

Abstract
Background
Plasma lipases and lipid transfer proteins are involved in the generation and speciation of high density lipoproteins. In this study we have examined the influence of plasma lipases and lipid transfer protein activities on the transfer of free cholesterol (FC) and phospholipids (PL) from lipid emulsion to human, rat and mouse lipoproteins. The effect of the lipases was verified by incubation of labeled (3H-FC,14C-PL) triglyceride rich emulsion with human plasma (control, post-heparin and post-heparin plus lipase inhibitor), rat plasma (control and post-heparin) and by the injection of the labeled lipid emulsion into control and heparinized functionally hepatectomized rats.
Results
In vitro, the lipase enriched plasma stimulated significantly the transfer of 14C-PL from emulsion to high density lipoprotein (p<0.001) but did not modify the transfer of 3H-FC. In hepatectomized rats, heparin stimulation of intravascular lipolysis increased the plasma removal of 14C-PL and the amount of 14C-PL found in the low density lipoprotein density fraction but not in the high density lipoprotein density fraction. The in vitro and in vivo experiments showed that free cholesterol and phospholipids were transferred from lipid emulsion to plasma lipoproteins independently from each other. The incubation of human plasma, control and control plus monoclonal antibody anti-cholesteryl ester transfer protein (CETP), with 14C-PL emulsion showed that CETP increases 14C-PL transfer to human HDL, since its partial inhibition by the anti-CETP antibody reduced significantly the 14C-PL transfer (p<0.05). However, comparing the nontransgenic (no CETP activity) with the CETP transgenic mouse plasma, no effect of CETP on the 14C-PL distribution in mice lipoproteins was observed.
Conclusions
It is concluded that: 1-intravascular lipases stimulate phospholipid transfer protein mediated phospholipid transfer, but not free cholesterol, from triglyceride rich particles to human high density lipoproteins and rat low density lipoproteins and high density lipoproteins; 2-free cholesterol and phospholipids are transferred from triglyceride rich particles to plasma lipoproteins by distinct mechanisms, and 3 - CETP also contributes to phospholipid transfer activity in human plasma but not in transgenic mice plasma, a species which has high levels of the specific phospholipid transfer protein activity.
Background
There have been plenty of epidemiological, clinical and experimental evidence that plasma high density lipoproteins levels are inversely correlated with the risk of atherosclerosis [1,2]. The contribution of enzymes and proteins associated with HDL to its process of generation and maturation have been extensively studied, both in vitro and in vivo.
The plasma cholesteryl ester transfer protein (CETP) modulates HDL levels and composition. It mediates the transfer of cholesteryl ester (CE) from HDL to triglyceride (TG) rich lipoproteins (LP), while TG is transferred in the opposite direction, to HDL. One way CE transfer from HDL to LDL may also occur [3]. CETP also promotes phospholipid (PL) transfer to human HDL [4,5,6]. CETP activity has been directly correlated with LDL cholesterol levels and inversely correlated with HDL cholesterol levels in human plasma [3,7].
PLTP, a specific phospholipid transfer protein, has been identified in human plasma [8,9] and in plasma of other vertebrate species [10]. It promotes the PL transfer from VLDL to HDL [11]. In addition to PL, PLTP transfers free cholesterol (FC) from PL/FC vesicles to HDL, although with a low efficiency [12]. Both, CETP and PLTP, can promote HDL remodelling. While CETP, together with hepatic lipoprotein lipase, stimulates the generation of small alpha-HDL, PLTP favours the emergence of large alpha-HDL particles [13]. Significantly higher levels of HDL-cholesterol were observed in human PLTP transgenic mice [14]. Furthermore, overexpression of human PLTP produced by recombinant adenovirus injection into mice, resulted in increased levels of prebeta-HDL, increased fractional catabolic rate and liver uptake of CE and PL from HDL [15].
After intravascular hydrolysis of TG rich LP by lipoprotein lipase (LPL), surface remnant components such as FC, PL and apoproteins may provide substrates for generation or modification of plasma HDL. Net transfer of PL and FC from chylomicrons and VLDL to HDL has previously been demonstrated in rats [16,17] and in human plasma after a fat meal [18,19] or during lipolysis [11]. The contribution of the lipolysed LP components to HDL formation has been reinforced by several studies where the activity of the enzyme lipoprotein lipase LPL was shown to correlate with HDL cholesterol levels in human plasma [20,21,22]. However, changes in the HDL-cholesterol concentration have not been observed in mice overexpressing LPL [23] or in LPL heterozygous knockout mice [24].
The metabolism of HDL in rats and mice differs significantly from that in humans. Part of the species differences observed in mice and in rats may result from their high levels of circulating lipases [25,26], lack of CETP [27] and high levels of PLTP [10]. Clee et al. [28] have shown that, in double transgenic mice overexpressing LPL and CETP, HDL cholesterol levels were significantly influenced by the LPL activity while no such correlation was observed in the absence of CETP expression.
In this study we have further evaluated the influence of plasma lipases and CETP on the free cholesterol and phospholipid transfer from triglyceride rich lipid emulsion similar to chylomicrons [29,30] to human, rat and mouse lipoproteins, in vitro and in vivo.
Results
The influence of the intravascular lipase activity on the phospholipids (PL) and free cholesterol (FC) transfer from lipid emulsion (EM) to plasma lipoproteins was evaluated in human control plasma, in lipase-enriched post-heparin plasma and in post-heparin plasma containing a lipase inhibitor, tetrahydrolipstatin (THL) [34]. PL transfer was determined after 30 minutes of incubation because Tall et al. [8] have demonstrated that PL net transfer reaches the maximum after this incubation period. The post-heparin plasma lipoprotein lipase (LPL) and hepatic lipoprotein lipase (HL) activities were 2527 ± 1353 and 3747 ± 2116 nmol of fatty acid/ml/h, respectively. The control and post-heparin + THL plasmas had no detectable lipase activity, showing that 2 mM THL completely inhibited the activities of the lipases present in the post-heparin plasma. Table 1 shows that 14C-PL was preferentially transferred to HDL (p<0.001) while 3H-FC was equally distributed to LDL and HDL fractions under all 3 plasma conditions. The transfer of 14C-PL from EM to HDL was significantly stimulated (+60%) by the increase in the lipases' activities in the post-heparin plasma (p<0.001). This stimulation of the PL transfer was abolished in the presence of THL (p<0.01). Considering that HL also has phospholipase activity, the integrity of the phospholipid transferred to HDL was checked by thin layer chromatography. Ninety eight % of the 14C-PL was recovered in the PL band and no radioactivity was detected in the fatty acid bands. The proportion of 3H-FC and 14C-PL found in HDL (0.4) was different from that found in LDL (2.3) or in the EM (1.5), which suggests that these components of the EM were independently transferred to the LP fractions and not as a surface unit detached from the lipolysed EM.
Table 1.
Influence of plasma lipases on the 3H-free cholesterol (FC) and 14C-phospholipid (PL) transfer from lipid emulsions to the human plasma lipoproteins.
| Fractions | Control | Post-heparin | Post-heparin + | THL | ||
| 3H-FC | 14C-PL | 3H-FC | 14C-PL | 3H-FC | 14C-PL | |
| LDL | 15 | 7 | 21 | 7 | 20 | 6 |
| (2-35) | (3-49) | (15-26) | (2-13) | (17-34) | (1-12) | |
| HDL | 13 | 31a | 18 | 50a,b | 15 | 39a,c |
| (7-28) | (18-47) | (12-36) | (35-68) | (8-33) | (23-48) | |
Percent transfer of 3H-FC and 14C-PL from lipid emulsion to LDL and HDL obtained from fasting human plasma control, post-heparin with and without addition of a lipase inhibitor, tetrahydrolipstatin (THL) incubated for 30 minutes at zero (ice) and at 37°C. Plasma lipoproteins were fractionated by FPLC. Percent transfer was calculated as the difference between values obtained at 37°C and 0°C. Results are expressed as median (range), n=12. Mann-Whitney test: a: p< 0.001 (HDL vs. LDL) ; b: p<0.001 (post-heparin vs. control) and c: p<0.01 (post-heparin vs. post-heparin+THL).
The effect of plasma lipases on the PL transfer to HDL was also verified by incubating labeled EM with plasma from control and heparinized rats. Figure 1 shows the 14C-PL distribution in the LP fractions at zero (ice) or after 30 minutes at 37°C. In the control rat plasma (Fig. 1, upper panel) at time zero, the distribution of 14C-PL was 60, 13, 22 and 6% for VLDL, LDL, HDL and a fraction smaller than HDL (<HDL), respectively, while after 30 minutes at 37°C, the observed distribution was 22, 14, 57 and 8% for VLDL, LDL, HDL and fraction <HDL, respectively. It is interesting that PL transfer to HDL can be observed even at time zero. This may represent the combination of the spontaneous and facilitated transfer that occurs during and even after loading the reaction mixture to the FPLC column run at room temperature. Furthermore, rat plasma contains circulating lipases [25,26] that could be stimulating PL transfer in the control plasma. In the rat post-heparin plasma (Fig. 1, lower panel), the 14C-PL transfer was so much accelerated that the distributions at zero and after 30 minutes of incubation at 37°C were almost identical, that is, 12, 29.5, 32.5 and 26.5% at zero and 7, 31, 29.5 and 32.5% at 37°C for VLDL, LDL, HDL and fraction <HDL, respectively. HDL and the fraction smaller than HDL together accounted for 59 to 62% of the 14C-PL. Thus, comparing the PL moiety that remained in the emulsion fraction in the control at 37°C (Fig 1 upper, 22%) with the post-heparin plasma at 37°C (Fig 1 lower, 7%), we conclude that PL transfer to LDL and HDL fractions is highly stimulated by the enrichment of the rat plasma with the vascular lipases. We did not investigate the chemical nature of the fraction smaller than HDL but we speculate that it derives from HDL phospholipids or from the emulsion surface itself, and might be lysophospholipids/albumin or phospholipids/apo AI complexes.
Figure 1.
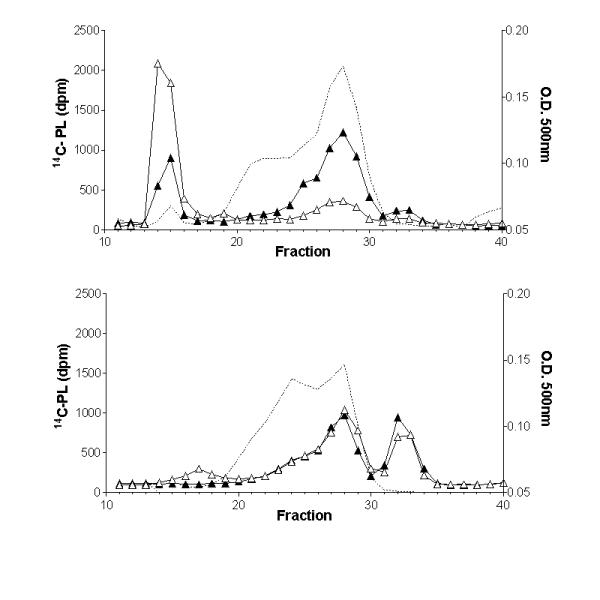
Influence of plasma lipases on the distribution of the 14C-PL from lipid emulsions into rat plasma lipoproteins. Control (upper panel) and heparin treated (lower panel) rat plasmas were incubated with lipid emulsion labeled with 14C-phospholipids (PL) at zero (on ice) (open triangles) or for 30 minutes at 37°C (closed triangles), and fractionated by fast protein liquid chromatography. Cholesterol mass in each fraction (O.D.500 nm) is shown in the dotted line profile.
In an attempt to evaluate the PL and FC transfer to HDL in a biological system, we injected the radioactive EM (3H-PL/14C -FC) into anesthetised functionally hepatectomized rats treated with saline (control) or heparin (LPL stimulated). The liver exclusion maneuver was done in order to minimize the differences in the residence time of the emulsion particles in the circulation of the control and LPL stimulated rats. The hepatectomy efficacy was verified by the amount of 3H-PL found in the livers at the end of the experiments; that was 1% and 3% of the injected dose in control and heparinized animals, respectively. There was a lower recovery of the labeled PL in the plasma of the heparinized animals (∼ 50%) than in control rats (∼ 90%). This suggests that the heparin stimulated lipolysis increased the plasma removal of PL by peripheral tissues. Table 2 shows the distribution of 3H-PL and 14C-FC from the EM into the plasma LP of control and heparinized hepatectomized rats. When intravascular lipolysis was stimulated by heparin, there was a 50% reduction of the 3H-PL content in the chylomicron (CM) + VLDL fraction (d<1.006) and a parallel increase in the LDL fraction that also includes remnant particles (d= 1.006 to 1.063). No significant difference was observed in the amount of 3H-PL found in the HDL fraction of control and heparinized hepatectomized rats. A similar result was observed in relation to the 14C-FC distribution, i.e., a reduction trend of 14C-FC in the CM+VLDL fraction (non significant) and a significant increase of 14C-FC transfer to the LDL fraction. However, in both cases, control and heparinized animals, the ratio 14C-FC/3H-PL differs markedly in the three LP fractions (Table 2). This indicates that, in vivo, the redistribution of the two emulsion surface lipids is independent one from another.
Table 2.
Influence of plasma lipases on the distribution of 14C-free cholesterol (FC) and 3H-phospholipid (PL) from lipid emulsions to plasma lipoproteins of hepatectomized rats in vivo.
| Fractions | Control | Heparinized | ||
| 14C-FC | 3H-PL | 14C-FC | 3H-PL | |
| CM+ VLDL | 61 | 36 | 47 | 16a |
| (53 - 74) | (27 - 61) | (43 - 55) | (14 - 22) | |
| LDL + remnants | 20 | 22 | 34b | 38c |
| (11 - 24) | (10 - 26) | (29 - 38) | (33 - 42) | |
| HDL | 19 | 42 | 18 | 46 |
| (15 - 23) | (29 - 47) | (14 - 22) | (40 - 51) | |
Percent distribution of 14C-FC and 3H-PL from lipid emulsion to VLDL, LDL and HDL plasma fractions from anesthetized hepatectomized rats treated with saline (control) or heparin after 30 minutes of an intra-arterial injection of the labeled emulsion. Lipoprotein fractions were obtained by ultracentrifugation. Results are expressed as median (range), n=4. Mann-Whitney test for heparin vs. control comparisons: a: 3H-PL - CM+VLDL, p < 0.03; b: 14C-FC - LDL, p < 0.03; c: 3H-PL - LDL, p < 0.03.
In order to verify the influence of CETP in the PL transfer to HDL, labeled emulsion was incubated with human plasma in the presence and absence of monoclonal antibody against CETP (TP2) and with plasma from CETP transgenic and non-transgenic mice. The CETP activity in these plasmas were 38 and 16% of cholesteryl ester transfer (4 h assay) respectively for human plasma without and with TP2, and 5 and 20% of cholesteryl ester transfer (2 h assay), respectively for nontransgenic and human CETP transgenic mice plasma. Table 3 shows that 14C-PL transfer from lipid emulsion to human HDL was reduced 40% (p<0.005) when CETP activity was inhibited 58%, indicating that CETP has PL transfer activity as previously reported [5]. However, the 14C-PL distribution in mice LP after incubation with the 14C-PL EM was almost identical for CETP transgenic and non-transgenic mice plasma (Fig. 2). The human CETP expressed in these mice might not efficiently interact with the endogenous mice HDL. To check this possibility an exogenous assay was performed using human HDL (240 ug cholesterol) as PL acceptor and minimal amounts of CETP transgenic and non-transgenic mice plasma (50 ul) as sources of CETP and of PLTP. Results obtained (data not shown) were identical to those of the endogenous assay (fig. 2). Therefore, human CETP in transgenic mice plasma does not contribute to PL transfer to either mice or human HDL.
Table 3.
Influence of CETP on the 14C-phospholipid (PL) transfer from lipid emulsions to the human plasma lipoproteins.
| Fractions | Control | +TP2 |
| LDL | 4 (0 - 6) | 0 (-5 - 1) |
| HDL | 30a (26 - 38) | 18a,b (16 - 27) |
Percent transfer of 14C-PL from lipid emulsion to LDL and HDL obtained from fasting human plasma without (control) and with CETP monoclonal antibody, TP2, incubated for 30 minutes at zero (ice) and at 37°C. Plasma lipoproteins were fractionated by FPLC. Percent transfer was calculated as the difference between values obtained at 37°C and 0°C. Results are expressed as median (range), n=6. Mann-Whitney test: a: p<0.001 (HDL vs. LDL), b: p<0.05 (TP2 vs. control).
Figure 2.
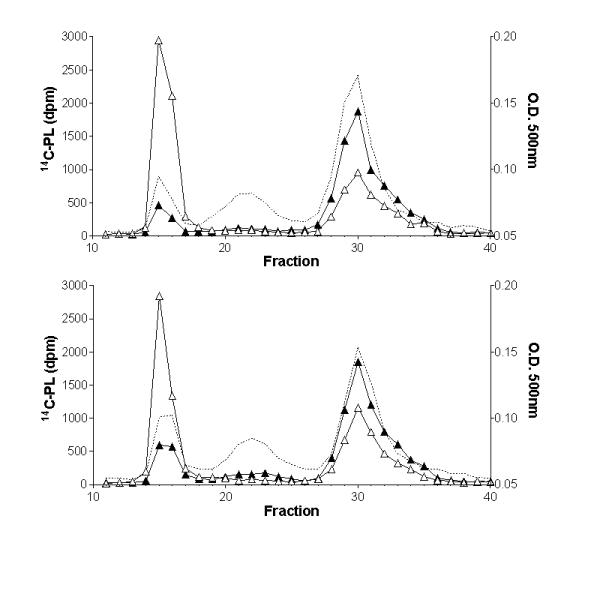
Influence of CETP on the distribution of the 14C-PL from lipid emulsions into mice plasma lipoproteins. Control non-transgenic (upper panel) and CETP transgenic (lower panel) mice plasmas were incubated with lipid emulsion labeled with 14C-phospholipids (PL) at zero (on ice) (open triangles) or for 30 minutes at 37°C (closed triangles), and fractionated by fast protein liquid chromatography. Cholesterol mass in each fraction (O.D.500 nm) is shown in the dotted line profile.
Discussion
Several studies, using different experimental approaches, have shown that the transfer of surface components of TG-rich lipoproteins during their intravascular metabolism is important to determine both level and chemical composition of the HDL subfractions. The present work has confirmed and extended previous observations showing that human and rat plasma lipoprotein lipases stimulate PLTP mediated PL transfer from TG-rich particles to HDL but do not influence the FC transfer process. Some of the previous studies [11,35,36,37,38] that used purified exogenous lipases in in vitro incubations with isolated LP displayed a potent stimulation of the PL transfer to HDL. In order to prepare a model that would mirror a physiological system more closely, we have used whole plasma and maximal endogenous lipases activity through heparin administration in vivo as well as in vitro. It is possible that other proteins released by heparin in the vascular bed may have played a role in stimulating PL transfer. The role of circulating lipases was confirmed through the use of a lipase inhibitor (THL), which abolished the stimulating effect of lipase-enriched plasma on the PL transfer to HDL (Tab. 1). According to these experiments it is likely that during fasting state, where no circulating lipases are detectable, all PL transfer results from the action of the lipid transfer proteins, PLTP and CETP, while in a post-prandial state, when lipases expectedly are more active, the PL transfer to HDL could be raised by 60% or more due to a greater substrate supply .
Noteworthy the in vitro PL transfer to HDL in the basal plasmas of rat (fig. 1, 82%) and mouse (fig. 2, 57%) was higher than in the human's (Tab. 1, 31%) and could be ascribed to the high levels of circulating lipases [26,39,40] and PLTP [10] found in those species. This could also explain the positive correlation between the lipoprotein lipase activity and HDL concentration in human plasma [20,21,22] but not in mice with genetically modified expression of LPL [23,24,28].
The in vivo studies showed that the EM PL transfer to HDL did not differ in control and in LPL stimulated (heparin treated) hepatectomized rats. Instead, the LDL density fraction was PL enriched in the heparinized animals. This could be explained by several and not exclusive possibilities. First, the lipolysis stimulation by heparin generates more remnants of the EM that would float in the same density range as LDL (1.006 - 1.063). Second, the rat plasma fraction smaller than HDL that appeared in the in vitro incubations with post-heparin plasma (Fig. 1) could also occur in vivo and float in the same density range as LDL (1.006 -1.063). In this regard, O'Meara et. al. [41] had shown that small HDL particles from heparinized hypertriglyceridemic subjects, identified in non-denaturing gel electrophoresis and by electron microscopy, floated after ultracentrifugation in a less dense range. Those authors had considered it an aberrant result of the ultracentrifugation technique. Third, the diminished availability of PL donor particles in the plasma of the heparinized rats (yield of 50%) as compared to control rats (yield of 90%) would be responsible for the apparent lack of stimulation of the PL transfer to the HDL fraction. Finally, other in vivo metabolic fates of PL would compete for the transfer process to plasma HDL particles in LPL stimulated animals.
The emulsion FC transfer to HDL was about 40% that of PL in both, in vitro (Tab. 1) and in vivo (Tab. 2) studies and it was not influenced by increased rate of intravascular lipolysis. These results suggest that the FC transfer is a slower, probably passive, process distinct from the facilitated PL transfer mechanism [8,9]. Also, these results challenge the possibility that new HDL particles are made from the EM surface peeling off during lipolysis because the relative PL/FC ratio was higher in HDL than in the CM+VLDL and emulsion fraction. Others also have shown that FC transfer to HDL is a slow process: FC increases in the HDL fraction only 5 to 8 h after a fat meal [19] or only after 2 h incubation of HDL with VLDL and purified rat heart LPL [36].
We have also confirmed previous studies [5] claiming that PL transfer from TG-rich particles to human HDL is facilitated by CETP, since its partial inhibition significantly reduced the PL transfer (Table 3). However, human CETP expressed in the transgenic mice plasma had no effect whatsoever on the PL transfer to mouse plasma lipoproteins or to human HDL. This lead us to admit that the mouse plasma PL transfer activity is so potent that some additional protein (CETP) with PL transfer activity would be irrelevant in an already saturated in vitro system.
In summary, the present findings indicate unequivocally the importance of the intravascular lipolytic mechanisms for the PLTP and CETP facilitated PL, but not FC, transfer process from TG-rich particles to HDL. PL enriched HDL would be more efficient in promoting FC efflux from cell membranes, hence accelerating the reverse cholesterol transport. These may provide the basis for the mechanism that accounts for the inverse correlation between HDL and TG plasma levels found in epidemiological studies in human populations as well as in several circumstances where plasma lipid levels are modified by pharmacological and dietary means.
Materials and Methods
Lipid emulsions
Cholesterol (FC), cholesteryl oleate (CO) and triolein (TO) were obtained from NuCheck Prep (Elysian, MN, USA) and lecithin (PL) from Lipid Products (Surrey,UK). They were more than 99% pure as tested by thin layer chromatography. Lipid mixtures (2% FC, 6% CO, 23%PL and 69%TO by weight) together with 130 μCi of L-α-dioleoyl [1-14C]-phosphatydylcholine and 25 μCi of [1,2-3H(N)]-cholesterol (New England Nuclear, Boston, MA, USA) were sonicated in 2.785 M NaCl solution (d=1.101 g/ml) utilizing a Branson Cell Disruptor (Branson Ultrasonics Corp., Danbury, CT, USA), model B30, 1 cm probe, with continuous output of 70-80 W, at aproximately 55°C, for 30 minutes, under N2 flow. Triglyceride-rich particles were purified after discontinuous gradient ultracentrifugation of NaCl solutions with densities 1.065, 1.020 and 1.006 g/ml. A first step of 12000 rpm for 15 min in a SW41 Beckman rotor at 22°C was performed to discard the floating coarse lipid. After replacing the 1.006 solution, the gradient was again centrifuged at 36000 rpm, for 30 min at 22°C and the emulsion particles were recovered from the top layer. The lipid emulsion composition achieved was: 1% FC, 4% CO, 14% PL and 81% TO. These particles resemble native chylomicrons [29]. By gel filtration (FPLC), 3H-FC, 14C-PL and triglyceride co-eluted as only one peak corresponding to the plasma VLDL size fraction (fractions # 13 to 17) on a HR 10/30 superose 6 column (Pharmacia Biotech, Uppsala, Sweden).
Sources of plasma
Human blood samples from 12 fasted healthy volunteers (5 men and 7 women, total cholesterol and triglycerides < 200 mg/dl), were drawn on EDTA, pre (basal) and 10 min after an I.V. bolus injection of heparin (100 U/kg BW). Male Wistar rats, approximately 300 g, had their carotid arteries cannulated under pentobarbital anesthesia. After recovery, they received saline (control) or heparin (LPL stimulated) and after 10 min they were exsanguinated on EDTA. Adult male C57Bl6 and human CETP transgenic mice (line 5203), derived from the colony of Dr AR Tall's Laboratory (Columbia University, NY, USA) were bled with heparinized hematocrit capillary tubes in the retro-orbital plexus under ketamine anesthesia (Vetarnacol, Konig, SP, Brazil). All plasmas were obtained by centrifugation at 2000 rpm in a Sorval RT6000B refrigerated centrifuge and freshly used.
Free cholesterol (FC) and phospholipid (PL) transfer assay
Control and treated plasmas (post-heparin, post-heparin + lipases inhibitor and control + CETP monoclonal antibody, TP2, provided by Dr. AR Tall) were incubated with lipid emulsion (∼ 700 μg of triolein/ml) labeled with 3H-FC (106 dpm/ml) and 14C-PL (2 × 105 dpm/ml) at zero (on ice, "time zero") and for 30 minutes at 37°C. Plasma lipoproteins were next separated by fast protein liquid chromatography (FPLC) as described by Jiao et al. [31]. Briefly, plasma samples (200 μl) were fractionated on a HR 10/30 Superose 6 column (Pharmacia Biotech, Uppsala, Sweden) using a constant flow of 0.5 ml/min of tris-buffered saline, pH 7.2. Sixty fractions of 0.5 ml were automatically collected. 3H- and 14C- dpm of each FPLC fraction was determined by liquid scintilation in a beta counter Beckman LS6000TA. As we have measured only radioactive FC and PL, the term "transfer" is used to describe either net transfer or exchange process among LP. Total cholesterol was also determined in the fractions #10 to 40 by an enzymatic assay in an automatic analyzer Cobas (F. Hoffman-La Roche, Basileia, Switzerland) using Boehringer Mannheim reagents (Mannheim, Germany).
In Vivo studies
Male Wistar rats, weighing ∼ 300 g were anesthetised with pentobarbital ip (50 mg/Kg BW). The right carotid artery was cannulated with a PE 50 siliconized catheter and after laparostomy, the liver hilum was ligated. Physiologic solution (control) or heparin (250 U/Kg BW) in a final volume of 0.25 ml was injected through the carotid catheter. After 10 minutes, labeled lipid emulsion (4 × 105 dpm of 3H-PL and 7 × 105 dpm of 14C-FC) was injected intra-arterially. After 30 minutes, the animals were exsanguinated by the carotid catheter and plasma lipoproteins were immediately separated by ultracentrifugation in a discontinuous gradient. Plasmas were adjusted to density (d) 1.21 g/ml with solid KBr and overlayed with solutions of d=1.063 and 1.006 g/ml and centrifuged for 24 h in a SW 41 rotor, at 4°C, 100000 × g, in a L8 Beckman ultracentrifuge. Lipoproteins fractions were collected from the top to bottom by vacuum as follow: 1.5 ml VLDL (d<1.006), 2.5 ml LDL (d = 1.006 - 1.063) and 7.5 ml HDL (d>1.063). Radioactivity was determined in aliquots from each lipoprotein fraction.
Intravascular Lipases Activity
Total lipase activity was determined according to Ehnholm & Kuusi [32]. Briefly, overnight fasted human plasmas, collected pre (basal) and 10 minutes after heparin I.V. injection (100 U/Kg body weight), were incubated with a 3H-triolein/arabic gum substrate ([9,10 3H (N)]-triolein, New England Nuclear, Boston, MA) in 0.2 M Tris-HCl buffer, pH 8.5, 37ºC, during 1 hour. Hepatic lipase (HL) activity was determined in tubes where the lipoprotein lipase (LPL) was inhibited by 2 M NaCl. The hydrolyzed labeled free fatty acids were extracted with methanol / chloroform / heptane (1.4 : 1.25 : 1), 0.14 M K2CO3 / H3BO3, pH 10.5, dried under N2, and their radioactivity was determined in a liquid scintillation solution in a LS6000 Beckman Beta Counter. The LPL activity was calculated as the difference between the total lipase and the hepatic lipase activities.
Cholesteryl ester transfer protein activity assay
A mixture of human VLDL and LDL protein (100 μg) were incubated with 10000 dpm of human HDL3 labeled with [14C]-cholesteryl ester (CE) [33] and 5 μl of diluted CETP transgenic mice plasma or undiluted human plasma as the source of CETP in a final volume of 100 μl. Blanks were prepared with tris/saline/EDTA buffer (10 mM/140 mM/1 mM), pH 7.4, and negative controls with non-transgenic mice plasma. The incubations were carried out at 37°C for 2 or 4 hours. After these periods, the apo B containing lipoproteins were precipitated using a mixture of 1.6% dextran sulfate / 1 M MgCl2 solution (1:1) and the radioactivity was measured in the remaining supernatant in scintillation solution Ultima Gold (Eastman Kodak Co., NY) in a LS6000 Beckman Beta Counter. The % CE transferred from [14C]-CE-HDL to VLDL+LDL was calculated as: (dpm in the blank tube - dpm in the plasma sample / dpm in the blank tube) × 100.
Statistical analysis
All comparisons were analysed by the non-parametric Mann-Whitney test using the GraphPad Prism, version 2.01 (1996) program. Differences were considered significant when p<0.05.
Acknowledgments
Acknowledgements
We are grateful to Dr. Alan R. Tall for kindly providing some human CETP transgenic mice and the CETP monoclonal antibody, TP2. This study was supported by Brazilian grants from FAPESP, CNPq and Pronex/FINEP. V.S.N was a MSc Student in the program of the Dept of Biochemistry, Escola Paulista de Medicina da Universidade Federal de São Paulo, SP, Brasil.
Contributor Information
Valéria S Nunes, Email: lipideq@usp.br.
Eder CR Quintão, Email: quintao@originet.com.br.
Patrícia M Cazita, Email: pmcazita@hotmail.com.
Lila M Harada, Email: lilamh@usp.br.
Eliana C de Faria, Email: cotta@fcm.unicamp.br.
Helena CF Oliveira, Email: ho98@unicamp.br.
References
- Backer G, Bacquer D, Konitzer M. Epidemiological aspects of high density lipoprotein cholesterol. Atherosclerosis. 1998;137:S1–S6. doi: 10.1016/S0021-9150(97)00310-9. [DOI] [PubMed] [Google Scholar]
- Stein O, Stein Y. Atheroprotective mechanisms of HDL. Atherosclerosis. 1999;144:285–301. doi: 10.1016/S0021-9150(99)00065-9. [DOI] [PubMed] [Google Scholar]
- Tall AR. Plasma lipid transfer proteins. Annu Rev Biochem. 1995;64:235–257. doi: 10.1146/annurev.biochem.64.1.235. [DOI] [PubMed] [Google Scholar]
- Hesler B, Tall AR, Swenson TL, Weech PK, Marcel YL, Milne RW. Monoclonal antibody to the Mr 74000 cholesterol ester transfer protein neutralize all of the cholesterol ester and triglyceride transfer activities in human plasma. J Biol Chem. 1988;263:5020–5023. [PubMed] [Google Scholar]
- Swenson TL, Brocia RW, Tall AR. Plasma cholesteryl ester transfer protein has binding sites for neutral lipids and phospholipids. J Biol Chem. 1988;263:5150–5157. [PubMed] [Google Scholar]
- Lagrost L, Athias A, Gambert P, Lallemant C. Comparative study of phospholipid transfer activities mediated by cholesteryl ester transfer protein and phospholipid transfer protein. J Lipid Res. 1994;35:825–835. [PubMed] [Google Scholar]
- Tato F, Vega GL, Grundy SM. Determinants of plasma HDL-cholesterol in hypertriglyceridemic patients. Arterioscler Thromb Vasc Biol. 1997;17:56–63. doi: 10.1161/01.atv.17.1.56. [DOI] [PubMed] [Google Scholar]
- Tall AR, Forester LR, Bongiovanni GL. Facilitation of phosphatidylcholine transfer into HDL lipoproteins by an apolipoprotein in the density 1.20 - 1.26 g/ml fraction of plasma. J Lipid Res. 1983;24:277–289. [PubMed] [Google Scholar]
- Albers JJ, Tollefson JH, Chen CH, Steinmetz A. Isolation and characterization of human plasma lipid transfer proteins. Arteriosclerosis. 1984;4:49–58. doi: 10.1161/01.atv.4.1.49. [DOI] [PubMed] [Google Scholar]
- Guyard-Dangremont V, Desrumaux C, Gambert P, Lallemant C, Lagrost L. Phospholipid and cholesteryl ester transfer activities in plasma from 14 vertebrate species. Relation to atherogenesis susceptibility. Comp Biochem Physiol Biochem Mol Biol. 1998;120:517–525. doi: 10.1016/S0305-0491(98)10038-X. [DOI] [PubMed] [Google Scholar]
- Tall AR, Krumholz S, Olivecrona T, Deckelbaum RJ. Plasma phospholipid transfer protein enhances transfer and exchange of phospholipids between VLDL and HDL lipoproteins during lipolysis. J Lipid Res. 1985;26:842–851. [PubMed] [Google Scholar]
- Nishida HI, Nishida T. Phospholipid transfer protein mediates transfer of not only phosphatidylcholine but also cholesterol from phosphatidylcholine-cholesterol vesicles to high density lipoproteins. J Biol Chem. 1997;272:6959–6964. doi: 10.1074/jbc.272.11.6959. [DOI] [PubMed] [Google Scholar]
- Lagrost L, Desrumaux C, Masson D, Deckert V, Gambert P. Structure and function of the plasma phospholipid transfer protein. Curr Opin Lipidol. 1998;9:203–209. doi: 10.1097/00041433-199806000-00004. [DOI] [PubMed] [Google Scholar]
- Albers JJ, Tu AY, Paigen B, Chen H, Cheung MC, Marcovina SM. Transgenic mice expressing human phospholipid transfer protein have increased HDL/non-HDL cholesterol ratio. Int J Clin Lab Res. 1996;26:262–267. doi: 10.1007/BF02602960. [DOI] [PubMed] [Google Scholar]
- Foger B, Santamarina-Fojo S, Shamburek RD, Parrot CL, Talley GD, Brewer HB., Jr Plasma phospholipid transfer protein. Adenovirus-mediated overexpression in mice leads to decreased plasma high density lipoprotein (HDL) and enhanced hepatic uptake of phospholipids and cholesteryl esters from HDL. J Biol Chem. 1997;272:27393–27400. doi: 10.1074/jbc.272.43.27393. [DOI] [PubMed] [Google Scholar]
- Redgrave TG, Small DM. Quantitation of the transfer of surface phospholipid of chylomicrons to the HDL lipoprotein fraction during the catabolism of chylomicrons in the rat. J Clin Invest. 1979;64:162–171. doi: 10.1172/JCI109435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tall AR, Green PH, Glickman RM, Riley JW. Metabolic fate of chylomicron phospholipids and apoproteins in the rat. J Clin Invest. 1979;64:977–989. doi: 10.1172/JCI109564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tall AR, Blum CB, Forester GP, Nelson CA. Changes in the distribution and composition of plasma HDL liproteins after ingestion of fat. J Biol Chem. 1982;257:198–207. [PubMed] [Google Scholar]
- Groot H, Scheek LM. Effects of fat ingestion on HDL profiles in human sera. J Lipid Res. 1984;25:684–692. [PubMed] [Google Scholar]
- Brunzell JD. Familial lipoprotein lipase deficiency and other causes of the chylomicronemia syndrome. Scriver, CR, Beaudet, AL, Sly, WS, ed Metabolic & Molecular Bases of Inherited Disease McGraw-Hill Inc, New York, 7th ed, 1995. pp. 1913–1932.
- Bijvoet S, Gagne SE, Moorjani S, Gagne C, Henderson HE, Fruchart JC, Dallongeville J, Alaupovic P, Prins M, Kastelein JJ, Hayden MR. Alterations in plasma lipoproteins and apolipoproteins before the age of 40 in heterozygotes for lipoprotein lipase deficiency. J Lipid Res. 1996;37:640–650. [PubMed] [Google Scholar]
- Kuusi T, Ehnholm C, Viikari J, Harkonen R, Vartiainen E, Puska P, Taskinen M-R. Postheparin plasma lipoprotein and hepatic lipase are determinants of hypo- and hyperalphalipoproteinemia. J Lipid Res. 1989;30:1117–1126. [PubMed] [Google Scholar]
- Liu S, Jirik FR, LeBoeuf RC, Henderson H, Castellani LW, Lusis AJ, Ma Y, Forsythe IJ, Zhang H, Kirk E. Alteration of lipid profiles in plasma of transgenic mice expressing human lipoprotein lipase. J Biol Chem. 1994;269:11417–11424. [PubMed] [Google Scholar]
- Weinstock PH, Bisgaier CL, Aalto-Setala K, Radner H, Ramakrishnan R, Levak-Frank S, Essenburg AD, Zechner R, Breslow JL. Severe hypertriglyceridemia, reduced high density lipoprotein, and neonatal death in lipoprotein lipase knockout mice. Mild hypertriglyceridemia with impaired very low density lipoprotein clearance in heterozygotes. J Clin Invest. 1995;96:2555–2568. doi: 10.1172/JCI118319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Applebaum-Bowden D, Kobayashi J, Kashyap VS, Brown DR, Berard A, Meyn S, Parrott C, Maeda N, Shamburek R, Brewer HB, Jr, Santamarina-Fojo S. Hepatic lipase gene therapy in hepatic lipase-deficient mice. Adenovirus-mediated replacement of a lipolytic enzyme to the vascular endothelium. J Clin Invest. 1996;97:799–805. doi: 10.1172/JCI118479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillett MP, Vieira EM, Dimenstein R. The phospholipase activities present in preheparin mouse plasma are inhibited by antiserum to hepatic lipase. Int J Biochem. 1993;25:449–453. doi: 10.1016/0020-711X(93)90639-V. [DOI] [PubMed] [Google Scholar]
- Ha YC, Barter PJ. Differences in plasma cholesteryl ester transfer activity in sixteen vertebrate species. Comp Biochem Physiol B. 1982;71:265–269. doi: 10.1016/0305-0491(82)90252-8. [DOI] [PubMed] [Google Scholar]
- Clee SM, Zhang H, Bissada N, Miao L, Ehrenborg E, Benlian P, Shen GX, Angel A, LeBoeuf RC, Hayden MR. Relationship between lipoprotein lipase and HDL lipoprotein cholesterol in mice: modulation by cholesteryl ester transfer protein and dietary status. J Lipid Res. 1997;38:2079–2089. [PubMed] [Google Scholar]
- Oliveira HCF, Hirata MH, Redgrave TG, Maranhão RC. Competition between chylomicrons and their remnants for plasma removal: a study with artificial emulsion models of chylomicrons. Biochim Biophys Acta. 1988;958:211–217. doi: 10.1016/0005-2760(88)90179-8. [DOI] [PubMed] [Google Scholar]
- Nakandakare ER, Lottenberg SA, Oliveira HCF, Bertolami MC, Vasconcelos KS, Sperotto G, Quintão EC. Simultaneous measurements of chylomicron lipolysis and remnant removal using a doubly labeled artificial lipid emulsion: studies in normolipidemic and hyperlipidemic subjects. J Lipid Res. 1994;35:143–152. [PubMed] [Google Scholar]
- Jiao S, Cole TG, Kitchens RT, Pfleger B, Schonfeld G. Genetic heterogeneity of lipoproteins in inbred strains of mice: analysis by gel-permeation chromatography. Metabolism. 1990;39:155–160. doi: 10.1016/0026-0495(90)90069-o. [DOI] [PubMed] [Google Scholar]
- Ehnholm C, Kuusi T. Preparation, characterization and measurement of hepatic lipase. Methods Enzymol. 1986;129:716–738. doi: 10.1016/0076-6879(86)29101-6. [DOI] [PubMed] [Google Scholar]
- Oliveira HCF, Quintão EC. 'In vitro' cholesteryl ester bidirectional flow between high-density lipoproteins and triglyceride-rich emulsions: effects of particle concentration and composition, cholesteryl ester transfer activity and oleic acid. J Biochem Biophys Methods. 1996;32:45–57. doi: 10.1016/0165-022X(94)00060-Q. [DOI] [PubMed] [Google Scholar]
- Huff MW, Miller DB, Wolf BM, Connelly PW, Sawyez CG. Uptake of hypertriglyceridemic VLDL and their remnants by HepG2 cells: the role of lipoprotein lipase, hepatic triglyceride lipase, and cell surface proteoglycans. J Lipid Res. 1997;38:1318–1333. [PubMed] [Google Scholar]
- Marques-Vidal P, Jauhiainen M, Metso J, Ehnholm C. Transformation of HDL2 particles by hepatic lipase and phospholipid transfer protein. Atherosclerosis. 1997;133:87–96. doi: 10.1016/S0021-9150(97)00120-2. [DOI] [PubMed] [Google Scholar]
- Murdoch SJ, Breckenridge WC. Effect of lipid transfer proteins on lipoprotein lipase induced transformation of VLDL and HDL. Biochim Biophys Acta. 1996;1303:222–232. doi: 10.1016/0005-2760(96)00105-1. [DOI] [PubMed] [Google Scholar]
- Murdoch SJ, Breckenridge WC. Influence of lipoprotein lipase and hepatic lipase on the transformation of VLDL and HDL during lipolysis of VLDL. Atherosclerosis. 1995;118:193–212. doi: 10.1016/0021-9150(95)05606-8. [DOI] [PubMed] [Google Scholar]
- Patsch JR, Gotto AM, Jr, Olivercrona T, Eisenberg S. Formation of HDL2- like particles during lipolysis of VLDL in vitro. Proc Natl Acad Sci USA. 1978;75:4519–4523. doi: 10.1073/pnas.75.9.4519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillett MP, Costa EM, Owen JS. The phospholipase activities present in preheparin mouse plasma are inhibited by antiserum to hepatic lipase. Biochim Biophys Acta. 1980;617:237–244. doi: 10.1016/0005-2760(80)90166-6. [DOI] [PubMed] [Google Scholar]
- Peterson J, Bengtsson-Olivecrona G, Olivecrona T. Mouse preheparin plasma contains high levels of hepatic lipase with low affinity for heparin. Biochim Biophys Acta. 1986;87:865–870. doi: 10.1016/0005-2760(86)90344-9. [DOI] [PubMed] [Google Scholar]
- O'Meara NM, Cabana VG, Lukens JR, Loharikar B, Forte TM, Polonsky KS, Getz GS. Heparin-induced lipolysis in hypertriglyceridemic subjects results in the formation of atypical HDL particle. J Lipid Res. 1994;35:2178–2190. [PubMed] [Google Scholar]