

Abstract
In an analysis of 47 aerobic myxobacterial strains, representing 19 genera in suborders Cystobacterineae, Nannocystineae, Sorangiineae, and a novel isolate, “Aetherobacter” SBSr008, an enormously diverse array of fatty acids (FAs) was found. The distribution of straight-chain fatty acids (SCFAs) and branched-chain fatty acids (BCFAs) supports the reported clustering of strains in the phylogenetic tree based on 16S rRNA genes. This finding additionally allows the prediction and assignment of the novel isolate SBSr008 into its corresponding taxon. Sorangiineae predominantly contains larger amounts of SCFA (57 to 84%) than BCFA. On the other hand, Cystobacterineae exhibit significant BCFA content (53 to 90%), with the exception of the genus Stigmatella. In Nannocystineae, the ratio of BCFA and SCFA seems dependent on the taxonomic clade. Myxobacteria could also be identified and classified by using their specific and predominant FAs as biomarkers. Nannocystineae is remarkably unique among the suborders for its absence of hydroxy FAs. After the identification of arachidonic (AA) FA in Phaselicystidaceae, eight additional polyunsaturated fatty acids (PUFAs) belonging to the omega-6 and omega-3 families were discovered. Here we present a comprehensive report of FAs found in aerobic myxobacteria. Gliding bacteria belonging to Flexibacter and Herpetosiphon were chosen for comparative analysis to determine their FA profiles in relation to the myxobacteria.
INTRODUCTION
Myxobacteria are one of nature's “talented” and widely distributed microorganisms commonly found in both terrestrial and aquatic ecosystems. They are Gram-negative, rod-shaped eubacteria famous for their unique developmental cycle, culminating in the formation of multicellular fruiting bodies (see Fig. S1 in the supplemental material), which serve as an important basis for myxobacterial classification (39). This group (Myxococcales) has also gained attention and fame for production of novel anti-infective drugs and chemotherapeutic agents with uncommon modes of action (53, 54). Surprisingly, myxobacteria were also described for their potential to produce polyunsaturated fatty acids (PUFAs). Although these are rare in bacteria, their production has already been detected in the marine myxobacterial genera Plesiocystis, Enhygromyxa, and Haliangium (8, 19, 20) and recently in the soil isolate Phaselicystis (12).
PUFAs are commercially valuable and essential for human health (50, 52). Omega-3 PUFAs, particularly eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), are of special significance to pharmaceutical and food industry. These fatty acids (FAs) are commonly used as supplements in food and dairy products, and as a drug (e.g., for treatment of hypertriglyceridemia). They play a role in the prevention of many heart-associated diseases (50) and are involved in many immune-inflammatory reactions (9) and brain development (14). Due to their significant benefits to human health, demands are constantly increasing (50). Alternative sources have been explored with various degrees of success, such as the straminopiles Schizochytrium, Thraustochytrium, and Saprolegniales; the fungal Entomophthorales (46); the marine piezophilic bacteria Shewanella, Colwellia, Moritella, and Psychroflexus (5, 36, 37); and poikilothermic animals, deep-sea fish, and arctic invertebrates (24, 56).
Surprisingly large amounts of EPA and DHA were found in one of our novel Sorangiineae isolates, “Aetherobacter” SBSr008 (49), described below, prompting us to determine their presence and distribution in aerobic myxobacteria and in morphologically related gliding, nonfruiting bacteria. In order to establish chemo-phylogeny correlations, different aspects of fatty acid content (e.g., FA types, FA ratio, and major markers) were considered in relation to taxonomic clades or taxa. The overall findings update and expand the previous study on FAs of Nannocystis (Nannocystineae), Myxococcus, Cystobacter, Stigmatella (Cystobacterineae), and Sorangium (Sorangiineae) (7). This study also aims to analyze, in addition to DHA and EPA, other polyunsaturated FAs found in myxobacteria.
MATERIALS AND METHODS
Bacterial strains and cultivation.
The type and neotype strains chosen to represent the whole aerobic myxobacteria are listed in Table S1 in the supplemental material. The majority were obtained from our collection at the Helmholtz Centre for Infection Research (HZI), Braunschweig, Germany. Archangium gephyra DSM2261T, Corallococcus coralloides DSM2259T, Cystobacter (Angiococcus) disciformis DSM52716T, Enhygromyxa salina DSM 15217T, Haliangium ochraceum DSM 14365T, Haliangium tepidum DSM 14436T, proposed neotype Melittangium lichenicola DSM14877T (30), Myxococcus fulvus DSM16525T, Myxococcus virescens DSM2260T, Myxococcus xanthus DSM16526T, Plesiocystis pacifica DSM14875T, Stigmatella aurantiaca DSM17044T, and Stigmatella erecta DSM16858T were purchased from the German Culture Collection (DSMZ) in Braunschweig. The novel isolate Aetherobacter SBSr008 was obtained from Helmholtz Institute for Pharmaceutical Research (HIPS) in Saarbrücken, Germany. Strains of the gliding bacteria Herpetosiphon (Hp g287, Hp g336, Hp g383, and Hp g454) and Flexibacter (Flex 23 and Flex 7014) were generously provided by Klaus Gerth, Microbial Drugs, HZI.
Myxobacteria were cultivated in 50 ml of medium in 300-ml flasks shaken at 170 rpm at 30°C (37°C for Haliangium tepidum). The media used are as follows: MD1 (45) for Archangium, Corallococcus, Hyalangium, Melittangium, Myxococcus, Nannocystis, Stigmatella, Kofleria, Cystobacter disciformis, and C. armeniaca; M medium (35) for all other Cystobacter strains; TS6 medium (1% tryptone [Difco], 0.6% soluble starch [Roth], 25 mM HEPES, pH 7.0) for Pyxidicoccus fallax; VY/2 (10) for Byssovorax (with maltose) (28, 40), Polyangium, Chondromyces, and Jahnella; HS medium (27) for Sorangium; VY/4-SWS (with 50% of the concentration of baker's yeast as in VY/2-SWS) (19, 20) for Enhygromyxa and Plesiocystis; and CY-SWS (18) for Haliangium. Strains of Flexibacter and Herpetosiphon were cultivated in LB broth and VY/2 agar, respectively. Phaselicystis flava and Aetherobacter SBSr008 were not cultivated in this study; instead, FA data for these species were obtained from earlier studies (11, 49).
Fatty acid extractions, GC-MS, and data analysis.
Cystobacter miniatus, M. lichenicola, and strains of Herpetosiphon did not grow well in their corresponding liquid media; hence, only their biomass scraped off on VY/2 agar plates was used for extraction. For other strains, 2-ml aliquots of broth cultures were centrifuged (5,000 rpm, 5 min, 20°C) and cell pellets were dried completely with a speed vacuum for 30 min at 60°C. FA extraction by the fatty acid methyl ester (FAME) method was performed accordingly (3), except that the whole-cell pellet was not resuspended in NaCl solution prior to drying in a vacuum concentrator. Gas chromatography-mass spectrometry (GC-MS) analysis of derivatized samples was also performed according to a previous study (3), with some changes in the parameter settings. The column temperature was kept at 130°C for 2.5 min and then increased to 240°C at 5°C/min. The mass selective detector was operated in scan mode, with a scanning mass range of m/z 40 to 500.
16S rRNA gene amplification and phylogenetic tree construction.
Extraction and purification of genomic DNA, amplification and sequencing of 16S rRNA genes, sequence alignments, calculations of distance matrices and bootstrap values for 1,000 replicates, and construction of phylogenetic tree were performed as previously described (12). The neighbor-joining trees in Fig. 2 are based on type strains, the holotype strain, and the novel isolate SBSr008. GenBank accession numbers of myxobacterial sequences and outgroup Desulfovibrio desulfuricans are listed in Table S1 in the supplemental material. The 11 sequenced strains (GenBank accession no. GU249615, GU207872 to GU207876, and GU207878 to GU207882) were previously described (12). To establish chemo-phylogeny correlations in the 16S DNA tree, different aspects of FA analysis were considered, which include their ratio (e.g., straight-chain FA [SCFA]/branched-chain FA [BCFA] ratio), types (hydroxy FA and PUFAs), and another determinant marker (C17:1 2-OH). In addition, major FAs common to specific genera or clades were assigned based on their highest (2–5) percentage values. The same considerations were applied in some species that do not share common major FAs.
Fig. 2.
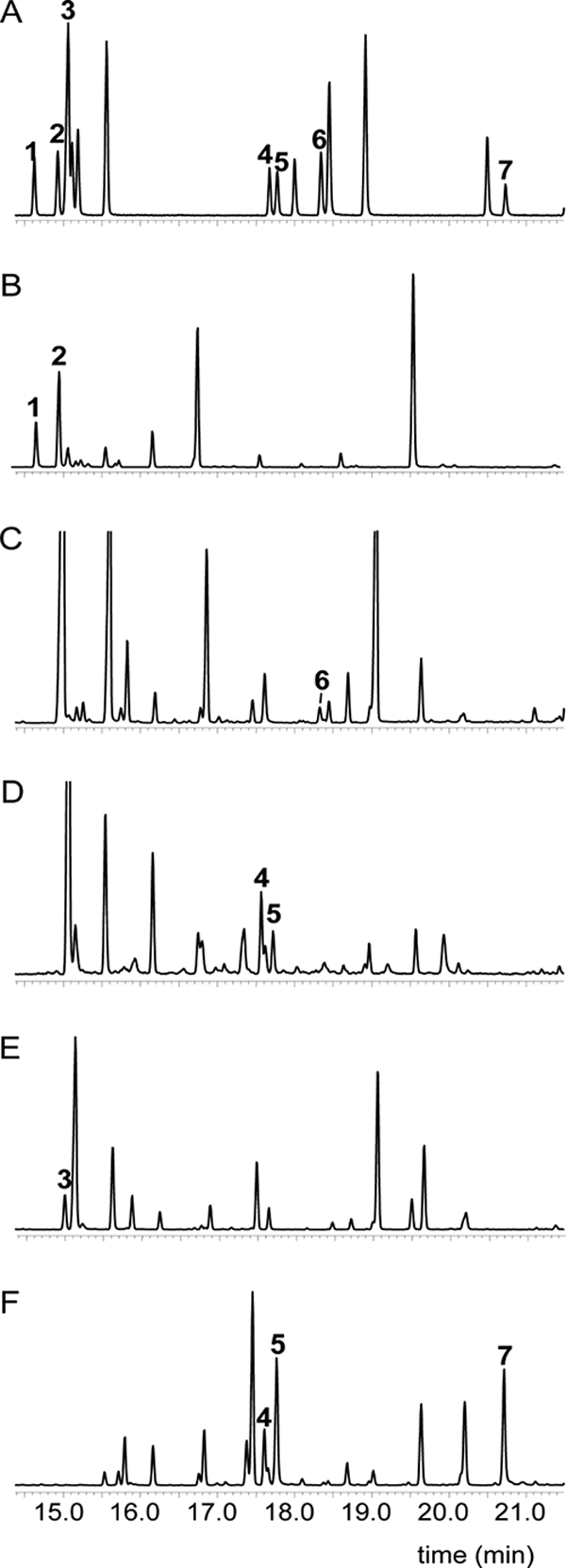
GC-MS chromatograms of PUFA-producing myxobacteria. (A) FAME reference standard mixture; (B) Myxococcus fulvus (ATCC 25946T) DSM1625T; (C) Sorangium cellulosum So ce1871T (DSM14627T); (D) Enhygromyxa salina (SHK-1T) DSM15217T; (E) Chondromyces crocatus SBCm010; (F) Aetherobacter SBSr008. PUFAs identified in myxobacteria: 1, C18:3ω6,9,12, all cis, γ-linolenic acid (GLA); 2, C18:2ω6,9, all cis, linoleic acid; 3, C18:3ω3,6,9, all cis, α-linolenic acid (ALA); 4, C20:4ω6,9,12,15, all cis, arachidonic acid (AA); 5, C20:5ω3,6,9,12,15, all cis, eicosapentaenoic acid (EPA); 6, C20:2ω6,9, all cis, eicosadienoic acid (EDA); and 7, C22:6ω3,6,9,12,15,18, all cis, docosahexaenoic acid (DHA).
RESULTS AND DISCUSSION
Numerous FAs were detected by GC-MS in all 47 strains of myxobacteria studied. Thirty-two were identified in Nannocystineae and 44 in Cystobacterineae. In Myxococcaceae alone, from the latter suborder, 34 were identified, approximately the same number as reported earlier (1, 2, 6). Most surprising was the discovery of 49 FAs in Sorangiineae. To our knowledge, the only previous extensive analytical studies of FA profiles in this suborder were performed on strains of Sorangium cellulosum (So ce14 [7] and AJ 13585 [8]), and Phaselicystis flava (11). In contrast to the majority of other bacteria, which have only simple (only a few types) FAs (25), myxobacteria appeared far more creative in their biosynthesis of diverse FAs, including those rarely encountered in nature.
Moreover, the occurrence of FA types (see Fig. S2 in the supplemental material) was discovered to be correlated with taxonomic placements of the genera of myxobacteria. SCFAs were dominant in Sorangiineae and BCFAs in Cystobacterineae (except in Stigmatella), and both were found in Nannocystineae. Within Nannocystineae, the total BCFA content was higher than SCFA in the Kofleriaceae-Haliangiaceae clade and vice versa in the Nannocystaceae clade.
Suborder Cystobacterineae.
The suborder Cystobacterineae has the highest number of known species (39), all of which are far easier to isolate and maintain in culture than their counterparts in Nannocystineae and Sorangiineae. It is remarkable for its large amount of BCFAs (53 to 90%), except for Stigmatella, whose FA diversity also diverged from most members of the Cystobacterineae in the phylogenetic tree. This suggests that it might need to be accommodated in a separate family, as already considered in the preceding phylogenetic paper (12).
Myxococcus, Corallococcus, and Pyxidicoccus clusters.
Certain species of Corallococcus and Myxococcus have been used as the preferred models in most FA biosynthesis (1, 4, 42) and developmental studies (2, 13, 26, 41). In Pyxidicoccus, the FA composition was evaluated here for the first time. These three genera formed a phylogenetically coherent group with 99.5% bootstrap support. The M. virescens-M. xanthus-M. macrosporus clade had a much higher ratio of BCFAs to SCFAs than the M. fulvus-M. stipitatus clade (Table 1); this was reflected in their phylogenetic divergence (Fig. 1).
Table 1.
Fatty acid distribution in Cystobacterineae genera Pyxidicoccus, Myxococcus, and Corallococcus
| FA type | % of FA ina: |
|||||||
|---|---|---|---|---|---|---|---|---|
| P. fallax | M. fulvus | M. stipitatus | M. virescens | M. xanthus | M. macrosporus | C. coralloides | C. exiguus | |
| SCFAs | ||||||||
| C13:0 | 0.21 | |||||||
| C14:0 | 4.59 | 2.75 | 6.36 | 4.70 | 4.93 | 4.60 | 0.21 | |
| C14:1ω5c | 1.03 | 0.51 | 0.41 | 1.27 | 0.75 | 0.61 | ||
| C15:0 | 4.02 | 0.77 | 1.59 | 1.36 | 0.29 | 0.26 | 0.10 | 0.06 |
| C15:1 | 5.92 | 0.54 | 0.35 | 3.95 | 0.34 | |||
| C16:0 | 4.20 | 9.31 | 14.59 | 1.03 | 1.44 | 5.42 | 1.11 | 0.91 |
| C16:1ω5c | 19.09 | 16.92 | 14.57 | 7.33 | 8.60 | 11.67 | 0.28 | 1.12 |
| C16:1ω9c | 0.11 | |||||||
| C16:1ω11c | 2.51 | 0.56 | 0.30 | 1.03 | ||||
| C17:0 | 0.07 | |||||||
| C18:0 | 1.40 | 0.56 | 0.73 | 0.34 | 0.74 | 0.42 | 1.13 | 0.79 |
| C18:1ω9c | 0.62 | |||||||
| PUFAs | ||||||||
| C16:2 | 2.55 | 1.86 | 1.21 | 3.45 | 2.00 | 3.15 | ||
| C18:2ω6,9, allcis | 2.70 | |||||||
| C18:3ω6,9,12, allcis | 1.25 | |||||||
| Hydroxy FAs | ||||||||
| C13:0 3-OH | ||||||||
| C14:0 3-OH | 0.34 | 0.11 | 0.27 | 0.13 | 0.38 | |||
| C15:0 3-OH | 0.07 | |||||||
| C16:0 2-OH | 0.16 | 0.17 | 0.13 | |||||
| C16:0 3-OH | 0.08 | 0.06 | 0.17 | 0.07 | 0.18 | |||
| Total SCFAs | 42.80 | 40.92 | 40.72 | 24.45 | 19.29 | 27.96 | 2.62 | 3.09 |
| BCFAs | ||||||||
| iso-C13:0 | 0.93 | 1.24 | 0.58 | 0.79 | 0.68 | 2.76 | 3.74 | |
| iso-C14:0 | 0.70 | 0.68 | ||||||
| iso-C15:0 | 44.43 | 23.12 | 32.01 | 55.25 | 63.48 | 42.59 | 33.90 | 36.45 |
| iso-C15:1ω9c | 0.95 | 0.36 | 0.89 | 0.38 | ||||
| iso-C16:0 | 1.32 | 0.25 | 0.42 | 0.18 | 0.19 | 0.40 | 2.99 | 1.76 |
| iso-C17:0 | 2.66 | 11.70 | 13.48 | 4.22 | 4.15 | 9.57 | 9.35 | 10.69 |
| iso-C17:1ω5c | 1.95 | 2.95 | 2.54 | 2.17 | 3.26 | 1.88 | 7.99 | 14.72 |
| iso-C17:1ω11c | 0.78 | 0.45 | 0.89 | 0.43 | 1.25 | 2.69 | 1.33 | |
| iso-C17:2ω5,11, all cis | 0.44 | 0.59 | 2.32 | 1.50 | 1.29 | 2.79 | 2.19 | |
| anteiso-C15:0 | 0.15 | |||||||
| Branched-chain hydroxy FAs | ||||||||
| iso-C15:0 3-OH | 0.55 | 2.48 | 0.81 | 2.15 | 1.77 | 2.30 | 2.40 | 1.32 |
| iso-C17:0 2-OH | 4.07 | 3.55 | 1.26 | 0.47 | 2.90 | 27.83 | 23.50 | |
| iso-C17:0 3-OH | 0.47 | 0.13 | 0.39 | |||||
| Branched-chain OAG FAs | ||||||||
| iso-C15:0 | 6.32 | 1.79 | 3.33 | 1.95 | 4.30 | 1.72 | ||
| Branched-chain DMA FA | ||||||||
| iso-C15:0 | 6.29 | 6.04 | 2.41 | 1.79 | 2.24 | 4.49 | 1.37 | |
| Total BCFAs | 57.20 | 59.08 | 59.28 | 75.55 | 80.71 | 72.04 | 97.38 | 96.91 |
Percentages of major fatty acids are distinguished in boldface type.
Fig. 1.

Chemo-phylogenetic tree of myxobacteria constructed by the neighbor-joining method based on 16S rRNA gene sequences and correlated with the fatty acid profile. The biomarker (major) FAs common for the genus are indicated after each set of bacterial initials on the right side of the tree (Ae, Aetherobacter; Ar, Archangium; By, Byssovorax; Cb, Cystobacter; Cc, Corallococcus; Cm, Chondromyces; En, Enhygromyxa; Hy, Hyalangium; Ja, Jahnella; Ko, Kofleria; Me, Melittangium; Mx, Myxococcus; Na, Nannocystis; Ph, Phaselicystis; Pl, Plesiocystis; Po, Polyangium; Px, Pyxidicoccus; Sg, Stigmatella; So, Sorangium) or have been specified for some species that do not agree (Cba, Cystobacter armeniaca; Cbm, Cystobacter miniatus; Ho, Haliangium ochraceum; Ht, H. tepidum; Mel, Melittangium lichenicola). The tree also highlights the dominant FAs in the three suborders. Fine dotted lines show the predominance of straight-chained fatty acids (SCFAs), while big dotted lines indicate the predominance of the branch-chained fatty acids (BCFAs). The vertical thick line shows the myxobacterial clusters devoid of hydroxy fatty acids, while the arrow line shows clusters with C17:1 2OH FA. The tree also localizes the production of omega-3 and omega-6 PUFA-producing strains. The PUFAs were abbreviated as follows: AA, arachidonic acid; ALA, α-linolenic acid; DHA, docosahexaenoic acid; EDA, eicosadienoic acid; EPA, eicosapentaenoic acid; GLA, γ-linolenic acid; LA, linoleic acid. Asterisks indicate the production of fatty acids in other isolates but only represented here by the type strain. iso fatty acids are indicated by “i.” The sequence of Desulfovibrio desulfuricans roots the tree. The numbers at branch points indicate the percentage of bootstrap support based on 1,000 resamplings. Only values greater than 60 are shown. Bar, 0.05 substitution per nucleotide position.
iso-C15:0 was found to be the major FA (23.1 to 63.5%) in Myxococcaceae, as also determined previously (6, 7, 34, 44, 51, 55), and was the FA in the largest amount observed in Myxococcus xanthus. This finding was in agreement with those for DK1622 (2) and several other strains (29, 31). The lower percentages in M. stipitatus-M. fulvus (23 to 32%) and Pyxidicoccus (44%) were reflected in the phylogenetic clustering. Corallococcus also had low iso-C15:0 (34 to 36%), and it is divergent from Myxococcus.
Straight-chain C16:1ω5c ranks as the second-most-abundant FA (7 to 19%) in Myxococcaceae, except Corallococcus, which contains a maximum of 1% (Table 1). Its low content affirms the previous reports (31, 48). On the other hand, the significant amounts (23.5 to 27.8%) of iso-C17:0 2-OH indicate that it could be a determinative marker for Corallococcus in Myxococcaceae family. Myxoccoccus had only 0.5 to 4% of this FA (Table 1), in contrast to the high value found in the M. fulvus Mx f2 non-type strain (7). This may be explained by strain-specific differences, as our findings on type strains are in perfect agreement with more recent studies (29, 31). Corallococcus also differed from Myxococcus in the absence of straight-chain hydroxy and O-alkylglycerol (OAG) FAs; however, both genera shared small amounts of iso-C17:1ω11c and diunsaturated iso-C17:2ω5,11, all cis.
C. exiguus differed from C. coralloides in the absence of iso-C15:0 OAG and dimethylacetal (DMA). The latter compound was shown to be derived from an aldehyde by a reduction process from iso-C15:0 and was found to increase during the first 24 h of development in Myxococcus xanthus DK1622 (40). OAG, on the other hand, was determined to be a monoacylglycerol derivative (MAG) compound (12). These iso-FAs, OAG and DMA, were shown to be important ether lipid-derived compounds contributing significantly to fruiting body formation in DK1622 (13, 41).
Archangium-Cystobacter cluster.
C16:1ω5c was the major FA (21 to 27%) in Archangium, in agreement with the reported amount found in A. gephyra strain 65 (55). It was also the major FA in Cystobacter, with the exception of C. armeniaca, C. miniatus, C. gracilis, and C. (Angiococcus) disciformis. These four isolates, as represented by type strains in this study (see Table S1 in the supplemental material), contain higher total BCFAs (70 to 75%), specifically iso-C15:0 (except C. gracilis) and iso-C17:0, lower C16:0, and show the presence of anteiso-C17:0 (Table 2). The latter FA has been reported in several marine isolates (8, 19), but it was also discovered here in Cystobacter spp. Cystobacter miniatus also differs from other Cystobacter species and even from other members of its suborder through its high (15.3%) C16:1ω7c content (Tables 1 to 3). The significant differences among FA profiles and polyphyletic position of these strains suggest their assignments to a novel genus, while C. disciformis should be reclassified back to its original genus, Angiococcus (22).
Table 2.
Fatty acid distribution among Cystobacterineae genera Archangium and Cystobacter
| FA type | % of FA ina: |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C. (Angiococcus) disciformis | A. gephyra | C. badius | C. ferrugineus | C. fuscus | C. gracilis | C. velatus | C. minus | C. violaceus | C. armeniaca | C. miniatus | |
| SCFAs | |||||||||||
| C14:0 | 1.99 | 1.09 | 4.60 | 7.03 | 5.95 | 0.26 | 10.26 | 5.68 | 2.91 | 0.66 | |
| C14:1ω5c | 0.29 | 1.58 | 0.70 | 1.53 | 0.60 | 0.65 | 0.07 | 0.19 | 0.11 | ||
| C15:0 | 0.13 | 0.30 | 0.21 | 0.50 | 0.23 | 0.52 | 0.33 | 0.36 | 0.26 | 3.73 | |
| C15:1 | 0.31 | 0.07 | 0.10 | 0.13 | 0.63 | ||||||
| C16:0 | 4.64 | 12.55 | 8.10 | 9.65 | 13.29 | 1.44 | 10.77 | 10.16 | 4.59 | 1.51 | 2.59 |
| C16:1ω5c | 15.81 | 24.55 | 22.70 | 25.42 | 22.34 | 13.07 | 20.62 | 26.86 | 27.48 | 6.12 | 1.31 |
| C16:1ω7c | 0.52 | 15.27 | |||||||||
| C16:1ω9c | 0.11 | 0.15 | |||||||||
| C16:1ω11c | 0.61 | ||||||||||
| C17:0 | 0.07 | 0.59 | |||||||||
| C18:0 | 0.75 | 7.78 | 0.67 | 0.47 | 1.24 | 0.51 | 0.28 | 0.30 | 0.50 | 0.64 | |
| C18:1 | 0.32 | 0.21 | |||||||||
| C18:1ω9c | 1.63 | ||||||||||
| PUFAs | |||||||||||
| C16:2 | 0.58 | 3.95 | |||||||||
| C18:2ω6,9, allcis | 0.27 | 0.55 | |||||||||
| C18:3ω6,9,12, allcis | 0.07 | 0.10 | 0.99 | ||||||||
| Hydroxy FAs | |||||||||||
| C14:0 3-OH | 0.23 | 0.66 | 0.17 | 0.77 | 0.16 | 0.47 | 0.37 | 0.25 | 0.22 | ||
| C16:0 2-OH | 0.30 | 0.56 | 1.10 | 1.35 | 1.11 | 1.66 | 0.83 | 1.38 | |||
| C16:0 3-OH | 0.19 | 0.16 | 0.11 | 0.40 | 0.07 | 0.22 | |||||
| OAG FAs | |||||||||||
| C16:0 | 0.13 | 0.51 | 0.30 | 0.28 | |||||||
| C16:1 | 6.41 | ||||||||||
| Total SCFAs | 25.34 | 45.96 | 39.35 | 45.34 | 47.21 | 25.68 | 46.15 | 44.75 | 38.14 | 10.44 | 30.61 |
| BCFAs | |||||||||||
| iso-C13:0 | 0.48 | 0.43 | 0.27 | 0.48 | 0.40 | 0.41 | 0.18 | 0.63 | |||
| iso-C14:0 | 0.16 | 0.19 | 0.43 | 0.12 | |||||||
| iso-C15:0 | 28.50 | 21.31 | 19.08 | 14.77 | 15.43 | 11.13 | 15.10 | 17.02 | 16.63 | 41.15 | 33.02 |
| iso-C16:0 | 2.07 | 2.13 | 4.85 | 2.04 | 3.92 | 11.87 | 4.13 | 1.89 | 1.70 | 1.54 | |
| iso-C17:0 | 11.30 | 4.99 | 5.55 | 4.27 | 2.81 | 10.44 | 3.19 | 6.51 | 3.84 | 14.84 | 13.65 |
| iso-C18:0 | 0.09 | ||||||||||
| iso-C17:1ω5c | 2.44 | 0.82 | 0.15 | 0.28 | 5.10 | 0.19 | 0.20 | 0.48 | 2.81 | ||
| iso-C17:1ω11c | 0.27 | ||||||||||
| iso-C17:2ω5,11, allcis | 0.23 | ||||||||||
| anteiso-C15:0 | 0.25 | 0.20 | |||||||||
| anteiso-C17:0 | 0.19 | 0.05 | 0.24 | 0.94 | 0.79 | ||||||
| Branched-chain OH FAs | |||||||||||
| iso-C15:0 3-OH | 1.11 | 1.59 | 2.42 | 0.50 | 1.81 | 1.02 | 1.03 | 1.19 | 0.90 | 2.22 | 0.71 |
| iso-C17:0 2-OH | 17.14 | 7.38 | 12.07 | 18.80 | 13.17 | 22.99 | 16.00 | 13.80 | 16.43 | 3.60 | |
| iso-C17:0 3-OH | 0.19 | ||||||||||
| Branched-chain OAG FA | |||||||||||
| iso-C15:0 | 2.79 | 6.04 | 9.04 | 9.63 | 11.47 | 1.95 | 13.68 | 5.74 | 8.36 | 9.35 | 10.36 |
| Branched-chain DMA FA | |||||||||||
| iso-C15:0 | 7.76 | 10.61 | 6.21 | 4.17 | 3.42 | 8.66 | 8.90 | 13.35 | 12.74 | 10.23 | |
| Total BCFAs | 74.66 | 54.04 | 60.65 | 54.66 | 52.79 | 74.32 | 53.85 | 55.25 | 61.86 | 89.56 | 69.39 |
Percentages of major fatty acids are distinguished in boldface type.
Table 3.
Fatty acid distribution in Cystobacterineae genera Stigmatella, Melittangium, and Hyalangium
| FA type | % of FA ina: |
||||||
|---|---|---|---|---|---|---|---|
| S. aurantiaca | S. erecta | S. hybrida | M. boletus SBMe003 | M. alboraceum | M. lichenicola | H. minutum | |
| SCFAs | |||||||
| C14:0 | 2.89 | 3.29 | 1.33 | 0.56 | 0.49 | ||
| C14:1ω5c | 0.17 | 0.29 | 0.58 | ||||
| C15:0 | 0.24 | 0.17 | 0.64 | ||||
| C15:1 | 0.14 | ||||||
| C15:1ω5c | 10.31 | ||||||
| C16:0 | 16.27 | 25.17 | 33.32 | 3.62 | 5.99 | 3.82 | 8.33 |
| C16:1ω5c | 11.83 | 8.14 | 4.99 | 3.56 | 4.66 | 18.93 | |
| C16:1ω7c | 32.25 | ||||||
| C16:1ω9c | 0.19 | 4.15 | 0.17 | 9.71 | |||
| C16:1ω11c | 0.33 | ||||||
| C17:0 | 0.07 | 0.30 | |||||
| C18:0 | 0.94 | 3.94 | 0.62 | 1.67 | 0.54 | 3.14 | 0.77 |
| C18:1 | 0.47 | ||||||
| C18:1ω9c | 1.53 | 5.64 | 1.42 | 1.48 | |||
| PUFAs | |||||||
| C16:2 | 13.04 | 0.62 | |||||
| C18:2 | 11.58 | 9.22 | |||||
| C18:2ω6,9, allcis | 14.76 | ||||||
| C18:3ω6,9,12, allcis | 3.96 | ||||||
| C18:3 | 1.75 | ||||||
| Hydroxy FAs | |||||||
| C14:0 3-OH | 0.29 | 0.28 | 0.13 | 0.24 | |||
| C16:0 2-OH | 0.46 | 0.28 | 1.44 | 0.06 | 0.31 | ||
| C16:0 3-OH | 0.16 | 0.10 | 0.20 | ||||
| OAG FAs | |||||||
| C15:0 | 0.50 | ||||||
| C16:0 | 2.93 | 1.26 | 2.30 | 4.00 | |||
| Total SCFAs | 50.00 | 62.85 | 76.52 | 26.43 | 27.62 | 30.21 | 35.23 |
| BCFAs | |||||||
| iso-C13:0 | 0.61 | 1.63 | 0.31 | ||||
| iso-C14:0 | 0.21 | ||||||
| iso-C15:0 | 16.50 | 13.82 | 5.55 | 31.59 | 27.51 | 29.62 | 20.98 |
| iso-C16:0 | 0.76 | 0.99 | 0.13 | 14.67 | 4.78 | ||
| iso-C17:0 | 8.90 | 4.11 | 9.65 | 7.71 | 21.85 | 8.96 | 14.65 |
| iso-C17:1ω5c | 0.33 | 0.84 | |||||
| iso-C17:1ω11c | 1.38 | 0.67 | |||||
| anteiso-C17:0 | 2.76 | ||||||
| Branched-chain hydroxy FAs | |||||||
| iso-C15:0 3-OH | 2.23 | 0.97 | 1.31 | 3.84 | 1.60 | 1.49 | 1.96 |
| iso-C17:0 2-OH | 7.17 | 3.61 | 6.59 | 3.73 | 4.72 | 6.99 | |
| iso-C17:0 3-OH | 0.86 | 0.17 | |||||
| Branched-chain OAG FA | |||||||
| iso-C15:0 | 5.48 | 9.79 | 7.45 | 6.38 | 6.02 | ||
| Branched-chain DMA FA | |||||||
| iso-C15:0 | 8.36 | 3.86 | 0.37 | 17.01 | 7.39 | 12.29 | 8.04 |
| Total BCFAs | 50.00 | 37.15 | 23.48 | 73.57 | 72.38 | 69.79 | 64.77 |
Percentages of major fatty acids are distinguished in boldface type.
Archangium is distinct from Cystobacter in its higher content of C18:0 (7.8%) and absence of straight-chain OAG and hydroxy FAs. However, the 1.2% C18:0 and 29.5% C19:0 found in A. gephyra strain 65 (55) were not detected in the type strain (DSM2261T), thus suggesting a case of strain variations.
Stigmatella-Hyalangium cluster.
The monophyletic position of the Stigmatella clade is reflected in its FA pattern. Stigmatella aurantiaca had equal amounts (50%) of total SCFAs and BCFAs, while S. erecta and S. hybrida had much higher total SCFAs (62.9 to 76.5%). This pattern for the latter two species was supported by a high bootstrap value (99.6%) and tree topology (Fig. 1). The three type species shared large amounts of C16:0, iso-C15:0, and iso-C17:0 FAs. The abundance of the latter two FAs in S. aurantiaca Sg a1 was previously reported (7). C16:1ω5c and iso-C17:0, both prominent in Cystobacter, were also found in considerable amounts in Stigmatella, but C16:1ω7c (32%) was unique to S. hybrida (Table 3).
The bifurcation of Hyalangium minutum NOCB-2T to Stigmatella was reflected in the ratio of SCFAs to BCFAs. The total level of BCFAs was much higher (65%), as in Cystobacter. The major FAs and their corresponding levels (iso-C15:0, 21%; C16:1ω5c, 19%; and iso-C17:0, 15%) likewise had counterparts in Cystobacter. Similarities in FA patterns and morphological characteristics of both genera reaffirm their placement in Cystobacteraceae.
Melittangium cluster.
The genus Melittangium appears polyphyletic (Fig. 1). Melittangium boletus strains Me b7 and Me b8T branch closely with C. miniatus, whereas Melittangium lichenicola is more closely affiliated with the majority of Cystobacter species. The third species, Melittangium alboraceum, was never cultivated (38). On a morphological basis, strain Me b7 closely matched M. alboraceum (47) and was therefore used to represent the taxon in FA analysis. The 16S rRNA gene sequence of the type strain (Me b8T) of M. boletus (39) was used for phylogenetic tree construction. However, strain Me b8T could not be revived (Klaus Gerth, personal communication) and therefore had to be replaced with strain SBMe003, which fits the description of M. boletus on the basis of its fruiting body structure (see Fig. S1c in the supplemental material) (32, 33) and bright yellow swarm (39).
Melittangium has many similarities to Cystobacter in their FA patterns: 70 to 74% of its FAs were BCFAs, of which iso-C15:0 was the highest (27.5 to 31.6%). iso-C15:0 DMA and iso-C17:0 were also present in significant amounts. The low content of C16:1ω5c (<5%) in Melittangium differentiates it from Cystobacter.
The similarity between Me b7 and Me b8, as represented by SBMe003, was reflected in tree topology (Fig. 1). Both contained nearly the same ratio of SCFAs and BCFAs (Table 3) and elevated amounts of iso-C15:0, iso-C15:0 DMA, and iso-C17:0. Phylogenetic and FA analyses suggest that Me b7, identified as M. alboraceum, (47), appears to be an M. boletus strain. We also agree that the described M. alboraceum strain (32) was just an immature fruiting stage of Chondromyces, as previously suggested (39).
Melittangium lichenicola appears distantly related from Me b7 and Me b8T in the phylogenetic tree (Fig. 1) and is divergent by the presence of large amounts of C15:1ω5c (10.3%) and iso-C16:0 (14.67%) and the absence of C16:1ω5c and iso-C15:0 OAG. FA C15:1ω5c was not detected in Me b7 and SBMe003 and may be considered an important taxonomic marker. Its paraphyletic affiliation with Cystobacter in the tree (Fig. 1) is manifested not only in the ratio of SCFAs to BCFAs but also in their similar amounts of unsaturated SCFAs—21.5% in M. lichenicola and 21.5 to 26.6% in Cystobacter spp. (C. velatus, C. ferrugineus, C. badius, and C. fuscus).
Suborder Nannocystineae.
The suborder Nannocystineae is a unique mixture of isolates grouped into two clusters—(i) the marine organisms Enhygromyxa and Plesiocystis, which are allied to the terrestrial nonhalophilic bacterium Nannocystis; and (ii) the Haliangium-Kofleria cluster (Fig. 1). Their phylogenetic placement in the same suborder could mean that these myxobacteria originated from a common ancestor but had developed convergent adaptations to different ecological niches in the course of evolution. This is further supported by the FA pattern.
Haliangium-Kofleria cluster.
iso-C16:0 and iso-C16:1 were the major BCFAs, with the amounts differing between species. Large amounts (5 to 12.6%) of iso-C17:0 were also found (Table 4). The type strains of Haliangium tepidum and H. ochraceum differed significantly in the ratios of BCFAs to SCFAs, suggesting that they should be in separate taxa. In H. tepidum (SMP-10T = DSM14436T), we found 13.7% SCFAs and 86.3% BCFAs, compared to 38.8% SCFAs and 60.4% BCFAs, as previously reported (8). It clusters with sister taxon Kofleria flava (Fig. 1), with both having comparable amounts of C16:0 (2.9% and 2.8%, respectively), much lower than the 15.1% reported for H. tepidum (8). In contrast, H. ochraceum had higher (18.4%) C16:0 and lower (4.8%) levels of iso-C15:0 OAG; our data suggest that the C16:0 content is less than half of the reported 38.3% (8). Although our study qualitatively reaffirms the presence of these FAs in type strains and supports their position in the phylogenetic tree, their amounts do not agree exactly with previous literature—perhaps a reflection of differences in cultivation media. The detection of 21 different FAs compared with 14 as previously reported in H. ochraceum (8) suggests that our medium supports more complex FA formation. Representatives of both studied genera Haliangium and Kofleria had small amounts (<3%) of anteiso-C17:0. It was demonstrated earlier that anteiso-branched acids serve as chemo-taxonomic markers for marine myxobacteria (8). This raises the question of the ancestor of the terrestrial Kofleria (e.g., strain Pl vt1T), which might be similar to the high-salt-tolerant genus Haliangium.
Table 4.
Fatty acid distribution among Nannocystineae type strains
| FA type | % of FA ina: |
||||||
|---|---|---|---|---|---|---|---|
| H. ochraceum | H. tepidum | E. salina | P. pacifica | K. flava | N. exedens | N. pusilla | |
| SCFAs | |||||||
| C13:0 | 0.56 | ||||||
| C14:0 | 0.18 | 0.79 | 0.41 | 17.33 | 10.59 | ||
| C14:1ω5c | 0.68 | 0.64 | |||||
| C15:0 | 0.63 | 0.45 | 0.18 | 0.35 | 2.37 | ||
| C15:1 | 2.12 | ||||||
| C16:0 | 18.41 | 2.89 | 10.69 | 6.67 | 2.81 | 12.01 | 6.00 |
| C16:1ω5c | 3.57 | 0.96 | 0.65 | 0.37 | 22.01 | 27.30 | |
| C16:1ω7c | 8.84 | 42.24 | 30.13 | ||||
| C16:1ω9c | 3.27 | 22.41 | 0.82 | 6.68 | |||
| C16:1ω11c | 4.41 | ||||||
| C17:0 | 1.81 | 0.39 | 0.20 | 0.73 | |||
| C17:1ω7c | 0.52 | 0.53 | 0.32 | ||||
| C18:0 | 4.81 | 2.23 | 5.88 | 3.19 | 0.76 | 7.75 | 3.05 |
| C18:1 | 2.23 | ||||||
| C18:1ω9c | 0.72 | 2.24 | 29.09 | 23.59 | |||
| PUFAs | |||||||
| C20:4ω6,9,12,15, allcis | 0.91 | 2.55 | |||||
| C20:5ω3,6,9,12,15, allcis | 1.44 | ||||||
| OAG FAs | |||||||
| C14:0 | 0.31 | ||||||
| C15:0 | 0.72 | 0.33 | 0.49 | ||||
| C16:0 | 6.98 | 0.18 | 1.48 | 0.67 | |||
| C16:1 | 0.20 | 2.93 | 6.55 | ||||
| Total SCFAs | 49.62 | 13.73 | 95.87 | 90.93 | 13.86 | 70.84 | 51.35 |
| BCFAs | |||||||
| iso-C14:0 | 0.15 | ||||||
| iso-C15:0 | 2.38 | 17.86 | 0.57 | 2.76 | 1.72 | 8.69 | 8.53 |
| iso-C16:0 | 25.45 | 14.02 | 1.20 | 2.46 | 34.14 | ||
| iso-C16:1 | 8.94 | 18.69 | 27.85 | ||||
| iso-C17:0 | 5.00 | 8.76 | 0.66 | 0.46 | 12.63 | 14.74 | 24.89 |
| iso-C17:1ω5c | 0.76 | ||||||
| iso-C17:1ω11c | 3.53 | 12.67 | |||||
| iso-C18:0 | 0.11 | 0.45 | |||||
| anteiso-C17:0 | 2.01 | 3.14 | 1.81 | ||||
| Branched-chain OAG FA | |||||||
| iso-C15:0 | 4.84 | 23.05 | 1.69 | 3.39 | 4.61 | ||
| Branched-chain DMA FA | |||||||
| iso-C15:0 | 1.65 | 2.79 | 2.20 | 2.56 | |||
| Total BCFAs | 50.38 | 86.27 | 4.13 | 9.07 | 86.14 | 29.16 | 48.65 |
Percentages of major fatty acids are distinguished in boldface type.
Enhygromyxa-Plesiocystis cluster.
In agreement with a previous study (20), hydroxy FAs were not detected, suggesting placement of Enhygromyxa and Plesiocystis in Nannocystineae and further justifying their topology and bootstrap support (100%) in the phylogenetic tree (Fig. 1). The predominance of SCFAs in both genera (19, 20) was also confirmed, but much higher values were obtained for the type strains of Enhygromyxa (95.9%) and Plesiocystis (90.9%), in comparison to 44.4% and 43.4% for Plesiocystis SIR-1T and SHI-1, respectively (20).
The major FAs detected in Enhygromyxa were iso-C15:0, iso-C16:0, and iso-C17:0 (20, 43), but here, a predominance of straight-chain C16:1ω7c (42%), C18:1ω9c (29%), and C16:0 (11%) was found. Although this study reproduced these findings with regard to detection of these FAs, the absence of quantitative data in previous studies prevents a true comparison. This study presents for the first time the complete FA data of E. salina DSM15217T (= SHK-1T). Plesiocystis was also reported to contain significant iso-C15:0 (32.3 to 35.6%) and iso-C16:0 (13.5 to 14.6%) (20), but <3% of both FAs were detected (Table 4), which can perhaps also be explained by differences in the cultivation media. Both genera contained C16:1ω7c (30 to 42%), C18:1ω9c (24 to 29%), and C16:0 (7 to 11%). C16:1ω9c (22.4%) was found only in Plesiocystis, and straight-chain OAG was found only in Enhygromyxa. It is possible that the FA C18:1ω9c is the marker by which these two low-salt-tolerant genera can be distinguished from the high-salt-tolerant genus Haliangium.
Nannocystis cluster.
Nannocystis phylogenetically clusters with “halotolerant” Enhygromyxa and Plesiocystis in Nannocystaceae (Fig. 1), although both differ significantly in cell morphology, source environment, and FA profile. Its predominant FAs were C16:1ω5c (22 to 27%), iso-C17:0 (15 to 25%), C14:0 (11 to 17%), and iso-C15:0 (nearly 9%). Nannocystis and Plesiocystis are distinguished in the suborder by the absence of straight-chain OAG FAs.
Nannocystis exedens differed from its sister taxon Nanocystis pusilla by production of less than 19% BCFA and the presence of C16:1ω9c. Both species also differed significantly in iso-C17:0 and iso-C17:1ω11c contents (Table 4).
Suborder Sorangiineae.
Of the six genera in the suborder Sorangiineae, only Sorangium, as represented by S. cellulosum strains So ce14 (7) and AJ 13585 (8), had previously been analyzed for FA content. In this study, all 12 species known to date in Byssovorax, Chondromyces, Jahnella, Phaselicystis, Polyangium, and Sorangium were covered. The new representative isolate Aetherobacter SBSr008 was also included in the analysis and in the phylogenetic tree (Fig. 1). SCFAs dominated over BCFAs, and out of a total of 49 FAs, 36 were identified as SCFAs. Sorangiineae thus appears to be the most complex among the myxobacteria with respect to SCFAs.
Polyangiaceae-Phaselicystidaceae cluster.
Straight-chain C16:1ω7c appears most abundant in Chondromyces-Jahnella (14 to 29%) and Polyangium (34 to 55%) clades, second to C16:1ω5c in Byssovorax (21%), and was not detectable in Sorangium and Phaselicystis. FA C16:1ω5c, though extremely rare in nature (26), was also comparatively high in Archangium, Nannocystis, and many Cystobacter strains. Its absence in Sorangium was unexpected, as this genus shares many characteristics with Byssovorax, like the ability to degrade cellulose. However, in Sorangiineae, Sorangium has the highest C16:0 (palmitic acid) content (20 to 25%).
A previous study showed that Sorangium (S. cellulosum So ce14) could be differentiated from Cystobacterineae through the absence of hydroxy FAs (7); however, we identified trace amounts of C16:0 2-OH and iso-C17:0 2-OH and 4.3 to 6.8% C17:1 2-OH in the type and reference strains (Table 5). The lack of detection of hydroxy-type FAs in So ce14 appears to reflect the differences in sensitivity of the analytical methods employed 30 years ago and does not seem to be associated with a particular strain. All other Sorangium cellulosum isolates in our collection analyzed for FAs produced hydroxy FAs (data not shown). C17:1 2-OH FA was higher in Chondromyces (9 to 15%) and Phaselicystis (25%). A total of 37.7% hydroxy FAs were found in the latter genus, while several others in small amounts were detected in other members of the Sorangiineae (Table 5).
Table 5.
Fatty acid distribution among Sorangiineae type strains
| FA type | % of FA ina: |
|||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. apiculatus | C. robustus | C. lanuginosus | C. pediculatus | C. robustus | P. fumosum | P. sorediatum | P. spumosum | S. cellulosum | S. cellulosum (“nigrum”)b | B. cruenta | Aetherobacterc | J. thaxteri | P. flavad | |
| SCFAs | ||||||||||||||
| C13:0 | 0.11 | 0.06 | 0.08 | |||||||||||
| C14:0 | 0.83 | 0.89 | 0.64 | 0.79 | 0.36 | 0.59 | 0.57 | 2.60 | 1.25 | 1.79 | 0.20 | 0.44 | 0.73 | |
| C14:1ω5c | 0.26 | 0.40 | 0.22 | 0.98 | 0.95 | 1.05 | 0.69 | |||||||
| C15:0 | 0.29 | 0.45 | 0.92 | 0.61 | 0.14 | 0.09 | 0.48 | 1.44 | 1.86 | 0.17 | ||||
| C15:1 | 0.31 | 0.41 | 0.06 | |||||||||||
| C15:1ω5c | 0.12 | |||||||||||||
| C16:0 | 9.46 | 16.42 | 4.51 | 11.72 | 3.39 | 5.65 | 5.96 | 4.17 | 19.60 | 24.85 | 4.98 | 2.14 | 8.29 | 6.01 |
| C16:0,9,10CH2 | 5.25 | 0.42 | ||||||||||||
| C16:1ω5c | 3.30 | 1.68 | 2.11 | 1.38 | 1.71 | 1.27 | 0.46 | 0.71 | 7.79 | 6.82 | 21.09 | 2.82 | 0.87 | |
| C16:1ω7c | 26.54 | 17.52 | 14.12 | 29.43 | 25.84 | 55.25 | 33.68 | 37.27 | 17.50 | 28.18 | ||||
| C16:1ω9c | 0.12 | 0.59 | ||||||||||||
| C16:1ω11c | 0.31 | 0.26 | ||||||||||||
| C17:0 | 1.99 | 4.47 | 3.98 | 2.18 | 0.66 | 1.19 | 0.26 | 1.58 | 0.24 | 1.59 | 1.31 | 0.87 | 5.53 | |
| C17:1ω7c | 0.31 | 0.50 | 0.51 | 0.45 | ||||||||||
| C18:0 | 2.94 | 2.53 | 2.11 | 4.84 | 4.82 | 1.75 | 2.67 | 3.18 | 4.61 | 8.18 | 1.97 | 1.43 | 7.05 | 3.22 |
| C18:1 | 25.80 | |||||||||||||
| C18:1ω9c | 3.02 | 0.33 | 7.47 | 1.85 | 20.23 | 5.40 | 1.29 | 25.53 | 2.59 | 1.45 | 13.90 | |||
| PUFAs | ||||||||||||||
| C16:2 | 0.74 | |||||||||||||
| C18:2 | 27.26 | |||||||||||||
| C18:2ω6,9c | 2.06 | 0.33 | 0.86 | 1.93 | 9.30 | 1.26 | 1.53 | 18.17 | 0.89 | |||||
| C18:3 | 0.03 | |||||||||||||
| C18:3ω6,9,12, allcis | 1.08 | |||||||||||||
| C20:2ω6,9, allcis | 1.09 | |||||||||||||
| C20:4ω6,9,12,15, allcis | 1.47 | 12.44 | ||||||||||||
| C20:5ω3,6,9,12,15, allcis | 10.90 | |||||||||||||
| C22:6ω3,6,9,12,15,18, allcis | 9.49 | |||||||||||||
| Hydroxy FAs | ||||||||||||||
| C16:0 2-OH | 1.45 | 1.44 | 0.17 | 0.84 | 0.50 | 0.56 | 0.08 | |||||||
| C16:1 2-OH | 1.69 | |||||||||||||
| C17:0 2-OH | 6.57 | |||||||||||||
| C17:1 2-OH | 8.87 | 14.88 | 11.27 | 10.98 | 11.38 | 0.62 | 2.86 | 5.94 | 6.77 | 4.31 | 5.24 | 13.50 | 2.09 | 25.19 |
| C18:0 2-OH | 0.32 | |||||||||||||
| C18:1 2-OH | 1.81 | |||||||||||||
| OAG FAs | ||||||||||||||
| C14:0 | 0.47 | 0.79 | ||||||||||||
| C15:0 | 1.27 | 1.45 | 1.58 | 0.61 | 0.66 | 0.37 | 0.77 | 10.21 | 0.16 | 1.43 | ||||
| C16:0 | 9.13 | 17.23 | 1.12 | 5.59 | 4.89 | 1.44 | 7.44 | 2.84 | 5.67 | 0.87 | 1.85 | 3.73 | 2.92 | |
| C16:1 | 11.86 | 0.38 | 5.33 | 1.42 | 0.61 | 4.86 | 0.52 | |||||||
| Total SCFAs | 83.26 | 81.05 | 57.22 | 77.81 | 83.77 | 71.38 | 60.97 | 80.72 | 78.17 | 72.28 | 73.25 | 63.13 | 80.86 | 68.73 |
| BCFAs | ||||||||||||||
| iso-C13:0 | 0.29 | 0.94 | 0.45 | 0.31 | 0.06 | |||||||||
| iso-C14:0 | 0.83 | 0.20 | ||||||||||||
| iso-C15:0 | 7.70 | 8.47 | 17.30 | 11.32 | 10.94 | 14.41 | 23.41 | 9.96 | 9.78 | 7.97 | 11.11 | 23.17 | 3.52 | 25.52 |
| iso-C16:0 | 0.76 | 0.18 | 8.98 | 1.78 | 0.10 | 1.44 | 0.82 | 0.58 | 0.41 | 2.25 | 0.52 | |||
| iso-C17:0 | 4.90 | 7.21 | 8.49 | 5.68 | 2.39 | 9.45 | 8.06 | 2.77 | 7.61 | 16.27 | 7.15 | 2.05 | 8.56 | 3.70 |
| iso-C17:1ω5c | 0.29 | 0.03 | ||||||||||||
| iso-C18:0 | 1.64 | 0.95 | 0.50 | 1.13 | ||||||||||
| anteiso-C17:0 | 0.50 | 0.21 | 0.20 | |||||||||||
| Branched-chain OH FAs | ||||||||||||||
| iso-C17:0 2-OH | 0.15 | 0.05 | 0.06 | 1.22 | 0.35 | 4.44 | 0.16 | |||||||
| iso-C17:1 2-OH | 1.89 | |||||||||||||
| iso-C17:0 3-OH | 0.18 | 0.23 | 0.11 | |||||||||||
| Branched-chain OAG FA | ||||||||||||||
| iso-C15:0 | 2.50 | 2.38 | 4.33 | 1.72 | 1.23 | 0.64 | 0.84 | 1.70 | 2.63 | 4.94 | 1.81 | 0.61 | ||
| Branched-chain DMA FA | ||||||||||||||
| iso-C15:0 | 0.55 | 0.48 | 0.43 | 0.63 | 1.57 | 3.32 | 5.85 | 5.65 | 9.40 | 0.15 | ||||
| Total BCFAs | 16.74 | 18.95 | 42.78 | 22.19 | 16.23 | 28.62 | 39.03 | 19.28 | 21.83 | 27.72 | 26.75 | 36.87 | 19.14 | 31.27 |
The characteristic anteiso FAs in “marine” myxobacteria (8, 19, 20) were also present in Chondromyces, Polyangium, and Jahnella, although trace amounts (<1%) of only anteiso-C17:0 were detected.
Although the proposed novel isolate Aetherobacter SBSr008 contained iso-C15:0 as the major FA, it lacked C18:1ω9c, and, unlike Chondromyces, Polyangium, Jahnella, and Byssovorax, it also lacked C16:1ω7c. In contrast to other members of Polyangiaceae, PUFAs constituted more than 20% of its total FAs and even higher levels in some strains within this cluster (49).
Gliding, nonfruiting bacteria: Herpetosiphon and Flexibacter.
Strains of Flexibacter (Flex 23 and Flex 7014) and Herpetosiphon (Hpg 287, Hpg 336, Hpg 383, and Hpg 454) have much simpler FA patterns (see Table S2 in the supplemental material) than myxobacteria. In Flexibacter, 14 FAs were identified, with roughly equal amounts of total SCFAs and BCFAs found in strains Flex 23 and Flex 7014. C16:1ω5c (34 to 40%) and iso-C15:0 (41 to 51%) were dominant in both strains. In a similar study, large amounts of C16:1ω5c were also detected in Flexibacter sp. strain Inp (21). PUFA production in the genus appears to be both species and strain specific. Neither EPA, known in one strain (17) and Flexibacter polymorphus (23), nor linoleic and linolenic acids, in strain Inp (21), were found in Flex 23 and Flex 7014. Arachidonic acid was reported recently in the gliding bacteria Aureispira marina and Aureispira maritima (15, 16).
The four strains of Herpetosiphon analyzed contained 4 to 10 straight-chain and even-numbered FAs, in agreement with a previous study (6). C18:1ω9c, C16:0, and C18:0 were dominant in all strains, except for C18:1ω9c in Hpg 383 (<3%). One PUFA, C18:2ω6,9, all cis, was also found in strains Hpg 287 and Hpg 336.
Hydroxy fatty acids.
The total hydroxy FAs were, on average, high in Cystobacterineae in comparison with Sorangiineae and absent in Nannocystineae. So far, only 2-OH and 3-OH hydroxy FAs were found in myxobacteria, agreeing with an earlier study (55). In Cystobacterineae, hydroxy BCFAs dominate over the hydroxy SCFAs. The straight-chain C17:1 2-OH was only found in Sorangiineae and appears to be an FA marker for the suborder. The presence of both the iso-branched and straight-chained C17:0 2-OH in Phaselicystis flava supports its current placement as a separate family (11).
iso-even and iso-odd BCFAs.
iso-odd BCFAs, though not necessarily found in larger total amounts, were more diversified than iso-even BCFAs. iso-C15:0 and iso-C17:0 serve as the major iso-odd FAs. The presence of iso-C15:0 has been shown to be crucial in fruiting body development in Myxococcus xanthus (41). Low levels of this FA (1.7 to 2.8%) in marine myxobacteria may account for their reported inability to form “true” fruiting bodies in aquatic environments (18, 19, 20). The formation of sporangioles containing ovoid spores in Haliangium tepidum SMP-10T (8), also reproducible in our study, may be associated with its high iso-C15:0 content (18%).
Of the 15 identified iso-FAs in the suborders, only 4 were iso-even (Table 4). Among those, the most common and abundant was iso-C16:0, while iso-C16:1 was determined to be exclusive to members of the Kofleria-“Haliangiaceae” cluster (Fig. 1) and therefore may be regarded as an important marker for the Kofleriaceae.
Saturated and unsaturated SCFAs.
Saturated SCFAs were, in general, more abundant than unsaturated SCFAs in Myxococcales. The larger amount of saturated SCFAs in Stigmatella (up to 37% in S. aurantiaca), marks its divergence from Cystobacter. In Nannocystineae, the Haliangium-Kofleria cluster had higher saturated than unsaturated SCFAs; this was the reverse in the Nannocystis-Enhygromyxa-Plesiocystis clade. The Nannocystis and Enhygromyxa-Plesiocystis clusters were differentiated by unsaturated SCFAs, of which the latter cluster contained as much as twice the amount (73 to 77%). The Sorangiineae generally contained larger amounts of unsaturated SCFAs (24 to 65%), except in Sorangium and the novel strain Aetherobacter SBSr008. Furthermore, Phaselicystidaceae differ from Polyangiaceae by the absence of unsaturated SCFA (Table 5), supporting their divergence as a family (Fig. 1).
Omega-6 PUFAs.
Using a FAME reference mixture (Fig. 2 A), PUFAs were identified in myxobacteria. Linoleic acid (LA; C18:2ω6,9, all cis) and γ-linolenic acid (GLA; C18:3ω6,9,12, all cis) (Fig. 2B) appear to be distributed among the Cystobacterineae and Sorangiineae suborders but were not found in Nannocystineae (Fig. 1).
PUFA C20:2ω6,9, all cis (eicosadienoic acid; EDA) (Fig. 2C) was also detected in myxobacteria, but only in Sorangium cellulosum So ce1851T (= DSM 14627T). We found later that almost half (45%) of the Sorangium strains in our collection were positive for EDA (R. Garcia et al., unpublished data); its absence in the reference isolate (So ce1654) suggests that EDA production is strain specific.
PUFA C20:4ω6,9,12,15, all cis (arachidonic acid; AA) was observed exclusively in Sorangiineae and Nannocystineae. Earlier, we reported its abundance in Phaselicystis flava (11) and have since detected it in small amounts in other strains belonging to the two suborders. The previously detected but unidentified C20:4 FA (19, 20) was confirmed and identified here as AA in the type strains of Plesiocystis pacifica and Enhygromyxa salina (Fig. 2D). However, the amount found in P. pacifica SIR-1T (DSM14875T) was low (2.6%) in comparison to the reported 14.1 to 17.5% (20). As pointed out in earlier sections, changes in the percentages of FA production may be attributed to differences in media and cultivation conditions.
Omega-3 PUFA.
Unlike omega-6 PUFAs, the omega-3 FAs were only discovered exclusively in Sorangiineae and Nannocystineae. PUFA C18:3ω3,6,9,12, all cis (α-linolenic acid; ALA) was present in some Chondromyces isolates (e.g., SBCm010) (Fig. 2E), but not in the five type strains studied. C20:5ω3,6,9,12,15, all cis (eicosapentaenoic acid; EPA), previously unknown in myxobacteria, was found initially in novel Sorangiineae isolates, such as SBSr008, SBSr002, and SBSr003 (49). Strain SBSr008 represents this cluster, with a significant level (10.9%) of EPA (Table 5 and Fig. 2F) compared with some Sorangium isolates (Garcia et al., unpublished). In Nannocystineae, only the Enhygromyxa salina type strain was found to produce EPA (Table 4). C22:6ω3,6,9,12,15,18, all cis (docosahexaenoic acid; DHA), also previously unknown in myxobacteria, was detected only in the novel Sorangiineae, such as SBSr008. We plan to propose the assignment of this unique strain, along with SBSr002 and SBSr003, which appear to be phylogenetically and morphologically closely related, to the novel genus, Aetherobacter. In a later screen with other novel isolates, we also discovered two additional PUFAs, identified as docosapentaenoic acid [C22:5(n-3)] and omega-6 homo-γ-linolenic acid [C20:3(n-6)] (data not shown).
Conclusions.
A comprehensive report of the cellular FA content in the order Myxococcales (myxobacteria) is presented here for the first time, covering most type strains and a representative novel isolate, to allow deduction of various FA correlations which might become useful for further follow-up work. Our study highlights the expanded FA profile of Sorangiineae and the discovery of PUFAs, particularly the omega-3 family.
Eight PUFAs, identified as linoleic acid, γ-linolenic acid, homo-γ-linolenic acid, eicosadienoic acid (all ω6), α-linolenic acid, eicosapentaenoic acid, docosapentaenoic acid, and docosahexaenoic acid (all ω3), were discovered for the first time in myxobacteria. Production of EPA appears restricted to certain genera in Sorangiineae and Nannocystineae. We described the extensive FA profile of Enhygromyxa salina and documented the production of EPA. Additionally, the discovery of DHA in the novel isolate Aetherobacter SBSr008 appears to be a unique characteristic exclusive to that genus. In our analysis, Herpetosiphon and Flexibacter (gliding, nonfruiting bacteria) show not only completely different FA profiles in comparison to myxobacteria, but also an absence of PUFAs, with the exception of linoleic acid in Herpetosiphon.
We have shown that myxobacteria could be potential sources of valuable omega-3 FAs for biotechnological and biopharmaceutical applications. Our overall study of their FA profiles shows complementarily with phylogeny findings and therefore might be regarded as a significant tool for the chemo-taxonomic classification of myxobactetria, especially for the discovery of novel PUFA producer strains.
Supplementary Material
ACKNOWLEDGMENTS
We sincerely thank Klaus Gerth (HZI) for the gliding bacteria and myxobacterial strains; Irineo J. Dogma, Jr., for critical comments and helpful discussions; and Janet Lei for proofreading of the manuscript.
This work was generously supported by German Bundesministerium für Bildung and Forschung (BMBF) grant 0315790 to InterMed Discovery GmbH and Scarland University.
Footnotes
Supplemental material for this article may be found at http://jb.asm.org/.
Published ahead of print on 11 February 2011.
REFERENCES
- 1. Bode H., Dickschat J., Kroppenstedt R., Schultz S., Müller R. 2005. Biosynthesis of iso-fatty acids in myxobacteria: iso-even fatty acids are derived by α-oxidation from iso-odd fatty acids. J. Am. Chem. Soc. 127:532–533 [DOI] [PubMed] [Google Scholar]
- 2. Bode H., et al. 2006. Straight-chain fatty acids are dispensable in the myxobacterium Myxococcus xanthus for vegetative growth and fruiting body formation. J. Bacteriol. 188:5632–5634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Bode H. B., et al. 2006. 3-Hydroxy-3-methylglutaryl-coenzyme A (CoA) synthase is involved in biosynthesis of isovaleryl-CoA in the myxobacterium Myxococcus xanthus during fruiting body formation. J. Bacteriol. 188:6524–6528 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Dickschat J., Bode H., Kroppenstedt R., Müller R., Schultz S. 2005. Biosynthesis of iso-fatty acids in myxobacteria. Org. Biomol. Chem. 3:2824–2831 [DOI] [PubMed] [Google Scholar]
- 5. Fang J., Kato C., Sato T., Chan O., McKay D. 2004. Biosynthesis and dietary uptake of polyunsaturated fatty acids by piezophilic bacteria. Comp. Biochem. Physiol. Part B. 137:455–461 [DOI] [PubMed] [Google Scholar]
- 6. Fautz E., Rosenfelder G., Grotjahn L. 1979. Iso-branched 2- and 3-hydroxy fatty acids as characteristic lipid constituents of some gliding bacteria. J. Bacteriol. 140:852–858 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Fautz E., Grotjahn L., Reichenbach H. 1981. Hydroxy fatty acids as valuable chemosystematic markers in gliding bacteria and flavobacteria, p. 127–133 In Reichenbach H., Weeks O. B. (ed.), The Flavobacterium-Cytophaga group. Verlag Chemie, Weinheim, Germany [Google Scholar]
- 8. Fudou R., Jojima Y., Iizuka T., Yamanaka S. 2002. Haliangium ochraceum gen. nov., sp. nov. and Haliangium tepidum sp. nov.: novel moderately halophilic myxobacteria isolated from coastal saline environments. J. Gen. Appl. Microbiol. 48:109–115 [DOI] [PubMed] [Google Scholar]
- 9. Funk C. D. 2001. Prostaglandins and leukotrienes: advances in eicosanoids biology. Science 294:1871–1875 [DOI] [PubMed] [Google Scholar]
- 10. Garcia R. O., Krug D., Müller R. 2009. Discovering natural products from myxobacteria with emphasis on rare producer strains in combination with improved analytical methods. Methods Enzymol. 458:59–91 [DOI] [PubMed] [Google Scholar]
- 11. Garcia R. O., Reichenbach H., Ring M. W., Müller R. 2009. Phaselicystis flava gen. nov., sp. nov., an arachidonic acid-containing soil myxobacterium, and the description of Phaselicystidaceae fam. nov. Int. J. Syst. Evol. Microbiol. 59:1524–1530 [DOI] [PubMed] [Google Scholar]
- 12. Garcia R. O., Gerth K., Stadler M., Dogma I. J., Jr., Müller R. 2010. Expanded phylogeny of myxobacteria and evidence for cultivation of the ‘unculturables’. Mol. Phylogenet. Evol. 57:878–887 [DOI] [PubMed] [Google Scholar]
- 13. Hoiczyk E., et al. 2009. Lipid body formation plays a central role in cell fate determination during developmental differentiation of Myxococcus xanthus. Mol. Microbiol. 74:497–517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Horrocks L. A., Yeo Y. K. 1999. Health benefits of docosahexaenoic acid (DHA). Pharmcol. Res. 40:211–225 [DOI] [PubMed] [Google Scholar]
- 15. Hosoya S., Arunpairojana V., Suwannachart C., Kantjana-Opas A., Yokota A. 2006. Aureispira marina gen. nov., sp. nov., a gliding, arachidonic acid-containing bacterium isolated from the southern coastline of Thailand. Int. J. Syst. Evol. Microbiol. 56:2931–2935 [DOI] [PubMed] [Google Scholar]
- 16. Hosoya S., Arunpairojana V., Suwannachart C., Kantjana-Opas A., Yokota A. 2007. Aureispira maritima sp. nov., isolated from marine barnacle debris. Int. J. Syst. Evol. Microbiol. 57:1948–1951 [DOI] [PubMed] [Google Scholar]
- 17. Hugenholtz P., Goebel B. M., Pace N. R. 1998. Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity. J. Bacteriol. 180:4765–4774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Iizuka T., Jojima Y., Fudou R., Yamanaka S. 1998. Isolation of myxobacteria from the marine environment. FEMS Microbiol. Lett. 169:317–322 [DOI] [PubMed] [Google Scholar]
- 19. Iizuka T., et al. 2003. Plesiocystis pacifica gen. nov., sp. nov., a marine myxobacterium that contains dihydrogenated menaquinone, isolated from the Pacific coasts of Japan. Int. J. Syst. Evol. Microbiol. 53:189–195 [DOI] [PubMed] [Google Scholar]
- 20. Iizuka T., et al. 2003. Enhygromyxa salina gen. nov., sp. nov., a slightly halophilic myxobacterium isolated from the coastal areas of Japan. Syst. Appl. Microbiol. 26:189–196 [DOI] [PubMed] [Google Scholar]
- 21. Intriago P., Floodgate G. D. 1991. Fatty acid composition of the estuarine Flexibacter sp. strain Inp: effect of salinity, temperature and carbon source for growth. J. Gen. Microbiol. 137:1503–1509 [Google Scholar]
- 22. Jahn E. 1924. Beitrage zur Botanischen Protistologie. I. Die Polyangiden. Gebrüder Borntraeger, Leipzig, Germany [Google Scholar]
- 23. Johns R. B., Perry G. C. 1977. Lipids of the marine bacterium Flexibacter polymorphus. Arch. Microbiol. 114:267–271 [Google Scholar]
- 24. Jøstensen J. P., Landfald B. 1997. High prevalence of polyunsaturated fatty acid-producing bacteria in arctic invertebrates. FEMS Microbiol. Lett. 151:95–101 [Google Scholar]
- 25. Kaneda T. 1991. Iso- and anteiso-fatty acids in bacteria: biosynthesis, function, and taxonomic significance. Microbiol. Rev. 55:288–302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Kearns D., et al. 2001. Identification of a developmental chemoattractant in Myxococcus xanthus through metabolic engineering. Proc. Natl. Acad. Sci. U. S. A. 98:13990–13994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Kopp M., et al. 2004. Critical variations of conjugational DNA transfer into secondary metabolite multiproducing Sorangium cellulosum strains So ce12 and So ce56: development of a mariner-based transposon mutagenesis system. J. Biotechnol. 107:29–40 [DOI] [PubMed] [Google Scholar]
- 28. Kunze B., et al. 2006. Cruentaren, a new antifungal salicylate-type macrolide from Byssovorax cruenta (Myxobacteria) with inhibitory effect on mitochondrial ATPase activity. J. Antibiot. 59:664–668 [DOI] [PubMed] [Google Scholar]
- 29. Lang E., Kroppenstedt R., Sträubler B., Stackebrandt E. 2008. Reclassification of Myxococcus flavescens Yamanaka et al. 1990VP as a later synonym of Myxococcus virescens Thaxter 1892AL. Int. J. Syst. Evol. Microbiol. 58:2607–2609 [DOI] [PubMed] [Google Scholar]
- 30. Lang E., Spröer C. 2008. Replacement of ATCC 25944T, the current type strain of Melittangium lichenicola, with ATCC 25946. Request for an opinion. Int. J. Syst. Evol. Microbiol. 58:2991–2992 [DOI] [PubMed] [Google Scholar]
- 31. Lang E., Stackebrandt E. 2009. Emended descriptions of the genera Myxococcus and Corallococcus, typification of the species Myxococcus stipitatus and Myxococcus macrosporus and a proposal that they be represented by neotype strains. Request for an opinion. Int. J. Syst. Evol. Microbiol. 59:2122–2128 [DOI] [PubMed] [Google Scholar]
- 32. McCurdy H. A. 1971. Studies on the taxonomy of the Myxobacterales. IV. Melittangium. Int. J. Syst. Bacteriol. 21:50–54 [Google Scholar]
- 33. McCurdy H. D. 1989. Family III. Cystobacteraceae, p. 2149–2158 In Staley J. T., Bryant M. P., Pfennig N., Holt J. G. (ed.), Bergey's manual of systematic bacteriology, vol. 3 Williams and Wilkins, Baltimore, MD [Google Scholar]
- 34. Monteoliva-Sanchez M., Ruiz C., Ramos-Cormenzana A. 1987. Cellular fatty acid composition of Corallococcus coralloides. Curr. Microbiol. 15:269–271 [Google Scholar]
- 35. Müller R., Gerth K. 2006. Development of simple media which allow investigations into the global regulation of chivosazol biosynthesis with Sorangium cellulosum So ce56. J. Biotechnol. 121:192–200 [DOI] [PubMed] [Google Scholar]
- 36. Nichols D., et al. 1999. Developments with Antarctic microorganisms: culture collections, bioactivity screening, taxonomy, PUFA production and cold-adapted enzymes. Curr. Opin. Biotechnol. 10:240–246 [DOI] [PubMed] [Google Scholar]
- 37. Nichols D., McMeekin T. 2002. Biomarker techniques to screen bacteria that produce polyunsaturated fatty acids. J. Microbiol. Methods 48:161–170 [DOI] [PubMed] [Google Scholar]
- 38. Peterson J. 1959. New species of myxobacteria from the bark of living trees. Mycologia 51:163–172 [Google Scholar]
- 39. Reichenbach H. 2005. Order VIII. Myxococcales, p. 1059–1144 In Brenner D. J., Krieg N. R., Staley J. T., Garrity G. M. (ed.), Bergey's manual of systematic bacteriology, vol. 2, part C. Springer, New York. NY [Google Scholar]
- 40. Reichenbach H., Lang E., Schumann P., Spröer C. 2006. Byssovorax cruenta gen. nov., sp nov., nom. rev., a cellulose-degrading myxobacterium: rediscovery of ‘Myxococcus cruentus’ Thaxter 1897. Int. J. Syst. Evol. Microbiol. 56:2357–2363 [DOI] [PubMed] [Google Scholar]
- 41. Ring M., et al. 2006. Novel iso-branched ether lipids as specific markers of developmental sporulation in the myxobacterium Myxococcus xanthus. J. Biol. Chem. 281:36691–36700 [DOI] [PubMed] [Google Scholar]
- 42. Ring M., Schwär G., Bode H. 2009. Biosynthesis of 2-hydroxy and iso-even fatty acids is connected to sphingolipid formation in myxobacteria. Chembiochem 10:2003–2010 [DOI] [PubMed] [Google Scholar]
- 43. Schäberle T. F., et al. 2010. Marine myxobacteria as a source of antibiotics—comparison of physiology, polyketide-type genes and antibiotic production of three new isolates of Enhygromyxa salina. Mar. Drugs 8:2466–2479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Schröder J., Reichenbach H. 1970. The fatty acid composition of vegetative cells and myxospores of Stigmatella aurantiaca (Myxobacterales). Arch. Mikrobiol. 71:384–390 [Google Scholar]
- 45. Shimkets L., Dworkin M., Reichenbach H. 2006. The myxobacteria, p. 31–115 In Dworkin M., Falkow S., Rosenberg E., Schleifer K.-H., Stackebrandt E. (ed.), The prokaryotes, 3rd ed., vol. 7. Springer, Berlin, Germany [Google Scholar]
- 46. Singh A., Wilson S., Ward O. P. 1996. Docosahexaenoic acid (DHA) production by Thraustochytrium sp., ATCC 20892. World J. Microbiol. Biotechnol. 12:76–81 [DOI] [PubMed] [Google Scholar]
- 47. Spröer C., Reichenbach H., Stackebrandt E. 1999. The correlation between morphological and phylogenetic classification of myxobacteria. Int. J. Syst. Bacteriol. 49:1255–1262 [DOI] [PubMed] [Google Scholar]
- 48. Stackebrandt E., et al. 2007. Taxonomic characterization of members of the genus Corallococcus: molecular divergence versus phenotypic coherency. Syst. Appl. Microbiol. 30:109–118 [DOI] [PubMed] [Google Scholar]
- 49. Stadler M., et al. June 2010. Production of omega-3 fatty acids by myxobacteria. International patent WO 2010/063451 A2. [Google Scholar]
- 50. Ward O., Singh A. 2005. Omega-3/6 fatty acids: alternative sources of production. Process Biochem. 40:3627–3652 [Google Scholar]
- 51. Ware J., Dworkin M. 1973. Fatty acids of Myxococcus xanthus. J. Bacteriol. 115:253–261 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Warude D., Joshi K., Harsulkar A. 2006. Polyunsaturated fatty acids: biotechnology. Crit. Rev. Biotechnol. 26:83–93 [DOI] [PubMed] [Google Scholar]
- 53. Weissman K. J., Müller R. 2010. Myxobacterial secondary metabolites: bioactivities and modes-of-action. Nat. Prod. Rep. 27:1276–1295 [DOI] [PubMed] [Google Scholar]
- 54. Wenzel S. C., Müller R. 2009. The biosynthetic potential of myxobacteria and their impact on drug discovery. Curr. Opin. Drug Discov. Devel. 12:220–230 [PubMed] [Google Scholar]
- 55. Yamanaka S., Fudo R., Kawaguchi A., Komagata K. 1988. Taxonomic significance of hydroxy fatty acids in myxobacteria with special reference to 2-hydroxy fatty acids in phospholipids. J. Gen. Appl. Microbiol. 34:57–66 [Google Scholar]
- 56. Yano Y., Nakayama A., Yoshida K. 1997. Distribution of polyunsaturated fatty acids in bacteria present in intestines of deep-sea fish and shallow-sea poikilothermic animals. Appl. Environ. Microbiol. 63:2572–2577 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.