


Abstract
Transcripts of typical dicot plant plastid genes undergo C→U RNA editing at approximately 30 locations, but there is no consensus sequence surrounding the C targets of editing. The cis-acting elements required for editing of the C located at tobacco rpoB editing site II were investigated by introducing translatable chimeric minigenes containing sequence –20 to +6 surrounding the C target of editing. When the –20 to +6 sequence specified by the homologous region present in the black pine chloroplast genome was incorporated, virtually no editing of the transcripts occurred in transgenic tobacco plastids. Nucleotides that differ between the black pine and tobacco sequence were tested for their role in C→U editing by designing chimeric genes containing one or more of these divergent nucleotides. Surprisingly, the divergent nucleotide that had the strongest negative effect on editing of the minigene transcript was located –20 nt 5′ to the C target of editing. Expression of transgene transcripts carrying the 27 nt sequence did not affect the editing extent of the endogenous rpoB transcripts, even though the chimeric transcripts were much more abundant than those of the endogenous gene. In plants carrying a 93 nt rpoB editing site sequence, transgene transcripts accumulated to a level three times greater than transgene transcripts in the plants carrying the 27 nt rpoB editing sites and resulted in editing of the endogenous transcripts from 100 to 50%. Both a lower affinity of the 27 nt site for a trans-acting factor and lower abundance of the transcript could explain why expression of minigene transcripts containing the 27 nt sequence did not affect endogenous editing.
INTRODUCTION
In vascular plant chloroplasts, approximately 30 Cs encoded in primary RNA transcripts are edited by conversion to Us. Proper functioning of most genes carrying C targets of editing (‘editing sites’) is likely to depend upon editing, as the accompanying codon changes usually result in conservation of protein sequence between species or create AUG initiation codons (1,2). In a few cases, mutant proteins specified by sequences not corrected by editing have been shown to result directly in a mutant phenotype (1,3).
There is no obvious conserved feature of the nucleotide sequences surrounding an edited C that can be used to predict the location of a C target of editing. Nevertheless, sequences surrounding such Cs have been shown to be important in editing site selection. Transgenic plants have been constructed in which the chloroplasts carry transgenes that incorporate a C target of editing and varying amounts of surrounding sequence. Relatively small regions of surrounding sequence, in one case as small as 16 nt 5′ and 6 nt 3′ to the C target of editing (4), have been shown to be sufficient to allow editing in tobacco chloroplasts in vivo. Deletion series involving psbL and ndhB editing sites indicate that more nucleotides are required 5′ than 3′ to support editing (4–6).
From chloroplast transformation experiments, some information is available about the requirements for cis-acting sequences, but little is known about the trans-acting elements that act upon the C targets of editing. The existence of site-specific trans-acting factors has been established, however, by careful analysis of the effect of expression of additional editing sites within transgenic chloroplasts. The first observations were made by Chaudhuri et al. (6), who found that transgenic expression of additional transcripts carrying the psbL editing site resulted in decreased editing of the unaltered endogenous psbL gene’s transcript. Editing of C targets present in transcripts of other chloroplast genes was unaffected. The decreased editing has been interpreted as a ‘competition effect’ for a limiting quantity of a site-specific trans-acting factor. A competition effect has not been observed in all chloroplast transgenics that have been produced to date. When Bock et al. (5) expressed certain ndhB editing sites with varying amounts of surrounding sequence in separate transgenic plants, no effect on the editing of the endogenous transcripts was observed.
To study the requirements for sequences surrounding a C target of editing, we have performed transgenic analysis of an editing site present in the rpoB genes of a number of different species. The rpoB gene encodes the β subunit of a chloroplast-specific, DNA-dependent RNA polymerase (7). We previously showed that a maize sequence spanning 93 nt surrounding this rpoB editing site in maize (referred to as site I in maize, but as site II in tobacco, located at nucleotide 473, amino acid 158) is sufficient to direct editing of the site in transgenic tobacco plants (8). We used the heterologous species approach further by testing whether the homologous sequence from the more distant species, Pinus thunbergii (black pine; 9), could direct editing in tobacco. We reasoned that a finding of non-editing of the pine sequence, which diverges at 4 nt from the homologous sequence in tobacco, could focus attention on critical nucleotides that are required for recognition of C targets of editing. Indeed, the black pine sequence was not a satisfactory editing substrate in tobacco, and we therefore tested a series of single, double and triple nucleotide changes in the tobacco sequence based on the black pine sequence to evaluate whether particular nucleotide changes interfered with editing of the tobacco rpoB site II sequence. Surprisingly, we found that the identity of the nucleotide 20 bases 5′ of the C target has a strong effect on extent of editing of the transgenic transcripts. We also found that expression of a 27 nt rpoB editing site within minigene transcripts did not affect the extent of editing of the endogenous rpoB gene. In contrast, the editing of endogenous rpoB transcripts is affected in plants in which a 93 nt rpoB editing site was highly expressed from a tobacco transgene.
MATERIALS AND METHODS
Plant materials
Tobacco plants (Nicotiana tabacum cv. Petit Havana) were grown under sterile conditions on MS-agar medium (10) containing 30 g/l sucrose. Transformed lines were rooted and propagated on the same medium. One plastid-transformed line, MR210, containing a 93 nt sequence surrounding an rpoB editing site, was propagated from seed derived from the MR210 transgenic plant described by Reed and Hanson (8).
Oligonucleotides
The following oligonucleotides were used in this study:
214f, CGCCATGGATCAGATTGGGGAGGAAGATCAGAATTTCTAGAAT;
215f, CGCCATGGTTCAGATTGGGGAAGAAGATCGA-AATTTCTAGAAT;
411f, GATCCCCATGGATCAGATTGGGGAGGAAGATCAGAATTTCTAGACGG;
412f, GATCCCCATGGTTCAGATTGGGGAGGAAGATCAGAATTTCTAGACGG;
413f, GATCCCCATGGATCAGATTGGGGAAGAAGATCAGAATTTCTAGACGG;
414f, GATCCCCATGGATCAGATTGGGGAGGAAGATCAAAATTTCTAGACGG;
415f, GATCCCCATGGTTCAGATTGGGGAAGAAGATCAGAATTTCTAGACGG;
416f, GATCCCCATGGTTCAGATTGGGGAGGAAGATCAAAATTTCTAGACGG;
417f, GATCCCCATGGATCAGATTGGGGAAGAAGATCAAAATTTCTAGACGG;
419f, GATCCCCATGGTTCAGATTGGGGAAGAAGATCAAAATTTCTAGACGG;
Trps16α3.1, CTACCCCCCTTTTTGTATTTCCTTAATTTATTTCC;
PPrrn2, AATACGAAGCGCTTGGATACAGTTGTAGGGA;
mzB5.1, TCACCTATTCTTCCGAATTATATGTATCCGCG;
T7HT, GTAATACGACTCACTATAGGGC;
PC1.1, TCTTGAACAACTTGGAGCCGGGCC;
PCα1.2, GAGGATAGCAAGTTCCAAATTCTGTCTCGG;
oNP90, CGAGCCCATATCCTTGCTTTTCTAT;
oNP91, GAAAGGGATGCTGTGTATGAATCACTCAC;
oNP92, CCACCTACACAAGCAAATTGTTGATAAA.
Construction of plastid transformation vectors
The forward oligonucleotides listed above (designated by the ‘f’) contain the tobacco rpoB editing site II with various nucleotide modifications (Fig. 1). Each oligonucleotide was annealed with its reverse complement (sequence not shown) and digested with restriction enzymes NcoI and XbaI, which cut at the 5′ and 3′ ends of the oligonucleotides, respectively. The resulting fragments were ligated into vector pLAA24A (11), which had been digested with the same enzymes to remove the uidA coding sequences (Fig. 2A). The resulting plasmids MR214, MR215, MR411–MR417 and MR419, contain the aadA spectinomycin resistance gene and test sequences flanked by trnV-rps12/7 plastid-derived sequences, which direct their insertion into the inverted-repeat region of the tobacco plastid genome. Plasmids were prepared by using the Qiagen purification system and sequenced to confirm that the inserts contained the correct sequences.
Figure 1.

Sequences surrounding rpoB editing site II. (A) Nucleotide sequence from tobacco and black pine located –20 to +6 relative to rpoB editing site II. The C→U editing site is indicated by an arrow. Differences between the two sequences are shown as bold in the black pine sequence. (B) Plasmid constructs containing test sequences were named as indicated. The bold nucleotides denote differences between the endogenous tobacco sequence (MR411) and the test sequence.
Figure 2.
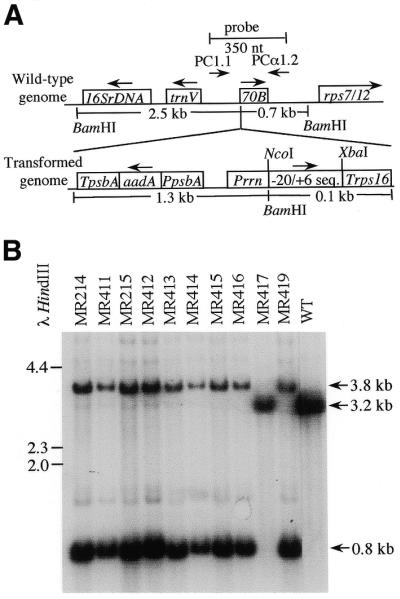
Analysis of DNA from transplastomic tobacco plants. (A) Schematic representation of the wild-type plastid genome and the inserted sequences in the transformed genome. –20/+6 sequence represents the region in which potentially edited test sequences were cloned. See Figure 1 for exact nucleotide sequence. (B) DNA was isolated from leaves of transformed and control plants (WT) and digested with BamHI. DNA gel blots were probed with 350 nt of plastid genome sequence surrounding the plasmid integration site. Sizes of λ DNA digested with HindIII are indicated on the left, in kb.
Plastid genome transformation
Either leaves from 6-month-old plants or 14-day-old seedlings of tobacco cultivar Petit Havana rooted in MS-agar medium were bombarded with plasmid-coated tungsten particles by using a model PDS1000/He Biolistic Particle Delivery System (Bio-Rad). Transformed tissue was regenerated essentially as described by Svab and Maliga (12). A 2 µg amount of plasmid DNA was precipitated on tungsten particles for each shot, and each plate was bombarded twice. DNA was isolated from transformed tissue by the CTAB (hexadecyltrimethylammonium bromide) method (13) and was analyzed for aadA and rpoB incorporation by PCR amplification.
DNA blot analysis
Total DNA (1 µg) from transformed or wild-type leaves was digested with BamHI, electrophoresed and blotted onto a positively charged nylon membrane (Amersham) using a Turboblotter (Schleicher and Schuell). The blot was hybridized with a 32P-labeled random primed (DECAprime II, Ambion) 350 nt genomic probe produced from PCR amplification of wild-type DNA using primers (PC1.1 and PCα1.2) surrounding the vector integration site in the chloroplast genome.
Analysis of editing in chloroplast transcripts
Total RNA was isolated from transformed and wild-type leaves using RNeasy Plant Mini Kit methods (Qiagen). Contaminating DNA was removed by a 30 min treatment with DNA-free reagent (Ambion). Reverse transcription (SuperScript II, Gibco BRL) using primers specific for transformed sequences (Trps16α3.1) or endogenous rpoB sequences (mzBα3.2) produced cDNA. The cDNA was PCR amplified using primers PPrrn2 and Trps16α3.1 for transformed sequences and primers mzB5.1 and mzBα3.2 for endogenous rpoB sequences surrounding the editing site. PCR products were cloned into plasmid pCR2.1-TOPO (Invitrogen) and individual clones were sequenced using BIG DYE terminators (PE Biosystems) and primer T7HT followed by analysis on ABI 3700 and ABI 377 DNA Sequencers (Cornell BioResource Center).
A restriction assay for editing extent was also performed on the RT–PCR products of the transgene transcripts. Two micrograms of RNA, extracted and DNase treated as above, was primed with 20 pmol of a degenerated hexamer (BRL) at 75°C for 10 min. The first strand cDNA was synthesized using the Omniscript RT kit (Qiagen). The transgene cDNA was then PCR amplified with the oligonucleotides Trps16α3.1 and PPrrn2. The PCR products were subsequently cloned in the pCR2.1-TOPO vector (Invitrogen). Single white colonies were cultured overnight and 10 µl of each culture was transfered to an individual well of a 96-well PCR microtiter plate. Each well was supplemented with 40 µl of a PCR mix [5 µl of 10× buffer (BRL), 1.5 µl of 50 mM MgCl2 (BRL), 0.5 µl Platinium Taq polymerase (5 U/µl, BRL), 1 µl of 10 mM dNTP, 1 µl of 10 µM Trps16α3.1, 1 µl of 10 µM PPrrn2 and 30 µl water]. A PCR reaction was then performed with a 58°C annealing step (PTC-200, MJ Research) and 10 µl of the PCR product was digested with 5 U of Sau3AI (NEB). After 6 h at 37°C the digest reaction was loaded on a 4% NuSieve agarose gel (FMC Bioproducts) and electophoresed in 1× TBE.
A third method to analyse the editing level of transcripts consisted of a poisoned primer extension performed on RT–PCR products. The first strand cDNA (made with degenerate hexamer) was amplified by PCR with the primers oNP91 and oNP92 surrounding editing site II. The amplification product (510 bp), was purified (QIAquick purification kit, Qiagen) quantified on agarose gel and then subjected to a primer extension reaction using radiolabeled oNP90. 100 ng of oNP90 was radiolabeled with 100 µCi of [γ-32P]ATP, using 10 U of T4 kinase (BRL), for 2 h at 37°C. The oligonucleotide was separated from the non-incorporated [γ-32P]ATP using a NucTrap column (Stratagene, La Jolla, CA). A 20 µl poisoned primer extension reaction contained 80–100 ng of RT–PCR template, 5 ng of labeled oNP90, 2 µl of 10× buffer (BRL), 0.6 µl of 50 mM MgCl2 (BRL), 0.2 µl Platinium Taq polymerase (5 U/µl, BRL), 50 µmol of each dATP, dTTP and dCTP (BRL) and 500 µmol ddGTP (Amersham Pharmacia Biotech). The reaction was amplified in a thermocyler (PTC-100, MJ Research) under the following conditions: 94°C for 2 min, followed by 50 cycles of 94°C for 15 s, 60°C for 30 s and 72°C for 15 s. The reaction was stopped by adding 5 µl of DNA sequencing stop solution (Promega), and then heated at 72°C for 2 min. Five microliters of each sample was loaded on a 12% acrylamide sequencing gel (SequaGel, National Diagnostics), and electrophoresed in 1× TBE for 2 h. The gel was exposed to a phosphorimager screen (Molecular Dynamics) for 2 h and was then scanned using a Storm 840 (Molecular Dynamics).
RNA blot analysis
A 10 µg amount of total RNA was electrophoresed on a 1.5% agarose–0.44 M formaldehyde–MOPS gel and blotted onto Hybond-N+ (Amersham) with a Turboblotter (Schleicher and Schuell). Hybridization was performed with an antisense probe matching the 27 nt sequence in construct 411, labeled using the Starfire system (Integrated DNA Technologies).
Nucleotide sequence accession numbers
The GenBank accession number for tobacco rpoB is X12745 and that for the P.thunbergii chloroplast genome is D17510.
RESULTS
Integration of rpoB editing site II into the tobacco chloroplast genome
Four C targets of editing have been identified within the chloroplast rpoB transcript (14). The vector pLAA24A was digested with NcoI and XbaI to remove the uidA coding sequences, which were replaced by oligonucleotides spanning the region –20 to +6 surrounding tobacco site II or the homologous chloroplast region from black pine (Fig. 2A). The resultant plasmids were named MR214 (tobacco) and MR215 (black pine). MR215 contains four nucleotide differences from the tobacco sequence (Fig. 1A). One such difference, the substitution of a G for the A immediately following the edited C in tobacco, is also present in the homologous maize rpoB sequence that previously was shown to be edited in tobacco (8), and therefore was excluded from further analysis. The rpoB test sequences are surrounded by regulatory regions from the chimeric Prrn operon promoter and the 3′ region of the rps16 ribosomal protein gene (Fig. 2A). The NcoI restriction site at the 5′ end of each insert contains an AUG start codon such that each construct contains a translatable minitransgene. The reading frame of the minigene has been shifted from the native frame, but this should have no effect on editing of the transcript since editing and translation are not linked in chloroplasts (15).
Transformation and selection of transformed plants
Tobacco leaves were bombarded with DNA-coated tungsten followed by selection on spectinomycin-containing regeneration medium. Homologous recombination between plasmid sequences and chloroplast DNA resulted in integration of the selectable marker aadA and test sequences into the chloroplast genome (Fig. 2A). Successive rounds of selection on spectinomycin-containing medium produced transformed tissue that expressed aadA resistance and the test sequences. Of 67 leaves bombarded with MR214, 53 spectinomycin-resistant shoots were obtained, of which one was a true transformant that had integrated the aadA resistance gene into its chloroplast genome. Following bombardment of 20 leaves with MR215, 30 spontaneous resistant plantlets and one transformant were obtained. These results represent a much greater incidence of development of spontaneous spectinomycin resistance during the selection process than we and others had previously observed (6,8).
In order to assess the degree of homoplasticity, total leaf DNA digested with BamHI was subjected to agarose gel electrophoresis, transferred to nylon membrane and probed with a 350 nt fragment homologous to sequences surrounding the plasmid integration site in the chloroplast genome (Fig. 2B). The transgenic plants show novel hybridization signals at 3.8 and 0.8 kb as well as a small amount of signal at 3.2 kb, which is characteristic of the wild-type untransformed genome. Either the plants are not entirely homoplastomic after >12 months on selective regeneration medium, or the probe is hybridizing with chloroplast DNA fragments that are present in the nucleus or mitochondrion (16). We were able to analyze RNA specifically expressed from the transformed sequences by amplifying cDNA with PCR primers derived from the sequences of the 5′ and 3′ regulatory regions surrounding the editing test sequences.
Analysis of transgenic transcripts carrying a 27 nt tobacco or black pine rpoB site II sequence
To evaluate the extent of editing of transgenic transcript, cDNA clones were produced by isolating total RNA from leaves from regenerating shoots, followed by RT–PCR amplification of cDNA. Individual clones of RT–PCR products were then sequenced. When 16 cDNA clones of MR214 transcripts were sequenced, nine exhibited the genomic sequence and a T was present in the remaining seven (Table 1), indicating that a 27 nt tobacco sequence was adequate to confer editing upon a chimeric transcript. MR215 transcripts exhibited almost no editing with a T present in only one of the 75 clones that were analyzed. We concluded that some or all of the nucleotide differences between the tobacco and black pine test sequences interfered with editing of rpoB site II.
Table 1. Analysis of rpoB site II editing in transplastomic plants.
| Construct | Transgene | Endogenous | Nucleotides | ||||
| |
edited |
edited |
–20 |
–7 |
+2 |
||
| MR214 | 7/16 | + | 27/28 | + | A | G | G |
| MR215 | 1/75 | – | 22/22 | + | T | A | A |
| MR411 | 17/96 | + | 37/42 | + | A | G | G |
| MR412 | 0/69 | – | 17/18 | + | T | G | G |
| MR413 | 29/91 | + | 22/24 | + | A | A | G |
| MR414 | 13/45 | + | 20/21 | + | A | G | A |
| MR415 | 2/45 | – | 25/28 | + | T | A | G |
| MR416 | 0/32 | – | 27/28 | + | T | G | A |
| MR417 | 9/35 | + | 22/23 | + | A | A | A |
| MR419 | 2/45 | – | 25/26 | + | T | A | A |
The number of edited rpoB site II transcripts is shown for both the transgene and the endogenous tobacco rpoB sequences. The nucleotide columns identify differences between tobacco (MR211, MR411) and black pine (MR215) rpoB sequences in the region –20 to +6; bold underlined nucleotides are those encoded in black pine. Editing status was derived both by sequencing of individual cDNA clones and by the restriction digestion assay (see Materials and Methods).
In the transformed plants, the rpoB –20 to +6 region was expressed as an additional copy of the editing site introduced into the inverted-repeat region of the chloroplast genome, while in each genome the endogenous rpoB gene is present in the single-copy region. Furthermore, the additional rpoB editing sites are present in a minigene controlled by the strong Prrn promoter. We examined editing of the endogenous tobacco rpoB transcript in order to determine whether the presence of additional transcripts carrying the editing site had affected the degree of editing of the normal rpoB transcripts. To amplify the endogenous transcripts specifically, oligonucleotide primers were used that were homologous to regions of rpoB not contained in the transgene. In the plants expressing MR214 and MR215, the endogenous rpoB site was highly edited, with no statistically significant difference in the extent of rpoB site II editing between non-transgenic wild-type plants, MR214 and MR215 (Table 1). Previously, we had observed that all sequenced cDNAs of endogenous rpoB from wild-type plants exhibit RNA editing (data not shown).
Integration of modified rpoB editing site II into the tobacco chloroplast genome
Because the black pine sequence was not edited in tobacco, although a tobacco region of the same size was recognized and edited when expressed from the minitransgene, we produced a series of plasmid constructs with single, double and triple nucleotide alterations (Fig. 1B). The nucleotide differences tested were three of the four differences between black pine and tobacco. We did not test the A→G difference at the +1 position, since the presence of G did not affect editing of the maize rpoB sequence in tobacco (8). Eight different plasmids were produced as described above using oligonucleotides encoding the altered sequences.
In an effort to increase the number of chloroplast transformants generated from tissue bombardment, we modified our protocol from bombarding 6-month-old leaf tissue to bombarding young tobacco seedlings. Originally, tobacco plants were grown for 6 months in Magenta boxes, and leaves were harvested when they were ∼4 × 5 cm. Improved results were obtained when we bombarded 0.5 × 0.5 cm leaves on 14-day-old seedlings. The total leaf area bombarded per seedling plate was ∼5 cm2 compared to 20 cm2 per plate of older leaves. On average, a single plate containing 10–12 tobacco seedlings produced one spectinomycin-resistant shoot. Approximately 50% of the shoots represent true transformants containing aadA sequences integrated into the chloroplast genome, while the other half result from spontaneous spectinomycin resistance that developed during selection. Starting with seedling leaves rather than older leaves represents a significant improvement in the ratio of transformed shoots to spontaneously-arising, spectinomycin-resistant shoots relative to the ratio we obtained previously (constructs MR214 and MR215 above; 8).
The degree of homoplasmy of the transformed plants was assessed by analyzing total DNA from regenerated shoots (Fig. 2B) as described above. With the exception of the shoots containing MR417, all of the transformed plants are >99% homoplastomic. Although a signal characteristic of the transgene is not evident in the MR417 lane of Figure 2B, these plants are transformed, as shown by a longer exposure of the membrane (data not shown).
Analysis of endogenous and modified rpoB transcripts in transformed tobacco plastids
In order to assess editing extent of the minitransgene transcripts, individual clones of cDNAs were produced from transformed plants and sequenced as described above or analyzed by restriction digestion with an enzyme that distinguishes cDNA from edited versus unedited transcripts (Fig. 3). There appeared to be two categories of chimeric transcripts—those that could be edited to a moderate degree and those that were rarely edited, if at all (Table 1). There is no statistically significant difference between the editing extents of the transcripts rated ‘plus’. The editing extents of the transcripts rated ‘minus’ also do not exhibit statistically significant differences, though a few edited transcripts were detected in three of the five ‘minus’ plants. Surprisingly, the nucleotide alteration most distant (at –20) from the C target of editing appears to exert the largest effect on editing efficiency. Minigene transcripts from any plant carrying the –20 T were very poorly edited, if at all, while plants carrying the A at –7 or +2 could be edited as efficiently as the wild-type sequence in the minigene transcripts.
Figure 3.

Restriction endonuclease assay to detect editing of the transgene transcripts. (A) Sau3AI (S) restriction map of the edited or non-edited RT–PCR products of the MR413 transgene with primers Trps16α3.1 and PPrrn2 (130 bp product). The size in bp is written under each restriction fragment. (B) NuSieve agarose gel (4%) of a Sau3AI restriction digest of 130 bp PCR product. PCR was performed on cloned and sequenced, edited (MR413T) or non-edited (MR413C) cDNA. Lanes 1–6 represent independent cloned RT–PCR products from a MR413 plant. Lane 5 corresponds to an edited RT–PCR product.
The extent of editing of the endogenous rpoB transcripts in plants carrying the 27 nt rpoB site II in chimeric transcripts was also assessed by sequencing individual cDNAs (Table 1), as described above. According to sequence analysis, the transcripts were all highly edited, with no statistically significant difference between extent of editing in different plants (Table 1). Extent of editing of endogenous transcripts in transgenic plants that had incorporated minigenes carrying either 27 or 93 nt sequences surrounding the rpoB site II were also assessed by a poisoned primer extension assay. According to this assay, the presence of minigenes carrying a 27 nt site did not affect endogenous rpoB editing (Fig. 4). However, as previously observed by cDNA sequencing (8), expression of the 93 nt rpoB site in chimeric transcripts resulted in reduction of endogenous rpoB site II editing to 50% (data not shown).
Figure 4.
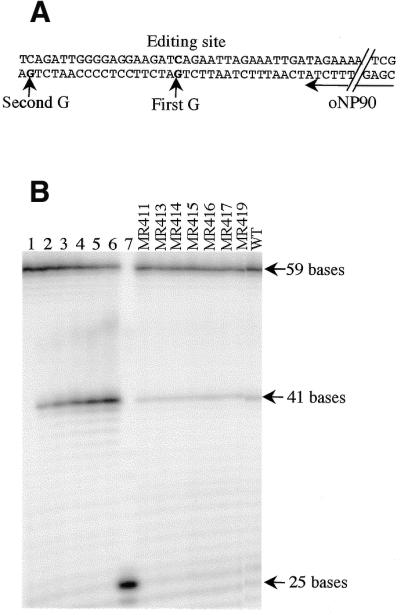
Poisoned primer extension assay to analyze endogenous rpoB transcript editing. (A) Portion of the sequence of the RT–PCR product (primers oNP91, oNP92, 510 bp amplicon) on which the poisoned primer extension with primer oNP90 (25 bp long) is performed. The primer extension is poisoned by addition of ddGTP. When the RNA is not edited, oNP90 is extended only to the first G (arrow); when the RNA is edited, the corresponding cDNA does not contain the first G, and oNP90 is extended to the second G (arrow). When extension is stopped at the first G, the length of the extended primer is 41 bases; if the extension goes to the second G, the length is 59 bases. (B) Autoradiograph of an acrylamide gel of primer extension products in which radiolabeled oNP90 was used with different RT–PCR templates. Lanes 1–6 consist of serial mixtures of PCR product (oNP91, oNP92) from a cloned edited and non-edited rpoB cDNA. Lane 1, 100 ng edited; lane 2, 80 ng edited plus 20 ng non-edited; lane 3, 60 ng edited plus 40 ng non-edited; lane 4, 40 ng edited plus 60 ng non-edited; lane 5, 20 ng edited plus 80 ng non-edited; and lane 6, 100 ng non-edited. Lane 7 is a control without template. In the next lanes, 80–100 ng of RT–PCR product (oNP91, oNP92) was used as template. The relative intensity of the 41 and 59 bases bands produced from total RNA isolated from transformed or control (WT) plants is an indication of the level of editing of the tobacco rpoB site II in the different plants tested.
Analysis of the level of rpoB RNAs present in transgenic plants
To observe the degree of expression of the minitransgene and endogenous rpoB RNA, total RNA was electrophoresed on an agarose gel, blotted, and probed with an oligonucleotide probe complementary to the 27 nt rpoB site II sequence incorporated into the minitransgene. The transformed plants exhibit a 0.2 kb transcript, as would be predicted from the size of the transgene including the 5′ and 3′ UTRs (Fig. 5). In addition, both the wild-type and transgenic plants express the endogenous rpoB messages of 1.8 and 3.2 kb (17). The transgenic transcripts are present at substantially higher levels than the endogenous transcript (Fig. 5). While some of the 200 nt signals in the different transgenic lines appear at different intensities, this does not reflect different transcript amounts in those cases where the oligonucleotide was not a perfect complement. For example, the weaker signal to the 200 nt transcript in MR415 may be explained because the transcript has 2 nt that are non-complementary to the 27 nt oligonucleotide probe. Nevertheless, it is quite obvious that transcripts in the MR210 line, which also exhibit 2 nt that are non-complementary to the probe, are far more abundant than the transgene transcripts in MR411, which carries perfectly complementary transcripts. Estimates from densitometry measurements indicate the minigene transcripts in MR210 are at least five times more abundant than those in MR411 (Fig. 5). Because both minigene cassettes carry the same promoter and terminator sequences, and differ only in the size of the coding region insertion, perhaps the smaller transcripts accumulate to a lower extent because of lower RNA stability.
Figure 5.
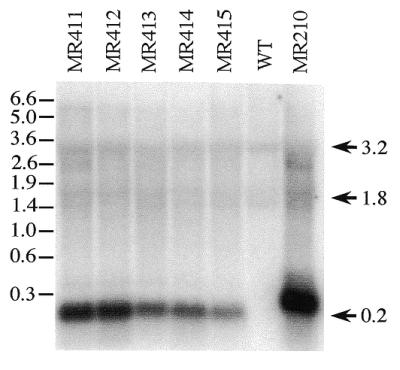
Analysis of RNA from transplastomic plants. Total RNA isolated from leaves of controls (WT) and transformed plants was probed with an oligonucleotide antisense to the 27 nt editing site sequence in MR411. Approximately 10 µg of each RNA sample was run per lane. Endogenous rpoB bands are under exposed in order to visualize the transgene. MR210 is transformed with the 93 nt rpoB editing site; all other transformants carry the 27 nt editing site. Sizes of RNA markers (Promega) are indicated on the left, in kb.
DISCUSSION
Because of the time and effort required to produce plants carrying transgenic chloroplasts, relatively few editing sites have been subjected to analysis of cis-acting elements in vivo. In addition to the rpoB editing site, studies have been performed on editing sites found in the psbL, ndhD and ndhB genes (4–6,18,19). With the exception of a region carrying tobacco ndhB editing sites II and III, which exhibited no editing when sequences 42 nt 5′ and 3′ of the C targets of editing were incorporated into a transgene, a relatively small amount of sequence information surrounding a C target of editing has been sufficient to direct editing of sites present in chimeric transgenes. In the case of the tobacco rpoB site II, only 20 nt 5′ and 6 nt 3′ of the C target were sufficient to allow editing, though the editing efficiency was much lower than that of the rpoB site II in normal, non-transgenic plants. Furthermore, the transcripts from the endogenous rpoB gene present in the transgenic plants carrying the rpoB gene were nearly completely edited. These data indicate that the minigene transcripts containing the 27 nt editing site are not as suitable a substrate for the editing machinery as the larger rpoB transcripts.
When plants carrying a 93 nt segment surrounding rpoB site II in transgenic tobacco plastids were previously analyzed (8), a greater degree of transgene transcript editing (∼50%) was observed than in the plants reported here that contain a 27 nt segment around rpoB. Furthermore, expressing the 93 nt segment results in reduced editing of the homologous site in the transcripts expressed from the endogenous rpoB genes. Diminished editing of the homologous endogenous site was also observed when psbL and ndhD editing sites were expressed from chimeric transgenes (4,6). These data have been interpreted as a ‘competition effect’ for site-specific trans-acting factors. When expression of a chimeric gene carrying a particular editing site results in reduced editing of the homologous endogenous site, but does not reduce editing at non-homologous sites, perhaps there are limiting quantities of a necessary trans-acting factor needed for editing of a particular site. In the case of the minigene transcripts carrying the 27 nt rpoB editing site II sequences, no competition effect was detected. This could be due to a lower affinity of the site-specific factor(s) for the 27 nt fragment so that more such factors were available for the editing of the endogenous transcript. Indeed, minigene transcripts carrying only the 27 nt editing site were edited at lower efficiency than the minigene transcripts carrying the 93 nt editing site.
Whether or not transgene transcripts exert a competition effect should also be dependent on the relative abundance of the transgene versus endogenous transcripts. Transcription of both the 93 and 27 nt editing sites occurred from a Prrn promoter present in the transgene. The 93 nt minigene transcript was much more abundant than the endogenous transcript, with a high ratio of transgene/endogenous RNA (8). Minigene transcripts carrying the 27 nt site also exhibited a high transgene/endogenous transcript ratio but the transgene transcripts are relatively less abundant than those in the plants carrying the 93 nt sequence (Fig. 5). Both the lower transcript abundance and a lower affinity for the 27 nt-containing transcript could explain why a competition effect was not observed. With a smaller number of transgene transcripts and lower affinity, there may then be sufficient trans-acting factor(s) to permit thorough editing of the endogenous rpoB transcripts.
In one other set of transgenic plants no competition effect was observed. These plants carried the ndhB editing sites expressed at a high level (5). Unfortunately, the relative ratio of the transgenic versus endogenous ndhB transcripts was not reported, so it is not known to what degree the expression of the transgene affected the total concentration of transcripts carrying the ndhB sites in the plastids. Possibly there are ndhB site-specific factors that are normally present in excess so that there are sufficient factors to allow efficient editing of both the transgene and the endogenous gene’s transcripts. Alternatively, editing of the ndhB sites may not require site-specific trans-acting factors at all. Without a competition effect, there is no evidence of such a factor.
Perhaps our most surprising observation in the present study is the strong effect of a single alteration –20 nt from the C target of editing. The presence of a T instead of the A present in the endogenous sequence virtually abolished editing. Most of the reported single nucleotide alterations that have strong effects on editing have either been immediately 5′ or 3′ to the C target (4). The most distant single nucleotide change reported previously exerting a strongly negative effect on editing is at –9 from ndhB site V (18). Because in neither the ndhB site V nor our rpoB site II was a competition effect observed on editing of the endogenous genes, we cannot conclude whether the single nucleotide alterations reduced the binding of a hypothetical required trans-acting factor, or changed the transcript into an unsuitable substrate for the editing enzyme, or both.
In our study, the –20 position was tested because of its divergence from the tobacco sequence in the unedited heterologous black pine rpoB sequence. It is unlikely that the –20 position would have been among the nucleotides tested in a set of transgenic plants had the data from the heterologous site not focused attention on the divergence at this location. Given the time-consuming nature of transgenic chloroplast experiments, choosing nucleotides for investigation based on a non-edited heterologous site may prove to be a valuable approach in analysis of additional editing sites.
Acknowledgments
ACKNOWLEDGEMENTS
We thank undergraduate lab assistants Melanie Irving and Danielle Knight for technical assistance, and Pal Maliga for providing the chloroplast transformation vector. This work was supported by NIH GM17743 NRSA postdoctoral grant to M.L.R and NIH grant GM50723 to M.R.H.
References
- 1.Bock, R., Kössel,H. and Maliga,P. (1994) Introduction of a heterologous editing site into the tobacco plastid genome: the lack of RNA editing leads to a mutant phenotype. EMBO J., 13, 4623–4628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Maier R.M., Neckermann,K., Igloi,G.L. and Kössel,H. (1995) Complete sequence of the maize chloroplast genome: gene content, hot-spots of divergence and fine tuning of genetic information by transcript editing. J. Mol. Biol., 251, 614–628. [DOI] [PubMed] [Google Scholar]
- 3.Zito F, Kuras,R., Choquet,Y., Kossel,H. and Wollman,F.A. (1997) Mutations of cytochrome b(6) in Chlamydomonas reinhardtii disclose the functional significance for a proline to leucine conversion by petB editing in maize and tobacco. Plant Mol. Biol., 33, 79–86. [DOI] [PubMed] [Google Scholar]
- 4.Chaudhuri S. and Maliga,P. (1996) Sequences directing C to U editing of the plastid psbL mRNA are located within a 22 nucleotide segment spanning the editing site. EMBO J., 21, 5958–5964. [PMC free article] [PubMed] [Google Scholar]
- 5.Bock R., Hermann,M. and Kössel,H. (1996) In vivo dissection of cis-acting determinants for plastid RNA editing. EMBO J., 15, 5052–5059. [PMC free article] [PubMed] [Google Scholar]
- 6.Chaudhuri S., Carrer,H. and Maliga,P. (1995) Site-specific factors mediate RNA editing of the psbL mRNA in tobacco plastids. EMBO J., 12, 2951–2957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Allison L.A., Simon,L.D. and Maliga,P. (1996) Deletion of rpoB reveals a second distinct transcription system in plastids of higher plants. EMBO J., 15, 2802–2809. [PMC free article] [PubMed] [Google Scholar]
- 8.Reed M.L. and Hanson,M.R. (1997) A heterologous maize rpoB editing site is recognized by transgenic tobacco chloroplasts. Mol. Cell. Biol., 17, 6948–6952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wakasugi T., Hirose,T., Horihata,M., Tsudzuk,T., Kössel,H. and Sugiura,M. (1996) Creation of a novel protein-coding region at the RNA level in black pine chloroplasts: The pattern of RNA editing in the gymnosperm chloroplast is different from that in angiosperms. Proc. Natl Acad. Sci. USA, 93, 8766–8770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Murashige T. and Skoog,F. (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant., 15, 473–497. [Google Scholar]
- 11.Zoubenko O.V., Allison,L.A., Svab,Z. and Maliga,P. (1994) Efficient targeting of foreign genes into the tobacco plastid genome. Nucleic Acids Res., 22, 3819–3824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Svab Z. and Maliga,P. (1993) High frequency plastid transformation in tobacco by selection for a chimeric aadA gene. Proc. Natl Acad. Sci. USA, 90, 913–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fulton T.M., Chunwongse,J. and Tanksley,S.D. (1995) Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep., 13, 207–209. [Google Scholar]
- 14.Hirose T., Kusumegi,T., Tsudzuki,T. and Sugiura,M. (1999) RNA editing sites in tobacco chloroplast transcripts: editing as a possible regulator of chloroplast RNA polymerase activity. Mol. Gen. Genet., 262, 462–467. [DOI] [PubMed] [Google Scholar]
- 15.Zeltz P., Hess,W.R., Neckerman,K., Börner,T. and Kössel,H. (1993) Editing of the chloroplast rpoB transcript is independent of chloroplast translation and shows different patterns in barley and maize. EMBO J., 12, 4291–4296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Maliga P. and Nixon,P.J. (1998) Judging the homoplastomic state of plastid transformants. Trends Plant Sci., 3, 376–377. [Google Scholar]
- 17.Hess W.R., Prombona,A., Fieder,B., Subraminian,A.R. and Börner,T. (1993) Chloroplast rps15 and the rpoB/C1/C2 gene cluster are strongly transcribed in ribosome-deficient plastids: evidence for a functioning non-chloroplast-encoded RNA polymerase. EMBO J., 12, 563–571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bock R., Hermann,M. and Fuchs,M. (1997) Identification of critical nucleotide positions for plastid RNA editing site recognition. RNA, 3, 1194–2000. [PMC free article] [PubMed] [Google Scholar]
- 19.Hermann M. and Bock,R. (1999) Transfer of plastid RNA-editing activity to novel sites suggests a critical role for spacing in editing-site recognition. Proc. Natl Acad. Sci. USA, 96, 4856–4861 [DOI] [PMC free article] [PubMed] [Google Scholar]