

Abstract
The vast majority of word meanings are learned simply by extracting them from context, rather than by rote memorization or explicit instruction. Although this skill is remarkable, little is known about the brain mechanisms involved. In the present study, ERPs were recorded as participants read stories in which pseudowords were presented multiple times, embedded in consistent, meaningful contexts (referred to as meaning condition, M+) or inconsistent, meaningless contexts (M−). Word learning was then assessed implicitly using a lexical decision task and explicitly through recall and recognition tasks. Overall, during story reading, M− words elicited a larger N400 than M+ words, suggesting that participants were better able to semantically integrate M+ words than M− words throughout the story. In addition, M+ words whose meanings were subsequently correctly recognized and recalled elicited a more positive ERP in a later time-window compared to M+ words whose meanings were incorrectly remembered, consistent with the idea that the late positive component (LPC) is an index of encoding processes. In the lexical decision task, no behavioral or electrophysiological evidence for implicit priming was found for M+ words. In contrast, during the explicit recognition task, M+ words showed a robust N400 effect. The N400 effect was dependent upon recognition performance, such that only correctly recognized M+ words elicited an N400. This pattern of results provides evidence that the explicit representations of word meanings can develop rapidly, while implicit representations may require more extensive exposure or more time to emerge.
Introduction
Humans efficiently and rapidly acquire the meanings of new words throughout their lives. During the school years, children learn approximately 3000 new words per year, and by adulthood vocabulary size has typically reached tens of thousands of words and may exceed 100,000 words in exceptional cases (Nagy & Herman, 1987; Sternberg, 1987). Even more remarkable is the fact that the majority of these words are learned incidentally through context, in the absence of any type of intentional instruction (Nagy & Herman, 1987; Stenberg, 1987). In addition, unlike many aspects of language processing, which show robust critical period effects, adults show relatively normal acquisition of novel semantic information late in life and continue to acquire new vocabulary in their native language throughout their lives (Newport, Bavelier, & Neville, 2001; Weber-Fox & Neville, 1996).
Despite this unusually prolonged plasticity profile, surprisingly few studies have focused on native language (L1) word learning in adults. Furthermore, although overt strategy and instruction play a minimal role in L1 vocabulary acquisition, much of what is known about the neural and cognitive processes of L1 word learning in adults comes from intentional and highly artificial training paradigms (e.g., Breitenstein et al., 2005, 2007; Clay et al., 2007; Perfetti et al., 2005). Thus, one major goal of the present study was to investigate neural mechanisms that support meaning acquisition in adulthood, using a task in which novel words are learned through contextual exposure as in natural language. We were especially interested in examining whether these newly acquired representations were encoded preferentially into implicit or explicit memory.
Although a variety of different terms have been used to describe the distinction between implicit and explicit learning processes, in the present paper we use the term incidental learning to describe a mode of learning in which individuals learn without intention, or when they learn one thing when their primary objective was to learn something else (cf. Ellis, 1994). In contrast, implicit memory is defined as knowledge that can facilitate processing without the need for conscious recollection, revealed by indirect memory tests such as semantic priming tasks. Explicit memory refers to knowledge that is accessible to awareness and is measured by direct memory tests such as recognition and recall (Ellis, 1994; Squire & Zola, 1996). The product of incidental learning may be either implicit or explicit, depending upon whether the material learned is accessible to awareness.
A great deal of evidence indicates that implicit and explicit memory are mediated by different neural systems. These forms of memory have been dissociated empirically in numerous tasks across different domains (e.g., Graf, 1987; Reber, Kassin, Lewis, & Cantor, 1980), and also show distinct neural indices at encoding and retrieval (e.g., Paller, Hutson, Miller, & Boehm, 2003; Rugg, 1998; Schott, Richardson-Klavehn, Heinze, & Duzel, 2002). Perhaps the most striking evidence, however, comes from amnesic patients, who show marked impairment on tests of explicit memory but normal or near-normal performance on implicit memory tasks (e.g., Cohen & Squire, 1980; Graf & Schacter, 1985; Milner, Corkin, & Teuber, 1968). Of relevance to the present study, previous research has found that amnesic patients are often capable of forming novel semantic or conceptual associations as measured by implicit memory tasks, despite having no explicit knowledge of these associations (Goshen-Gottstein, Moscovitch, & Melo, 2000; Moscovitch, 1986; Verfaellie, Martin, Page & Keane, 2006; Gabrieli, Keane, Zarella, & Poldrack, 1997). These findings indicate that new semantic information may be encoded into implicit memory in the absence of explicit awareness, at least under some circumstances.
Behavioral Studies of Word Learning
Although relatively little attention to date has been directed towards L1 word learning in adults, a small number of behavioral studies have investigated the conditions necessary for novel words to become integrated into existing semantic networks. Breitenstein and colleages (2007) trained participants on an incidental vocabulary task, which involved a higher co-occurrence of "correct" arbitrary object and novel word pairings as compared to "incorrect" pairings. After five training sessions, novel words induced a cross-modal semantic priming effect in a semantic classification task equal to that elicited by real related words, suggesting that novel words may be represented similarly to existing words after sufficient associative training. Dumay, Gaskell and Feng (2004) presented participants with novel words embedded in isolated sentences over two successive days of training. Novel words did not semantically prime associated category names on a subsequent lexical decision task either immediately or 24 hours after training, but elicited significant priming one week later. Converging results were found by Clay, Bowers, Davis, and Hanley (2007), who presented participants with novel words that were repeatedly paired with both short definitions as well as pictures, in a single study session. At test, participants were required to name familiar pictures in English that were presented simultaneously with familiar words, trained novel words, or untrained novel words. An automatic Stroop-like picture-word interference effect to studied novel words was not immediately observed following the study phase, but emerged after a one-week delay despite the absence of any additional study. Findings from all of these studies are consistent with the idea that semantic integration processes occur over a relatively protracted period of time, and that implicit representations of new semantic information may form only after a period of incubation. In addition, providing meaningful semantic information about new words appears to be an important factor in the integration of novel words into existing conceptual and lexical networks.
Neural Substrates of Word Lexicalization
A recent neurocomputational account of word learning proposes that lexical acquisition occurs in two stages (Davis & Gaskell, 2009). The first stage involves rapid initial familiarization, mediated by the hippocampus. The second stage is one of slow offline lexical consolidation, mediated by neocortical areas. This model is consistent not only with behavioral studies of spoken word learning, but also with recent neuroimaging evidence. For example, two recent fMRI word learning studies demonstrated that novel vocabulary items initially elicit an elevated hippocampal response, followed by a subsequent decline over later presentations, and that the extent of this activity correlates with behavioral proficiency (Breitenstein et al., 2005; Davis, Di Betta, Macdonald, & Gaskell, 2008). These findings suggest that the hippocampus plays an important role in the successful learning of novel words, particularly during initial acquisition. Breitenstein and colleagues also found that increasing vocabulary proficiency was associated with intercorrelated activity in the hippocampus and left neocortical regions, providing evidence that successful acquisition of a new lexicon depends on an interaction between the hippocampus and neocortical regions. Finally, circumscribed neocortical regions show similar levels of activation to unfamiliar novel words and words that were learned on the day of scanning (Davis et al., 2008; Davis & Gaskell, 2009), but show significantly reduced activity to words that were learned on the previous day (Davis et al., 2008), supporting the idea that representations encoded by neocortical regions require time to develop.1 Although Davis & Gaskell's model focuses on acquisition of word form, a similar two-stage process may also mediate acquisition of word meaning, an idea that is consistent with results from semantic word-learning studies (Breitenstein et al., 2005; Clay et al., 2007; Dumay et al., 2004).
Electrophysiology of Word Learning
Event-related potential (ERP) studies of word learning have focused upon one particular brainwave component, the N400. The N400 is a negative-going component that peaks approximately 400 ms post-stimulus, with a posterior and bilateral distribution, whose amplitude varies as an inverse function of the subject’s expectancy for the upcoming word of a sentence (Kutas & Hillyard, 1980, 1984). Words that are semantically unexpected elicit larger amplitude N400 responses than words that are more expected given the preceding sentence context, leading to the hypothesis that the N400 reflects semantic processes of lexical integration (Friederici, Pfeifer, & Hahne, 1993; Holcomb & Neville, 1991; Kutas, Van Petten, & Besson, 1988).
Previous ERP studies in both L1 and L2 have found that the N400 is a sensitive index of word learning. McLaughlin and colleagues (2004) demonstrated that native English speakers learning French showed an N400 effect to French pseudowords relative to French words after only 14 hours of classroom instruction. This pseudoword effect occurred even while participants performed at chance levels when making explicit L2 word-nonword judgments. L1 learning can also result in rapid neural change. Perfetti, Wlotko, and Hart (2005) used flashcards to train participants on the definitions of rare unknown words in a 45-minute study session. Following training, participants performed a semantic decision task in which they indicated whether pairs of words were semantically related or not. An N400 effect was evoked by the unrelated trained words relative to the related trained words, and this effect was larger for more skilled comprehenders. Finally, Mestres-Misse, Rodriguez-Fornells, and Munte (2007) presented triplets of sentences with novel or real words in the terminal position of each sentence. The ERP response to novel words presented in meaningful contexts words became indistinguishable from the response evoked by real words over the three presentations, an effect which did not occur for novel words presented in meaningless contexts. In addition, an N400 effect was observed during a subsequent generalization task, in which participants were presented with the learned novel words and their real-word counterparts. These studies demonstrate that the N400 is an effective marker of semantic word learning.
The present study examined the neural signatures of incidental meaning acquisition in a passive task designed to closely mimic natural language learning, focusing in particular on the N400. ERPs were recorded as participants read simple fiction stories in which novel pseudowords were presented multiple times, embedded in consistent, meaningful contexts (referred to as meaning condition, M+) or inconsistent, meaningless contexts (M−). We hypothesized that acquiring the meanings of novel words should be indexed by a gradual reduction in the N400 over time, reflecting increasing facilitation of semantic integration processes. Therefore, the N400 elicited by M+ words was predicted to show a greater reduction over time than the response evoked by M− words, for which no meaning assignment was possible. We also hypothesized that the N400 to real words should be reduced relative to both M+ and M− words, reflecting overall facilitated semantic integration, and consistent with previous findings showing that pseudowords elicit a greater N400 component than real words (Holcomb & Neville, 1990, Holcomb, 1993; Bentin et al., 1999).
We also investigated whether novel words acquired during the learning phase were encoded preferentially by implicit or explicit memory systems. To this end, we examined behavioral and electrophysiological indices of word learning on an indirect memory test, namely lexical decision, and on two direct memory tasks, recall and recognition. In addition, we addressed whether the N400 effect depends upon explicit semantic knowledge by comparing ERP responses to words whose meanings were correctly identified with words whose meanings could not be identified, at both exposure and test. We hypothesized that if the N400 indexes semantic integration processes that are dependent upon explicit knowledge of word meanings, an N400 should be elicited only to words that were correctly recognized. In contrast, if the N400 reflects implicit learning and continuous change that occur independently of explicit knowledge, we might expect to observe an N400 to words that were both correctly and incorrectly identified. Finally, by comparing the N400 to real words relative to M+ words, we examined whether the neural substrates underlying these effects were largely overlapping or distinct. We predicted that these effects should be fairly similar, consistent with previous findings showing that the N400 can index word learning at very early stages of acquisition (McLaughlin et al., 2004; Perfetti et al., 2005; Mestres-Misses et al., 2007).
Methods
Participants
Twenty-one monolingual native English speakers (10 female) were recruited at the University of Oregon to participate in the experiment. One male participant was excluded from all analyses due to below-chance M+ word recognition performance. Participants were between 18–30 years old (M = 22.4, SD=2.9), were right-handed, had normal vision, and had no history of neurological problems. Participants were run in two sessions of approximately 2.5 hours each, conducted within the same week. In each session, they read two of the four stories and completed the corresponding lexical decision tasks, recognition tasks, and recall tasks, described in detail below. Participants were paid $10/hour.
Experimental Tasks
The story task consisted of reading four simple fiction stories, ranging from approximately 4000 to 5000 words in length, presented to participants on a computer monitor. Within each story, novel pseudowords were presented ten times, embedded in consistent, meaningful contexts (referred to as meaning condition, M+) or inconsistent, meaningless contexts (M− condition). In the M+ condition, each novel pseudoword consistently replaced a real English word that was originally in the story, while in the M− condition, each novel pseudoword replaced a different word at each presentation and thus had no consistent meaning. As a control condition, real English words (R condition) were also presented ten times throughout each story. All critical words occurred in mid-sentence positions. Table 1 presents examples of sentences embedded with critical words illustrating these three conditions. A total of 20 critical words, divided evenly among the three conditions, were presented throughout each story, yielding 26–27 trials per condition over the four stories. Conditions were counterbalanced across participants, such that each critical word position was experienced as an R word for some participants, an M+ word for other participants, and an M− word for a third group of participants. All critical words were concrete nouns of high frequency (mean 132.6 per one million words, Kucera & Francis, 1967). Novel pseudowords were matched phonologically to R words for place of articulation, number of syllables, and number of letters, and were assigned randomly (N = 8) or pseudorandomly (N = 12) to the M+ and M− conditions across participants in order to eliminate chance baseline word differences between conditions. Stories were presented one word at a time using semi-variable Serial Visual Presentation, a procedure designed for presenting written texts in EEG experiments at a natural reading rate (Otten and Van Berkum, 2008). Critical words were always presented at durations of 400 ms, separated by inter-word intervals of 107 ms. To ensure adequate comprehension, participants were required to answer two-alternative multiple-choice comprehension questions about the story's content at regular intervals. No overt instructions were given about the presence of the novel pseudowords.
Table 1.
Example of sentences presented during the story task, embedded with Real, M+, and M− critical words. Critical words are displayed in bold text.
| R condition: |
| Several white fluffy clouds spotted the clear blue sky. |
| Thunder rumbled and low grey clouds gathered over the horizon. |
| Philip unearthed a rusty yet usable knife and quickly pocketed it. |
| Philip sharpened the blade of the knife carefully. |
| M+ condition: |
| Several white fluffy meeves spotted the clear blue sky. |
| Thunder rumbled and low grey meeves gathered over the horizon. |
| Philip unearthed a rusty yet usable yepal and quickly pocketed it. |
| Philip sharpened the blade of the yepal carefully. |
| M− condition: |
| Several white fluffy meeves spotted the clear blue sky. |
| Thunder rumbled and low grey yepals gathered over the horizon. |
| Philip unearthed a rusty yet usable meeve and quickly pocketed it. |
| Philip sharpened the blade of the yepal carefully. |
Upon completion of each story, participants first completed a lexical decision task, designed to assess implicit learning of M+ words, and then completed a recognition task followed by a free recall task, both designed to assess explicit learning of M+ words. For the lexical decision task, participants were instructed to read both the prime and the target of each trial, and to respond as quickly and accurately as possible to the target by pressing a button labeled "Word" if the target was an English word, and to press a different button labeled "Nonword" if the target was not an English word. The prime was a critical word that had been presented during the preceding story (from either the R, M+ or M− condition), while the target was either a legal English word or a pseudoword formed in accordance with the orthographic and phonological rules of English. English word targets and pseudoword targets occurred with equal frequency. Half of the legal English word targets were close semantic associates of the prime (when the prime was an R or M+ word), while the other half of the targets were semantically unrelated to the prime. (Each M− prime also preceded two English word targets, though these targets could not be considered either related or unrelated to the prime). Unrelated word pairs were created by rearranging related primes and targets so that there was no semantic relationship between the two words. Word and pseudoword targets were matched on number of syllables and word length. Each critical word was presented a total of four times, once preceding a related target, once preceding an unrelated target, and twice preceding a nonword, for a total of 80 trials per block. Trial order was counterbalanced across participants, such that half were first exposed to the related pair of a particular word, while the other half first saw the same word in the unrelated condition. Each trial began with the presentation of a fixation box that stayed on the screen until 1200 ms after the onset of the final word. One thousand ms after the onset of this fixation box, the prime word was presented for 200 ms, followed by 300 ms of blank, followed by a 200 ms presentation of the target word. The next trial began 3 sec after the participant's response. Reaction time data from the lexical decision task were analyzed using a repeated-measures ANOVA with prime condition (R, M+) and target condition (related, unrelated, nonword) as factors.
For the recognition task, participants again read both a prime and target word, and were told to press a button labeled "Related" if they judged the word pairs to be semantically related or equivalent, and a button labeled "Unrelated" if they judged that the two words were not semantically related. Accuracy was emphasized over speed. The prime was a critical word from the R or M+ conditions, while the target was a word that was either semantically related or unrelated to the prime. Critical words from the M− condition were not presented, as these words had not been assigned semantic meaning during the story task. Related word pairs for the M+ condition were created by pairing each M+ word with its real-word counterpart, while unrelated word pairs were created by combining each M+ prime with the real-word counterpart of a different M+ word. For words from the R condition, related word pairs were created by pairing each R target with either a synonym or close semantic associate (e.g. bed - cot), while unrelated targets were created by matching each R prime to the synonym or close semantic associate of a different R word. Again, trial order was counterbalanced across participants, such that half of participants first saw the related pair of a particular word, while the other half were first exposed to the unrelated pair. A word was considered to be correctly recognized if performance was correct for both its related and unrelated trials. Stimulus timing was identical to that of the lexical decision task.
Finally, for the free recall task, participants were given a sheet of paper listing all the M+ and M− pseudowords that had been presented during the preceding story and told to fill in a corresponding real word for each pseudoword shown. They were encouraged to guess if necessary but told that they could leave a blank next to any word whose meaning they did not know.
ERP Recording and Analysis
EEG data were collected throughout the story tasks, lexical decision tasks, and recognition tasks. EEG was recorded at a sampling rate of 1024 Hz from 32 Ag-AgCl-tipped electrodes attached to an electrode cap using the 10/20 system. Recordings were made with the Active-Two system (Biosemi, Amsterdam, Netherlands), which does not require impedance measurements, an online reference, or gain adjustments. Additional electrodes were placed on the left and right mastoid, at the outer canthi of both eyes and below the right eye. Scalp signals were recorded relative to the Common Mode Sense (CMS) active electrode and then re-referenced off-line to the algebraic average of the left and right mastoid. Left and right horizontal eye channels were re-referenced to one another, and the vertical eye channel was re-referenced to FP1.
ERP analyses were carried out using EEGLAB (Delorme & Makeig, 2004). For each paradigm for which EEG data were collected (story task, lexical decision task, and recognition task), the four experimental blocks were merged into one data file. Data were down-sampled to 256 Hz to speed computation and band-pass filtered from 0.1 to 40 Hz. Next, epochs time-locked to critical word onset were extracted from −100 to 1000 msec. Trials containing large or paroxysmal artifacts or movement artifacts were identified by visual inspection and removed from further analysis. Data from the story task for all participants (N = 20), and from the lexical decision (N = 16) and recognition tasks (N = 6) for participants who occasionally blinked during the critical epochs, were then submitted to the extended runica routine of EEGLAB software. Ocular artifacts were identified from scalp topographies and the component time series and removed. ICA-cleaned data were subjected to a final manual artifact correction step to detect any residual or atypical ocular artifacts not removed completely with ICA. For some participants (lexical decision task N = 4; recognition task N = 14), ICA did not converge on clean ocular artifact components due to low numbers of vertical or horizontal eye movements or blinks. For these data, ocular artifacts were detected and removed manually by inspecting eye-channels for deflections and polarity inversions with scalp channels.
Based on visual inspection of the waveforms as well as on previous studies (Kutas and Hillyard, 1980, 1984; Van Petten et al., 1991), the N400 time window was selected as 300 to 500 ms poststimulus, and the late positive component (LPC) time window was selected as 500 to 900 ms poststimulus. Waveforms were quantified by measuring mean voltages within the selected latency windows, relative to a 100 msec prestimulus baseline. Repeated-measures analyses of variance (ANOVA) were conducted separately for each task. In the story task, factors were presentation (1st to 10th), condition (R, M+, M−), and three topographical factors [hemisphere (left, right), anterior/posterior (frontal, fronto-temporal, temporal, central, parietal, occipital), and lateral/medial (lateral, medial)]. Topographical factors were included in order to examine and compare the distributions of different ERP effects. In the lexical decision task, factors included prime condition (R, M+), target condition (related, unrelated, nonword), and the three topographical factors. In the recognition task, factors were prime condition (R, M+), relatedness condition (related, unrelated), and the three topographical factors. To compare the distribution of the real and M+ N400 effects in the recognition task, differences waves (computed by subtracting the related ERP from the unrelated EPP between 300–500 ms poststimulus) were normalized (McCarthy & Wood, 1985) and then subjected to a repeated-measures ANOVA that included prime condition (R, M+) and the three topographical measures as factors. To examine the effect of correct subsequent recognition on the ERP over the ten exposure periods of M+ critical words, a repeated-measures ANOVA with five factors [presentation, subsequent recognition (recognized, unrecognized), and the three topographical factors] was carried out on the story task data. A similar analysis was performed to examine the effect of subsequent recall. Data from the recognition task for M+ words was also divided by recognition performance using a repeated-measures ANOVA with five factors [subsequent recognition, relatedness condition (related, unrelated), and the three topographical factors)]. Only participants who had a sufficient number of trials in both correct and incorrect bins (≥ 5 trials) were included in these analyses (recognition N = 14; recall N = 15).2 An M+ word was considered correctly recalled when a participant provided its hidden real-word counterpart or a closely-related semantic associate that could also have reasonably fit the given context. An M+ word was defined as correctly recognized when a participant both endorsed the correct real-word counterpart and rejected a false unrelated target (chance performance = 25%). For all analyses, separate tests were run for midline electrodes (Fz, Cz, Pz, Oz, CP1, CP2) that could not be included in the main factorial topographical ANOVA. Both results from the main factorial ANOVA as well as the midline analyses are reported.; Greenhause-Geisser corrections are reported for factors with more than two levels.
Because the N400 and LPC are maximal at central midline sites (e.g., Kutas & Hillyard, 1980; Van Petten et al., 1991), Cz was chosen as a representative electrode across the ten presentations of the story task in the depicted figures (Figures 1 & 3). Representative central and parietal sites, where ERP effects were maximal, were chosen for figures illustrating data from the lexical decision and recognition tasks (Figures 4–6).
Figure 1.
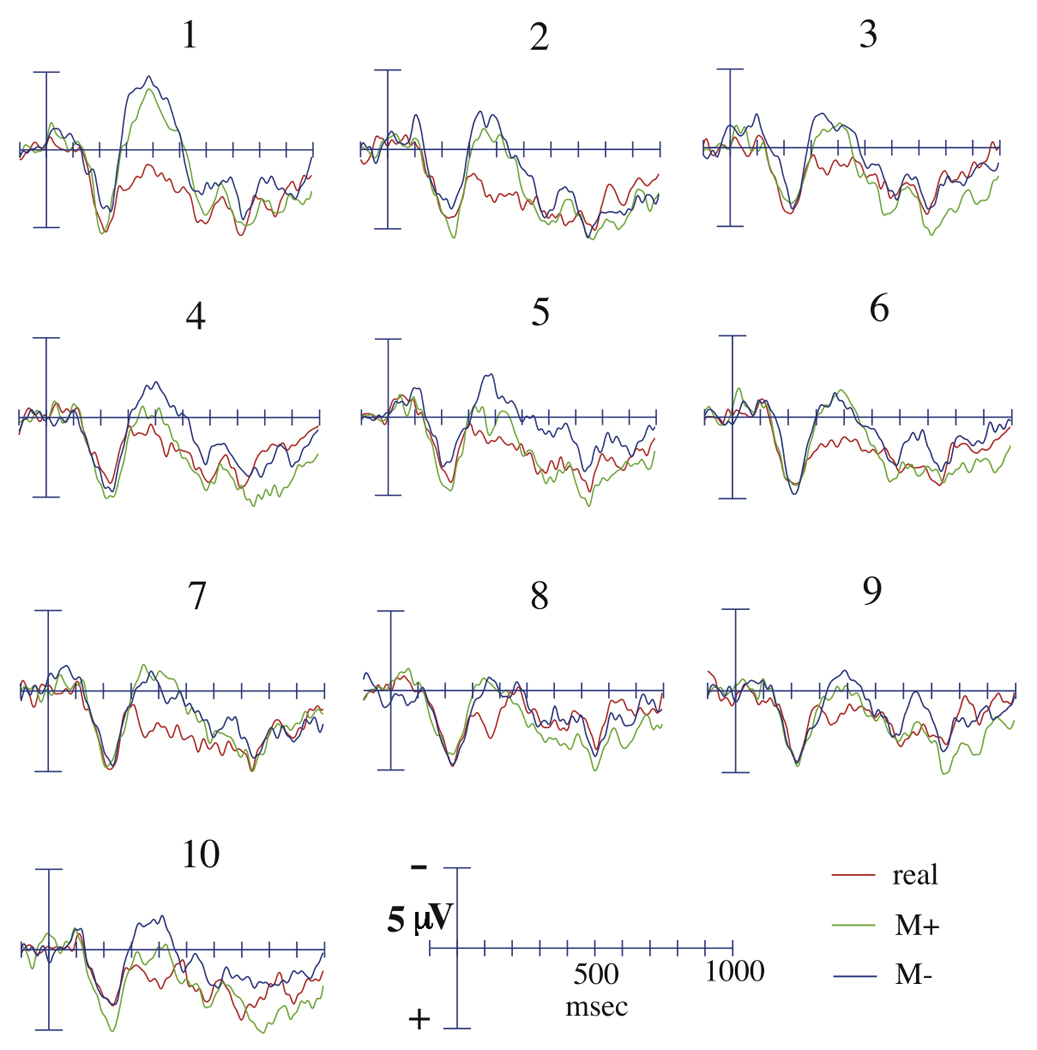
Grand average ERPs at the Cz electrode to real, M+, and M− critical words, divided across the ten presentations in the story task.
Figure 3.
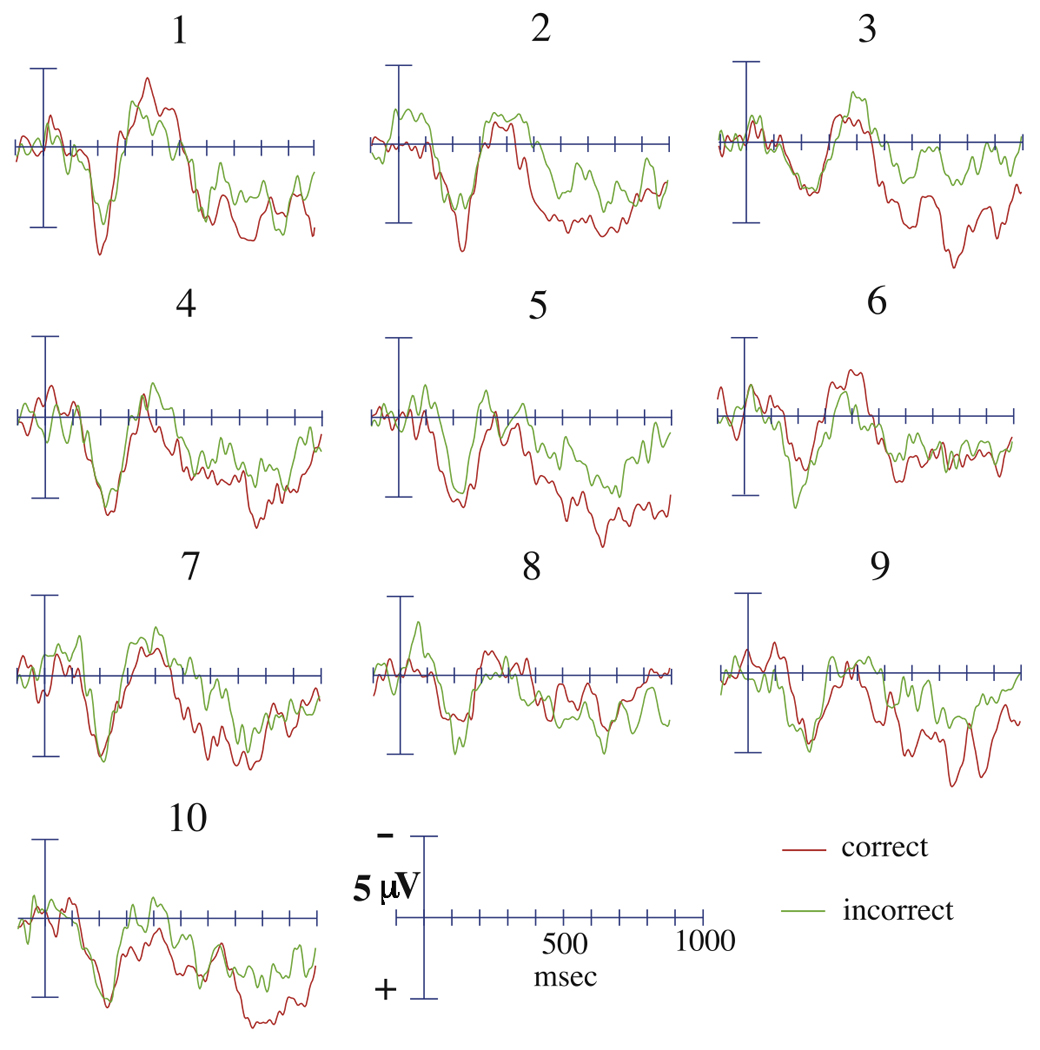
Grand average ERPs to M+ critical words at the Cz electrode across the ten presentations of the story task, divided as a function of subsequent recall.
Figure 4.
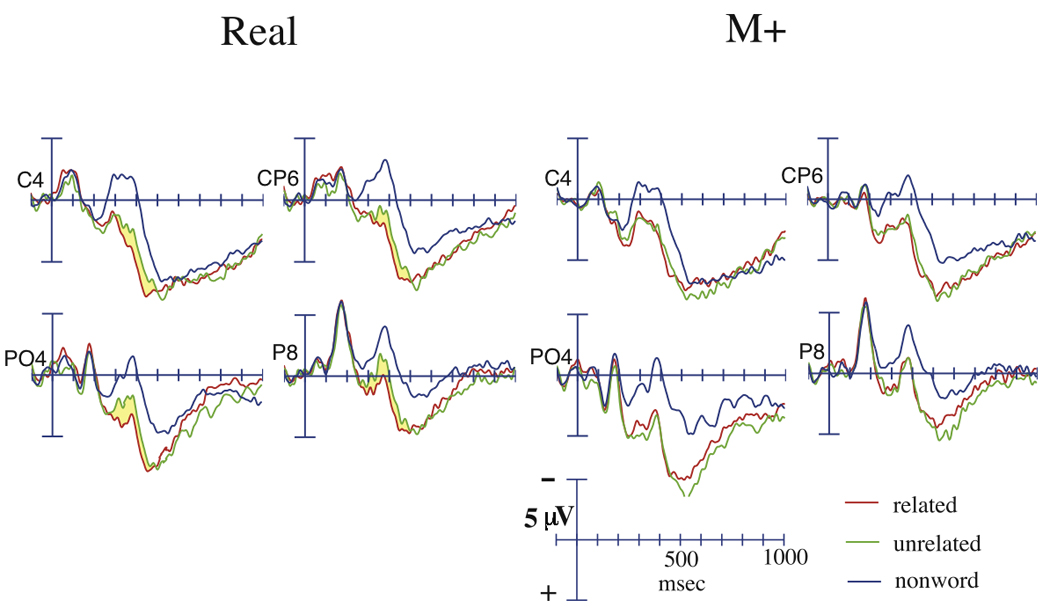
Grand average ERPs to targets in the lexical decision task, at representative right-hemisphere central and parietal sites. Targets were either nonwords or were real English words preceded by either semantically related or unrelated primes. Targets preceded by real English primes are shown on the left, and targets preceded by M+ primes are shown on the right.
Figure 6.
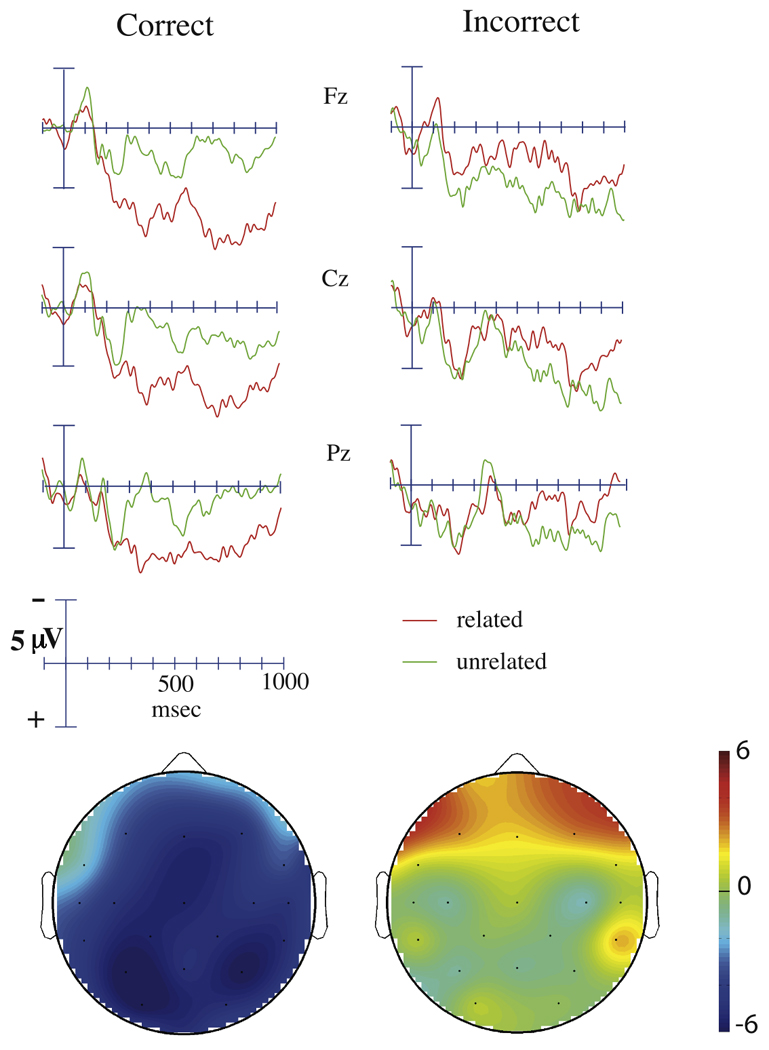
Grand average ERPs and topographical voltage maps to targets preceded by M+ word primes in the recognition task, divided as a function of subsequent recognition. Targets preceded by M+ primes that were subsequently recognized (in both related and unrelated trials) are shown on the left. Targets preceded by M+ primes that were not subsequently unrecognized in both trials appear on the right. The effects shown in the voltage maps were computed by subtracting the ERP to unrelated targets from the ERP to related targets between 300 to 500 ms post-stimulus. The scale for each effect is identical.
Results
Behavioral Results
Average accuracy to multiple-choice comprehension questions during the story task was 94.3 % (SD = 7.4%).
Analysis of median reaction times in the lexical decision task revealed that participants responded significantly more quickly to real word targets than to nonword targets (F(2,38) = 13.9, p < 0.001; Word > Nonword contrast: F(1,19) = 17.3, p = 0.001). A marginally significant effect of semantic relatedness on reaction time was found (Related > Unrelated contrast: F(1,19) = 3.09, p = 0.095). Follow-up analyses indicated that semantic relatedness tended to speed reaction times in the R condition (F(1,19) = 3.46, p = 0.078; Related Targets: Median = 565.5 ms, SD = 97.9; Unrelated Targets: Median = 596.6 ms, SD = 145.1 ms), but not in the M+ condition (F(1,19) = 0.31, p = 0.58; Related Targets: Median = 592.5 ms, SD = 122.2; Unrelated Targets: 598.9 ms, SD = 134.3 ms). Average recognition accuracy of the meanings of M+ words was 72.4% (SD = 19.7%), while average recall accuracy was 63.8% (SD = 23.8%).
ERP Results
Story task
N400 time-window, all trials
During the 300 to 500 msec time-window, the negativity to all words decreased as a function of repetition (main analysis: F(9, 171) = 4.94, p < 0.001; midline sites: F(9, 171) = 5.39, p < 0.001). The change in ERP response over the ten presentations to R words, M+ words, and M− words differed significantly (main analysis: (F(18, 342) = 2.32, p = 0.027; midline sites: F(18, 342) = 2.31, p = 0.002; Figures 1 and 2). Contrasts revealed that the N400 elicited by M+ and M− words decreased linearly over the ten presentations relative to the N400 elicited by R words (main analysis: F(1,19) = 14.16, p = 0.001; midline sites: (F(1,19) = 16.16, p = 0.001), and that the N400 to M+ words decreased cubically relative to the N400 to M− words (main analysis: F(1,19) = 5.32, p = 0.033; midline sites: (F(1,19) = 2.98, p = 0.10). The N400 to R words did not differ as a function of presentation (main analysis: F(9, 171) = 1.05, p = 0.40; midline sites: F(9, 171) = 1.30, p = 0.27). A main effect of condition was revealed across all presentations and topographical factors (main analysis: F(2, 38) = 29.12, p < 0.001; midline sites: F(2, 38) = 20.55, p < 0.001). Specifically, R words elicited a significantly reduced N400 relative to the novel pseudowords (main analysis: F(1) = 37.21, p < 0.001; midline sites: F(1) = 24.25, p < 0.001), and M+ words elicited a significantly reduced N400 relative to M− words (main analysis: F(1) = 7.33, p = 0.014; midline sites: F(1) = 8.31, p = 0.010). Follow-up analyses revealed that there was no significant difference in the ERP evoked by M+ and M− words at the first presentation (main analysis: F(1,19) = 1.06, p = 0.32; midline sites: F(1, 19) = 1.64, p = 0.22), but that by the second presentation M− words evoked a marginally larger N400 relative to M+ words (main analysis: F(1,19) = 2.93, p = 0.10; Condition × Hemisphere × Laterality: F(1,19) = 6.26, p = 0.022), fully significant over left lateral sites (F(1, 19 = 4.88, p = 0.040), though not over midline sites (F(1,19) = 2.33, p = 0.14). This condition difference persisted across the final nine presentations over all electrodes (Condition effect, collapsed across final nine presentations: main analysis: F(1,19) = 7.67, p = 0.012; midline sites F(1,19) = 7.82, p = 0.012).
Figure 2.
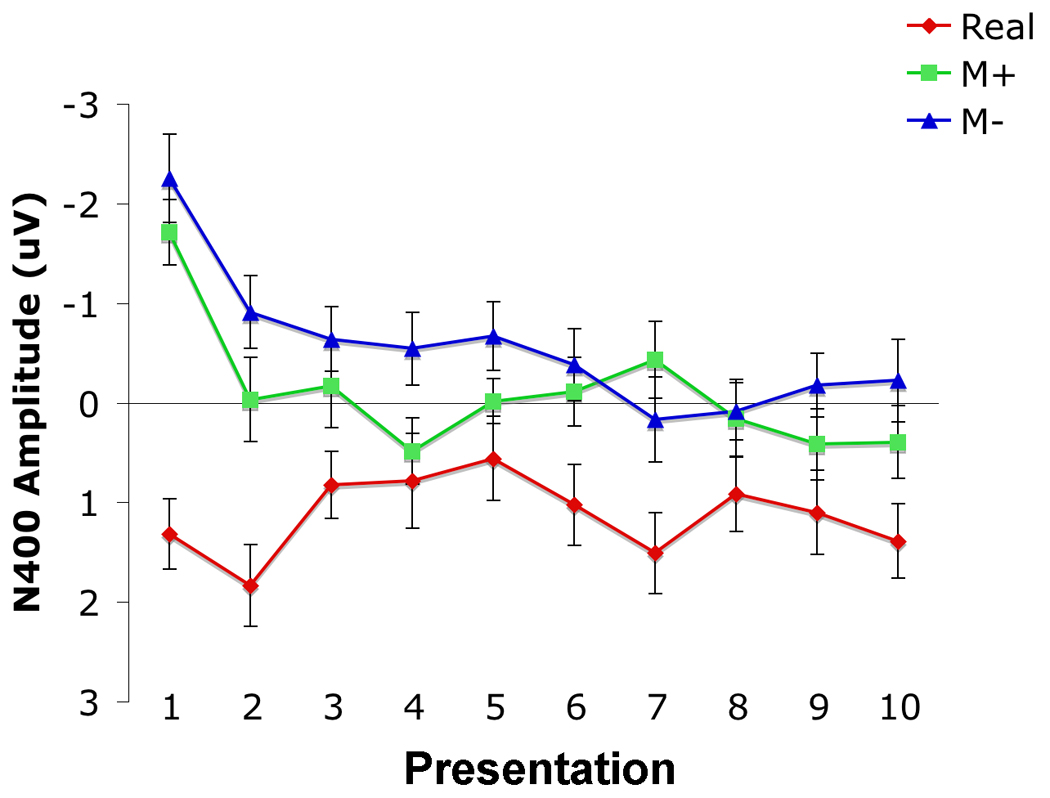
Mean amplitude plots of the N400 response across all electrodes included in the main factorial ANOVA to real, M+ and M− critical words in the story task, as a function of presentation. Mean amplitude measurements were computed from 300 to 500 msec post-stimulus. Negative is plotted upward. Error bars indicate standard errors of the mean.
N400 time-window, divided by subsequent M+ recognition and recall performance
When M+ critical trials were divided by recognition performance, no main effect of recognition was revealed (main analysis: F(1,13) = 0.84, p = 0.38; midline sites: F(1,13) = 1.65, p = 0.22). Similarly, when M+ critical trials were divided by recall performance, no main effect of recall was found (main analysis: F(1,14) = 1.70, p = 0.21; midline sites: F(1,14) = 1.66, p = 0.22; Figure 3). Neither recognition nor recall performance interacted with any factorial factor (all p values > 0.3).
LPC time-window, all trials
During the later 500 to 900 msec time-window, a main effect of condition was revealed (main analysis: F(2, 38) = 4.40, p = 0.035; midline sites: F(2, 38) = 4.12, p = 0.044; Figure 1). While R words did not elicit a significantly different LPC compared to M+ and M− words (main analysis: F(1,19) = 2.30, p = 0.15; midline sites: F(1,19) = 2.38, p = 0.14), the LPC elicited by M+ words was significantly greater than that elicited by M− words (main analysis: F(1,19) = 12.80, p = 0.002; midline sites: F(1,19) = 12.1, p = 0.002), or by R words (main analysis: F(1,19) = 7.64, p = 0.012; midline sites: F(1,19) = 7.43, p = 0.013). Hypothesis-driven follow-up analyses, designed to examine the time-course over which these condition differences emerged, indicated that there was no significant difference in the ERP evoked by M+ and M− words at the first presentation (main analysis: F(1,19) = 1.47, p = 0.24; midline sites: F(1,19) = 1.27, p = 0.27) or at the second presentation (main analysis: F(1,19) = 0.79, p = 0.39; midline sites: F(1,19) = 0.99, p = 0.33), but that a condition difference emerged by the third presentation (main analysis: F(1,19) = 5.29, p = 0.033; midline sites: F(1,19) = 3.29, p = 0.086) and persisted across the final eight presentations (Condition effect, collapsed across final eight presentations: main analysis: F(1, 19) = 12.24, p = 0.002; midline sites: F(1, 19) = 11.33, p = 0.003). The LPC enhancement was maximal over posterior and medial sites (Condition × Anterior/Posterior: F(10,190) = 27.1, p < 0.001; Condition × Laterality: F(2,28) = 4.77, p = 0.026; Condition × Anterior/Posterior × Laterality: F(10,190) = 4.66, p < 0.001).
LPC time-window, divided by subsequent M+ recognition and recall performance
During the 500 to 900 msec time-window, no main effect of recognition on M+ trials was found (main analysis: F(1,13) = 1.04, p = 0.33; midline sites: F(1,13) = 2.45, p = 0.14). Recognition did not interact with any distributional factor (all p values > 0.2). In contrast, when M+ trials were divided by subsequent recall performance, a highly significant main effect of recall performance was revealed (main analysis: F(1,14) = 29.81, p < 0.001; midline sites: F(1,14) = 30.4, p < 0.001; Figure 3), such that M+ words whose meanings were correctly recalled at test showed a significantly more positive ERP. This effect was largest over posterior sites (Subsequent Recall × Anterior/Posterior: F(5, 70) = 5.16, p = 0.004). Follow-up analyses designed to ensure that this effect could not be attributed to baseline differences in the word forms between conditions confirmed that there were no differences in the ERP evoked by correctly-recalled versus incorrectly-recalled M+ words at the initial presentation (main analysis: F(1,14) = 0.31, p = 0.56; midline sites: F(1,14) = 0.45, p = 0.51).
Lexical Decision Task
During the 300 to 500 msec time-window, across both the R and M+ prime condition, a main effect of target condition was revealed, indicating that nonword targets elicited a significantly larger N400 than word targets (main analysis: F(2, 38) = 37.5, p < 0.001; midline sites: F(2, 38) = 33.1, p < 0.001; Contrasts: main analysis: F(1,19) = 45.1, p < 0.001; midline sites: F(1,19) = 40.5, p < 0.001; Figure 4). Unrelated targets elicited a significantly larger N400 than related targets at temporal, central, and parietal rows (Target Condition × Quadratic Anterior/Posterior contrast: F(1,19) = 4.80, p = 0.041; Follow-up Analysis: F(1,19) = 4.48, p = 0.048), and a marginally larger N400 at midline sites (F(1,19) = 3.71, p = 0.069). To investigate whether targets preceded by both R and M+ primes showed this N400 effect, separate analyses were run for each prime condition. Targets preceded by R primes showed a nearly significant N400 effect across all electrodes (main analysis: F(1,19) = 4.26, p = 0.053; midline sites: F(1,19) = 3.50, p = 0.077), while targets preceded by M+ primes did not show a significant effect (main analysis: F(1,19) = 0.005, p = 0.94; midline sites: F(1,19) = 0.30, p = 0.59).
Recognition Task
All trials
Across both prime conditions (R and M+), unrelated targets elicited a larger N400 than related targets (main analysis: F(1,19) = 20.8, p < 0.001; midline sites: F(1,19) = 33.7, p < 0.001; Figure 5), an effect which was maximal over posterior, medial, and right hemisphere sites (Relatedness × Anterior/Posterior: F(5, 95) = 15.06, p < 0.001; Relatedness × Laterality: F(1,19) = 10.80, p = 0.004; Relatedness × Hemisphere × Laterality: F(1,19) = 7.95, p = 0.011; Relatedness × Anterior/Posterior × Laterality: F(5,95) = 6.23, p < 0.001). No main effect of N400 amplitude across prime condition was found (main analysis: F(1, 19) = 0.902, p = 0.35; midline sites: F(1,19) = 0.314, p = 0.58), and topographical analyses of normalized data revealed no significant differences in the distribution of these effects (all p values > 0.2; Figure 5). To confirm that the N400 effect was present in both prime conditions, independent analyses were run for each prime condition, confirming that both R trials (main analysis: F(1,19) = 23.6, p < 0.001; midline sites: F(1,19) = 29.3, p < 0.001) as well as M+ trials (main analysis: F(1, 19) = 9.00, p = 0.007; midline sites: F(1,19) = 18.4, p < 0.001) showed significant N400 effects.
Figure 5.
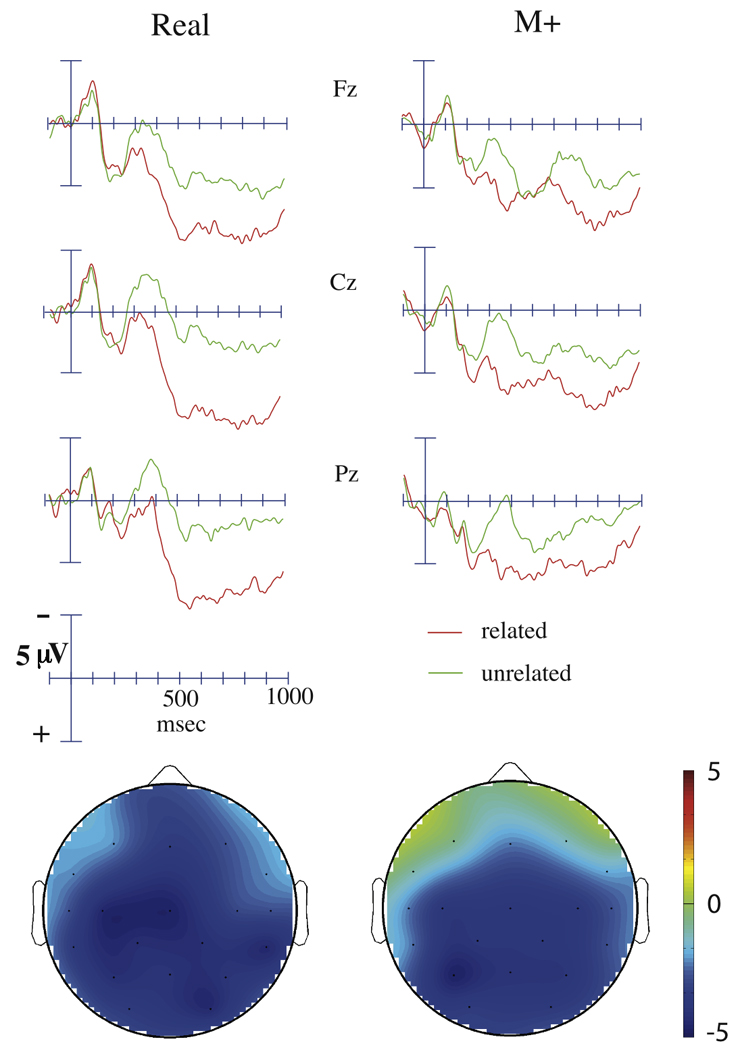
Grand average ERPs to targets, depicted at midline sites, and topographical voltage maps in the recognition task. Targets were real English words preceded by either semantically related or unrelated primes. Targets preceded by real English primes are shown at the left, while targets preceded by M+ primes are shown on the right. The effects depicted in the voltage maps were computed by subtracting the ERP to unrelated targets from the ERP to related targets between 300 to 500 ms post-stimulus. The scale for each of these effects is identical.
M+ trials divided by performance
The N400 effect elicited by targets preceded by M+ primes whose meanings were subsequently recognized was significantly larger than the N400 effect elicited by targets preceded by primes whose meanings were subsequently unrecognized (main analysis: F(1, 13) = 7.14, p = 0.019; midline sites: F(1,13) = 9.95, p = 0.008; Figure 6). Follow-up analyses indicated that correctly-recognized M+ primes elicited a significant N400 effect (main analysis: F(1, 13) = 26.0, p < 0.001; midline sites: F(1,13) = 23.9, p < 0.001) that was largest over medial and posterior sites (Relatedness × Anterior/Posterior: F(5, 65) = 4.45, p = 0.023; Relatedness × Laterality: F(1, 13) = 5.91, p = 0.030; Relatedness × Anterior/Posterior × Laterality: F(5, 65) = 4.98, p = 0.006; Figure 6). In contrast, subsequently unrecognized M+ primes did not elicit a significant N400 effect at any scalp site (main analysis: F(1,13) = 0.24, p = 0.63; midline sites: F(1,13) = 0.006, p = 0.94).
Discussion
Story Task
During the story task, consistent with our hypotheses, M+ words showed a significant reduction in the N400 across the ten presentations. Although M− words also showed a reduction in the N400, likely due to the effects of repetition (e.g., Besson, Kutas, & Van Petten, 1992; Mitchell, Andrews, & Ward, 1993; Van Petten, Kutas, Kluender, Mitchiner, & McIsaac, 1991), M+ words showed a decrease beyond that observed to M− words, indicating that some proportion of this effect can be attributed to semantic integration due to meaning acquisition. This effect emerged rapidly, with amplitude differences appearing between the two conditions as early as the second presentation, converging with previous work that suggests that at least some aspects of meaning acquisition occur with remarkable speed (Mestres-Misse, et al., 2007). In sum, the reduction in the N400 effect over time represents an electrophysiological index of incidental meaning acquisition as occurs during natural language processing.
Although M+ words elicited a reduced N400 compared to M− words overall, the differences in N400 amplitude between the two conditions followed a cubic function, rather than a linear one. Visual inspection of mean amplitudes in the 300–500 msec latency window over the ten presentations further demonstrates that the expected effect was greatly reduced and even reversed over the sixth through the eight presentation (Figure 2). One possible explanation for this apparent anomaly is that meaning acquisition of the novel words interacted with repetition over the ten presentations. Word repetition is thought to exert its effects by increasing the baseline activation level of primed representations, leading to a reduction in the N400 even when words are semantically incongruent with the preceding context. This effect appears to be additive; previous research has indicated that N400 amplitude is driven lower by multiple repetitions compared to a single repetition, and that congruency effects may be quickly masked by repetition effects (Besson, et al., 1992; Van Petten, et al., 1991). After five presentations in our task, baseline activation of the novel pseudowords may have increased to such a level that increased facilitation of semantic processing due to M+ word learning exerted no additional effect on the ERP. In the final two presentations, when participants' representations of the M+ word were presumably the most developed, the difference in N400 response between M+ and M− words began to re-emerge. This finding suggests that after ten exposures, despite powerful repetition effects, differences in conceptual representations between M+ and M− words were sufficiently robust to produce a difference in N400 amplitude.
Word condition effects were also found in the later time-window (500–900 msec), with M+ words eliciting a significantly larger LPC component compared to both M− words and real words (Fig. 1). An enhanced late positivity is often elicited by word repetition (e.g., Bentin & Peled, 1990; Besson, et al., 1992; Van Petten, et al., 1991) and has been proposed to reflect the extended retrieval of semantic and episodic information from long-term memory and the integration of that information into working memory (Van Petten, et al., 1991). Thus, one reasonable explanation for our finding is that the enhanced LPC component evoked by M+ words reflects participants' attempts to retrieve prior instances of each word and link them to the present context. This process is not necessary for real words, whose meanings have already been firmly established in semantic memory; prior instances of these words in the preceding story context do not need to be retrieved in order for participants to understand their meanings. Similarly, retrieving previous presentations of a M− word serves no useful purpose in contributing to comprehension of its meaning in the current phrase. Because M− words occurred in inconsistent contexts, no comprehensive representations of meaning are available for retrieval and integration into the present framework. Our finding that LPC differences between M+ and M− words did not emerge until the third presentation, the first point at which more than one presentation of a given word had previously occurred, supports this interpretation. While for an M+ word both prior instances of the word would contribute to the development of a cohesive meaning representation and might be retrieved to aid in current comprehension processes, for a M− word the two previous occurrences would conflict with one another and likely could not be retrieved as a coherent representation. Thus the late enhanced positivity to M+ words is likely an index of extensive encoding, integration, and recall processes that are key in the development of stable, long-term meaning representations.
The N400 and the LPC show different effects as a function of subsequent memory. The N400 to M+ words was not significantly impacted by either subsequent recognition or recall of these words, suggesting that there is no reliable link between the processes reflected by the N400 during learning and those that support later memory. This finding is supported by prior studies that found no relationship between N400 amplitude and subsequent recognition performance (Besson, et al., 1992; Neville, Kutas, Chesney, & Schmidt, 1986; Rugg & Doyle, 1992). In contrast, the amplitude of the LPC to M+ words in our study was highly predictive of subsequent recall performance, consistent with a number of previous findings (e.g., Neville, et al., 1986; Paller, 1990; Paller, Kutas, & Mayes, 1987; Paller, McCarthy, & Wood, 1988; Schott, et al., 2002; Roeder, Rosler, and Neville, 2001). This positivity at encoding for subsequently-remembered items, commonly termed the "difference due to later memory effect" (Dm effect), has been proposed to index elaborative encoding and consolidation processes engaged for the formation of distinctive memory traces, processes that are related to those indexed by the LPC (Besson, et al., 1992; Mitchell, et al., 1993; Van Petten, et al., 1991). Our finding that LPC amplitude did not reliably predict recognition performance converges with previous demonstrations of larger Dm amplitude differences for free recall compared to recognition and cued recall (Paller, 1988; 1990). These data provide further support for the idea that the Dm effect reflects encoding strength of words due to explicit elaborative processes (Paller et al, 1988). Cues given in recognition tests aid retrieval and make recognition less dependent upon elaborative processes, so that sorting trials on the basis of recall rather than recognition may be more sensitive to differences in encoding strength. The present study replicates and extends these findings, demonstrating that the processes indexed by the Dm are at play not only in the processing of isolated words in lists, but also in more natural language processing tasks.
In summary, the ERPs elicited by M+ words over multiple presentations suggests that two distinct processes support the development and encoding of novel meaning representations. In the first time-window, a reduction in the N400 to these words across multiple representations is indicative of increasing facilitation in semantic expectancy and semantic integration processes over time. The modulation in this online measure of language processing indicates that newly acquired semantic representations can be retrieved and processed within the same general time window as existing words. This effect appears to index processes that are independent of those that support subsequent memory. During the later time-window, an enhanced positivity (LPC) to M+ words appears to reflect extended processing that follows the completion of semantic analysis, in which prior instances of a given word are retrieved from long-term memory and integrated into the present context. These processes likely play a critical role in the development of comprehensive semantic representations of novel words, as well as in the encoding of these representations into long-term memory. This interpretation is further supported by the finding that the amplitude of the late positivity elicited by these words predicts subsequent recall. Thus, our data indicate that both language-related processes of semantic integration, indexed by the N400, and memory-related processes of encoding, elaboration, and retrieval, indexed by the LPC and the Dm effects, contribute to the development and maintenance of new meaning representations. The present study extends previous work on the functional significance of these ERP components by using a language-processing task that is more natural than those that have been previously employed to investigate these questions.
Lexical Decision and Recognition Tasks
In the lexical decision task, real word primes tended to speed behavioral responses to semantically related targets relative to unrelated targets, while this semantic priming effect was not observed to targets preceded by M+ words. Electrophysiologically, a similar pattern was revealed; targets preceded by real word primes showed a nearly significant N400 reduction across all electrodes, while targets preceded by related M+ primes elicited no trace of an N400 reduction. In contrast, in the recognition task, targets preceded by both real and M+ primes exhibited highly significant N400 effects with similar distributions. Behavioral recognition accuracy for the meanings of M+ words was reasonably high (mean = 72.4%), indicating that a moderate degree of explicit word learning took place. Thus, M+ words showed neither behavioral nor ERP evidence of implicit memory, but demonstrated behavioral and ERP effects of explicit memory.
Although null effects must be carefully interpreted, both behavioral and ERP data suggest that while explicit representations of M+ words developed rapidly, implicit meaning representations had not yet formed and were not available for processing under the present experimental conditions. Behaviorally, no semantic priming effect emerged, despite the fact that participants performed quite accurately in the explicit recognition task and had clearly learned the meanings of most of the words. At the neural level, the N400 was elicited only under conditions of intentional recollection, when explicit memory traces for M+ words were called upon. In the lexical decision task, in which response speed to related targets should have been facilitated by implicit memory for M+ word meanings while explicit memory for these words did not need to be summoned, no N400 effect was observed. We had hypothesized that adopting a natural and incidental language processing task could potentially minimize the role of explicit encoding strategies while simultaneously facilitating the operation of implicit learning processes. However, it appears that implicit memory for the meanings of novel words develops only after more extensive exposure and/or a longer period of incubation than used in the present study, regardless of the nature of the task. Even under more incidental learning conditions, explicit memory systems still appears to play a dominant role in initial vocabulary acquisition.
Although we did not directly test whether implicit representations in our participants developed after a period of incubation, our results are consistent with the Davis and Gaskell's (2009) proposal that word learning occurs in two stages: an initial stage of rapid familiarization, followed by a second stage of slower lexical consolidation. With further experimentation, this two-stage model may be extended to apply not only to the acquisition of word form, but also to semantic learning. The present findings are also consistent with a small number of behavioral word learning studies whose data shed light on development of implicit and explicit knowledge of novel words, reviewed previously in the introduction. For example, Clay et al. (2007) demonstrated that while most participants achieved an accuracy level of at least 85% in the second half of the study phase, showing high accuracy in explicit judgments of whether new word matched corresponding descriptions or pictures, an automatic interference effect emerged only one week after initial training. Similarly, participants in Dumay et al.'s (2004) study generated the meanings of 30% of novel words in a free association task administered on the day of training, demonstrating a reasonable amount of explicit knowledge, but showed semantic priming effects on a lexical decision task only after a one-week delay. These data suggest that implicit priming effects in our participants might have emerged several days after the initial experimental session, though future research will be necessary to confirm this hypothesis.
The present paradigm was limited by the necessity of revealing to participants, once they had read the first narrative, that their explicit memory for the novel words would be subsequently tested. Further, informal post-session interviews with participants indicated that many of them engaged in intentional strategies to infer and remember the meanings of M+ words. One possibility, then, is that participants learned the first set of novel words incidentally, but learned the last three sets of words intentionally after realizing that they would later be asked about the words' meanings. To investigate this issue, we compared the first block of data to each of the last three blocks. However, we found no evidence of either behavioral or electrophysiological differences between the first story block and any of the last three, suggesting that participants performed the tasks similarly even after becoming aware that they would be tested on the novel words. Most compellingly, in terms of recognition and recall, participants scored neither better nor worse in the first block compared to the following three. Therefore, the use of any intentional strategies does not appear to have been driven by participants' awareness that their recognition and recall memory would later be tested. Rather, it appears that explicit memory plays an important role in semantic word learning, even in a relatively natural language acquisition environment.
Consistent with the idea that M+ word learning was initially mediated by explicit memory systems, dividing M+ trials as a function of recognition performance in the recognition task revealed that only correctly recognized M+ words elicited an N400 effect. Targets preceded by M+ words whose meanings were incorrectly identified evoked no N400 effect. These results shed light on the functional significance of the N400, indicating that this component is an index of semantic integration processes that are dependent upon explicit knowledge of a word's meaning. This conclusion converges with a recent study from our lab, in which we demonstrated that words occurring within the attentional blink period elicit an N400 effect only if they can be correctly reported (Batterink, Karns, Yamada, & Neville, in press). The present study is also consistent with another recent study that investigated whether an N400 is elicited by semantic anomalies that are difficult to detect, such as the one that occurs in the phrase "after an air crash, where should the survivors be buried?" (Sanford, Leuthold, Bohan, & Sanford, in press). The authors found that difficult anomalies did not produce an N400 effect, while easily detectable control anomalies did, suggesting that the N400 is not an index of unconscious recognition but rather reflects processes that are dependent upon online awareness. A final study with which our results corroborate demonstrated that only well-formed and constrained representations of novel words evoked the N400 (Borovsky, Elman, & Kutas, 2008). These researchers found that novel words reduced the N400 amplitude to plausible verbs presented in subsequent test sentences only if the novel word had appeared in a highly constraining context during initial learning. This finding suggests that a relatively precise representation of a word's meaning is necessary to modulate the N400 effect. In the present study, participants may have acquired the general "gist" of a subset of M+ words--learning, for example, that a given word was a concrete noun with certain semantic features--without being able to correctly identify the word in the recognition task. Consistent with Borovsky et al.'s result, this type of vague representation was not found to be sufficient to support the facilitation of semantic integration processes indexed by the N400.
Our finding that the N400 is dependent upon explicit awareness of a word's meaning stands in contrast to results from McLaughlin and colleagues' (2004) study, in which beginning French learners showed an N400 effect to French pseudowords despite performing at chance levels in an explicit lexical decision task. These inconsistent results may be better understood when framed in the ongoing debate between the lexical access view and the semantic integration view of the N400 (e.g., Lau, Phillips & Poeppel, 2008). The lexical view proposes that the N400 reflects facilitated activation of a lexical item's representation in long-term memory. Lexical access can proceed without awareness, as shown by masked semantic priming studies (Grossi, 2006; Kiefer, 2002; Deacon, Hewitt, Yang & Nagata, 2000), and thus may not depend upon the retrieval of an explicit representation of a word's meaning. In contrast, the integration view posits that the N400 indexes the semantic integration of an incoming word with the current context. This process depends upon awareness (Brown & Hagoort, 1993), and presumably relies upon explicit knowledge of the word's meaning. While these two accounts are often pitted against each other, a more comprehensive account might concede that there are two N400 subtypes, one reflecting lexical access and another indexing semantic integration. The enhanced N400 found to pseudowords relative to real words may primarily reflect increased difficulty with lexical access, while the N400 effect to unrelated versus related words may predominantly reflect differences in semantic integration processes. This hypothesis predicts that the N400 lexicality effect may be observed in the absence of explicit knowledge of a word's meaning whereas the N400 relatedness effect depends upon an explicit representation of a word's meaning. This prediction is consistent with McLaughlin et al.'s findings. In that study, an N400 to French pseudowords was elicited even when d' for the lexical decision task was at zero; in contrast, N400 differences between related and unrelated word targets in that study did not emerge until participants exhibited improved overt knowledge of French words (d' = 0.5). One previous study that directly compared the distribution of these two N400 effects found that the lexicality N400 was significantly more frontal relative to the centroparietally distributed relatedness N400 (Chwilla, Brown, & Hagoort, 1995). This finding provides further evidence that these N400 subtypes may be indexing non-identical neural processes.
In summary, data from the lexical decision and recognition tasks suggest that explicit representation of novel words learned from a natural language context can develop with remarkable speed, eliciting a robust N400 effect after only ten exposures. These effects also showed very similar distributions, suggesting that the neural substrates underlying the processing of existing words are very quickly recruited to process newly acquired words. In contrast, no behavioral or ERP evidence of implicit knowledge of new words was revealed, suggesting that implicit representations may develop much more slowly, perhaps requiring more extensive exposure and/or a longer incubation period than used in the present study. Lastly, our finding that only correctly recognized M+ words elicit an N400 effect suggests that this component is an index of semantic integration processes that are dependent upon explicit knowledge of a word's meaning.
Footnotes
Data from one behavioral study, in which participants were exposed to novel words either in the morning or the evening and then tested 12 hours later, suggest that this process of consolidation depends critically on sleep, rather than simply the passage of time (Dumay & Gaskell, 2007).
This criterion was based on signal-to-noise of the individual averages as well as of the grand averages. Grand averages using a more stringent criterion were first constructed, using only participants with 9 or more trials in each bin (recognition N = 10, recall N = 11). However, individual averages of four borderline participants who had a more uneven distribution of trials in correct and incorrect bins were reasonably clean, and including these participants improved the signal-to-noise ratio of the grand averages. A minimum of five trials per subject has been used previously as a cutoff value in N400 studies (Rolke, Heil, Streb, and Henninghausen, 2001).
References
- Batterink L, Karns CM, Yamada Y, Neville HJ. The Role of Awareness in Semantic and Syntactic Processing: An ERP Attentional Blink Study. Journal of cognitive neuroscience. doi: 10.1162/jocn.2009.21361. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bentin S, Peled B-S. The contribution of task-related factors to ERP repetition effects at short and long lags. Memory & Cognition. 1990;18(4):359–366. doi: 10.3758/bf03197125. [DOI] [PubMed] [Google Scholar]
- Bentin S, Mouchetant-Rostaing Y, Giard MH, Echallier JF, Pernier J. ERP manifestations of processing printed words at different psycholinguistic levels: time course and scalp distribution. Journal of Cognitive Neuroscience. 1999;11:235–260. doi: 10.1162/089892999563373. 1999. [DOI] [PubMed] [Google Scholar]
- Borovsky A, Elman J, Kutas M. Getting the Gist is not Enough: An ERP Investigation of Word Learning from Context; Paper presented at the Proceedings of the Cognitive Science Society.2008. [Google Scholar]
- Breitenstein C, Jansen A, Deppe M, Foerster A-F, Sommer J, Wolbers T, et al. Hippocampus activity differentiates good from poor learners of a novel lexicon. NeuroImage. 2005;25(3):958–968. doi: 10.1016/j.neuroimage.2004.12.019. [DOI] [PubMed] [Google Scholar]
- Breitenstein C, Zwitserlood P, de Vries M, Feldhues C, Knecht S, Dobel C. Five days versus a lifetime: Intense associative vocabulary training generates lexically integrated words. Restorative neurology and neuroscience. 2007;25(5–6):493–500. [PubMed] [Google Scholar]
- Brown C, Hagoort P. The processing nature of the N400: Evidence from masked priming. Journal of Cognitive Neuroscience. 1993;5:34–44. doi: 10.1162/jocn.1993.5.1.34. [DOI] [PubMed] [Google Scholar]
- Chwilla DJ, Brown CM, Hagoort P. The N400 as a function of the level of processing. Psychophysiology. 1995;32(3):274–285. doi: 10.1111/j.1469-8986.1995.tb02956.x. [DOI] [PubMed] [Google Scholar]
- Clay F, Bowers JS, Davis CJ, Hanley DA. Teaching adults new words: The role of practice and consolidation. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2007;33(5):970–976. doi: 10.1037/0278-7393.33.5.970. [DOI] [PubMed] [Google Scholar]
- Cohen NJ, Squire LR. Preserved learning and retention of pattern-analyzing skill in amnesia: Dissociation of knowing how and knowing that. Science. 1980;210(4466):207. doi: 10.1126/science.7414331. [DOI] [PubMed] [Google Scholar]
- Delorme A, Makeig S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics. Journal of Neuroscience Methods. 2004;134:9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Davis M, Di Betta A, Macdonald M, Gaskell M. Learning and Consolidation of Novel Spoken Words. Journal of cognitive neuroscience. 2008;21(4):803–820. doi: 10.1162/jocn.2009.21059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis M, Gaskell M. A complementary systems account of word learning: neural and behavioural evidence. Philosophical transactions - Royal Society. Biological sciences. 2009;364(1536):3773–3800. doi: 10.1098/rstb.2009.0111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deacon D, Hewitt S, Yang C, Nagata M. Event-related potential indices of semantic priming using masked and unmasked words: Evidence that the N400 does not reflect a post-lexical process. Cognitive Brain Research. 2000;9:137–146. doi: 10.1016/s0926-6410(99)00050-6. [DOI] [PubMed] [Google Scholar]
- Dumay N, Gaskell MG. Sleep-Associated Changes in the Mental Representation of Spoken Words. Psychological Science. 2007;18(1):35–39. doi: 10.1111/j.1467-9280.2007.01845.x. [DOI] [PubMed] [Google Scholar]
- Dumay N, Gaskell MG, Feng X. A Day in the Life of a Spoken Word. In: Forbus K, Gentner D, Regier T, editors. Proceedings of the Twenty-Sixth Annual Conference of the Cognitive Science Society. Mahwah, NJ: Erlbaum; 2004. pp. 339–344. [Google Scholar]
- Ellis NC. Consciousness in second language learning: Psychological perspectives on the role of conscious processes in vocabulary acquisition. AILA Review. 1994;11:37–56. [Google Scholar]
- Friederici A, Pfeifer E, Hahne A. Event-related brain potentials during natural speech processing: Effects of semantic, morphological and syntactic violations. Cognitive Brain Research. 1993;1(3):183–192. doi: 10.1016/0926-6410(93)90026-2. [DOI] [PubMed] [Google Scholar]
- Gabrieli JDE, Keane MM, Zarella MM, Poldrack RA. Preservation of implicit memory for new associations in global amnesia. Psychological Science. 1997;8(4):326–329. [Google Scholar]
- Goshen-Gottstein Y, Moscovitch M, Melo B. Intact implicit memory for newly formed verbal associations in amnesic patients following single study trials. Neuropsychology. 2000;14(4):570–578. doi: 10.1037//0894-4105.14.4.570. [DOI] [PubMed] [Google Scholar]
- Graf Selective effects of interference on implicit and explicit memory for new associations. Journal of experimental psychology. Learning, memory, and cognition. 1987;13(1):45. [Google Scholar]
- Graf P, Schacter DL. Implicit and explicit memory for new associations in normal and amnesic subjects. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1985;11(3):501–518. doi: 10.1037//0278-7393.11.3.501. [DOI] [PubMed] [Google Scholar]
- Grossi G. Relatedness proportion effects on masked associative priming: An ERP study. Psychophysiology. 2006;43:21–30. doi: 10.1111/j.1469-8986.2006.00383.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Semantic priming and stimulus degradation: implications for the role of the N400 in language processing. Psychophysiology. 1993;30:47–61. doi: 10.1111/j.1469-8986.1993.tb03204.x. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ, Neville HJ. Auditory and visual semantic priming in lexical decision: a comparison using event-related brain potentials. Language and Cognitive Processes. 1990;5:281–312. [Google Scholar]
- Holcomb PJ, Neville HJ. Natural speech processing: An analysis using event-related brain potentials. Psychobiology. 1991;19(4):286. [Google Scholar]
- Keane MM, Gabrieli JDE, Monti LA, Fleischman DA, Cantor JM, Noland JS. Intact and impaired conceptual memory processes in amnesia. Neuropsychology. 1997;11(1):59–69. doi: 10.1037//0894-4105.11.1.59. [DOI] [PubMed] [Google Scholar]
- Kiefer M. The N400 is modulated by unconsciously perceived masked words: Further evidence for an automatic spreading activation account of N400 priming effects. Cognitive Brain Research. 2002;13:27–39. doi: 10.1016/s0926-6410(01)00085-4. [DOI] [PubMed] [Google Scholar]
- Kucera H, Francis WN. Computational Analysis of Present-Day American English. Providence: Brown University Press; 1967. [Google Scholar]
- Kutas M, Hillyard S. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science. 1980;207(4427):203. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Kutas M, Hillyard S. Brain potentials during reading reflect word expectancy and semantic association. Nature. 1984;307(5947):161. doi: 10.1038/307161a0. [DOI] [PubMed] [Google Scholar]
- Kutas M, Van Petten C, Besson M. Event-related potential asymmetries during the reading of sentences. Electroencephalography and clinical neurophysiology. 1988;69(3):218. doi: 10.1016/0013-4694(88)90131-9. [DOI] [PubMed] [Google Scholar]
- Lau E, Phillips C, Poeppel D. A cortical network for semantics: (de)constructing the N400. Nature Reviews Neuroscience. 2008;9:920–933. doi: 10.1038/nrn2532. [DOI] [PubMed] [Google Scholar]
- McCarthy G, Wood CC. Scalp distributions of event-related potentials: An ambiguity associated with analysis of variance models. Electroencephalography and Clinical Neurophysiology. 1985;62:203–208. doi: 10.1016/0168-5597(85)90015-2. [DOI] [PubMed] [Google Scholar]
- McLaughlin J, Osterhout L, Kim A. Neural correlates of second-language word learning: Minimal instruction produces rapid change. Nature Neuroscience. 2004;7(7):703–704. doi: 10.1038/nn1264. [DOI] [PubMed] [Google Scholar]
- Mestres-Misse A, Rodriguez-Fornells A, Munte TF. Watching the brain during meaning acquisition. Cerebral Cortex. 2007;17(8):1858–1866. doi: 10.1093/cercor/bhl094. [DOI] [PubMed] [Google Scholar]
- Milner B, Corkin S, Teuber HL. Further analysis of the hippocampal amnesic syndrome: 14-year follow-up study of HM. Neuropsychologia. 1968;6(3):215. [Google Scholar]
- Mitchell P, Andrews S, Ward P. An event-related potential study of semantic congruity and repetition in a sentence-reading task : effects of context change. Psychophysiology. 1993;30(5):496–509. doi: 10.1111/j.1469-8986.1993.tb02073.x. [DOI] [PubMed] [Google Scholar]
- Moscovitch M. Memory as assessed by recognition and reading time in normal and memory-impaired people with Alzheimer's disease and other neurological disorders. Journal of experimental psychology. General. 1986;115(4):331. doi: 10.1037//0096-3445.115.4.331. [DOI] [PubMed] [Google Scholar]
- Nagy WE, Herman PA. The nature of vocabulary acquisition. Hillsdale, NJ, England: Lawrence Erlbaum Associates, Inc.; 1987. Breadth and depth of vocabulary knowledge: Implications for acquisition and instruction; pp. 19–35. [Google Scholar]
- Neville HJ, Kutas M, Chesney G, Schmidt AL. Event-related brain potentials during initial encoding and recognition memory of congruous and incongruous words. Journal of Memory and Language. 1986;25(1):75–92. [Google Scholar]
- Newport EL, Bavelier D, Neville HJ. Language, brain, and cognitive development: Essays in honor of Jacques Mehler. Cambridge, MA, US: The MIT Press; 2001. Critical thinking about critical periods: Perspectives on a critical period for language acquisition; pp. 481–502. [Google Scholar]
- Paller KA. Recall and stem-completion priming have different electrophysiological correlates and are modified differentially by directed forgetting. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1990;16(6):1021–1032. doi: 10.1037//0278-7393.16.6.1021. [DOI] [PubMed] [Google Scholar]
- Paller KA, Hutson CA, Miller BB, Boehm SG. Neural Manifestations of Memory with and without Awareness. Neuron. 2003;38(3):507–516. doi: 10.1016/s0896-6273(03)00198-3. [DOI] [PubMed] [Google Scholar]
- Paller KA, Kutas M, Mayes AR. Neural correlates of encoding in an incidental learning paradigm. Electroencephalography & Clinical Neurophysiology. 1987;67(4):360–371. doi: 10.1016/0013-4694(87)90124-6. [DOI] [PubMed] [Google Scholar]
- Paller KA, McCarthy G, Wood CC. ERPs predictive of subsequent recall and recognition performance. Biological Psychology. 1988;26(1–3):269–276. doi: 10.1016/0301-0511(88)90023-3. [DOI] [PubMed] [Google Scholar]
- Perfetti CA. Word learning and individual differences in word learning reflected in event-related potentials. Journal of experimental psychology. Learning, memory, and cognition. 2005;31(6):1281. doi: 10.1037/0278-7393.31.6.1281. [DOI] [PubMed] [Google Scholar]
- Roeder B, Rosler F, Neville H. Auditory memory in congenitally blind adults: a behavioral-electrophysiological investigation. Cognitive Brain Research. 2001;11:289–303. doi: 10.1016/s0926-6410(01)00002-7. [DOI] [PubMed] [Google Scholar]
- Rolke B, Heil M, Streb J, Hennighausen E. Missed prime words within the attentional blink evoke an N400 semantic priming effect. Psychophysiology. 2001;38:165–174. [PubMed] [Google Scholar]
- Rugg MD. Dissociation of the neural correlates of implicit and explicit memory. Nature. 1998;392(6676):595. doi: 10.1038/33396. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Doyle MC. Event-Related Potentials and Recognition Memory for Low- and High-Frequency Words. Journal of cognitive neuroscience. 1992;4(1):69–79. doi: 10.1162/jocn.1992.4.1.69. [DOI] [PubMed] [Google Scholar]
- Sanford AJ, Leuthold H, Bohan J, Sanford AJS. Anomalies at the Borderline of Awareness: An ERP Study. Journal of cognitive neuroscience. doi: 10.1162/jocn.2009.21370. (in press) [DOI] [PubMed] [Google Scholar]
- Schott B, Richardson-Klavehn A, Heinze H-J, Duzel E. Perceptual priming versus explicit memory: Dissociable neural correlates at encoding. Journal of cognitive neuroscience. 2002;14(4):578–592. doi: 10.1162/08989290260045828. [DOI] [PubMed] [Google Scholar]
- Shelton JR, Martin RC. How semantic is automatic semantic priming? Journal of Experimental Psychology: Learning, Memory, and Cognition. 1992;18(6):1191–1210. doi: 10.1037//0278-7393.18.6.1191. [DOI] [PubMed] [Google Scholar]
- Squire LR, Zola SM. Structure and function of declarative and nondeclarative memory systems. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(24):13515. doi: 10.1073/pnas.93.24.13515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sternberg RJ. The nature of vocabulary acquisition. Hillsdale, NJ, England: Lawrence Erlbaum Associates, Inc.; 1987. Most vocabulary is learned from context; pp. 89–105. [Google Scholar]
- Van Petten C, Kutas M, Kluender R, Mitchiner M, McIsaac H. Fractionating the word repetition effect with event-related potentials. Journal of cognitive neuroscience. 1991;3(2):131–150. doi: 10.1162/jocn.1991.3.2.131. [DOI] [PubMed] [Google Scholar]
- Verfaellie M, Martin E, Page K, Keane M. Implicit memory for novel conceputal associations in amnesia. Cognitive Affective Behavioral Neuroscience. 2006;6(20):91–101. doi: 10.3758/cabn.6.2.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber-Fox C, Neville HJ. Maturational constraints on functional specializations for language processing: ERP and behavioral evidence in bilingual speakers. Journal of Cognitive Neuroscience. 1996;8(3):231. doi: 10.1162/jocn.1996.8.3.231. [DOI] [PubMed] [Google Scholar]