

Abstract
Background:
Subarachnoid hemorrhage (SAH) is associated with severe vasospasm caused by a variety of neurochemical mechanisms. The anterior choroid arteries (AChAs) are innervated by vasodilated fibers of the trigeminal ganglion (TGG). The goal of this study was to determine whether there is a relationship between the neuron density of the TGG and the severity of AChAs vasospasm with SAH.
Methods:
Thirty-two rabbits were used for the study; eight served as the baseline control group, seven as a SHAM group, with injections of 1 cc of isotonic saline solution, and 17 rabbits were included in the experimental SAH group, with injection of homologous blood into the cisterna magna. After 10 days, the histopathology of the AChAs and TGGs were examined. The AChAs vasospasm index (VSI) of the external/internal diameter and the neuron density of the ophthalmic root of the TGGs were evaluated stereologically. The AChAs VSI was preferred -- a measure of the degree of vasospasm. As the VSI increased, the degree of arterial vasospasm increased. The results were statistically analyzed.
Results:
The mean AChAs VSI was significantly higher and the mean neuronal density of the ophthalmic root of the TGG was significantly lower in the group with severe vasospasm associated with SAH compared to the controls, SHAM, and the group with mild vasospasm associated with SAH (P< 0.05). The ophthalmic root of the TGG neuron density in the 7 rabbits that developed severe vasospasm was statistically less than that observed in the 10 rabbits with mild vasospasm. There was a linear relationship between the low neuronal density in the ophthalmic root of the TGG and the severity of the AChA vasospasm.
Conclusions:
The trigeminal ganglion neuron density may be an important factor in the regulation of AChAs diameter and cerebral blood flow. Low neuron density of the ophthalmic root of the TGG may play a role in the pathogenesis of AChAs vasospasm associated with SAH.
Keywords: Anterior choroid artery, neuron density, subarachnoid hemorrhage, trigeminal ganglion, vasospasm, vasospasm index
INTRODUCTION
Cerebral vasospasm is a serious complication of subarachnoid hemorrhage (SAH); delayed narrowing of the large capacity arteries and cerebral vasospasm are associated with significant morbidity and mortality following a SAH.[6] The anterior choroid artery (AChA) is the most important branch of the supraclinoid segment of the ICA. It supplies the optic tract, uncus, lateral part of the geniculate body, posterior two-thirds of the posterior limbs of the internal capsule, optic radiation, and choroid plexus of the lateral ventricles. The classical features of AChA occlusion include contralateral hemiplegia, hemianesthesia and hemianopsia. Vasospasm of the choroid arteries may be severe when a SAH causes damage to the choroid plexus. This can affect structures that play an important role in immune, endocrine, detoxifying, thermoregulatory, and secretory functions of the brain; such abnormalities are associated with a poor prognosis.[10,23,25,30,31] The response of AChAs, in a rabbit model of SAH, was evaluated in this study.
The cerebral arteries are innervated by several systems that contribute to the autonomic control of cerebral blood flow. Parasympathetic fibers affect vasodilation and sympathetic fibers cause vasospasm of the cerebral arteries.[16] Cerebrovascular sensory nerves originate mainly from the first division of ipsilateral trigeminal ganglion (TGG), known as the trigemino-cerebrovascular system.[2,19,32] Trigeminal sensory nerves project to the ipsilateral ICA, MCA, ACA, the rostral part of the basilar artery, the PCA, and the AChA.[1]
Neuro-humoral mechanisms have been suggested as important factors associated with the cerebral artery vasospasm after a subarachnoid hemorrhage.[5,6,22,29] The goal of this study was to determine the relationship between the number of neurons in the ophthalmic root of the TGG and changes in the diameter of the AChA, associated with SAH-induced vasospasm.
MATERIALS AND METHODS
Thirty-two anesthetized, adult male New Zealand rabbits were used for this study. The Ethics Committee of Atatürk University, Medical Faculty, approved the animal protocols. Animal care and experimental protocols were conducted according to the guidelines set forth by the same ethics committee. The animals were randomly divided into three groups: SAH (n=17), isotonic saline solution (SHAM; n=7), and control (n=8) groups. A balanced, injectable anesthetic was used to reduce pain and mortality. Anesthesia was induced with isoflurane, given by a facemask, and 0.2 mL/kg of the anesthetic combination of Ketamine HCL, 150 mg/1.5 mL; Xylazine HCL, 30 mg/1.5 mL; and distilled water, 1 mL, was subcutaneously injected prior to the surgery. All animals were monitored for changes in the electrocardiogram, respiration patterns, and blood oxygen concentration during the experiment. All parameters were recorded by a camera and analyzed by physicians that did not know to which experimental group the individual animals belonged. During the procedure, a dose of 0.1 mL/kg of the anesthetic combination was used when required. In 17 (n=17) of the animals, autologous blood (0.5 mL) was taken from the auricular artery. While the head of the animal was held in a hyperflexed position, the posterior notch of the foramen magnum was identified, and the cisterna magna was entered; CSF was then aspirated. When the identification of the cisterna magna was confirmed, blood was injected using a 22-gauge needle, over about 1 minute, in the SAH group; 1 mL of isotonic saline solution was injected in the same way in the seven (n=7) animals of the SHAM group. The remaining eight (n=8) animals not subjected to this procedure were considered the control group. The animals were followed for 10 days without any medical treatment and then killed. The time was selected based on relief of the vasospasm. The number of ganglion neurons is thought to be associated with vasospasm. The effects of SAH on vasospasm were studied by the removal of all AChAs and trigeminal ganglia, bilaterally, for histological examination. Specimens were stored in a 10% formalin solution for 7 days, after which 5 μm tissue sections were cut and stained with hematoxylin and eosin.
To estimate the neuron density of the ophthalmic division of the trigeminal nerve, all trigeminal roots, together with their ganglions, were extracted bilaterally. The specimens were then horizontally embedded in paraffin blocks to evaluate all roots during the histopathological examination. The physical dissector method[27] was used to evaluate the number of neurons in the TGG. The advantages of this method are that it easily estimates particle number; can be readily performed; is intuitively simple; is free from assumptions about particle shape, size, and orientation; and is unaffected by overprotection and truncation. Two consecutive sections (dissector pairs) obtained from tissue samples with named references were mounted on each slide. The reference and look-up sections were reversed to double the number of dissector pairs; this eliminated the need for obtaining new sections. The mean neuronal density of the ophthalmic root of the TGG (NvGN) per mm3 was estimated using the following formula: NvGN = ∑Q – N/t × A, where ∑Q – N is the total number of counted neurons appearing only in the reference sections, t is the section thickness, and A and B are the areas of the counting frames [Figure 1a,b]. The Cavalieri volume estimation method was used to obtain the total number of neurons in each specimen.[8,12,27] The total number of neurons was calculated by multiplying the volume (mm3) and the numerical density of neurons in each TGG. In addition, the mean number of neurons in the ophthalmic division of the TGG was counted bilaterally in every animal [Table 1].
Figure 1.
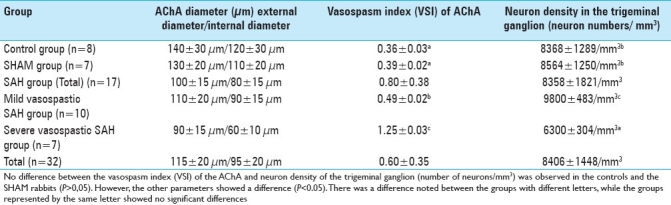
(a, b) Stereologic cell counts of the ophthalmic division of the TGG in a rabbit. Use of the physical dissector method where the micrographs of the same fields of view (a, b) were taken from two parallel, adjacent thin sections separated by 5 μm. The numerical density of the neurons was calculated as NvGN= ∑Q – N/t × A. In this application, the nucleoli marked as ‘2,3,4,8’ are dissector particles in A. Section B shows them as they disappeared. The nucleoli marked as ‘1,5,6,7’ do not represent dissector particles in A. Section B shows ‘1 and 8’ as they disappeared (H and E, 100, LM)
Table 1.
Vasospasm index of the AChAs and the neuronal density of the ophthalmic root of the trigeminal ganglion

The AChAs were obtained from the coronal brain sections at the level of the AChAs entering into the lateral ventricles. They were also stained with H&E. For the calculation of the vasospasm index of the AChAs, all AChAs were accepted as a cylinder, in view of their morphological characteristics; simple geometric formulas were used to estimate their wall ring values. As a measure of the degree of vasospasm, the use of the AChAs vasospasm index was preferred to use of only measurements of the lumen radius; it can be readily performed, is intuitively simple, more reliable, free from assumptions about vessel diameter of various segments and is unaffected by overestimation errors of the radius of the AChAs. The AChAs of all animals were cut 20 segments away from the point where the internal carotid arteries entered the choroid plexus. Then, 20 histopathology sections, 5 μm apart, were obtained, using a microtome, for each designation, and are represented by the lines 1, 2, 3,… and 20. The average of 10 diameters, of 10 cross-sectional areas, was recorded as the mean diameter. A single line in the figures represents one of them. The mean external and internal (luminal) diameters of each section was measured; the external radius is represented by R1 and the internal radius is represented by r1. The mean external radius of the anterior choroid arteries was calculated as R1 = R1 + R2 + R3 +….R20/20; the lumen radius was calculated as r1 = r1 + r2 + r3 +….r20/20 [Figure 2]. The wall ring surface values were calculated using the following formula: S1 = πR12 – πr12. The lumen surface area was calculated using the same method. Therefore, the lumen surface value (S2) = πr12. The vasospasm index (VI) was calculated as the proportion of S1/S2 . The vasospasm index (VI) =S1/S2 = πR12 – πr12/πr12 = π(R12 – r12)/πr12 = R12 – r12/r12.
Figure 2.
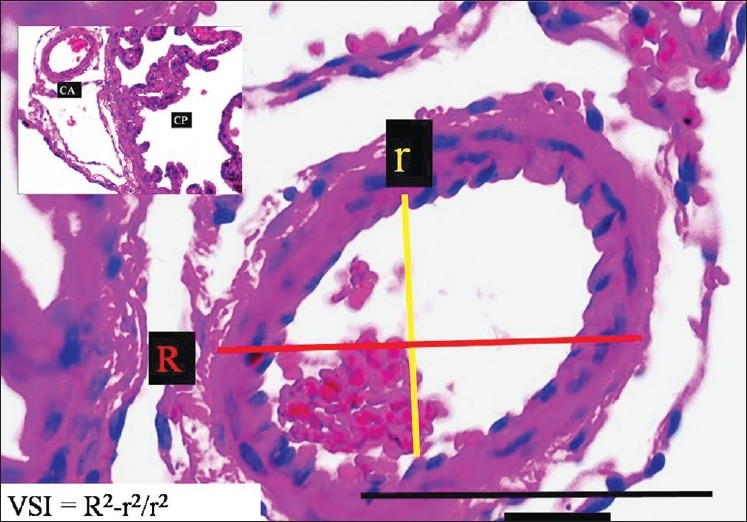
The histopathological appearance of the right AChA at the entrance level in a normal rabbit (LM, H and E, ×100). On the left side, minimal forms of CP are observed (LM, H and E, ×20)
In summary, VI= (R12 – r12)/r12. In addition, the vasospasm index of AChAs in the rabbits with SAH was calculated as the same manner; VI= (R22 – r22)/r22 . As the VSI increases, the degree of arterial vasospasm also increases. Vasospasm was defined as mild with a reduction of 25% of the normal diameter of any arterial segment of AChA and as severe vasospasm with a decrease of 40%.[13,14]
The differences between the VSI of AChAs and neuron density of the ophthalmic root of the TGG were compared statistically. For the statistical analysis, SPSS ver. 15.0 was used. The mean ± standard deviation of the variables is reported. Since the data showed a normal distribution, intergroup differences were assessed using a one-way ANOVA. The presence of homogeneous variance necessitated the use of the Tukey test for comparisons between two groups. A P<0.05 was accepted as statistically significant.
RESULTS
Two of the animals (n=2) died within the first week and the remaining animals (n=30) were followed for 10 days and then killed. The following clinical findings were frequently observed during the premortem period of the dead animals and the five living animals: signs of meningeal irritation, consciousness, seizures, fever, apnea, cardiac arrhythmia, and breathing disturbances.
TGGs were identified from the trigeminal impressions located on the upper surface of the petrous bones. They were fusiform shaped, and the volumes were estimated to be 2 × 1.5 × 2 mm3 . All trigeminal nerves have three main branches: the ophthalmic, mandibular, and maxillary. The neuron density of only the ophthalmic root division of the TGGs was examined [Figure 1a,b].
Morphological examinations of the brains showed the AChAs at the cisternal segment, extending from its origin to the choroid fissure, and the plexus segment, extending from the choroid fissure to the area where it enters into the choroid plexus of the lateral ventricles. The mean diameter of the AChA examined was 0.35±0.10 mm. The AChA convolutions were more prominent in the animals with severe vasospasm associated with SAH compared to the SHAM, control, and mild vasospasm associated with SAH groups.
To estimate the AChA volume, square-lined glass plates were used and photographs were taken under the microscope during histopathological examinations. The inner elastic membrane (IEM) was less convoluted and the luminal surface area was greater in the SHAM, control [Figure 2], and mild vasospasm associated SAH groups [Figure 3]. The following features were observed specifically in the severe vasospasm associated SAH group: arachnoiditis, pia-arachnoid adhesions, AChA narrowing, IEM convolutions, intimal edema formation and endothelial cell shrinkage, desquamation, and loss [Figure 4].
Figure 3.
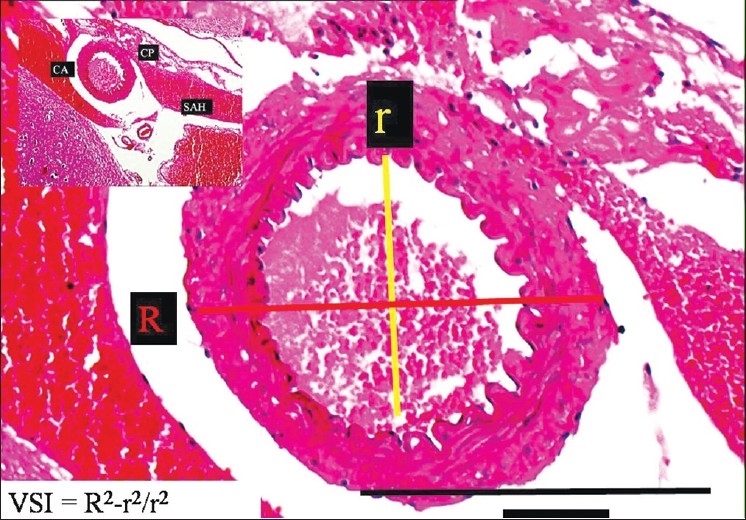
The histopathological appearance of the right AChA at the postorigin level in a SAH rabbit model (LM, H and E, ×100). On the left side, minimal forms of CP are observed (LM, H and E, ×20) (mild vasospasm associated SAH group)
Figure 4.
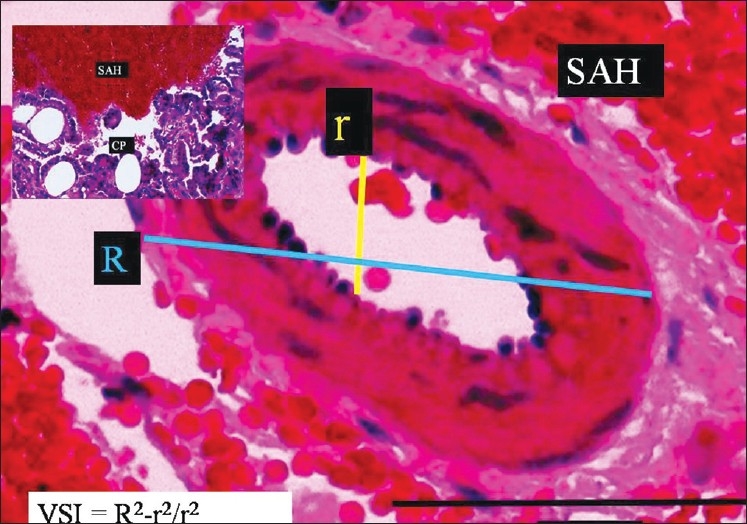
The histopathological appearance of the right AChA at the postorigin level in a SAH rabbit model (LM, H and E, ×100). On the left side, minimal forms of CP are observed (LM, H and E, ×20) (severe vasospasm associated SAH group)
The mean external diameter/internal diameter of the AChAs using the segmental model was estimated as 115±20 μm/95±20 μm; the mean vasospasm index of the AChAs was 0.46; the mean neuronal density of the ophthalmic root of the TGGs was 8290±1480 neurons/mm3 for all animals (n=32). The mean external diameter/internal diameter of the AChAs was estimated as 140±30 μm/120±30 μm, and the mean vasospasm index of the AChAs was 0.36. The mean neuronal density of the ophthalmic root of the TGGs was 8350±390 neurons/mm 3 in the control group (n=8). The mean external diameter/internal diameter of the AChAs was estimated as 130±20 μm/110±20 μm, and the mean vasospasm index of the AChAs was 0.39. The mean neuronal density of the ophthalmic root of the TGGs was 8550±650 neurons/mm 3 in the SHAM group (n=7). The mean external diameter/internal diameter of the AChAs was estimated as 100±15 μm/80±15 μm, and the mean vasospasm index of the AChAs was 0.56. The mean neuronal density of the ophthalmic root of the TGGs was 8200±600 neurons/mm3 in the SAH group (n=17). Severe vasospasms were observed in seven rabbits with SAH, and mild vasospasms were observed in the remaining 10 rabbits with SAH. The mean external diameter/internal diameter of the AChAs was estimated as 110±20 μm/90±15 μm, and the mean vasospasm index of the AChAs was 0.49. The mean neuronal density of the ophthalmic root of the TGGs was 9800±724 neurons/mm3 for the mild vasospasm with the SAH group (n=10). However, the mean external diameter/internal diameter of the AChAs was estimated as 90±15 μm/60±10 μm, and the mean vasospasm index of the AChAs was 1.25. The mean neuronal density of the ophthalmic root of the TGGs was 6342±557 neurons/mm3 in the severe vasospasm with the SAH group (n=7),
The estimated mean AChAs VSI was significantly higher and the mean neuronal density of the ophthalmic root of the TGG was significantly lower in the severe vasospasm associated with the SAH group when compared to the control, SHAM, and mild vasospasm associated with SAH groups (P< 0.05) [Table 1]. The ophthalmic root of the TGG neuron density in the seven rabbits that developed severe vasospasm was significantly less than observed in the 10 rabbits with mild vasospasm. The results showed an invers relationship between low neuron density in the ophthalmic root of the TGG and severity of the AChA vasospasm [Figure 5].
Figure 5.
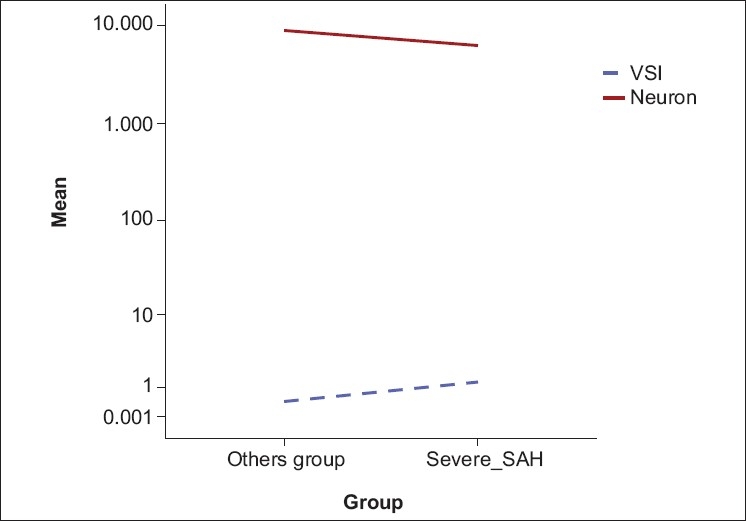
There was an inverse correlation between low neuron density in the TGG and the severity of the AChA vasospasm (r=-0.55, <0.05, Pearson correlation analysis)
DISCUSSION
Vasospasm is pathophysiologically characterized by narrowing of the vascular lumen; this may develop following a SAH. Vasospasm after a SAH is associated with significant morbidity and mortality.[7,16,17] It is widely believed that blood, or its degradation products, causes vasospasm by directly affecting the walls of the cerebral arteries.[29] The pathogenesis of the arterial narrowing that occurs following a SAH is not fully understood.
Numerous neuronal, humoral, and chemical factors are involved in cerebrovascular innervations. Parasympathetic fibers have vasodilation effects, and sympathetic fibers have vasospastic effects on cerebral arteries. A variety of autonomic nerve fibers provide neural innervation to cerebral vascular structures. The postganglionic fibers, of the ciliary ganglion, of the third cranial nerve, the sphenopalatine ganglion of the seventh cranial nerve, the otic ganglion of the ninth cranial nerve, and the ganglion of the fifth cranial nerve, are involved in the parasympathetic outflow, which causes vasodilation of the cerebral arteries.[16,20] The sympathetic innervations, from the postganglionic fibers of the superior sympathetic ganglion, which contribute to the carotid plexus, cause vasospasm of the cerebral vessels.[14]
The extracerebral and intracerebral cranial circulations are innervated by trigeminal nerve fibers. The branches of the ophthalmic division of the trigeminal nerve provide the primary innervations of the large cerebral arteries. The trigeminal nerve fibers release transmitters that cause vasodilation of the cerebral arteries. Direct stimulation of the trigeminal nerve results in ipsilateral cerebral vasodilation after a SAH.[9,11,28] Unilateral postganglionic trigeminal lesions result in ipsilateral constriction of the cerebral arteries and consequent decrease of the CBF; however, pre-ganglionic lesions do not affect the baseline diameter of cerebral arteries.[26,29]
Vasoactive neurotransmitters, neuromodulators, and neurochemicals are released from trigeminal nerve terminals; these compounds affect large intracranial and extracranial blood vessels.[20] Aydin et al.[3] reported that decreased neuron density in the petrosal ganglion was an etiologic factor associated with high blood pressure. Onder et al.[22] reported that decreased neuron density of the TGG was associated with a lower basilar artery volume and that a decreased number of neurons in the TGG resulted in a poorer prognosis of rabbits with a SAH. In their study, the animals that died in the SAH group had the lowest number of neurons in the TGG. In this study, two animals in the group with severe vasospasm associated with a SAH died, and the number of neurons in their TGG was significantly lower than in the controls, SHAM, and mild vasospasm associated SAH groups. In addition, Kanat et al.[15] reported that the neuron density of the C3 dorsal root ganglia (C3DRG) might be an important factor in the regulation of the anterior spinal artery (ASA) volume and spinal cord blood flow. The low neuron density of C3DRG was suggested to be an important factor associated with the pathogenesis of severe ASA vasospasm associated with a SAH.
The goal of this study was to investigate the potential relationship between the neuron density of the ophthalmic root of the TGG and the severity of AChA vasospasm in rabbits, following a SAH. The two major findings of this investigation were (i) a correlation between the neuron density of the ophthalmic root of the TGG and ACA volume, and (ii) the association of a low neuron density, of the TGG, with the pathogenesis of ACA vasospasm after a SAH. In order to evaluate the severity of AChA vasospasm associated with a SAH, the vasospasm index of the AChA was used as the preferred parameter for assessment rather than the radius of the vessel lumen. As the VSI increased, the degree of arterial vasospasm also increased.
The results of this study showed that the mean AChA VSI was higher in animals with lower neuron density in the trigeminal ganglia. A correlation was found between the neuron density of the ophthalmic division of the trigeminal ganglion and the degree of AChA vasospasm (P<0.05). In particular, the ophthalmic root of the TGG neuron density of the seven rabbits that developed severe vasospasm was significantly lower than in the 10 rabbits that had mild vasospasm. Therefore, a lower neuron density in the TGG might be associated with reduced vasodilation of the AChAs. Compounds associated with vasodilation are synthesized by TGG neurons and secreted from the nerve terminals that end in the cerebral arteries. A reduced number of neurons may result in deficient quantities of vasodilator molecules in the trigeminal ganglia and/or cerebrovascular sensory nerves, which affects the trigemino-cerebrovascular system and the AChAs, and might subsequently increase the severity of vasospasm in the AChAs after a SAH. Therefore, novel approaches to improve cerebral blood flow, under these conditions, are being investigated.
CONCLUSION
The number of neurons in the ophthalmic root of the TGG may play an important role in the regulation of cerebral artery vasospasm (AChAs as well) and, therefore, in the regulation of the cerebral circulation. In rabbits, a significant reduction of the number of the ophthalmic root of the TGG neurons, after a SAH, impaired the sensory innervation of cerebral arteries and the AChAs, and might play an important role in the development of cerebral vasospasm. The results of this study showed that the pathological processes involved in vasospasm, associated with a SAH, significantly affected the choroid arteries. In addition, neurological deficits, resulting in ischemic injury and apoptosis of the choroid plexus, were observed.
Footnotes
Available FREE in open access from: http://www.surgicalneurologyint.com/text.asp?2011/2/1/77/82084
Contributor Information
Adem Yilmaz, Email: ademyilmaz70@yahoo.com.
Cemal Gündoğdu, Email: cemgun@yahoo.com.
Mehmet Dumlu Aydin, Email: nmda11@hotmail.com.
Murat Musluman, Email: mmusluman@yahoo.com.
Ayhan Kanat, Email: ayhankanat@yahoo.com.
Yunus Aydin, Email: ynaydin@hotmail.com.
REFERENCES
- 1.Arbab MA, Wiklund L, Svendgaard NA. Origin and distribution of cerebral vascular innervation from superior cervical, trigeminal and spinal ganglia investigated with retrograde and anterograde WGA-HRP tracing in the rat. Neuroscience. 1986;19:695–708. doi: 10.1016/0306-4522(86)90293-9. [DOI] [PubMed] [Google Scholar]
- 2.Atalay B, Bolay H, Dalkara T, Soylemezoglu F, Oge K, Ozcan OE. Transcorneal stimulation of trigeminal nerve afferents to increase cerebral blood flow in rats with cerebral vasospasm: A noninvasive method to activate the trigeminovascular reflex. J Neurosurg. 2002;97:1179–83. doi: 10.3171/jns.2002.97.5.1179. [DOI] [PubMed] [Google Scholar]
- 3.Aydin MD, Bayram E, Atalay C, Aydin N, Erdogan AR, Gundogdu C, et al. The role of neuron numbers of the petrozal ganglion in the determinetion of blood pressure. An experimental study. Minim Invasive Neurosurg. 2006;49:359–61. doi: 10.1055/s-2006-955071. [DOI] [PubMed] [Google Scholar]
- 4.Bederson JB, Connolly ES, Jr, Batjer HH, Dacey RG, Dion JE, Diringer MN, et al. American Heart Association. Guidelines for the management of aneurysmal subarachnoid hemorrhage: A statement for healthcare professionals from a special writing group of the Stroke Council, American Heart Association. Stroke. 2009;40:994–1025. doi: 10.1161/STROKEAHA.108.191395. [DOI] [PubMed] [Google Scholar]
- 5.Bederson JB, Guarino L, Germano IM. Cortical blood flow and cerebral perfusion pressure in a new noncraniotomy model of subarachnoid hemorrhage in the rat. Stroke. 1995;26:1086–91. doi: 10.1161/01.str.26.6.1086. [DOI] [PubMed] [Google Scholar]
- 6.Cholet N, Bonvento G, Seylaz J. Effect of neuronal NO synthase inhibition on the cerebral vasodilatory response to somatosensory stimulation. Brain Res. 1996;708:197–200. doi: 10.1016/0006-8993(95)01387-3. [DOI] [PubMed] [Google Scholar]
- 7.Clozel M, Watanebe H. BQ-123, a peptidic endothelin -a receptor antagonist, prevents the early cerebral vasospazm following subarachnoid hemorrhage after intracisternal but not intravenous injection. Life Sci. 1993;52:825–34. doi: 10.1016/0024-3205(93)90081-d. [DOI] [PubMed] [Google Scholar]
- 8.Cruz-Orive LM, Weibel ER. Recent stereological methods for cell biology: A brief survey. Am J Physiol. 1990;258:L148–56. doi: 10.1152/ajplung.1990.258.4.L148. [DOI] [PubMed] [Google Scholar]
- 9.Edvinsson L, Mulder H, Goadsby PJ, Uddman R. Calcitonin gene-related peptide and nitric oxide in the trigeminal ganglion: Cerebral vasodilatation from trigeminal nerve stimulation involves mainly calcitonin gene-related peptide. J Auton Nerv Syst. 1998;70:15–22. doi: 10.1016/s0165-1838(98)00033-2. [DOI] [PubMed] [Google Scholar]
- 10.Ennis SR, Keep RF. The effects of cerebral ischemia on the rat choroid plexus. J Cereb Blood Flow Metab. 2006;26:675–83. doi: 10.1038/sj.jcbfm.9600224. [DOI] [PubMed] [Google Scholar]
- 11.Goadsby PJ, Knight YE, Hoskin KL, Butler P. Stimulation of an intracranial trigeminally-innervated structure selectively increases cerebral blood flow. Brain Res. 1997;751:247–52. doi: 10.1016/s0006-8993(96)01344-3. [DOI] [PubMed] [Google Scholar]
- 12.Gundersen HJ, Bendtsen TF, Korbo L, Marcussen N, Møller A, Nielsen K, et al. Some new, simple and efficient stereological methods and their use in pathological research and diagnosis. APMIS. 1988;96:379–94. doi: 10.1111/j.1699-0463.1988.tb05320.x. [DOI] [PubMed] [Google Scholar]
- 13.Hara H, Zhang QJ, Kuroyanagi T, Kobayashi S. Parasympathetic cerebrovascular innervation: An anterograde tracing from the sphenopalatine ganglion in the rat. Neurosurgery. 1993;32:822–7. doi: 10.1227/00006123-199305000-00016. [DOI] [PubMed] [Google Scholar]
- 14.Jakobsen M. Role of initial brain ischemia in subarachnoid hemorrhage following aneurysm rupture: A pathophysiological survey. Acta Neurol Scand. 1992;141:1–33. [PubMed] [Google Scholar]
- 15.Kanat A, Yilmaz A, Aydin MD, Musluman M, Altas S, Gursan N. Role of degenerated neuron density of dorsal root ganglion on anterior spinal artery vasospasm in subarachnoid hemorrhage: Experimental study. Acta Neurochir (Wien) 2010;152:2167–72. doi: 10.1007/s00701-010-0793-4. [DOI] [PubMed] [Google Scholar]
- 16.Kassell NF, Sasaki T, Colohan AR, Nazar G. Cerebral vasospasm following aneurysmal subaracnoid hemorrhage. Stroke. 1985;16:562–72. doi: 10.1161/01.str.16.4.562. [DOI] [PubMed] [Google Scholar]
- 17.Lehmann RM, Kassell NF, Nazar G. Morphometric method in the study of vasospasm. In: Wilkins RH, editor. Cerebral vasospasm. New York: Raven Press; 1998. p. 367. [Google Scholar]
- 18.Lindegaard KF, Nornes H, Bakke SJ, Sorteberg W, Nakstad P. Cerebral vasospasm diagnosis by means of angiography and blood velocity measurements. Acta Neurochir (Wien) 1989;100:12–24. doi: 10.1007/BF01405268. [DOI] [PubMed] [Google Scholar]
- 19.Liu-Chen LY, Mayberg MR, Moskowitz MA. Immunohistochemical evidence for a substance P-containing trigeminovascular pathway to pial arteries in cats. Brain Res. 1983;268:162–6. doi: 10.1016/0006-8993(83)90402-x. [DOI] [PubMed] [Google Scholar]
- 20.Moskowitz MA, Reinhard JF, Jr, Romero J, Melamed E, Pettibone DJ. Neurotransmitters and the fifth cranial nerve: Is there a relation to the headache phase of migraine? Lancet. 1979;2:883–5. doi: 10.1016/s0140-6736(79)92692-8. [DOI] [PubMed] [Google Scholar]
- 21.Newell DW, Grady MS, Eskridge JM, Winn HR. Distribution of angiographic vasospasm after subarachnoid hemorrhage: Implications for diagnosis by transcranial Doppler ultrasonography. Neurosurgery. 1990;27:574–7. [PubMed] [Google Scholar]
- 22.Onder A, Serarslan Y, Aydin MD, Aydin N, Ulvi H, Kotan D, et al. The role of trigeminal ganglion neuron density in the prevention of subarachnoid hemorrhage-induced basilar artery vasospasm: An experimental study. Neurosurg Q. 2009;19:264–9. [Google Scholar]
- 23.Rhoton AL, Jr, Fujii K, Fradd B. Microsurgical anatomy of the anterior choroidal artery. Surg Neurol. 1979;12:171–87. [PubMed] [Google Scholar]
- 24.Rosner SS, Rhoton AL, Jr, Ono M, Barry M. Microsurgical anatomy of the anterior perforating arteries. J Neurosurg. 1984;61:468–85. doi: 10.3171/jns.1984.61.3.0468. [DOI] [PubMed] [Google Scholar]
- 25.Saeki N, Rhoton AL., Jr Microsurgical anatomy of the upper basilar artery and the posterior circle of Willis. J Neurosurg. 1977;46:563–78. doi: 10.3171/jns.1977.46.5.0563. [DOI] [PubMed] [Google Scholar]
- 26.Shiokawa Y, Svendgaard NA. Cerebrovascular sensory innervation involved in the development of cerebral vasospasm following a subarachnoid hemorrhage. J Auton Nerv Syst. 1994;49:167–70. doi: 10.1016/0165-1838(94)90107-4. [DOI] [PubMed] [Google Scholar]
- 27.Sterio DC. The unbiased estimation of number and sizes of arbitrary particles using the disector. J Microsc. 1984;134:127–36. doi: 10.1111/j.1365-2818.1984.tb02501.x. [DOI] [PubMed] [Google Scholar]
- 28.Svendgaard NA, Arbab MA, Delgado-Zygmunt T, Shiokawa Y. Trigeminal afferents and brainstem centers involved in the occurrence of cerebral vasospasm. Neurol Res. 1996;18:394–400. doi: 10.1080/01616412.1996.11740443. [DOI] [PubMed] [Google Scholar]
- 29.Svendgaard NA, Göksel M, Westring S. Trigeminal nerve and brainstem catecholamine systems in cerebral vasospasm. Neurol Med Chir (Tokyo) 1998;38:146–51. doi: 10.2176/nmc.38.suppl_146. [DOI] [PubMed] [Google Scholar]
- 30.Wolburg H, Paulus W. Choroid plexus: Biology and pathology. Acta Neuropathol. 2010;119:75–88. doi: 10.1007/s00401-009-0627-8. [DOI] [PubMed] [Google Scholar]
- 31.Yasargil MG, Antic J, Laciga R, Jain KK, Hodosh RM, Smith RD. Microsurgical pterional approach to aneurysms of the basilar bifurcation. Surg Neurol. 1976;6:83–91. [PubMed] [Google Scholar]
- 32.Zhang QJ, Hara H, Kobayashi S. Distribution patterns of sensory innervation from the trigeminal ganglion to cerebral arteries in rabbits studied by wheat germ agglutinin- conjugated horseradisg peroxidase anterogrde tracing. Neurosurgery. 1993;32:993–9. doi: 10.1227/00006123-199306000-00019. [DOI] [PubMed] [Google Scholar]