


Abstract
Three different base paired stems form between U2 and U6 snRNA over the course of the mRNA splicing reaction (helices I, II and III). One possible function of U2/U6 helix II is to facilitate subsequent U2/U6 helix I and III interactions, which participate directly in catalysis. Using an in vitro trans-splicing assay, we investigated the function of sequences located just upstream from the branch site (BS). We find that these upstream sequences are essential for stable binding of U2 to the branch region, and for U2/U6 helix II formation, but not for initial U2/BS pairing. We also show that non-functional upstream sequences cause U2 snRNA stem–loop IIa to be exposed to dimethylsulfate modification, perhaps reflecting a U2 snRNA conformational change and/or loss of SF3b proteins. Our data suggest that initial binding of U2 snRNP to the BS region must be stabilized by an interaction with upstream sequences before U2/U6 helix II can form or U2 stem–loop IIa can participate in spliceosome assembly.
INTRODUCTION
Introns are spliced out of mRNA precursors by two sequential transesterification reactions. These reactions are catalyzed by the spliceosome, a large multicomponent complex of small nuclear ribonucleoprotein particles (snRNPs) and numerous proteins (reviewed in 1–6). Splicing of major class U2-dependent introns involves sequential and coordinated binding of five snRNPs (U1, U2, U4, U5 and U6) to the mRNA precursor. In the conventional view, U1 first binds across the exon/intron boundary at the 5′-end of introns [5′ splice site (SS); 7–9], leading to formation of an ATP-independent complex committed to splicing (complex E in mammalian extract and complexes CC1 and CC2 in yeast extract; 10–13). Upon addition of ATP, complex E assembles into complex A, U2 stably associates with the branch site (BS) and U1 dissociates from the 5′ SS (14–18). Lastly, the U4/U5/U6 tri-snRNP complex assembles with complex A to form complex B, which is transformed into complex C by the first transesterification and into complex D by the second (19–22). However, an exciting alternative is that the U1 and U4/U5/U6 snRNPs work together in an ATP-dependent process to identify the 5′ SS (23), independent of U2 snRNP binding to the 3′ SS (24–28). This would imply that formation of complex B reflects stabilization of U4/U5/U6 tri-snRNP already bound to the spliceosome, not recruitment of free U4/U5/U6 tri-snRNP.
Although the BS is highly degenerate in mammals (24), pairing of U2 with the BS is required to bulge the residue (almost always an adenosine) that initiates the first transesterification reaction (25–32). U2 binding to the BS occurs in multiple steps. In yeast, the BS is first recognized by a cooperative interaction between two proteins, mBBP/SF1 and U2AF65 (33); this recognition takes place in a commitment complex (CC2) that appears to be the homolog of the E complex (13,32,33). U2 snRNP can associate with complex E independently of the branch site in the absence of ATP (34); in the presence of ATP, complex E is transformed into complex A where U2 recognizes the BS directly by base pairing (35,36). Although U2 can bind the BS in the absence of ATP (37,38), the ATP dependence of U2 snRNP recruitment and prespliceosome assembly are normally assured by PRP5, a member of the ATP-dependent RNA helicase family, working under the control of CUS2 (the yeast homolog of human Tat-SF1) (38,39).
In mammals, two intron elements located on either side of the BS regulate stable U2/BS binding: (i) the polypyrimidine tract, located downstream of the BS, binds U2AF65 and facilitates U2/BS binding (33,40,41) and (ii) the anchoring site, a 20 nt region upstream of the BS, binds two U2 snRNP-associated protein complexes (18) known as SF3a and SF3b (42,43; for review see 44). SF3a and SF3b proteins are highly conserved between mammals and yeast, and are essential for assembly of complex A (18,42,43,45–49). An antisense oligonucleotide directed against the anchoring site blocks assembly of complex A, suggesting an essential role for this region in stable association of U2 snRNP with the BS (18); however, mutations in the anchoring site apparently have no effect on cis-splicing (18). Thus, the anchoring site binds SF3 proteins and is essential for complex A assembly, but appears to function in a sequence-independent fashion.
When bound to the BS, U2 forms three different base paired interactions with U6 (helices I, II and III; Fig. 1A). Helix II, which forms between the 5′-end of U2 and the 3′-end of U6, is conserved through evolution (50), and is functionally important in mammals (51,52) although entirely dispensable in yeast (50,53). Helix II may position U2 and U6 snRNA to facilitate the other two U2/U6 interactions. Helix I (subdivided into helix Ia and Ib), which forms between U2 residues just 5′ to the BS recognition sequence and U6 residues just 3′ to the invariant ACAGAGA sequence, is essential in yeast (54,55) and mammals (56). Although helix Ib initially appeared to be functionally redundant with helix II in yeast (53), helix Ib has subsequently been found to play a role in 5′ SS selection that cannot be performed by helix II (57). Helix III, which is essential in mammals, may help to juxtapose the 5′ SS and BS before the first catalytic step (56) but the role of helix III in yeast is still unclear (39,58).
Figure 1.


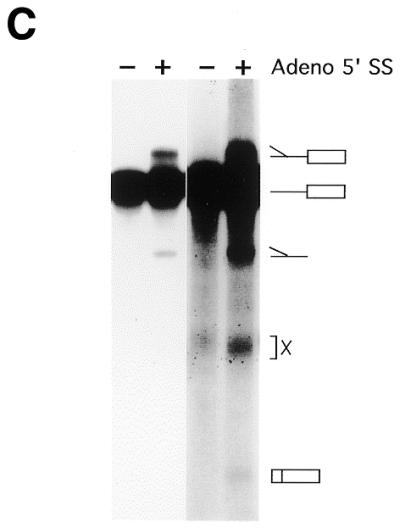
Substrates, crosslinks and trans-splicing assay. (A) U2 snRNA interactions with U6 snRNA and the mRNA precursor. U snRNA stem–loops are numbered conventionally (4); U2/U6 helices I, II and III are indicated as hI, hII and hIII; the branch site is denoted BS; and the exons are boxed. Partial, complete or alternative base pairing is denoted by parallel lines, loops and bulges by curved lines. U2 snRNA can in principle pair with the BS or with U6 to extend helix III. Adapted from (88). (B) trans-splicing substrates. The Adeno 5′ SS is a synthetic oligonucleotide with a 3′ terminal deoxythymidine to prevent degradation. The Adeno 3′ RNA contains the anchoring site (AS), the most probable BS (underlined; see 24, fig. 3), polypyrimidine tract (Py), 3′ splice site (3′ SS) and 3′ exon. (C) In vitro trans-splicing. 32P-labeled Adeno 3′ RNA was incubated with unlabeled Adeno 5′ SS oligonucleotide in HeLa nuclear extract at 30°C for 120 min. RNA was purified and resolved by denaturing 12% PAGE. The positions of Y-branched splicing intermediate, Adeno 3′ RNA, Y-branched intron and X are shown on the right. X is an exonuclease degradation product of Adeno 3′ RNA resulting from protection in the BS region (89–93). Left to right, lanes 1–4. Splicing intermediates are seen most easily on short exposure (lanes 1 and 2), ligated exons on long exposure (lanes 3 and 4).
We decided to examine the role of sequences upstream from the BS because deletion of this region inhibited formation of complex A in a trans-splicing assay (G.Ast and A.M.Weiner, submitted for publication) as originally observed for cis-splicing (18). In addition, because the U2 snRNP-associated SF3 proteins interact with the anchoring site (18), we were also intrigued by the observation that SF3 proteins (59) can suppress mutations in U2 stem–loop IIa, suggesting that SF3 proteins and/or the U2 snRNP-associated CUS2 protein (60) could mediate a U2 conformational change between alternative phylogenetically conserved structures (49,59–62). Consistent with these earlier observations, we show here that (i) functional sequences upstream of the branch site are required for stable U2/BS association in complex A and for formation of U2/U6 helix II; and (ii) non-functional upstream sequences not only fail to support stable assembly of complex A, but cause misfolding of U2 stem–loop IIa.
MATERIALS AND METHODS
Oligonucleotides and plasmids
Oligonucleotides were synthesized by J.Flory (Oligos R Us, Yale School of Medicine) on an Applied Biosystems machine. RNA oligonucleotides were deprotected, desalted and purified by denaturing 20% PAGE. To generate the four Adeno 3′ RNA templates, a 146 bp fragment from clone pSpAde (63; GenBank accession no. J01917), extending from 23 bp upstream of the branch site to 106 bp downstream of the 3′ SS, was amplified by PCR using either of two 5′ primers containing a KpnI site (5′-GGTACCGTTTCCTTGAT-3′ to generate fragment UP, 5′-GGTACCGTCATACTTATC-3′ to generate fragment –UP) and a single 3′ primer containing an EcoRI site (5′-GAATTCCAACCAGTTCTAC-3′) where restriction sites are underlined. The –UP fragment lacks the anchoring site located in the 5′-most 15 bp of the UP fragment. The amplified fragments were digested with EcoRI and KpnI, and cloned into Bluescript SK+ (Stratagene) to generate constructs UP and –UP. The UP-GG+100 construct is identical to –UP, but contains, just upstream of the insert, an inverted duplication of the Bluescript polylinker fragment (KpnI to EcoRI) accidentally generated during cloning. Construct UP-GG was derived from UP-GG+100 by deleting the 54 bp SalI fragment from the duplicated polylinkers. Construct UP-GA was derived from UP-GG by site-directed mutagenesis (64). All constructs were verified by sequencing. Constructs were linearized with ScaI, and RNA substrates prepared by runoff transcription using T7 RNA polymerase and labeled UTP. The resulting RNA transcripts were 99 (WT), 126 (UP-GG and UP-GA) and 181 nt (UP-GG+100). Although upstream sequences in UP-GG+100 could in principle be sequestered by formation of a stem–loop between inverted repeat sequences, there is ample evidence that nuclear extracts melt RNA secondary structure. For example, antisense DNA oligonucleotides, or even 2′-O-methyl RNA oligonucleotides, do not protect mRNA precursors from T1 digestion in nuclear extract (G.Ast, unpublished observations).
Trans-splicing reactions, crosslinking assays and spliceosomal complex analysis
Standard reactions (12.5 µl) containing 60% (v/v) HeLa nuclear extract, 2.2 mM MgCl2, 0.5 mM ATP, 20 mM creatine phosphate and 6 × 104 c.p.m. 3′-32P-labeled substrate were preincubated for 20 min at 30°C. Spliceosome complexes were analyzed by native 4% PAGE as described (20) but using 0.5 mg/ml heparin instead of 5 mg/ml in the loading buffer. Crosslinking was performed as described (50,65–69). AMT psoralen was added (20 µg/ml) and the reactions irradiated at 365 nm for 10 min at 4°C at a distance of 5 cm from the light source (Chromato-UVE Transilluminator, Model C-62; Ultra-Violet Products).
RNase H digestion and primer extension
Standard RNase H digestion reactions (20 µl) contained crosslinked RNA (10 000 c.p.m.), 40 mM Tris–HCl pH 7.5, 50 mM KCl, 5 mM MgCl2, carrier RNA (40 ng/µl) and DNA oligonucleotide (250 ng/µl). Reactions were denatured for 3 min at 90°C, annealed by cooling to 37°C over a period of 30 min in a heat block and digested with RNase H (2 µl of 1 U/µl; Boehringer) for 35 min at 37°C. Reactions were stopped by heating to 90°C for 3 min, followed by ethanol precipitation. Primer extension was performed as described (67–69) under standard conditions (50 mM Tris–HCl, 8 mM MgCl2, 30 mM KCl, 1 mM DTT pH 8.5) in 10 µl reactions containing 1 µl of a 100-fold dilution of total RNA from deproteinized nuclear extract.
Dimethylsulfate (DMS) modification
Chemical modification with DMS was carried out as described (69) except that incubation with DMS was for 12 min.
RESULTS
Upstream sequences stabilize the U2/BS interaction and are essential for U2/U6 helix II formation
As documented in detail elsewhere (G.Ast and A.M.Weiner, submitted for publication), trans-splicing can take place in HeLa nuclear extract between an Adeno 5′ SS RNA oligonucleotide and an Adeno 3′ RNA substrate extending from the 3′ exon to sequences known as the intron anchoring site upstream from the branch site (Fig. 1B and C) (18).
To examine the function of these sequences in trans-splicing, this region of the Adeno 3′ RNA was mutated to two different sequences, one G-rich and the other G+A-rich (Fig. 1B, UP-GG and UP-GA, respectively). To understand why the UP-GG and UP-GA substrates failed to trans-splice (data not shown), we incubated the 32P-labeled Adeno 3′ substrates (Adeno 3′, UP-GG and UP-GA) in HeLa nuclear extract for 20 min at 30°C with or without Adeno 5′ SS RNA. The reaction was then divided in two; half was resolved by native PAGE to examine spliceosomal complex assembly (Fig. 2A) while the other half was crosslinked with psoralen, deproteinized and the RNAs analyzed by denaturing PAGE to examine base pairing of U2 with the BS and with U6 snRNA (Fig. 2B) (50,65–69).
Figure 2.
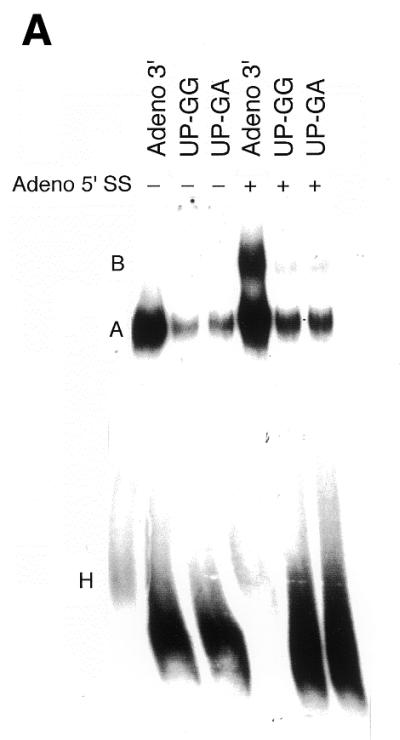
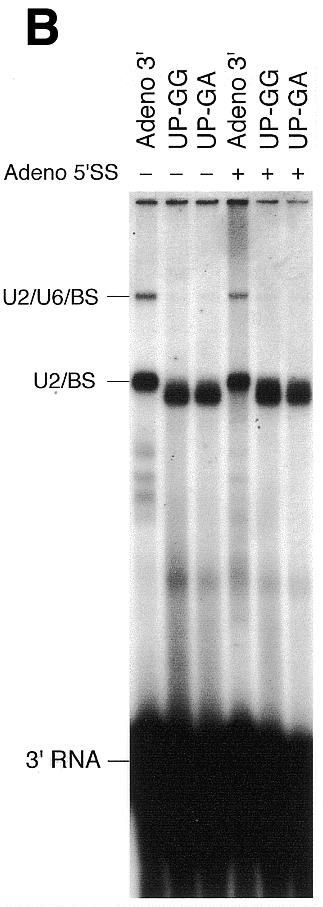
Sequences upstream of the branch site are essential for complex A assembly and formation of U2/U6 helix II within the trans-spliceosome. (A) Upstream mutations affect assembly of the Adeno 3′ RNA substrate into complex A. Nuclear extracts were incubated with 32P-labeled Adeno 3′ RNA, or the UP-GG and UP-GA substrates (10 ng), with (+) or without (–) unlabeled Adeno 5′ SS oligonucleotide (440 ng), under trans-splicing conditions at 30°C for 20 min (G.Ast and A.M.Weiner, submitted for publication). RNP complexes were then resolved by native 4% PAGE. The positions of the H, A and B (U2/U4/U5/U6/3′ RNA) complexes are indicated on the left. Formation of complex A was both ATP- and temperature-dependent (data not shown). Left to right, lanes 1–6. (B) Upstream mutations affect the U2/U6 helix II interaction. After incubation as in (A), reactions were crosslinked with psoralen on ice, and the deproteinized RNAs resolved by denaturing 5% PAGE. The identities of crosslinked products obtained with the Adeno 3′ RNA substrate are indicated on the left (see Fig. 4) (70). The identity of the U2/BS crosslinks obtained with the UP-GG and UP-GA substrates was confirmed by RNase H protection with a DNA oligonucleotide complementary to U2 positions 28–42; cleavage of both complexes with a DNA oligonucleotide complementary to the 3′ SS served as a positive control (Fig. 4 and data not shown). The wild-type and mutant U2/BS crosslinks differ in size because the wild-type Adeno 3′ RNA substrate has 27 additional 5′ terminal nucleotides. Left to right, lanes 1–6.
Native PAGE analysis of complexes formed in the absence of the Adeno 5′ SS revealed that UP-GG and UP-GA mutations reduced complex A (U2/3′ RNA) by >6-fold (87% in Fig. 2A, average of 82%) compared to the wild-type Adeno 3′ substrate as judged by densitometry (Fig. 2A, lanes 1–3). In the presence of unlabeled Adeno 5′ SS, complex A progressed to complex B (U2/U4/U5/U6/3′ RNA) by addition of the U4/U5/U6 (Fig. 2A, lanes 4–6). However, replacing the normal upstream sequences with G-rich or G+A-rich sequences (UP-GG or UP-GA) not only reduced assembly of complex A by 4-fold (74%) in the presence of Adeno 5′ SS, but severely inhibited progression to complex B: 25% of complex A advanced to complex B using the Adeno 3′ substrate, but only 5% using the upstream mutants (Fig. 2A, lanes 5 and 6). Conceivably, complex A contains a mixture of complexes representing different stages of U2 assembly on the mRNA precursor; the mutant UP-GG and UP-GA substrates might then arrest a subset of these complexes prior to the stabilization of U2/BS interaction, or increase the fraction of incompetent complexes that cannot progress to complex B.
To determine whether upstream mutations affect formation or stability of complexes A and B, we examined base pairing of U2 with the branch site and with U6 snRNA. Half of each reaction shown in Figure 2A was crosslinked with psoralen, and the deproteinized RNAs resolved by denaturing 6% PAGE (Fig. 2B). As previously observed (70), two crosslinked products were formed: U2 singly crosslinked to the BS sequence (U2/BS) and U2 doubly crosslinked to both the branch site and U6 (U2/U6/BS; Fig. 2B, lane 1). Several conclusions can be drawn by comparing Figure 2A and B. First, the UP-GG and UP-GA mutations severely reduced complex A when assayed by native PAGE (Fig. 2A) but had no effect on the U2/BS crosslink in the absence or presence of a 5′ SS (Fig. 2B). Thus, upstream sequences stabilize, but are not required for, the U2/BS interaction. Secondly, although complex B is unstable to native PAGE in the absence of a 5′ SS (Fig. 2A, compare lanes 1 and 4) (70), the U2/U6/BS double crosslink can still be observed in the absence of a 5′ SS (Fig. 2B, lanes 1 and 4). Thus, the 5′ SS is not required for the U2/U6/BS double crosslink. Thirdly, the UP-GG and UP-GA mutants abolish the U2/U6/BS crosslink without affecting the U2/BS crosslink (Fig. 2B, compare lanes 2 and 5, and lanes 3 and 6, with lanes 1 and 4). Thus, sequences upstream of the branch site, although unnecessary for the U2/BS interaction, are essential for the U2/U6 helix II interaction (see Fig. 4 for identification of the U2/U6 helix II and U2/BS interactions). Fourthly, since the U2/U6/BS crosslink is abolished by the upstream mutants but the U2/BS crosslink is not, U2/U6 helix II must form after the U2/BS interaction, consistent with previous observations that U2 snRNP appears to bind the branch site independently of other snRNPs (reviewed in 1,3,4). Although based on a trans-splicing assay, these conclusions may also apply to normal cis-splicing: upstream sequences would stabilize binding of U2 to the branch site, and stable binding of U2 to the branch site would be required to stabilize U4/U5/U6 already bound to complex A (23) before U2/U6 helix II could form.
Figure 4.
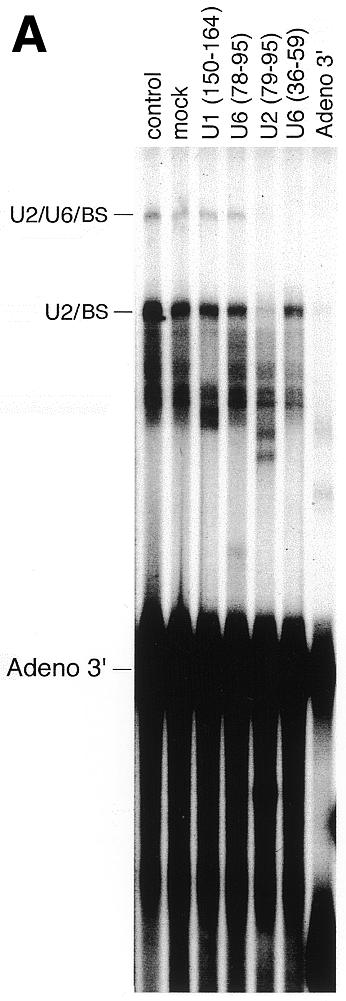
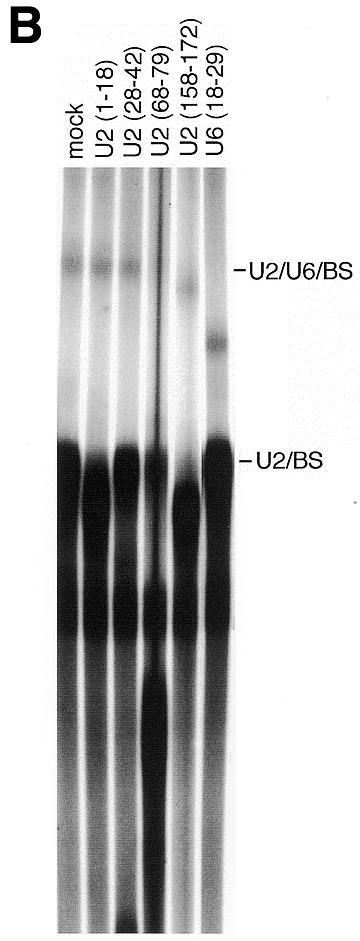
The U2/U6/BS crosslink contains U2/U6 helix II. (A and B) Unlabeled Adeno 5′ SS and 32P-labeled Adeno 3′ RNA were incubated in nuclear extract as in Figure 2A, and crosslinked as in Figure 2B. RNA was then purified, subjected to RNase H cleavage targeted by the indicated oligodeoxynucleotides, and the products resolved by denaturing 5% PAGE. The positions of the U2/BS and U2/U6/BS crosslinks, and the 3′ RNA substrate, are indicated. (A) Left to right, lanes 1–7. (B) Left to right, lanes 1–6.
Strictly speaking, we do not know whether the effects of the UP-GG and UP-GA mutations (Fig. 2) reflect the absence of a positive upstream sequence element required for normal spliceosome function, or the presence of negative (‘poisonous’) upstream sequences that prevent normal function. Indeed, the accumulation of non-specific complex H observed with both the UP-GG and UP-GA mutants (Fig. 2A) could indicate that formation of complex A and/or subsequent steps in spliceosome assembly is blocked by binding of hnRNP proteins to purine-rich upstream sequences. However, inhibition caused by binding of hnRNP (or other) proteins to purine-rich upstream sequences would be formally and functionally analogous to inhibition caused by binding of an antisense oligonucleotide, as originally used to demonstrate the importance of the anchoring site just upstream from the branch site (18). Thus, whether the purine-rich upstream sequences bind inhibitory proteins, or fail to interact with the correct spliceosomal components, our data underscore the importance of sequences located upstream from the BS, and suggest roles for this region in the early steps of spliceosome assembly.
To determine whether the UP-GG mutant can be rescued by additional 5′ sequence, we performed the same experiment as in Figure 2 using UP-GG+100, a derivative of UP-GG containing ∼100 additional upstream nucleotides (see Fig. 1B); however, this had no obvious effect other than to increase the fraction of substrate remaining in the non-specific complex H (Fig. 3A and B, and data not shown). Curiously, two different U2/BS crosslinks are sometimes observed (Fig. 3B) and both are resistant to RNase H digestion targeted by U2(28-42) (Fig. 4B). The existence of two crosslinks may indicate that the U2/BS interaction changes as splicing progresses; however, the lower U2/BS crosslink band was not consistently observed, and may depend on nuclear extract, psoralen batch or minor differences in experimental protocol.
Figure 3.
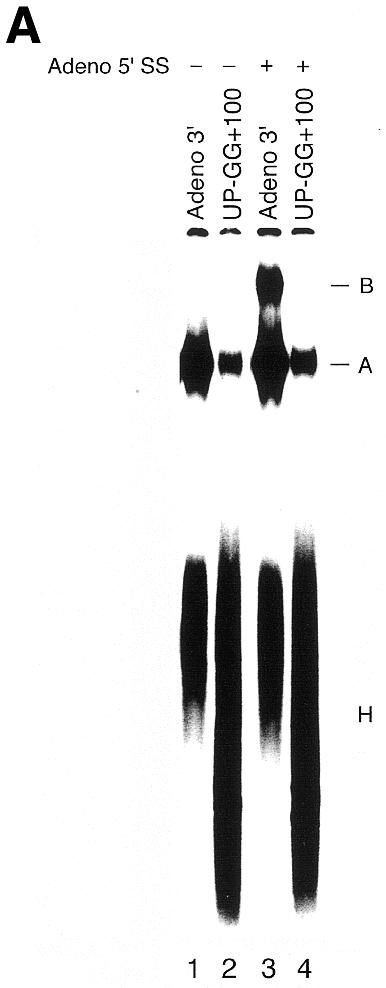
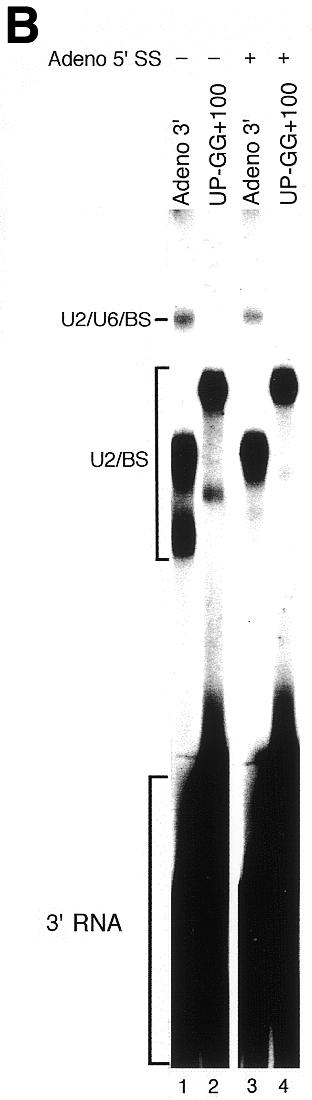
The UP-GG upstream mutant cannot be rescued by additional intron sequences. (A and B) As in Figure 2A and B, but using a UP-GG derivative containing 100 additional 5′ nucleotides. The Adeno 3′ and UP-GG+100 substrates differ in size (see Materials and Methods) causing equivalent crosslinked products to have different gel mobilities, as indicated on left.
The U2/U6/BS crosslink contains U2/U6 helix II
U2 bound to the BS can form three base pairing interactions with U6 (Fig. 1A): helix I between positions 23–25 of U2 and 49–51 of U6 (54–56); helix II between positions 3–8 of U2 and 90–95 of U6 (50–52); and helix III between positions 36–49 of U2 and 30–42 of U6 (56). We used an RNase H cleavage assay to determine which of these interactions are responsible for the observed U2/U6/BS crosslinks (Fig. 4). Uniformly labeled Adeno 3′ RNA was incubated in nuclear extract, crosslinked with psoralen on ice, and the RNA then deproteinized and purified as in Figure 2B. The U2/U6/BS crosslink is resistant to RNase H digestion targeted by oligodeoxynucleotides U6(78-95) and U2(1-18) (Fig. 4A and B, respectively), indicating that U2/U6 helix II forms within the U2/U6/BS complex. In contrast, the U2/U6/BS crosslink is digested when RNase H is targeted by oligodeoxynucleotides U2(79-95) and U6(36-59) (Fig. 4A), or U2(68-79), U2(158-172) and U6(18-29) (Fig. 4B), indicating that U6 sequences which participate in helix I and III are not protected from digestion. As expected, the U2/BS crosslink is digested when RNase H is targeted by U2(1-18) because the 5′-end of U2 is not protected by U2/U6 helix II as it is in the U2/U6/BS crosslink (Fig. 4B). Also, as expected if U2 is base paired to the BS within the crosslinked U2/BS and U2/U6/BS species, U2 in both crosslinked species is protected from digestion targeted by the U2(28-42) oligodeoxynucleotide complementary to the branch site binding region (Fig. 4B).
Non-functional upstream sequences induce a conformational change in U2 stem–loop IIa
Three observations led us to suspect that binding of U2 snRNP to upstream sequences, including the anchoring site, might induce a U2 snRNA conformational change: (i) functional upstream sequences stabilize U2/BS binding in complex A (compare Fig. 2A and B) and induce U2/U6 helix II formation (Fig. 4); (ii) a region within the upstream sequences called the anchoring site is bound in complex A by intrinsic U2 snRNP proteins known as SF3 (18); and (iii) yeast SF3 proteins can functionally suppress mutations in U2 snRNA stem–loop IIa (49,59,61,62).
We used DMS modification to detemine whether U2 snRNP undergoes a conformational change when bound to the Adeno 3′ RNA or the UP-GG mutant (Fig. 5A). The Adeno 3′ RNA substrate (lanes 7 and 9) or the UP-GG mutant (lanes 8 and 10) were incubated in nuclear extract for 20 min at 30°C with (lanes 9 and 10) or without (lanes 7 and 8) addition of the Adeno 5′ SS (Fig. 5A). The reactions were then modified with DMS, RNA was purified and U2 modification sites determined by primer extension using a labeled oligodeoxynucleotide complementary to U2(79-98). Unexpectedly, strong blocks to primer extension were seen at U2 nucleotides A57, A59, U60 and U65 with the UP-GG mutant, but not with the Adeno 3′ RNA (Fig. 5A, compare lanes 8 and 10 with lanes 7 and 9; shown schematically in Fig. 5B). Weak blocks were also seen at U2 nucleotides U54 and G63. All of these blocks were independent of the Adeno 5′ SS (Fig. 5A, compare lanes 8 and 10) indicating that U2 stem–loop IIa partially unfolds, misfolds or interacts differently with proteins when complex A assembles on the UP-GG mutant.
Figure 5.
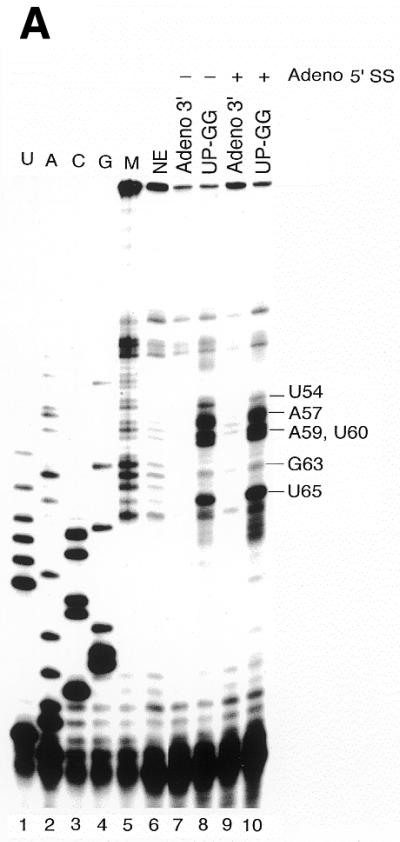


U2 snRNP undergoes a conformational change upon binding to the mutant UP-GG substrate containing G-rich sequences upstream of the BS. (A) Primer extension assay for U2 snRNA conformational changes. Nuclear extracts were incubated with wild-type Adeno 3′ RNA or the UP-GG mutant under trans-splicing conditions for 20 min at 30°C, with or without the Adeno 5′ SS. The reactions were treated for 12 min at room temperature with DMS (2 µl of DMS in dioxane, 5% v/v). DMS modification sites were detected by primer extension using purified RNA as template and U2(79-98) as primer. Modification sites were localized by comparison to a sequencing ladder generated with the same primer (lanes 1–6). (B) The DMS reactivity of nucleotides in U2 stem–loop IIa is denoted by arrowheads, with larger arrowheads indicating greater reactivity. For reference, the alternative U2 conformation (39,49) is shown schematically. (C) The U2/AS interaction, possibly involving U2 snRNP component SF3b, stabilizes the initial U2/BS interaction and allows formation of U2/U6 helix II.
Changes in DMS reactivity within stem–loop IIa could in principle reflect altered RNA conformation and/or altered protein binding. When U2 snRNP assembles onto the mutant UP-GG substrate, the reactivity of U65 at the base of stem IIa increases dramatically, but the reactivity of other nucleotides within stem IIa is almost unchanged. Thus, increased reactivity throughout stem–loop IIa may reflect loss of proteins such as SF3b that bind the stem–loop, rather than a significant conformational change within the RNA itself. We conclude that the non-functional UP-GG mutant substrate not only destabilizes complex B (compare Fig. 2A and B) and prevents U2/U6 helix II from forming (Fig. 2B) but causes U2 snRNP to undergo an aberrant conformational change that unshields or unfolds U2 stem–loop IIa (Fig. 5A).
DISCUSSION
Using a trans-splicing assay, we have found that (i) functional sequences upstream from the BS are essential for stable U2/BS pairing in complex A, but not for initial U2/BS pairing; (ii) functional upstream sequences are required to form U2/U6 helix II; and (iii) non-functional upstream sequences cause U2 stem–loop IIa to become accessible to DMS modification, perhaps reflecting loss of SF3b proteins and/or a U2 snRNA conformational change.
Assuming that the trans-splicing assay resembles certain aspects of cis-splicing (see below for qualification), our results suggest a multistep model for U2/BS binding (Fig. 5C). First, U2 snRNP pairs with the BS in a low affinity complex that is unstable in the absence of functional upstream sequences (Fig. 2A and B). Secondly, the low affinity U2/BS complex is stabilized by additional interaction(s) requiring functional upstream sequences (Fig. 2A and B). And thirdly, interaction with upstream sequences triggers a U2 snRNP conformational change leading to U2/U6 helix II when the upstream sequences are functional (see Fig. 1A), but to an aberrant conformational change when upstream sequences are non-functional (Fig. 5A).
U2/U6 helix II has been known for some time (39,50–52,56–58). It is also known that U2 snRNP can bind independently of other snRNPs to the BS (for examples see 37,38) and to the 5′ SS (34), and that the U4/U5/U6 snRNP can bind to the 5′ SS independently of U2 snRNP (23,71). Yet it is still unclear whether U2 must bind to the BS before U2/U6 helix II can form. Our data demonstrate that, at least in a trans-splicing system, U2/U6 helix II does not form until after U2 snRNA pairs with the BS (Fig. 2A and B). Thus, if the mammalian spliceosome is assembled stepwise, and not by binding of the mRNA precursor to a preassembled penta-snRNP complex, formation of U2/U6 helix II may (i) consolidate a bridging interaction between U4/U5/U6 bound to the 5′ SS and U2 bound to the BS, and (ii) prepare U2 snRNP for a subsequent conformational change involving stem–loop IIa (Fig. 5 and discussion below).
Why are upstream sequences required to form U2/U6 helix II? One attractive scenario involves the SF3a and SF3b components of the large SF3 protein complex that is tightly associated with U2 snRNP. SF3 proteins are highly conserved from yeast to humans, and are essential for splicing (18,42–46,48,49,72). SF3a associates primarily with the 3′ half of U2 snRNA, and SF3b with the 5′ half including U2 stem–loops Ib and IIa (73). Intriguingly, SF3 proteins in complex A can also bind to a 20 nt region of the mRNA precursor, known as the anchoring site, that is located just upstream of the BS (18). Moreover, SF3 components can functionally suppress U2 stem–loop IIa mutations in yeast (49,59,61,62). Thus, SF3b proteins might interact simultaneously with U2 snRNA stem–loop IIa and with the anchoring site on the mRNA precursor; alternatively, SF3b might be partially transferred from U2 snRNA to the anchoring site on the mRNA precursor (Fig. 5C). In either case, the SF3b/anchoring site interaction could trigger a U2 snRNP conformational change, allowing U2/U6 helix II to form when the upstream sequences are functional, but leading to an aberrant conformational change when the sequences are non-functional (Fig. 5A). The non-essential CUS2 protein, which possesses an RNA recognition motif (RRM) and interacts genetically with U2 stem–loop IIa, could also participate in this U2 conformation change (38,60).
U2 snRNP appears to exist in two conformations: the familiar conformation required for spliceosome assembly where U2 stem–loop IIa is intact (74), and a phylogenetically conserved alternative conformation where stem–loop IIa is disrupted in order to pair with a sequence immediately downstream from stem–loop IIb (39,74) (Fig. 5B). The alternative conformation is likely to be biologically significant because the potential for alternative base pairing is conserved from yeast to humans (74). Yet, despite prodigious effort (39,49,59), there has been no direct evidence that the alternative U2 conformation is required for, or occurs during, the normal mRNA splicing reaction. Judging by increased reactivity of U65, U2 loop IIa may be partially exposed in the aberrant U2 snRNP conformation brought about by trans-spliceosome assembly on the non-functional UP-GG mutant (Fig. 5A). Thus, the altered U2 snRNP conformation caused by a non-functional anchoring site, and the phylogenetically conserved alternative U2 snRNA conformation, could be functionally related. For example, the aberrant U2 conformation might resemble a structural intermediate normally formed as U2 snRNA refolds into the phylogenetically conserved alternative conformation.
We do not know whether the G-rich UP-GG and UP-GA mutants inhibit trans-splicing non-specifically by binding hnRNP proteins (consistent with the observation that the non-functional mutants increase formation of the non-specific complex H; Figs 2A and 3A) or by preventing upstream sequences from interacting with U2 snRNP or other splicing factors. Nor do we know whether functional upstream sequences are defined by primary sequence, base composition or both. These uncertainties, however, do not affect our conclusion that upstream sequences are required to stabilize U2 binding to the BS, and to allow formation of U2/U6 helix II.
Our data are consistent with evidence that antisense oligonucleotides directed against the anchoring site can perturb (72) or block splicing (18). Surprisingly, however, mutations in the anchoring site had no apparent effect on cis-splicing of an Adeno mRNA precursor (18). This could mean that interactions of SF3b or other factors with upstream sequences are redundant in a cis-splicing assay, but not in a more stringent trans-splicing assay where the 5′ and 3′ splice sites are held together solely by spliceosomal interactions. In cis-splicing, the commitment complex brings the 5′ SS and the branch region into proximity before U2 snRNP associates with the BS (13,32,33,75). In trans-splicing, 5′ SS selection occurs after U2 snRNP is stably bound to the BS on the 3′ RNA substrate (76). Thus, binding of U2 snRNP to upstream sequences may stabilize the assembling trans-spliceosome until the 5′ SS can bind, partially compensating for loss of bridging interactions between the 5′ and 3′ SS. The dependence of our trans-splicing system on upstream sequences resembles other experimental tools, such as including an enhancer within the 3′ exon (77) or a 5′ SS downstream of the 3′ exon (76), that facilitate in vitro trans-splicing by exploiting normal (but partially redundant) spliceosomal interactions.
Although bridging interactions between the 5′ and 3′ SS normally stabilize the earliest steps of spliceosome assembly (10–13), these interactions are not absolutely essential for initial binding of the snRNPs: the U4/U5/U6 complex can bind the 5′ SS independently of U2 snRNP and the BS (23,71), U1 and U2 snRNP can associate with a 5′ SS RNA independent of the BS (75,78), and U2 can bind the 3′ RNA substrate independently of U1 snRNP and the 5′ SS (1,37,70). The absence of bridging interactions in our trans-splicing assay may explain why addition of U4/U5/U6 to the U2/3′ RNA complex (trans-splicing complex A) is unstable (Fig. 2A, lane 1) despite formation of an extended U2/U6 helix II as judged by psoralen crosslinking (Fig. 2B, lane 1); complex B (U2/U4/U5/U6/5′ SS/3′ RNA) would be stabilized only when bridging interactions were restored by addition of a 5′ SS (Fig. 2A, compare lanes 1 and 4).
Our data suggest that interaction of spliceosomal components with upstream sequences is one of many weak or redundant spliceosomal interactions that are essential only under special conditions. Other examples include U2/U6 stem II, which is dispensable for splicing in yeast (53,57) but not for accurate 5′ SS selection (57); U2/U6 stem II and III interactions, which are required for splicing of some mammalian introns but not others (51,52,56,79), and pairing between the invariant loop of U5 and the last 2 or 3 nt of the 5′ exon (80–85), which is dispensable for both steps of human (86) and the first step of yeast in vitro splicing (87). The existence of redundant interactions suggests that splicing of every intron may differ in detail, relying to different extents on a variety of redundant spliceosomal interactions. Indeed, trans-splicing of mammalian mRNAs could be viewed as a synthetic lethal strategy for biochemists: an experimental tool to identify ‘optimal’ spliceosomal interactions that are redundant in normal cis-splicing, but revealed when cis-splicing is partially disabled by physically separating the 5′ and 3′ RNA substrates.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Arnold D.Bailey for discussions and critical reading of the manuscript. This work was funded by NIH awards GM31073 and GM57606 to A.M.W.
References
- 1.Moore M.J., Query,C.C. and Sharp,P.A. (1993) Splicing of precursors to mRNA by the spliceosome. In Gesteland,R.F. and Atkins,J.F. (eds), The RNA World. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 303–357.
- 2.Horowitz D.S. and Krainer,A.R. (1993) Mechanisms for selecting 5′ splice sites in mammalian pre-mRNA splicing. Trends Genet., 10, 100–106. [DOI] [PubMed] [Google Scholar]
- 3.Will C.L. and Lührmann,R. (1997) Protein functions in pre-mRNA splicing. Curr. Opin. Cell Biol., 9, 320–328. [DOI] [PubMed] [Google Scholar]
- 4.Staley J.P. and Guthrie,C. (1998) Mechanical devices of the spliceosome: motors, clocks, springs, and things. Cell, 92, 315–326. [DOI] [PubMed] [Google Scholar]
- 5.Collins C.A. and Guthrie,C. (2000) The question remains: is the spliceosome a ribozyme? Nat. Struct. Biol., 7, 850–854. [DOI] [PubMed] [Google Scholar]
- 6.Reed R. (2000) Mechanisms of fidelity in pre-mRNA splicing. Curr. Opin. Cell Biol., 12, 340–345. [DOI] [PubMed] [Google Scholar]
- 7.Zhuang Y. and Weiner,A.M. (1986) A compensatory base change in U1 snRNA suppresses a 5′ splice site mutation. Cell, 46, 827–835. [DOI] [PubMed] [Google Scholar]
- 8.Seraphin B., Kretzner,L. and Rosbash,M. (1988) A U1 snRNA: pre-mRNA base pairing interaction is required early in yeast spliceosome assembly but does not uniquely define the 5′ cleavage site. EMBO J., 7, 2533–2538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Siliciano P.G. and Guthrie,C. (1988) 5′ splice site selection in yeast: genetic alterations in base pairing with U1 reveal additional requirements. Genes Dev., 2, 1258–1267. [DOI] [PubMed] [Google Scholar]
- 10.Ruby S.W. and Abelson,J. (1988) An early hierarchic role of U1 small nuclear ribonucleoprotein in spliceosome assembly. Science, 242, 1028–1035. [DOI] [PubMed] [Google Scholar]
- 11.Seraphin B. and Rosbash,M. (1989) Identification of functional U1 snRNA-pre-mRNA complexes committed to splicing assembly and splicing. Cell, 59, 349–358. [DOI] [PubMed] [Google Scholar]
- 12.Michaud S. and Reed,R. (1993) A functional association between the 5′ and 3′ splice sites is established in the earliest prespliceosome complex (E) in mammals. Genes Dev., 7, 1008–1020. [DOI] [PubMed] [Google Scholar]
- 13.Abovich N. and Rosbash,M. (1997) Cross-intron bridging interactions in the yeast commitment complex are conserved in mammals. Cell, 89, 403–412. [DOI] [PubMed] [Google Scholar]
- 14.Seraphin B. and Rosbash,M. (1990) Exon mutations uncouple 5′ splice site selection from U1 snRNA pairing. Cell, 63, 619–629. [DOI] [PubMed] [Google Scholar]
- 15.Michaud S. and Reed,R. (1991) An ATP-independent complex commits pre-mRNA to the mammalian spliceosome assembly pathway. Genes Dev., 5, 2534–2546. [DOI] [PubMed] [Google Scholar]
- 16.Liao X.C., Colot,H.V., Wang,Y. and Rosbash,M. (1992) Requirements for U2 snRNP addition to yeast pre-mRNA. Nucleic Acids Res., 20, 4237–4245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Champion-Arnaud P. Gozani,O., Palandjian,L. and Reed,R. (1995) Accumulation of a novel spliceosomal complex on pre-mRNAs containing branch site mutations. Mol. Cell. Biol., 15, 5750–5756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gozani O., Feld,R. and Reed,R. (1996) Evidence that sequence-independent binding of highly conserved U2 snRNP proteins upstream of the branch site is required for assembly of spliceosomal complex A. Genes Dev., 10, 233–243. [DOI] [PubMed] [Google Scholar]
- 19.Konarska M.M. and Sharp,P.A. (1986) Electrophoretic separation of complexes involved in the splicing of precursors to mRNAs. Cell, 46, 845–855. [DOI] [PubMed] [Google Scholar]
- 20.Konarska M.M. and Sharp,P.A. (1987) Interactions between small nuclear ribonucleoprotein particles in formation of spliceosomes. Cell, 49, 763–774. [DOI] [PubMed] [Google Scholar]
- 21.Konforti B.B., Koziolkiewicz,M.J. and Konarska,M.M. (1993) Disruption of base pairing between the 5′ splice site and the 5′ end of U1 snRNA is required for spliceosome assembly. Cell, 75, 863–873. [DOI] [PubMed] [Google Scholar]
- 22.Konforti B.B. and Konarska,M.M. (1995) A short 5′ splice site RNA oligo can participate in both steps of splicing in mammalian extracts. RNA, 1, 815–827. [PMC free article] [PubMed] [Google Scholar]
- 23.Maroney P.A., Romfo,C.M. and Nilsen,T.W. (2000) Functional recognition of 5′ splice site by U4/U6.U5 tri-snRNP defines a novel ATP-dependent step in early spliceosome assembly. Mol. Cell ., 6, 317–328. [DOI] [PubMed] [Google Scholar]
- 24.Burge C.B., Tuschl,T. and Sharp,P.A. (1999) Splicing of precursors to mRNAs by the spliceosomes. In Gesteland,R.F., Cech,T.R. and Atkins,J.F. (eds), The RNA World II. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
- 25.Ares M.,Jr (1986) U2 RNA from yeast is unexpectedly large and contains homology to vertebrate U4, U5 and U6 small nuclear RNAs. Cell, 47, 49–59. [DOI] [PubMed] [Google Scholar]
- 26.Parker R., Siliciano,P.G. and Guthrie,C. (1987) Recognition of the TACTAAC box during mRNA splicing in yeast involves base pairing to the U2-like snRNA. Cell, 49, 229–239. [DOI] [PubMed] [Google Scholar]
- 27.Wu J. and Manley,J.L. (1989) Mammalian pre-mRNA branch site selection by U2 snRNP involves base-pairing. Genes Dev., 3, 1553–1561. [DOI] [PubMed] [Google Scholar]
- 28.Zhuang Y. and Weiner,A.M. (1989) A compensatory base change in human U2 snRNA suppresses a globin branch site mutation. Genes Dev., 3, 1545–1552. [DOI] [PubMed] [Google Scholar]
- 29.Query C.C., Moore,M.J. and Sharp,P.A. (1994) Branch nucleophile selection in pre-mRNA splicing: evidence for the bulged duplex model. Genes Dev., 8, 587–597. [DOI] [PubMed] [Google Scholar]
- 30.Pascolo E. and Seraphin,B. (1997) The branchpoint residue is recognized during commitment complex formation before being bulged out of the U2 snRNA-pre-mRNA duplex. Mol. Cell. Biol., 17, 3469–3476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Berglund J.A., Abovich,N. and Rosbash,M. (1998) A cooperative interaction between U2AF65 and mBBP/SF1 facilitates branchpoint region recognition. Genes Dev., 12, 858–867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Arning S., Gruter,P., Bilbe,G. and Krämer,A. (1996) Mammalian splicing factor SF1 is encoded by variant cDNAs and binds to RNA. RNA, 2, 794–810. [PMC free article] [PubMed] [Google Scholar]
- 33.Berglund J.A., Chua,K., Abovich,N., Reed,R. and Rosbash,M. (1997) The splicing factor BBP interacts specifically with the pre-mRNA branchpoint sequence UACUAAC. Cell, 89, 781–787. [DOI] [PubMed] [Google Scholar]
- 34.Das R., Zhou,Z. and Reed,R. (2000) Functional association of U2 snRNP with the ATP-independent spliceosomal complex E. Mol. Cell ., 5, 779–787. [DOI] [PubMed] [Google Scholar]
- 35.Seraphin B. and Rosbash,M. (1991) The yeast branchpoint sequence is not required for the formation of stable U1 snRNA-pre-mRNA complex and is recognized in the absence of U2 snRNA. EMBO J., 10, 1209–1216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Abovich N., Liao,X.C. and Rosbash,M. (1994) The yeast MUD2 protein: an interaction with PRP11 defines a bridge between commitment complexes and U2 snRNP addition. Genes Dev., 8, 843–854. [DOI] [PubMed] [Google Scholar]
- 37.Query C.C., McCaw,P.S. and Sharp,P.A. (1997) A minimal spliceosomal complex A recognizes the branch site and polypyrimidine tract. Mol. Cell. Biol., 17, 2944–2953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Perriman R. and Ares,M.,Jr (2000) ATP can be dispensable for prespliceosome formation in yeast. Genes Dev., 14, 97–107. [PMC free article] [PubMed] [Google Scholar]
- 39.Yan D. and Ares,M.,Jr (1996) Invariant U2 RNA sequences bordering the branchpoint recognition region are essential for interaction with yeast SF3a and SF3b subunits. Mol. Cell. Biol., 16, 818–828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ruskin B., Zamore,P.D. and Green,M.R. (1988) A factor, U2AF, is required for U2 snRNP binding and splicing complex assembly. Cell, 52, 207–219. [DOI] [PubMed] [Google Scholar]
- 41.Zamore P.D. and Green,M.R. (1989) Identification, purification, and biochemical characterization of U2 small nuclear ribonucleoprotein auxiliary factor. Proc. Natl Acad. Sci. USA, 86, 9243–9247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Behrens S.E., Tyc,K., Kastner,B., Reichelt,J. and Lührmann,R. (1993) Small nuclear ribonucleoprotein (RNP) U2 contains numerous additional proteins and has a bipartite RNP structure under splicing conditions. Mol. Cell. Biol., 13, 307–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Brosi R., Groning,K., Behrens,S.E., Lührmann,R. and Krämer,A. (1993) Interaction of mammalian splicing factor SF3a with U2 snRNP and relation of its 60-kD subunit to yeast PRP9. Science, 262,102–105. [DOI] [PubMed] [Google Scholar]
- 44.Krämer A. (1996) The structure and function of proteins involved in mammalian pre-mRNA splicing. Annu. Rev. Biochem., 65, 367–409. [DOI] [PubMed] [Google Scholar]
- 45.Bennett M. and Reed,R. (1993) Correspondence between a mammalian spliceosome component and an essential yeast splicing factor. Science, 262,105–108. [DOI] [PubMed] [Google Scholar]
- 46.Legrain P. and Chapon,C. (1993) Interaction between PRP11 and SPP91 yeast splicing factors and characterization of a PRP9–PRP11–SPP91 complex. Science, 262, 108–110. [DOI] [PubMed] [Google Scholar]
- 47.Behrens S.E., Galisson,F., Legrain,P. and Lührmann,R. (1993) Evidence that the 60-kDa protein of 17S U2 small nuclear ribonucleoprotein is immunologically and functionally related to the yeast PRP9 splicing factor and is required for the efficient formation of prespliceosomes. Proc. Natl Acad. Sci. USA, 90, 8229–8233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Brosi R., Hauri,H.P. and Krämer,A. (1993) Separation of splicing factor SF3 into two components and purification of SF3a activity. J. Biol. Chem., 268, 17640–17646. [PubMed] [Google Scholar]
- 49.Wells S.E. and Ares,M.,Jr (1994) Interactions between highly conserved U2 small nuclear RNA structures and Prp5p, Prp9p, Prp11p, and Prp21p proteins are required to ensure integrity of the U2 small nuclear ribonucleoprotein in Saccharomyces cerevisiae. Mol. Cell. Biol., 14, 6337–6349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hausner T.P., Giglio,L.M. and Weiner,A.M. (1990) Evidence for base pairing between mammalian U2 and U6 small nuclear ribonucleoprotein particles. Genes Dev., 4, 2146–2156. [DOI] [PubMed] [Google Scholar]
- 51.Datta B. and Weiner,A.M. (1991) Genetic evidence for base pairing between U2 and U6 snRNAs in mammalian mRNA splicing. Nature, 352, 821–824. [DOI] [PubMed] [Google Scholar]
- 52.Wu J. and Manley,J.L. (1991) Base-pairing between U2 and U6 snRNAs is necessary for splicing of a mammalian pre-mRNA. Nature, 352, 818–821. [DOI] [PubMed] [Google Scholar]
- 53.Field D.J. and Friesen,J.D. (1996) Functionally redundant interactions between U2 and U6 spliceosomal snRNAs. Genes Dev., 10, 489–501. [DOI] [PubMed] [Google Scholar]
- 54.Madhani H.D. and Guthrie,C. (1992) A novel base-pairing interaction between U2 and U6 snRNAs suggests a mechanism for the catalytic activation of the spliceosome. Cell, 71, 803–817. [DOI] [PubMed] [Google Scholar]
- 55.Madhani H.D. and Guthrie,C. (1994) Dynamic RNA–RNA interactions in the spliceosome. Annu. Rev. Genet., 28, 1–26. [DOI] [PubMed] [Google Scholar]
- 56.Sun J.-S. and Manley,J.L. (1995) A novel U2–U6 snRNA structure is necessary for mammalian mRNA splicing. Genes Dev., 9, 843–854. [DOI] [PubMed] [Google Scholar]
- 57.Luukkonen B.G. and Seraphin,B. (1998) A role for U2/U6 helix Ib in 5′ splice site selection. RNA, 4, 915–927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Li Z. and Brow,D.A. (1996) A spontaneous duplication in U6 spliceosomal RNA uncouples the early and late functions of the ACAGA element in vivo. RNA, 2, 879–894. [PMC free article] [PubMed] [Google Scholar]
- 59.Zavanelli M.I. and Ares,M.,Jr (1991) Efficient association of U2 snRNPs with pre-mRNA requires an essential U2 RNA structural element. Genes Dev., 5, 2521–2533. [DOI] [PubMed] [Google Scholar]
- 60.Yan D., Perriman,R., Igel,H., Howe,K.J., Neville,M. and Ares,M.,Jr (1998) CUS2, a yeast homolog of human Tat-SF1, rescues function of misfolded U2 through an unusual RNA recognition motif. Mol. Cell. Biol., 18, 5000–5009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ruby S.W., Chang,T.-H. and Abelson,J. (1993) Four yeast spliceosomal proteins (PRP5, PRP9, PRP11, and PRP21) interact to promote U2 snRNP binding to pre-mRNA. Genes Dev., 7, 1909–1925. [DOI] [PubMed] [Google Scholar]
- 62.Wells S.E., Neville,M., Haynes,M., Wang,J., Igel,H. and Ares,M.,Jr (1996) CUS1, a suppressor of cold-sensitive U2 snRNA mutations, is a novel yeast splicing factor homologous to human SAP 145. Genes Dev., 10, 220–232. [DOI] [PubMed] [Google Scholar]
- 63.Solnick D. (1985) Trans splicing of mRNA precursors. Cell, 42, 157–164. [DOI] [PubMed] [Google Scholar]
- 64.Barettino D., Feigenbutz,M., Valcarcel,R. and Stunnenberg,H.G. (1994) Improved method for PCR-mediated site-directed mutagenesis. Nucleic Acids Res., 22, 541–542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wassarman D.A. and Steitz,J.A. (1992) Interactions of small nuclear RNAs with precursor messenger RNA during in vitro splicing. Science, 257, 1918–1925. [DOI] [PubMed] [Google Scholar]
- 66.Wassarman D.A. (1993) Psoralen crosslinking of small RNAs in vitro. Mol. Biol. Rep., 17, 143–151. [DOI] [PubMed] [Google Scholar]
- 67.Ast G. and Weiner,A.M. (1996) A U1/U4/U5 snRNP complex induced by a 2′-O-methyl-oligonucleotide complementary to U5 snRNA. Science, 272, 881–884. [DOI] [PubMed] [Google Scholar]
- 68.Ast G. and Weiner,A.M. (1997) A novel U1/U5 interaction suggests that U1 directs U5 small nuclear RNA to the 5′ splice site. RNA, 3, 371–381.9085844 [Google Scholar]
- 69.Ast G. and Weiner,A.M. (1997) Antisense oligonucleotide binding to U5 snRNP induces a conformational change that exposes the conserved loop of U5 snRNA. Nucleic Acids Res., 25, 3508–3513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Tarn W.-Y. and Steitz,J.A. (1995) Modulation of 5′ splice site choice in pre-messenger RNA by two distinct steps. Proc. Natl Acad. Sci. USA, 92, 2504–2508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Konforti B.B. and Konarska,M.M. (1994) U4/U5/U6 snRNP recognizes the 5′ splice site in the absence of U2 snRNP. Genes Dev., 8, 1962–1973. [DOI] [PubMed] [Google Scholar]
- 72.Dominski Z. and Kole,R. (1994) Identification and characterization by antisense oligonucleotides of exon and intron sequences required for splicing. Mol. Cell. Biol., 14, 7445–7454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Krämer A., Gruter,P., Groning,K. and Kastner,B. (1999) Combined biochemical and electron microscopic analyses reveal the architecture of the mammalian U2 snRNP. J. Cell Biol., 145, 1355–1368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ares M.,Jr and Igel,A.H. (1990) Lethal and temperature sensitive mutations and their suppressors identify an essential structural element in U2 small nuclear RNA. Genes Dev., 4, 2132–2145. [DOI] [PubMed] [Google Scholar]
- 75.Hong W., Bennett,M., Xiao,Y., Kramer,R.F., Wang,C. and Reed,R. (1997) Association of U2 snRNP with the spliceosomal complex E. Nucleic Acids Res., 25, 354–361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Chiara M.D. and Reed,R. (1995) A two-step mechanism for 5′ and 3′ splice-site pairing. Nature, 375, 510–513. [DOI] [PubMed] [Google Scholar]
- 77.Bruzik J.P. and Maniatis,T. (1995) Enhancer-dependent interaction between 5′ and 3′ splice sites in trans. Proc. Natl Acad. Sci. USA, 92, 7056–7059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Daugeron M.-C., Tazi,J., Jeanteur,P., Brunel,C. and Cathala,G. (1992) U1–U2 snRNPs interaction induced by an RNA complementary to the 5′ end sequence of U1 snRNA. Nucleic Acids Res., 20, 3625–3630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Fu X.-Y. and Manley,J.L. (1987) Factors influencing alternative splice site utilization in vivo. Mol. Cell. Biol., 7, 738–748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Newman A.J. and Norman,C. (1991) Mutations in yeast U5 snRNA alter the specificity of 5′ splice site cleavage. Cell, 65, 115–123. [DOI] [PubMed] [Google Scholar]
- 81.Newman A.J. and Norman,C. (1992) U5 snRNA interacts with exon sequences at 5′ and 3′ splice sites. Cell, 68, 743–753. [DOI] [PubMed] [Google Scholar]
- 82.Wyatt J.R., Sontheimer,E.J. and Steitz,J.A. (1992) Site-specific cross-linking of mammalian U5 snRNP to the 5′ splice site before the first step of pre-mRNA splicing. Genes Dev., 6, 2542–2553. [DOI] [PubMed] [Google Scholar]
- 83.Cortes J.J., Sontheimer,E.J., Seiwert,S.D. and Steitz,J.A. (1993) Mutations in the conserved loop of human U5 snRNA generate use of novel cryptic 5′ splice sites in vivo. EMBO J., 12, 5181–5189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sontheimer E.J. and Steitz,J.A. (1993) The U5 and U6 snRNAs as active site components of spliceosome. Science, 262, 1989–1996. [DOI] [PubMed] [Google Scholar]
- 85.Newman A.J., Teigelkamp,S. and Beggs,J.D. (1995) snRNA interactions at the 5′ and 3′ splice sites monitored by photoactivated crosslinking in yeast spliceosomes. RNA, 1, 968–980. [PMC free article] [PubMed] [Google Scholar]
- 86.Segault V., Will,C.L., Polycarpou-Schwarz,M., Mattaj,I.W., Branlant,C. and Lührmann,R. (1999) Conserved loop I of U5 small nuclear RNA is dispensable for both catalytic steps of pre-mRNA splicing in HeLa nuclear extract. Mol. Cell. Biol., 19, 2782–2790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.O’Keefe R.T., Norman,C. and Newman,A.J. (1996) The invariant U5 snRNA loop 1 sequence is dispensable for the first catalytic step of pre-mRNA splicing in yeast. Cell, 86, 679–689. [DOI] [PubMed] [Google Scholar]
- 88.Valadkhan S. and Manley,J.L. (2000) A tertiary interaction detected in a human U2-U6 snRNA complex assembled in vitro resembles a genetically proven interaction in yeast. RNA, 6, 206–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ruskin B., Krainer,A.R., Maniatis,T. and Green,M.R. (1984) Excision of an intact intron as a novel lariat structure during pre-mRNA splicing in vitro. Cell, 38, 317–331. [DOI] [PubMed] [Google Scholar]
- 90.Reed R. (1989) The organization of 3′ splice-site sequences in mammalian introns. Genes Dev., 3, 2113–2123. [DOI] [PubMed] [Google Scholar]
- 91.Krainer A.R., Maniatis,T., Ruskin,B. and Green,M.R. (1984) Normal and mutant human β-globin pre-mRNAs are faithfully and efficiently spliced in vitro. Cell, 36, 993–1005. [DOI] [PubMed] [Google Scholar]
- 92.Noble J.C., Ge,H., Chaudhuri,M. and Manley,J.L. (1989) Factor interactions with the simian virus 40 early pre-mRNA influence branch site selection and alternative splicing. Mol. Cell. Biol., 9, 2007–2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Anderson K. and Moore,M.J. (1997) Bimolecular exon ligation by the human spliceosome. Science, 276, 1712–1716. [DOI] [PubMed] [Google Scholar]