat mother following handling of young. Animal Behaviour. 1974;22:679–681. [Google Scholar]
- Levine S. Infantile experience and resistence to physiological stress. Science. 1957;126:405–406. doi: 10.1126/science.126.3270.405. [DOI] [PubMed] [Google Scholar]
- Levine S. The effects of infantile experience on adult behavior. In: Bachrach AJ, editor. Experimental foundations of clinical psychology. New York: Basic Books; 1962. pp. 139–169. [Google Scholar]
- Levine S. Influence of psychological variables on the activity of the hypothalamic-pituitary-adrenal axis. European Journal of Pharmacology. 2000;405(1–3):149–160. doi: 10.1016/s0014-2999(00)00548-3. [DOI] [PubMed] [Google Scholar]
- Levine S, Alpert M, Lewis GW. Infantile experience and the maturation of the pituitary adrenal axis. Science. 1957;126(3287):1347. doi: 10.1126/science.126.3287.1347. [DOI] [PubMed] [Google Scholar]
- Levine S, Chevalier JA, Korchin SJ. The effects of early shock and handling on later avoidance learning. Journal of Personality. 1956;24(4):475–4493. doi: 10.1111/j.1467-6494.1956.tb01283.x. [DOI] [PubMed] [Google Scholar]
- Levine S, Mody T. The long-term psychobiological consequences of intermittent postnatal separation in the squirrel monkey. Neuroscience and Biobehavioral Reviews. 2003;27(1–2):83–89. doi: 10.1016/s0149-7634(03)00011-3. [DOI] [PubMed] [Google Scholar]
- Liu D, Diorio J, Tannenbaum B, Caldji C, Francis D, Freedman A, et al. Maternal care, hippocampal glucocorticoid receptors, and hypothalamic-pituitary-adrenal responses to stress. Science. 1997;277(5332):1659–1662. doi: 10.1126/science.277.5332.1659. [DOI] [PubMed] [Google Scholar]
- Lyons DM, Afarian H, Schatzberg AF, Sawyer-Glover A, Moseley ME. Experience-dependent asymmetric variation in primate prefrontal morphology. Behavioral Brain Research. 2002;136(1):51–59. doi: 10.1016/s0166-4328(02)00100-6. [DOI] [PubMed] [Google Scholar]
- Lyons DM, Banks EM. Ultrasounds in neonatal rats: Novel, predator and conspecific odor cues. Developmental Psychobiology. 1982;15(5):455–460. doi: 10.1002/dev.420150507. [DOI] [PubMed] [Google Scholar]
- Lyons DM, Kim S, Schatzberg AF, Levine S. Postnatal foraging demands alter adrenocortical activity and psychosocial development. Developmental Psychobiology. 1998;32(4):285–291. [PubMed] [Google Scholar]
- Lyons DM, Lopez JM, Yang C, Schatzberg AF. Stress-level cortisol treatment impairs inhibitory control of behavior in monkeys. Journal of Neuroscience. 2000;20(20):7816–7821. doi: 10.1523/JNEUROSCI.20-20-07816.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons DM, Martel FL, Levine S, Risch NJ, Schatzberg AF. Postnatal experiences and genetic effects on squirrel monkey social affinities and emotional distress. Hormones and Behavior. 1999;36(3):266–275. doi: 10.1006/hbeh.1999.1547. [DOI] [PubMed] [Google Scholar]
- Lyons DM, Parker KJ. Stress inoculation-induced indications of resilience in monkeys. Journal of Traumatic Stress. 2007;20(4):423–433. doi: 10.1002/jts.20265. [DOI] [PubMed] [Google Scholar]
- Lyons DM, Parker KJ, Katz M, Schatzberg AF. Developmental cascades linking stress inoculation, arousal regulation, and resilience. Frontiers in Behavioral Neuroscience. 2009;3:32. doi: 10.3389/neuro.08.032.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons DM, Yang C, Mobley BW, Nickerson JT, Schatzberg AF. Early environmental regulation of glucocorticoid feedback sensitivity in young adult monkeys. Journal of Neuroendocrinology. 2000;12(8):723–728. doi: 10.1046/j.1365-2826.2000.00505.x. [DOI] [PubMed] [Google Scholar]
- Macri S, Wurbel H. Developmental plasticity of HPA and fear responses in rats: A critical review of the maternal mediation hypothesis. Hormones and Behavior. 2006;50(5):667–680. doi: 10.1016/j.yhbeh.2006.06.015. [DOI] [PubMed] [Google Scholar]
- Masten AS. Ordinary magic. Resilience processes in development. American Psychologist. 2001;56(3):227–238. doi: 10.1037//0003-066x.56.3.227. [DOI] [PubMed] [Google Scholar]
- Matsuo K, Nicoletti M, Nemoto K, Hatch JP, Peluso MA, Nery FG, et al. A voxel-based morphometry study of frontal gray matter correlates of impulsivity. Human Brain Mapping. 2009;30(4):1188–1195. doi: 10.1002/hbm.20588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meaney MJ, Diorio J, Francis D, Widdowson J, LaPlante P, Caldji C, et al. Early environmental regulation of forebrain glucocorticoid receptor gene expression: Implications for adrenocortical responses to stress. Developmental Neuroscience. 1996;18(1–2):49–72. doi: 10.1159/000111395. [DOI] [PubMed] [Google Scholar]
- Meichenbaum D. Stress inoculation training: A preventative and treatment approach. In: Lehrer PM, Woolfolk RL, Sime WE, editors. Principles and practice of stress management. 3. New York, NY: Guildford Press; 2007. pp. 497–518. [Google Scholar]
- Milad MR, Quinn BT, Pitman RK, Orr SP, Fischl B, Rauch SL. Thickness of ventromedial prefrontal cortex in humans is correlated with extinction memory. Proceedings of the National Academy of Sciences USA. 2005;102(30):10706–10711. doi: 10.1073/pnas.0502441102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moncek F, Duncko R, Johansson BB, Jezova D. Effect of environmental enrichment on stress related systems in rats. Journal of Neuroendocrinology. 2004;16(5):423–431. doi: 10.1111/j.1365-2826.2004.01173.x. [DOI] [PubMed] [Google Scholar]
- Mortimer JT, Staff J. Early work as a source of developmental discontinuity during the transition to adulthood. Development and Psychopathology. 2004;16(4):1047–1070. doi: 10.1017/s0954579404040131. [DOI] [PubMed] [Google Scholar]
- Parker KJ, Buckmaster CL, Justus KR, Schatzberg AF, Lyons DM. Mild early life stress enhances prefrontal-dependent response inhibition in monkeys. Biological Psychiatry. 2005;57(8):848–855. doi: 10.1016/j.biopsych.2004.12.024. [DOI] [PubMed] [Google Scholar]
- Parker KJ, Buckmaster CL, Schatzberg AF, Lyons DM. Prospective investigation of stress inoculation in young monkeys. Archives of General Psychiatry. 2004;61(9):933–941. doi: 10.1001/archpsyc.61.9.933. [DOI] [PubMed] [Google Scholar]
- Parker KJ, Buckmaster CL, Sundlass K, Schatzberg AF, Lyons DM. Maternal mediation, stress inoculation, and the development of neuroendocrine stress resistance in primates. Proceedings of the National Academy of Sciences USA. 2006;103(8):3000–3005. doi: 10.1073/pnas.0506571103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker KJ, Rainwater KL, Buckmaster CL, Schatzberg AF, Lindley SE, Lyons DM. Early life stress and novelty seeking behavior in adolescent monkeys. Psychoneuroendocrinology. 2007;32(7):785–792. doi: 10.1016/j.psyneuen.2007.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pryce CR, Bettschen D, Feldon J. Comparison of the effects of early handling and early deprivation on maternal care in the rat. Developmental Psychobiology. 2001;38(4):239–251. doi: 10.1002/dev.1018. [DOI] [PubMed] [Google Scholar]
- Pryce CR, Dettling A, Spengler M, Spaete C, Feldon J. Evidence for altered monoamine activity and emotional and cognitive disturbance in marmoset monkeys exposed to early life stress. Annals of the New York Academy of Sciences. 2004;1032:245–249. doi: 10.1196/annals.1314.030. [DOI] [PubMed] [Google Scholar]
- Quirk GJ, Mueller D. Neural mechanisms of extinction learning and retrieval. Neuropsychopharmacology. 2008;33(1):56–72. doi: 10.1038/sj.npp.1301555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Repetti RL, Taylor SE, Seeman TE. Risky families: Family social environments and the mental and physical health of offspring. Psychological Bulletin. 2002;128(2):330–366. [PubMed] [Google Scholar]
- Roder EL, Timmermans PJ, Vossen JM. Effects of rearing and exposure condition upon the acquisition of phobic behaviour in cynomolgus monkeys. Behaviour Research and Therapy. 1989;27(3):221–231. doi: 10.1016/0005-7967(89)90040-5. [DOI] [PubMed] [Google Scholar]
- Rosenberg KM, Denenberg VH, Zarrow MX. Mice (Mus musculus) reared with rat aunts: The role of rat-mouse contact in mediating behavioural and physiological changes in the mouse. Animal Behaviour. 1970;18(1):138–143. doi: 10.1016/0003-3472(70)90082-5. [DOI] [PubMed] [Google Scholar]
- Rosenzweig MR. Effects of differential experience on the brain and behavior. Developmental Neuropsychology. 2003;24(2–3):523–540. doi: 10.1080/87565641.2003.9651909. [DOI] [PubMed] [Google Scholar]
- Rutter M. Implications of resilience concepts for scientific understanding. Annals of the New York Academy of Sciences. 2006;1094:1–12. doi: 10.1196/annals.1376.002. [DOI] [PubMed] [Google Scholar]
- Sackett GP. Exploratory behavior of rhesus monkeys as a function of rearing conditions and sex. Developmental Psychology. 1972;6:260–270. [Google Scholar]
- Schrijver NC, Pallier PN, Brown VJ, Wurbel H. Double dissociation of social and environmental stimulation on spatial learning and reversal learning in rats. Behavioral Brain Research. 2004;152(2):307–314. doi: 10.1016/j.bbr.2003.10.016. [DOI] [PubMed] [Google Scholar]
- Smotherman WP, Bell RW. Maternal mediation of early experiences. In: Smotherman WP, Bell RW, editors. Maternal influences and early behavior. New York: Spectrum Publications; 1980. [Google Scholar]
- Stetz MC, Thomas ML, Russo MB, Stetz TA, Wildzunas RM, McDonald JJ, et al. Stress, mental health, and cognition: A brief review of relationships and countermeasures. Aviation, Space, and Environmental Medicine. 2007;78 (5 suppl):B252–B260. [PubMed] [Google Scholar]
- Tang AC, Akers KG, Reeb BC, Romeo RD, McEwen BS. Programming social, cognitive, and neuroendocrine development by early exposure to novelty. Proceedings of the National Academy of Sciences USA. 2006;103(42):15716–15721. doi: 10.1073/pnas.0607374103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tronick E. The inherent stress of normal daily life and social interaction leads to the development of coping and resilience, and variation in resilience in infants and young children: Comments on the papers of Suomi and Klebanov & Brooks-Gunn. Annals of the New York Academy of Sciences. 2006;1094:83–104. doi: 10.1196/annals.1376.008. [DOI] [PubMed] [Google Scholar]
- Turner RJ, Lloyd DA. Cumulative adversity and drug dependence in young adults: Racial/ethnic contrasts. Addiction. 2003;98(3):305–315. doi: 10.1046/j.1360-0443.2003.00312.x. [DOI] [PubMed] [Google Scholar]
- Wallis JD, Dias R, Robbins TW, Roberts AC. Dissociable contributions of the orbitofrontal and lateral prefrontal cortex of the marmoset to performance on a detour reaching task. European Journal of Neuroscience. 2001;13(9):1797–1808. doi: 10.1046/j.0953-816x.2001.01546.x. [DOI] [PubMed] [Google Scholar]
- Yamasue H, Abe O, Suga M, Yamada H, Inoue H, Tochigi M, et al. Gender-common and -specific neuroanatomical basis of human anxiety-related personality traits. Cerebral Cortex. 2008;18(1):46–52. doi: 10.1093/cercor/bhm030. [DOI] [PubMed] [Google Scholar]