


Abstract
The yeast heat shock transcription factor (HSF) belongs to the winged helix family of proteins. HSF binds DNA as a trimer, and additional trimers can bind DNA co-operatively. Unlike other winged helix–turn–helix proteins, HSF’s wing does not appear to contact DNA, as based on a previously solved crystal structure. Instead, the structure implies that the wing is involved in protein–protein interactions, possibly within a trimer or between adjacent trimers. To understand the function of the wing in the HSF DNA-binding domain, a Saccharomyces cerevisiae strain was created that expresses a wingless HSF protein. This strain grows normally at 30°C, but shows a decrease in reporter gene expression during constitutive and heat-shocked conditions. Removal of the wing does not affect the stability or trimeric nature of a protein fragment containing the DNA-binding and trimerization domains. Removal of the wing does result in a decrease in DNA-binding affinity. This defect was mainly observed in the ability to form the first trimer-bound complex, as the formation of larger complexes is unaffected by the deletion. Our results suggest that the wing is not involved in the highly co-operative nature of HSF binding, but may be important in stabilizing the first trimer bound to DNA.
INTRODUCTION
Winged helix proteins are a growing subfamily of the helix–turn–helix DNA-binding proteins, of which many have been studied by X-ray crystallography and NMR spectroscopy (1–4). Among the characteristics associated with the DNA-binding domains of winged helix proteins are an extended loop (or wing) sequence found C-terminal to the DNA-recognition helix (5,6). The DNA-recognition helix makes sequence-specific contacts with the major groove of DNA while the wing typically contacts DNA along the phosphodiester backbone and, in some cases, the minor groove. However, exceptions to this scenario exist. Recently, the structure of hRFX1 bound to DNA was solved, showing that its wing contacts the major groove, supplying most of the protein contacts with DNA, while the DNA-recognition helix makes only one contact with the minor groove (4).
The yeast heat shock transcription factor (HSF) was also found to belong to the family of winged helix proteins, though it contains unique features (7). First, the winged helix DNA-binding proteins commonly bind DNA as monomers, dimers or tetramers, but HSF binds as a trimer with strong co-operativity between trimers. Secondly, a crystal structure of the yeast HSF DNA-binding domain in complex with DNA showed that the wing had no observable DNA contacts (7). This raised the question of the wing’s possible role in DNA binding.
Crystallographic and NMR studies have shown that most residues of the yeast HSF wing were dynamic and presumably flexible in both the apo and complex structures (7–10). However, one model predicted that the wing provides an interface for protein–protein contacts when bound to DNA (7). A more recently solved crystal structure of the HSF DNA-binding domain contained a complete ordered structure of the wing, though not in complex with DNA (11). Implications from this structure predict that the wing may provide a large surface area for interaction with a second DNA-binding domain. This interaction could be important for contacts within a single trimer or contacts between two trimers to promote co-operative binding.
Yeast HSF readily forms trimers in solution through interactions of its oligomerization domain predicted to form a three-stranded coiled-coil (12,13). The yeast HSF trimers constitutively bind DNA upstream of heat shock promoters at sequences called heat shock elements (HSEs), which are comprised of multiple inverted nGAAn tandem repeats (14,15). Two adjacent repeats can exist in either a head-to-head (nGAAnnTTCn) or tail-to-tail (nTTCnnGAAn) orientation (16). DNA-binding studies with HSF have shown that affinity depends on the number and sequence of repeats present on DNA, with high-affinity binding occurring when at least three repeats are present (16,17). Typical gel shift assays show multiple bound complexes on DNA, illustrating the highly co-operative nature of HSF (18,19).
Interestingly, not all HSFs contain a wing sequence within the DNA-binding domain. HSFs from plants, such as Lycopersicon peruvianum (tomato) and Arabidopsis thaliana, lack a wing motif in the DNA-binding domain (20,21). HSFs from both of these organisms specifically bind DNA with high affinity and form multiple bound complexes on DNA. The A.thaliana HSF was also determined to exist as a trimer in solution, although the oligomeric state of the DNA-bound complexes has not been determined.
In this study, we attempt to understand the role of the wing in yeast HSF by creating wing-deletion mutants. A yeast strain expressing the mutant protein was tested for HSF-dependent expression from a reporter plasmid. Fragments of wild-type and wingless proteins were purified and used in DNA binding assays. The data presented here show that the wing is not critical for the stability of the DNA-binding domain, nor is it important for the formation of a trimeric complex in solution. The wing is also not important for co-operativity between two adjacent trimers when bound to DNA. Removal of the wing does reduce transcriptional activity of HSF under constitutive and heat-induced conditions and also decreases the affinity of a single trimer to DNA.
MATERIALS AND METHODS
Proteins
HSF from Saccharomyces cerevisiae was used for the in vivo assays because of the ease of genetics in that system. Kluyveromyces lactis HSF fragments were used for the in vitro studies because of the structural data available for that protein, as well as ease of overexpression and purification (8,22,23). The core regions containing the DNA-binding and trimerization domains are highly conserved between the two yeast species and data suggest that the two proteins are homologous both structurally and functionally (8,23,24).
Plasmids
Plasmid pHN1031 is a derivative of the CEN/ARS plasmid pRS414 (Stratagene) and expresses wild-type S.cerevisiae HSF from its natural promoter (25). The wingless version of pHN1031 was created by site-directed mutagenesis, using primer SH50 (5′-TGGATGGCATAAAGTTGATCCTGATAAGTGGCAATTTG-3′) to delete residues 239–249, and substituting Ser250 and Asp251 with aspartate and proline, respectively. The latter substitutions were designed to mimic the corresponding residues in the naturally wingless L.peruvianum HSF (20) (see Fig. 1 for comparison).
Figure 1.

Amino acid sequence comparison of the DNA-binding domains from the two yeast strains used in this study along with the naturally wingless L.peruvianum (tomato) sequence. The sequences of the two wingless HSF mutants used in this study are also included. Secondary structural characteristics are included below the sequence, and were based on the crystal structure of the K.lactis DNA-binding domain (8).
Plasmid pHN208 was previously described and is a pET-3b derivative containing the DNA-binding and trimerization domains of K.lactis HSF (amino acids 192–394) (18). The wingless mutation was introduced into pHN208 by mega-primer PCR mutagenesis (26,27), using primers JH10 (5′-CCGGTGATGCCGGCCACG-3′) and JH26 (5′-GGATATCCGGATATAGTTCC-3′) as the upstream and downstream primers, respectively, and primer MPC100 (5′-AACATGTATGGGTGGCACAAAGTGGATCCTAGCAGATGGGAATTTGAG-3′) as the mutagenic primer. The mutations deleted residues Gln261 to Asn271, and substituted Asn272 and Asp273 with aspartate and proline, respectively, to create plasmid pHN845 (see Fig. 1 for sequence comparisons).
The K.lactis DNA-binding domains were expressed as GST fusions from derivatives of plasmid pGEX-2tk (Pharmacia). The wild-type DNA-binding domain plasmid (pHN281R) was previously described (7). The wingless version was created by PCR amplification of the DNA-binding domain sequence from pHN845, using primers that created SmaI and EcoRI sites at the 5′- and 3′-ends of the fragment, respectively. The fragment was then cut with the restriction enzymes and ligated into SmaI/EcoRI-cleaved pHN281R plasmid. The presence of the proper mutations for all plasmids was confirmed by nucleotide sequencing.
Western analysis
Western analyses of wild-type and wingless HSF proteins were performed as previously described (23). Briefly, yeast strains were grown to saturation on selective synthetic media at 30°C, then 1.5 ml of cells was harvested by centrifugation and resuspended in 0.125 M Tris pH 6.8, 4% (w/v) SDS, 20% (v/v) glycerol and 1% (v/v) β-mercaptoethanol. Cells were lysed by boiling and vortexing in the presence of glass beads, and cell debris was removed by centrifugation. Cell extracts were electrophoresed on a 4–20% (w/v) denaturing gradient gel and electroblotted overnight onto an Immobilon filter (Millipore). The filter was probed with rabbit anti-HSF antibody (a gift from Peter Sorger, Massachusetts Institute of Technology) and then analyzed by the ECL western blotting detection system (Amersham Pharmacia).
β-Galactosidase assay
Assays used a yeast tester strain that is a haploid derivative of S.cerevisiae W303 (MATa ade2-1 trp1 can1-100 leu2,3-112 his3-11,15 ura3), which contained a chromosomal disruption of the HSF gene (HSFΔ2::LEU2) and harbors a version of the plasmid YCp50 that contains the wild-type HSF gene under the control of a GAL1 promoter (25,28). The CEN/ARS plasmids expressing wild-type or wingless HSF from the natural HSF promoter were introduced into this strain. The transformants were then cured of the pGAL1-HSF plasmid by growth in the presence of 5-fluoroorotic acid and uracil (29). The resultant strains were transformed with plasmid pHSE2-βgal, which contains the β-galactosidase gene downstream from a synthetic heat shock promoter (30). Cells were grown at 30°C until OD600 was 0.1–0.3 in selective synthetic media. Heat-induced samples were then incubated at 42°C for 30 min in a shaking water bath followed by recovery growth at 30°C for 90 min. Cells (3 ml) were centrifuged and resuspended in 200 µl of Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, 50 mM β-mercaptoethanol) plus 10% Triton X-100 and 200 µl glass beads (425–600 µm; Sigma). Cells were incubated on ice for 10 min, vortexed for 10 min at 4°C, and then centrifuged for 10 min to remove cell debris. The protein concentration of the crude extract was measured by Bio-Rad Bradford assay. β-Galactosidase levels were measured by the ONPG assay (31). Either 100 µl (non-induced cells) or 50 µl (heat-induced cells) were added to Z buffer to a final volume of 500 µl to which 100 µl of 4 mg/ml ONPG (in 0.1 M KPO4) was added. The mixture was incubated at 28°C for at least 30 min, the reaction terminated by the addition of 250 µl 1 M Na2CO3, and the extent of the reaction was measured by absorbance at 420 nm. β-Galactosidase activity was calculated as [(OD420 × 500)/(t × v × p)] × 0.5, where t is time (in min), v is volume of extract (in ml) and p is protein concentration (in mg/ml).
Protein purification
Wild-type and wingless HSF protein fragments containing the DNA-binding and trimerization domains were expressed and purified identically. Expression plasmids were transformed into BL21(DE3) cells harboring a derivative of pACYC177 containing lacIQ (32,33). Cells were grown in Terrific Broth at 37°C until OD600 was 0.2, and then induced by the addition of IPTG to a final concentration of 2 mM. Cells were grown for 2 h, harvested by centrifugation and stored as frozen suspensions of ~1 g wet cells per 2 ml of a solution of 200 mM NaCl, 50 mM Tris pH 7.5, 10% glycerol and 5 mM MgCl2. Cells were then thawed, supplemented with 1 µg/ml aprotinin, 2 µg/ml pepstatin and 1 µg/ml leupeptin, incubated on ice for 30 min with 200 µg/ml lysozyme, disrupted by sonication and centrifuged at 39 000 g for 15 min at 4°C. The high-speed supernatant was diluted 4-fold with 50 mM Tris pH 7.5 and loaded onto a heparin–agarose column (Sigma). The column was developed with a gradient from 50 to 1250 mM NaCl in 50 mM Tris pH 7.5. The proteins eluted over a NaCl concentration range from 0.6 to 1.0 M.
Fractions from the heparin column with the highest concentration of protein were diluted with 50 mM Tris pH 7.5 to a final NaCl concentration of 50 mM, and loaded onto a sulfopropyl column (Waters). The column was developed with a gradient from 50 to 600 mM NaCl in 50 mM Tris pH 7.5 and the protein eluted between 200 and 250 mM NaCl. At this point the proteins appeared to be ∼90% pure from Coomassie-stained SDS–PAGE gels. Protein concentrations were determined using an extinction coefficient of 33 700 cm–1M–1, which was calculated from the tyrosine and tryptophan content of the proteins.
The K.lactis HSF DNA-binding domain proteins were expressed as GST fusions. Cleavage of the fusion proteins and subsequent purification of the isolated DNA-binding domains has been previously described (7,23).
Circular dichroism (CD) spectroscopy
CD spectra and thermal melting of the wild-type and wingless HSF DNA-binding domains were performed as previously described (23). Briefly, lyophilized proteins were resuspended in water and diluted to 20 µM in 20 mM sodium phosphate buffer pH 7.6. CD spectra were recorded on an AVIV 60 DS spectropolarimeter. For the wavelength scans, CD measurements were taken at 25°C at 1 nm intervals using 0.1 cm path length, 1 s time constant and 1.5 nm bandwidth. The CD signal was plotted as mean residue ellipticity (θres) in order to allow direct comparison of the CD spectra independent of the size of the protein (34). The thermal melting experiment measured the CD signal at 222 nm as the temperature was raised in steps of 2°C, with a 4 min equilibration time, 10 s averaging time and 1.5 nm bandwidth.
Analytical ultracentrifugation
Sedimentation equilibrium experiments were done using a Beckman XL-A analytical ultracentrifuge. The proteins were in 50 mM HEPES pH 8.0, 50 mM NaCl, with starting concentrations from 0.2 to 10 µM. The experiment was done at 25°C and 20 000 r.p.m., with the concentration gradient determined by the absorption at 280 nm.
Gel shift assays
Binding reactions were conducted similar to those previously described (22). Briefly, reactions (20 µl) contained the indicated amount of protein with ~2 fmol end-labeled DNA fragment, 50 mM HEPES pH 8.0, 100 mM NaCl, 1 mM EDTA, 10% glycerol, 500 µg/ml BSA and 25 µg/ml poly(dI–dC). After a 10 min incubation at 22°C, the samples were fractionated on a 6% polyacrylamide gel at 12 V/cm in 0.25× TBE (23 mM Tris–borate pH 8.3, 0.5 mM EDTA) for 2 h at 22°C. Electrophoretic bands were analyzed and quantified by a Molecular Dynamics PhosphorImager and MD ImageQuant software, and apparent dissociation constants were calculated as previously described (22,23). The various short duplex DNA substrates used in this study are shown in Table 1. The longer 6-box substrate was a 154-bp XhoI/SpeI fragment from a pBL3 derivative (23) and had the HSE sequence aGAAtaTTCtaGAAtaTTCtaGAAtaTTCa. All substrates were kinase end-labeled with [32P]ATP.
Table 1. DNA substrates used in the gel shift studies.
| Substrate |
Sequence (5′–3′) |
| 3-box |
gccaGAAcaTTCcaGAAcacc |
| 4-box HH |
gccaTTCcaGAAcaTTCcaGAAcacc |
| 4-box TT |
gccaGAAcaTTCcaGAAcaTCCcacc |
| 2-box HH |
gatgccGAAccTTCcatgcc |
| 2-box TT | CatgccTTCccGAAcatgcc |
One strand of the duplex substrates is shown. Opposite strands were complementary and formed blunt-ended substrates. GAA binding sites are capitalized.
RESULTS
Wingless HSF allows normal growth but is defective in HSF-mediated transcription
The HSF is an essential protein for S.cerevisiae. We first determined if a wingless HSF mutant allows growth of S.cerevisiae by using a tester strain that has the genomic HSF gene disrupted, with growth dependent on a plasmid-produced HSF. We compared the ability of the tester strain to grow in the presence of a CEN/ARS plasmid containing either wild-type or wingless HSF expressed from the natural HSF promoter. The wingless mutant was created by a deletion in HSF that removed the 10 residues comprising just the wing and substituted two residues at the C-terminal end of the deletion to mimic the amino acid sequence of the naturally wingless L.peruvianum (tomato) HSF (20) (see Fig. 1). The strain containing the wingless HSF expression plasmid grew normally on plates at 30°C, indicating that the wingless HSF mutant still contained enough activity to maintain viability (data not shown). The growth curves for the two strains in selective synthetic media at 30°C were compared (Fig. 2A). Doubling times were nearly identical between the two strains (~110 and 120 min for wild-type and wingless HSF strains, respectively), suggesting that the wingless HSF does not impede growth under normal conditions. Western analysis showed that HSF levels were similar between the wild-type and wingless HSF strains grown at 30°C (Fig. 2B).
Figure 2.

Growth curve and western analysis of S.cerevisiae strains producing wild-type and wingless HSF. (A) Growth of S.cerevisiae strains producing wild-type (squares) or wingless (circles) HSF in selective synthetic media at 30°C. (B) Western analysis of HSF protein produced from S.cerevisiae strains producing wild-type (wt) or wingless (wg–) HSF. Western analysis used a rabbit anti-HSF antibody (obtained from Peter Sorger).
Transcriptional activation by wild-type and wingless HSF was measured using a pHSE2-βgal reporter plasmid (30,35). This plasmid contains a synthetic heat shock promoter upstream of the β-galactosidase gene. The HSF-regulated promoter is considered to be a strong promoter that is occupied constitutively by HSF (36,37). Transcription from the promoter can then be measured indirectly by β-galactosidase activity from crude cell extracts. Cells were grown continuously at 30°C to measure constitutive HSF activity, and were incubated at 42°C for 30 min to measure heat-induced activity. The wingless HSF caused a dramatic decrease in constitutive expression of β-galactosidase, resulting in about one-third the transcriptional activity compared to wild-type HSF (Table 2). This defect was also observed with heat-induced cells, as cells containing wingless HSF only reached transcriptional levels of approximately one-third wild-type levels. Interestingly, the fold-induction after heat treatment did not seem to be affected by the absence of the wing, as both strains increased transcription ~7-fold. These results indicate that the wing of the DNA-binding domain is involved in maintaining constitutive levels of transcription by HSF, although the mutant still has enough activity to allow normal growth.
Table 2. β-Galactosidase assay results with S.cerevisiae expressing wild-type or wingless HSF.
| Yeast strain |
Constitutive |
Heat-induced |
Fold-induction |
| Wild-type HSF |
11.4 ± 3.0 |
82.5 ± 27.2 |
7.3 |
| Wingless HSF | 3.8 ± 1.0 | 25.8 ± 3.6 | 6.8 |
The β-galactosidase units for each experiment are shown as determined by procedures described in Materials and Methods. Fold-induction was calculated by dividing the heat-induced values by the constitutive values.
Removal of the wing motif does not affect structure of the DNA-binding domain or trimerization by HSF
We used CD spectroscopy to determine if the wingless deletion causes any gross changes in secondary structure that would affect protein function. CD spectroscopy provides a convenient measure of secondary structure characteristics of peptides in solution (38). The wild-type and wingless DNA-binding domains had similar spectral features in the α-helix/β-sheet region of 200–220 nm (Fig. 3A), indicating that the proteins were well-folded and contained mixed αβ secondary structure characteristics, consistent with previous structural studies of the HSF DNA-binding domain (8–10). The wingless and wild-type proteins do differ slightly in the 230 nm range (Fig. 3A). Positive CD signals at 230 nm have been correlated with tryptophan residues (39). As there are two tryptophan residues on either side of the wing, it is likely that removal of the wing could affect the far-UV spectra without affecting secondary structure. We also obtained thermal melting profiles of each protein by measuring the loss of CD signal at 222 nm as the temperature was increased. Both proteins gave similar melting profiles, with the Tm of each protein near 57°C (Fig. 3B). Denaturation was reversible for both proteins upon cooling. These results support the idea that removal of the wing had little effect on protein stability and folding, thus suggesting that any phenotypic difference by the mutant is likely caused by a change in macromolecular interactions.
Figure 3.

Wild-type and wingless HSF DNA-binding domains have similar secondary structure and thermal melting profiles. (A) CD analysis was performed for the wild-type (squares) and wingless (circles) DNA-binding domain. (B) Proteins underwent thermal melting followed by CD measurements at 222 nm. The fraction of the protein folded is shown for the wild-type (squares) and wingless (circles) protein.
We tested if the wingless mutation affected the ability of HSF to form trimers in solution. Wild-type and wingless protein fragments containing the DNA-binding and trimerization domains underwent analytical ultracentrifugation. The molecular mass of the wild-type protein, as determined by the sedimentation equilibrium, was 70.4 kDa, agreeing with the predicted molecular mass of a trimer of 71.9 kDa. The wingless HSF had a molecular mass of 71.0 kDa, also near the predicted size of its trimer complex (68.7 kDa). This provides evidence that the wingless deletion within the DNA-binding domain does not have an effect on trimer formation, thus eliminating this possibility as the cause for the transcription defect observed in vivo.
Wingless HSF shows decreased affinity to HSEs
Gel mobility shift assays were conducted with wild-type and mutant K.lactis HSF fragments containing the DNA-binding and trimerization domains. DNA binding was first assayed using a 3-box DNA substrate (see Table 1). Binding by wild-type HSF gave an apparent KD of 8.5 × 10–10 M, while the wingless HSF showed a considerable decrease in binding affinity, with an apparent KD of 5.1 × 10–9 M (Fig. 4). Differences in binding were more evident in the formation of the first trimer-bound complex, as the wingless protein readily formed the higher migrating species (Fig. 4A, lanes 1 and 2), comparable to wild-type (Fig. 4A, lanes 9 and 10).
Figure 4.
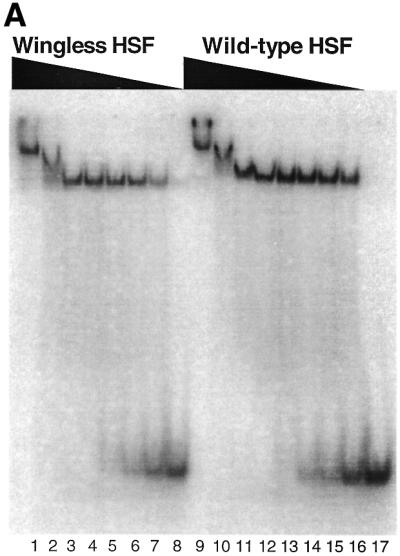

Wingless HSF binds DNA with less affinity than wild-type. (A) Gel shift assay with wingless (lanes 1–8) and wild-type (lanes 9–16) HSF fragments binding to a 3-box template. Samples contained increasing amounts of protein going from right to left, with the highest protein amount of 2 µM (lanes 1 and 9) and undergoing 3-fold dilutions for each subsequent lane. Lane 17 contained only labeled DNA substrate. The binding curves from the quantitation of gel shift assays using wild-type (B) or wingless (C) HSF are shown. Three separate experiments were quantitated as described in Materials and Methods. The apparent KD value is given for each plot.
To compare the co-operative nature of wild-type and wingless HSF, we conducted gel shift assays using a larger DNA fragment that contained a 6-box HSE. Previously, studies showed that a minimum of five repeats were required to observe full co-operative interactions by HSF (17). With the 6-box DNA substrate, we observed little difference in the binding pattern of the larger complexes between wild-type and wingless HSF (Fig. 5). For each protein, nearly all of the substrate migrated with the larger complexes at 4 nM protein concentration (compare lane 4 with lane 12). This indicates that the wing is not required for co-operative binding by HSF, and may not be involved in interactions between adjacent HSF trimers.
Figure 5.
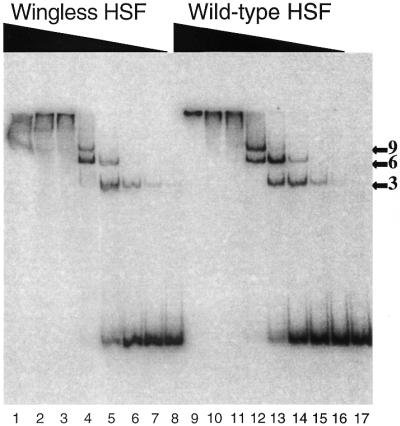
Wingless HSF binds with similar co-operativity as wild-type. Gel shift assay was conducted with a 6-box HSE DNA substrate. Samples contained increasing amounts of protein going from right to left, with the highest protein amount of 1 µM (lanes 1 and 9) and undergoing 4-fold dilutions for each subsequent lane. Lane 17 contains the unbound DNA substrate. Labels to the right of the gel indicate complexes formed by one (3), two (6) or three (9) bound trimers (as determined from previous studies; 18,19).
The wingless binding defect is not caused by a defect in head-to-head or tail-to-tail specific binding
Even though co-operativity between trimers was not affected by removal of the wing, we wanted to determine if the wing was involved in trimer binding in a particular orientation (i.e. head-to-head or tail-to-tail). Crystallographic and modeling studies suggest that this may be the case. The structure of two DNA-binding domains bound to DNA in the tail-to-tail orientation was previously solved (7). The model of the dimer-bound complex structure was used to create a 4-box model (Fig. 6), which suggests that a possible interaction between the wings may occur between two DNA-binding domains bound in the head-to-head orientation. In this orientation, the wings are in close proximity and may provide important contacts to stabilize the trimer on DNA. This implies that the absence of the wing may affect binding differently, depending on the orientation of the binding site.
Figure 6.

Model of the 4-box HSF DBD–DNA complex with a central head-to-head binding site (7). The wing motif is shown in red and includes the dotted line depicting the residues that remained unstructured. The DNA-binding domains depicted in the upper left and lower right corner of the figure would form a head-to-head interaction.
This prediction was tested by comparing binding of the proteins on substrates containing nGAAn repeats in each orientation (Table 1). First, we tested 2-box substrates, but did not find a significant difference between the two possible orientations (~2.5 × 10–7 and 7.5 × 10–7 M apparent KD values for head-to-head and tail-to-tail substrates, respectively, for both proteins). However, the affinity of wild-type and wingless HSF to these substrates was much reduced compared to the 3-box substrate (~50–900-fold decrease in apparent KD), which may have masked any differences caused by the binding orientation.
Therefore, we measured binding on 4-box substrates containing central head-to-head or tail-to-tail binding sites. Binding by wild-type protein did not seem to be affected by the orientation of the binding sites, as the KD was similar for both substrates, and comparable to binding to the 3-box substrates (Fig. 7A and B). The apparent binding affinity by wingless HSF was decreased with both substrates, though not as considerably as with the 3-box DNA (Fig. 7C and D). Binding to the head-to-head substrate by wingless gave a KD of 2.9 × 10–9 M, ~3-fold less than wild-type HSF (9.4 × 10–10 M). Wingless protein bound the tail-to-tail substrate with a KD of 1.8 × 10–9 M, or ~2-fold less than wild-type (8.2 × 10–10 M). These results did not agree with our prediction that absence of the wing would affect binding only to a head-to-head substrate, as we still observed a decrease in binding by the wingless protein with the tail-to-tail substrate. One explanation is that each 4-box substrate still contains both head-to-head and tail-to-tail binding sites. Therefore, these substrates will not completely limit binding only to a particular orientation. This would once again mask any defect occurring by the absence of the wing for DNA binding to a particular orientation.
Figure 7.

Wingless HSF binds with less affinity than wild-type to head-to-head and tail-to-tail DNA substrates. Binding plots are shown for wingless and wild-type HSF fragments binding to 4-box DNA substrates. Reactions were conducted and assayed in a similar manner to those in Figure 4. Reactions used either wild-type protein (A and B) or wingless protein (C and D) with either 4-box HH (A and C) or 4-box TT (B and D) substrates. The apparent KD for each plot is included.
We attempted to observe differences in head-to-head and tail-to-tail binding by measuring binding to the 2-box substrates by each DNA-binding domain (minus the trimerization domain). Although binding by the DNA-binding domain fragment was poor, no differences were observed with head-to-head or tail-to-tail binding by wild-type and wingless DNA-binding domains (data not shown). From these experiments, we conclude that the wing is not specifically involved with head-to-head or tail-to-tail binding. However, the wing is required for optimal binding to HSEs, and may explain the decrease in gene expression in vivo by the wingless HSF mutant.
DISCUSSION
The wings of winged helix proteins are commonly used to recognize and contact DNA in a specific manner, sometimes even inducing conformational changes in DNA. However, structural studies of a similar motif in yeast HSF predicted that the wing did not make specific DNA contacts, but may play a role in protein–protein interactions required for heat-induced gene regulation (7). Yeast HSF constitutively binds DNA as trimers and additional trimers bind in a highly co-operative fashion. The wing may play a critical role in binding of the first complex, or it may aid in the co-operative interactions between trimers.
We created an HSF mutant that contained a deletion of the wing motif. In vivo studies showed that the wingless HSF was defective in transcriptional activation, during both constitutive and heat-induced growth. However, both strains had a similar increase in induction after heat treatment (~7-fold). This suggests that wing removal primarily affects constitutive transcription and is not specific for the mechanism of heat-induced activation of HSF. Wild-type and wingless HSF protein fragments were purified and CD spectroscopy was used to determine that the wingless mutation did not affect the secondary structure or thermal stability of the DNA-binding domain. This suggests that any phenotypic effects were not caused by gross structural changes induced by the deletion. In addition, analytical ultracentrifugation results showed that trimerization was also unaffected by the wing deletion.
Gel shift assays showed that the defect in activation correlated with a decrease in DNA-binding affinity, with the mutant HSF having an ~6-fold decrease in binding affinity compared to the wild-type protein. The most dramatic difference in binding affinity was seen with formation of the first bound complex, formed by one trimer bound to DNA. Complexes formed by co-operative interactions did not seem to be affected by the mutation, suggesting that the wing is not involved in interactions between adjacent HSF trimers. Several experiments were performed to determine if this deficiency was caused by a specific defect in head-to-head or tail-to-tail interactions essential for optimal binding. However, using various DNA substrates and protein fragments, we were unable to correlate the decrease in binding to a defect in required interactions for a particular orientation.
We found that the wing is important for optimal binding of an HSF trimer to DNA. Previous structural evidence may give some insight about the role of the wing in stabilizing the DNA-bound form of an HSF trimer. The 4-box model predicts that the wings of two monomers in the head-to-head orientation are in close proximity and may provide areas of protein–protein contact (7). However, the 4-box model was created using the predicted structure of two DNA-binding domains bound in the tail-to-tail orientation. Also, although no specific DNA contacts by the wing were observed in the crystal complex, many residues of the wing were in a disordered state. Thus, this model may be a bit misleading, especially if the wing plays a separate role in the head-to-head complex, such as making important contacts with DNA. Protein–protein and protein–DNA interactions by the wing may be important for optimal binding of an HSF trimer to DNA. Clearly, additional structural data of the head-to-head bound complex, along with the fully ordered wing motif, is required to understand the role of the wing in HSF trimer binding.
Alternatively, or additionally, the wing may have another role within the general transcription complex to transcribe heat shock genes. We predict that within a single HSF trimer, only two wings from the DNA-binding domains would be buried in the complex with DNA, while the third wing would be exposed. Our experimental data suggests that the exposed wing is not involved in co-operative binding between adjacent trimers since the wingless protein formed the larger complexes in gel shift assays at a similar affinity as wild-type protein. Also, the wing is not involved specifically in heat-induced activation, as the wingless mutant showed a similar decrease in the levels of constitutive and heat-induced transcription. One possibility is that the wing may be involved more generally in transcription through interactions with other protein subunits of the general transcription complex. A defect in interacting with the general transcription machinery, along with a defect in DNA binding, may explain the transcription defect observed in vivo. Again, further studies are necessary for understanding the possible roles of the HSF wing motif.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Jim Lear for help with the analytical ultracentrifugation experiment, Bill DeGrado for use of his CD spectrometer, Giovanna Ghirlanda for help with the CD melt analysis and Peter Sorger for the anti-HSF antibody used in the western analysis. This work was supported by NIH grant GM44086.
References
- 1.Clark K.L., Halay,E.D., Lai,E. and Burley,S.K. (1993) Co-crystal structure of the HNF-3/fork head DNA-recognition motif resembles histone H5. Nature, 364, 412–420. [DOI] [PubMed] [Google Scholar]
- 2.Clubb R.T., Mizuuchi,M., Huth,J.R., Omichinski,J.G., Savilahti,H., Mizuuchi,K., Clore,G.M. and Gronenborn,A.M. (1996) The wing of the enhancer-binding domain of Mu phage transposase is flexible and is essential for efficient transposition. Proc. Natl Acad. Sci. USA, 93, 1146–1150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jin C., Marsden,I., Chen,X. and Liao,X. (1999) Dynamic DNA contacts observed in the NMR structure of winged helix protein–DNA complex. J. Mol. Biol., 289, 683–690. [DOI] [PubMed] [Google Scholar]
- 4.Gajiwala K.S., Chen,H., Cornille,F., Roques,B.P., Reith,W., Mach,B. and Burley,S.K. (2000) Structure of the winged-helix protein hRFX1 reveals a new mode of DNA binding. Nature, 403, 916–921. [DOI] [PubMed] [Google Scholar]
- 5.Gajiwala K.S. and Burley,S.K. (2000) Winged helix proteins. Curr. Opin. Struct. Biol., 10, 110–116. [DOI] [PubMed] [Google Scholar]
- 6.Wolberger C. and Campbell,R. (2000) New perch for the winged helix. Nat. Struct. Biol., 7, 261–262. [DOI] [PubMed] [Google Scholar]
- 7.Littlefield O. and Nelson,H.C.M. (1999) A new use for the ‘wing’ of the ‘winged’ helix–turn–helix motif in the HSF–DNA cocrystal. Nat. Struct. Biol., 6, 464–470. [DOI] [PubMed] [Google Scholar]
- 8.Harrison C.J., Bohm,A.A. and Nelson,H.C. (1994) Crystal structure of the DNA binding domain of the heat shock transcription factor. Science, 263, 224–227. [DOI] [PubMed] [Google Scholar]
- 9.Damberger F.F., Pelton,J.G., Harrison,C.J., Nelson,H.C. and Wemmer,D.E. (1994) Solution structure of the DNA-binding domain of the heat shock transcription factor determined by multidimensional heteronuclear magnetic resonance spectroscopy. Protein Sci., 3, 1806–1821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Damberger F.F., Pelton,J.G., Liu,C., Cho,H., Harrison,C.J., Nelson,H.C. and Wemmer,D.E. (1995) Refined solution structure and dynamics of the DNA-binding domain of the heat shock factor from Kluyveromyces lactis. J. Mol. Biol., 254, 704–719. [DOI] [PubMed] [Google Scholar]
- 11.Hardy J.A. and Nelson,H.C.M. (2000) Proline in α-helical kink is required for folding kinetics but not for kinked structure, function, or stability of heat shock transcription factor. Protein Sci., 9, 2128–2141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Peteranderl R. and Nelson,H.C. (1992) Trimerization of the heat shock transcription factor by a triple-stranded α-helical coiled-coil. Biochemistry, 31, 12272–12276. [DOI] [PubMed] [Google Scholar]
- 13.Peteranderl R., Rabenstein,M., Shin,Y.K., Liu,C.W., Wemmer,D.E., King,D.S. and Nelson,H.C. (1999) Biochemical and biophysical characterization of the trimerization domain from the heat shock transcription factor. Biochemistry, 38, 3559–3569. [DOI] [PubMed] [Google Scholar]
- 14.Xiao H. and Lis,J.T. (1988) Germline transformation used to define key features of heat-shock response elements. Science, 239, 1139–1142. [DOI] [PubMed] [Google Scholar]
- 15.Amin J., Fernandez,M., Ananthan,J., Lis,J.T. and Voellmy,R. (1994) Cooperative binding of heat shock transcription factor to the Hsp70 promoter in vivo and in vitro. J. Biol. Chem., 269, 4804–4811. [PubMed] [Google Scholar]
- 16.Perisic O., Xiao,H. and Lis,J.T. (1989) Stable binding of Drosophila heat shock factor to head-to-head and tail-to-tail repeats of a conserved 5 bp recognition unit. Cell, 59, 797–806. [DOI] [PubMed] [Google Scholar]
- 17.Xiao H., Perisic,O. and Lis,J.T. (1991) Cooperative binding of Drosophila heat shock factor to arrays of a conserved 5 bp unit. Cell, 64, 585–593. [DOI] [PubMed] [Google Scholar]
- 18.Rye H.S., Drees,B.L., Nelson,H.C.M. and Glazer,A.N. (1993) Stable fluorescent dye–DNA complexes in high sensitivity detection of protein–DNA interactions. Application to heat shock transcription factor. J. Biol. Chem., 268, 25229–25238. [PubMed] [Google Scholar]
- 19.Drees B.L., Rye,H.S., Glazer,A.N. and Nelson,H.C.M. (1996) Environment-sensitive labels in multiplex fluorescence analyses of protein–DNA complexes. J. Biol. Chem., 271, 32168–32173. [DOI] [PubMed] [Google Scholar]
- 20.Scharf K.D., Rose,S., Zott,W., Schoffl,F. and Nover,L. (1990) Three tomato genes code for heat stress transcription factors with a region of remarkable homology to the DNA-binding domain of the yeast HSF. EMBO J., 9, 4495–4501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hubel A. and Schoffl,F. (1994) Arabidopsis heat shock factor: isolation and characterization of the gene and the recombinant protein. Plant Mol. Biol., 26, 353–362. [DOI] [PubMed] [Google Scholar]
- 22.Drees B.L., Grotkopp,E.K. and Nelson,H.C. (1997) The GCN4 leucine zipper can functionally substitute for the heat shock transcription factor’s trimerization domain. J. Mol. Biol., 273, 61–74. [DOI] [PubMed] [Google Scholar]
- 23.Hardy J.A., Walsh,S.T. and Nelson,H.C.M. (2000) Role of an α-helical bulge in the yeast heat shock transcription factor. J. Mol. Biol., 295, 393–409. [DOI] [PubMed] [Google Scholar]
- 24.Jakobsen B.K. and Pelham,H.R. (1991) A conserved heptapeptide restrains the activity of the yeast heat shock transcription factor. EMBO J., 10, 369–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hubl S.T., Owens,J.C. and Nelson,H.C.M. (1994) Mutational analysis of the DNA-binding domain of yeast heat shock transcription factor. Nat. Struct. Biol., 1, 615–620. [DOI] [PubMed] [Google Scholar]
- 26.Picard V., Ersdal-Badju,E., Lu,A. and Bock,S.C. (1994) A rapid and efficient one-tube PCR-based mutagenesis technique using Pfu DNA polymerase. Nucleic Acids Res., 22, 2587–2591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Datta A.K. (1995) Efficient amplification using ‘megaprimer’ by asymmetric polymerase chain reaction. Nucleic Acids Res., 23, 4530–4531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sorger P.K. and Pelham,H.R. (1988) Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell, 54, 855–864. [DOI] [PubMed] [Google Scholar]
- 29.Sikorski R.S. and Boeke,J.D. (1991) In vitro mutagenesis and plasmid shuffling: from cloned gene to mutant yeast. Methods Enzymol., 194, 302–318. [DOI] [PubMed] [Google Scholar]
- 30.Sorger P.K. and Pelham,H.R. (1987) Purification and characterization of a heat-shock element binding protein from yeast. EMBO J., 6, 3035–3041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ausubel F.M., Brent,R., Kingston,R.E., Moore,D.D., Seidman,J.G., Smith,J.A. and Struhl,K. (1994) Current Protocols in Molecular Biology, Vols 1 and 2. Greene Publishing Associates and John Wiley & Sons, New York, NY.
- 32.Stark M.J. (1987) Multicopy expression vectors carrying the lac repressor gene for regulated high-level expression of genes in Escherichia coli. Gene, 51, 255–267. [DOI] [PubMed] [Google Scholar]
- 33.Studier F.W., Rosenberg,A.H., Dunn,J.J. and Dubendorff,J.W. (1989) Use of T7 RNA polymerase to direct expression of cloned genes. Methods Enzymol., 185, 60–89. [DOI] [PubMed] [Google Scholar]
- 34.Flick K.E., Gonzalez,L.,Jr, Harrison,C.J. and Nelson,H.C. (1994) Yeast heat shock transcription factor contains a flexible linker between the DNA-binding and trimerization domains. Implications for DNA binding by trimeric proteins. J. Biol. Chem., 269, 12475–12481. [PubMed] [Google Scholar]
- 35.Sorger P.K. (1990) Yeast heat shock factor contains separable transient and sustained response transcriptional activators. Cell, 62, 793–805. [DOI] [PubMed] [Google Scholar]
- 36.Jakobsen B.K. and Pelham,H.R. (1988) Constitutive binding of yeast heat shock factor to DNA in vivo. Mol. Cell Biol., 8, 5040–5042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gross D.S., English,K.E., Collins,K.W. and Lee,S.W. (1990) Genomic footprinting of the yeast HSP82 promoter reveals marked distortion of the DNA helix and constitutive occupancy of heat shock and TATA elements. J. Mol. Biol., 216, 611–631. [DOI] [PubMed] [Google Scholar]
- 38.Pelton J.T. and McLean,L.R. (2000) Spectroscopic methods for analysis of protein secondary structure. Anal. Biochem., 277, 167–176. [DOI] [PubMed] [Google Scholar]
- 39.Woody R.W. (1994) Contributions of tryptophan side chains to the far-ultraviolet circular dichroism of proteins. Eur. Biophys. J., 23, 253–262. [DOI] [PubMed] [Google Scholar]