

Abstract
Background
Of the animals typically used to study fertilization-induced calcium dynamics, none is as accessible to genetics and molecular biology as the model organism Caenorhabditis elegans. Motivated by the experimental possibilities inherent in using such a well-established model organism, we have characterized fertilization-induced calcium dynamics in C. elegans.
Results
Owing to the transparency of the nematode, we have been able to study the calcium signal in C. elegans fertilization in vivo by monitoring the fluorescence of calcium indicator dyes that we introduce into the cytosol of oocytes. In C. elegans, fertilization induces a single calcium transient that is initiated soon after oocyte entry into the spermatheca, the compartment that contains sperm. Therefore, it is likely that the calcium transient is initiated by contact with sperm. This calcium elevation spreads throughout the oocyte, and decays monotonically after which the cytosolic calcium concentration returns to that preceding fertilization. Only this single calcium transient is observed.
Conclusion
Development of a technique to study fertilization induced calcium transients opens several experimental possibilities, e.g., identification of the signaling events intervening sperm binding and calcium elevation, identifying the possible roles of the calcium elevation such as the completion of meiosis, the formation of the eggshell, and the establishing of the embryo's axis of symmetry.
Introduction
In all animals, fertilization generates a pattern of intracellular calcium dynamics within the oocyte that constitutes an essential trigger for normal development. The spatiotemporal properties of the calcium dynamics differ among animals, e.g., echinoderms, fish, and frogs have single calcium transients whereas ascidians, nemerteans, and mammals have multiple calcium oscillations [1]. Fertilization-induced calcium dynamics are mediated by release of internal calcium stores by inositol 1,4,5-triphosphate (IP3). In echinoderms and ascidians, the signaling pathway between sperm-egg fusion and the production of IP3 requires phospholipase Cγ and a Src family kinase, but little is known about the earliest events in this pathway [2,3,4].
In the C. elegans hermaphrodite, oocytes are formed by budding from a syncytium. Afterwards, they undergo maturation, ovulation, and are fertilized internally in a single-file, assembly-line-like process. An oocyte arrests at diakinesis of prophase I upon reaching the entrance to the spermatheca. The nuclear envelope breaks down ~ 6 min before the mature oocyte enters the spermatheca where its leading edge engulfs a single sperm. Eggshell formation is initiated and meiosis I and II and completed immediately follow fertilization. The newly fertilized egg remains in the spermatheca ~ 3-5 min before it is pushed out of the spermatheca and into the uterus [5,6]. The first cleavage in embryonic development occurs ~ 40 min after fertilization. Figure 1 is a DIC image of the posterior arm of the gonad showing the syncytial gonad, developing oocytes, spermatheca, and fertilized eggs within the uterus.
Figure 1.

A DIC image of the poserior arm of the gonad. Oocytes are formed by budding from the syncytial gonad, each bud carrying a single nucleus. Oocytes develop in single-file, and oocytes at various stages of development are visible. Oocytes are fertilized upon entrance to the spermatheca. After leaving the spermatheca, embryos develop in the uterus awaiting expulsion through the vulva. We inject dyes into the syncytial gonad. After recovery, the fluorescence pattern of the dyes becomes that of figure 2.
Here, we describe fertilization-induced calcium dynamics in the nematode C. elegans (see [7] for a description of an earlier study of the same). The benefit of continued use of C. elegans to study fertilization-induced calcium dynamics is the existence of powerful genetic tools, e.g., techniques for forward and reverse genetics, and an entirely sequenced genome. Molecular genetic analysis of the signaling pathway mediating fertilization-induced calcium dynamics may be possible using C. elegans.
Results and discussion
We tracked intracellular [Ca++] in oocytes by monitoring the fluorescence of the indicator dye Calcium Green-1 dextran, 10,000 MW (Molecular Probes). We microinjected the syncytial gonad with a solution of dye (100 μM dissolved in dH2O) using conventional techniques (see Materials and Methods; also [8]). As individual oocytes are formed, they incorporate dye in their cytoplasm (see Fig 2). We estimate dye concentrations in oocytes to reach 10 to 50 μM with this protocol (see Materials and Methods). Since these dye-filled oocytes are successfully fertilized and give rise to viable larvae, these dye concentrations are non-toxic and probably do not significantly perturb [Ca++] transients.
Figure 2.
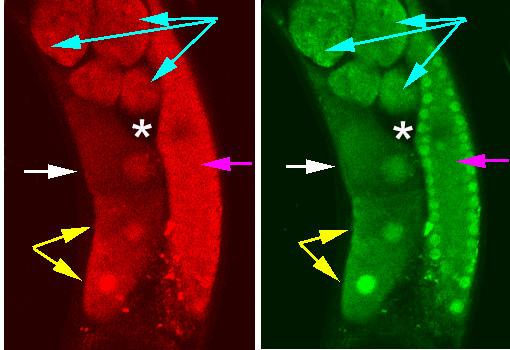
A portion of the posterior arm of the gonad. Fluorescence of tetramethylrhodamine, dextran 10,000 MW (red image) and Calcium Green-1, dextran 10,000 MW (green image) simultaneously measured in an injected animal. A single z-section constitutes this confocal image. The white arrow indicates the proximal oocyte, i.e., the oocyte nearest the entrance to the spermatheca. Blue arrows indicate developing embryos within the uterus. Pink arrows indicate the syncytial gonad in which pachytene nuclei line the walls of the syncytium. Yellow arrows denote maturing oocytes. The asterisk (*) denotes the spermatheca.
We coinjected Calcium Green dextran with rhodamine dextran, also 10,000 MW. As in other oocytes [9], both were about 50% brighter in the nucleus than in cytoplasm. This difference is due to the absence of organelles in the nucleus, compared to the significant excluded volume by the abundant yolk particles in the cytoplasm. Dye distributes uniformly throughout the oocyte following breakdown of the nuclear envelope (see Fig. 3).
Figure 3.

Fluorescence of Calcium Green-1, dextran 10,000 MW in a fertilized oocyte monitored by CCD camera. Images are presented at 3 sec intervals. The oocyte that is fertilized is denoted by the arrow. In this image, the uterus is above the fertilized oocyte, and contains a 2-celled embryo. An immature oocyte is below the fertilized oocyte, and has not yet undergone nuclear envelope breakdown. At t = 6 sec, the mature oocyte begins to enter the spermatheca through the sphincter at its entrance (at this point the oocyte is not yet encased in eggshell and retains flexibility). Between t = 12 sec and t = 15 sec, the leading edge of the oocyte has engulfed a sperm leading to a local ~ 30% increase in fluorescence, i.e., cytosolic [Ca++]. As the oocyte enters the spermatheca, the [Ca++] elevation spreads throughout the cell. At t = 30 sec, the entire oocyte has entered the spermatheca, and the oocyte has become rigid as a result of the formation of eggshell.
The unfertilized oocyte enters the spermatheca by squeezing through the constriction at its entrance. Fertilization induces a rise in cytosolic calcium within the oocyte that we detect from the increase in fluorescence of Calcium Green-1 (Fig. 3). The calcium elevation initiates at a point at the edge of the oocyte that first enters the spermatheca. Since sperm normally enter the oocyte at the end that first penetrates the spermatheca [5], we presume that the point of sperm entry is also the point of initiation of the calcium elevation. The point of sperm entry also sets the anteroposterior axis of the embryo [10], raising the possibility that calcium signaling is involved in axis determination. Caveat: Since it is not possible to visualize sperm fusion and image calcium simultaneously, it is not possible to formally rule out other causative agents for the calcium elevation; however, it seems highly unlikely given the ubiquity of the fertilization induced calcium elevation in other animals and the characteristics of the process in C. elegans.
Under the conditions described above, we typically measured peak increases in Calcium Green-1 fluorescence of ~ 30%. There is no corresponding change in the fluorescence of tetramethylrhodamine, confirming that the rise in Calcium Green-1 fluorescence is due to an increase in cytosolic [Ca++], and not due, for example, to motion artifacts. Although it is difficult to know the resting intracellular [Ca++] and the fluorescence properties of Calcium Green-1 in cytosol, assuming a resting concentration of 100 nM and the properties of Calcium Green-1 published in the Molecular Probes catalogue, the increase in Calcium Green-1 fluorescence indicates an elevation to ~ 250 nM. Assuming a resting concentration of 50 nM, the elevation would be to ~ 125 nM. The resting concentration is plausibly in the range of 50-100 nM (a range typical of most cell types). Also, although on the low side, the calcium elevation is of the same order of magnitude to other species, e.g., mollusks ~ 250 nM, annelids ~ 500 nm (see [1], for an exhaustive comparative analysis). In any event, the dynamics of the fertilization induced calcium elevation is clearly evident (see Fig 3 and 4, and Additional Material: Movie 1) although we provide only a rough quantitation of its amplitude.
Figure 4.

Ratiometric imaging of calcium elevation obtained by confocal microscopic measurement of fluorescence of coinjected tetramethylrhodamine, dextran 10,000 MW and Calcium Green-1, dextran 10,000 MW. Single z-sections constitute these images. These ratiometric images were obtained by calculating Δf/f at each pixel (fCaGreen-fRhodamine)/fRhodamine) which is the fractional increase in the fluorescence of the green channel (Calcium Green-1) over the red channel (tetramethylrhodamine); in all experiments, there was no measurable variation in the amplitude of the tetramethylrhodamine fluorescence (data not shown) making it useful for these ratiometric calculations). The inset is a plot of the average Δf/f within the area of the fertilized oocyte. The fertilized oocyte is denoted by the arrow. The empty spermatheca is denoted by the arrowhead (the redness of the spermatheca is an artifact of the ratio calculation; where there is no dye, green background fluorescence exceeds red background fluorescence, the area outside the worm was painted black for presentation purposes). At t = 3 min, the oocyte has completely entered the spermatheca and has been fertilized. The oocyte has entered the uterus at t = 9 min (the actual entry was missed between scan intervals). After ~ 12 min, cytosolic [Ca++] essentially reaches the level prior to fertilization. At t ~ 40 min, the first cell division takes place (data not shown), thus the calcium dynamic imaged here can be considered physiologically normal and not perturbative.
The elevation spreads rapidly throughout the oocyte as it completes entry into the spermatheca. The process of oocyte entry and calcium elevation spread takes 2-4 seconds. After fertilization, the calcium elevation drops monotonically to the level prior to fertilization. The time constant for the decay of the calcium elevation τ = 6 +/- 2 min in the three oocytes that we studied until the first cell division (measured by fitting an exponential exp(-t/τ) to the decay; one such experiment is in Fig. 4). Both the amplitude and duration of the single calcium transient seem comparable to those in other species [1].
In most animal eggs, the fertilization induced calcium elevation is due to the release of calcium from internal stores by an IP3. C. elegans has one gene encoding an IP3 receptor, itr-1. Worms homozygous for the null allele of the IP3 receptor, itr-1 (n2559) are sterile [11]. This sterility may be a direct result of the disruption of fertilization induced calcium signaling in the oocyte. It should now be possible to utilize the genetic tools available in C. elegans to study fertilization induced calcium signaling, and in particular to elucidate the signaling pathway intervening the fusion of sperm with oocyte and the rise in intracellular [Ca++].
Conclusion
This technique for measuring fertilization induced calcium transients provides a new experimental method in the study of C. elegans. A large number of existing mutants with fertilization defects can now be assayed for possible defects in the calcium transient [12]. Forward genetic (gene knockout and RNAi) methods available in C. elegans should permit testing proteins hypothesized to be involved in this critical step in embryogenesis.
Materials and Methods
Nematode strains & culture conditions
Methods for C. elegans culture and manipulation were as described in [13]. Worms were grown in the same room as the one used for imaging (20-23°C) since they seem to resist fertilizing and laying eggs at other than their growth temperature. The strain used was CB4845 unc-119(e2498) [14]. We used the unc-119(e2498) mutant since its near immobility facilitates extended observation.
Microinjection techniques were adapted from [8]. We inserted a needle (produced by pulling 1 mm OD glass capillary and filled with fluorescent dyes at a concentration of 100 μM in dH2O) into one of the 2 gonad arms of a young adult worm immobilized on an injection pad (pads are produced by drying a flattened drop of 2% agarose solution on a clean cover slip). Fluid is expelled into the gonad by moderately pressurizing the needle (we used an Eppendorf Transjector). Fluid can easily be observed entering and filling the gonad during needle pressurization. We aimed to 1/2 fill the gonad with fluid, and thus we presume the final dye concentration in the gonad (and the oocytes that are formed by budding from the gonad) to be ~ 10-50 μM. Worms were allowed to recover for ~ 2 hr after injection before imaging. This procedure did not kill the worm, or seem to damage the reproductive tissue. In every instance (dozens of individual experiments and many dozens of injected worms), viable dye-filled oocytes were produced. The dye-filled oocytes produced by this procedure eventually hatch into dye-filled larvae.
To image fertilization, the worms were then mounted in drops of M9 supplemented with 10 mg/mL serotonin on pads of 2% agarose. Serotonin promotes egg laying and seems to promote fertilization [15]. Sometimes, sloppy injections result in dye leaked into compartments outside the oocytes and syncytium. As previously noted with other injected dyes, dye that leaks outside the reproductive tissue accumulates in the coelomocytes [16]. We disposed of poorly injected worms. Finally, a coverslip was placed on the slide and sealed using nail polish.
Microscopy
Confocal microscopy (Figs. 2 &4) was performed using a Zeiss 510 confocal system. Video microscopy (Fig. 3) was performed using an Olympix cooled CCD camera mounted on an Olympus BX50WI. Average pixel intensities within the contours of individual fertilized oocytes were measured using Scion Image.
We observed calcium transients in 8/12 fertilized oocytes. We limited observation prior to fertilization to about 20 min. We found that it is critical to limit the animal's exposure to light and heat. Carelessness in this regard seemed to damage animals and oocytes, arresting fertilization and egg-laying. We routinely used an IR filter to block heat from the tungsten illumination. With video microscopy we used 1 % transmission neutral density filters to block most of the excitation light from the mercury arc source. Since the confocal microscope was used to assess the slow decline of the calcium transient, we typically scanned at a rate of 1 scan cycle per minute, and attenuated the laser to the lowest usable settings. The efforts taken to reduce light exposure also eliminated problems of photobleaching.
Supplementary Material
Quicktime movie corresponding to Figure 3 (see Figure 3 legend for details). Frames are continuously presented; each frame represents 0.3 s.
Acknowledgments
Acknowledgements
We began these experiments during the 1999 CSHL C. elegans course. We thank Karel Svoboda for providing access and support with imaging equipment. ADTS warmly thanks Mark Terasaki and Rindy Jaffe for teaching him about calcium imaging and fertilization; this paper would not exist without their coaching. Some strains were obtained from the C. elegans Genetic Stock Center, which is funded by a grant from the NIH National Center for Research Support. ADTS is an Amgen Fellow of the Life Sciences Research Foundation.
Contributor Information
Aravinthan DT Samuel, Email: asamuel@fas.harvard.edu.
Venkatesh N Murthy, Email: vnmurthy@fas.harvard.edu.
Michael O Hengartner, Email: hengartn@cshl.org.
References
- Stricker S. Comparative biology of calcium signaling during fertilization and egg activation in animals. Dev Biol. 1999;211:157–176. doi: 10.1006/dbio.1999.9340. [DOI] [PubMed] [Google Scholar]
- Carroll DJ, Albay DT, Terasaki M, Jaffe LA, Foltz KR. Identification of PLCγ-dependent and -independent events during fertilization of sea urchin eggs. Dev Biol. 1999;206:232–247. doi: 10.1006/dbio.1998.9145. [DOI] [PubMed] [Google Scholar]
- Carroll DJ, Ramarao CS, Mehlmann LM, Roche S, Terasaki M, Jaffe LA. Calcium release at fertilization in starfish eggs is mediated by phospholipase Cγ. J Cell Biol. 1997;138:1303–1311. doi: 10.1083/jcb.138.6.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giusti AF, Carroll DJ, Abassi YA, Terasaki M, Foltz KR, Jaffe LA. Requirement of a Src family kinase for initiating calcium release at fertilization in starfish eggs. J Biol Chem. 1999;274:29318–29322. doi: 10.1074/jbc.274.41.29318. [DOI] [PubMed] [Google Scholar]
- Kemphues KJ, Strome S. Fertilization and establishment of polarity in the embryo. in C elegans II (Edited by DL Riddle, T Blumenthal, BJ Meyer, and JR Priess) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 1997:335–360. [PubMed] [Google Scholar]
- McCarter J, Bartlett B, Dang T, Schedl T. On the control of oocyte meiotic maturation and ovulation in Caenorhabditis elegans. Dev Biol. 1999;205:111–128. doi: 10.1006/dbio.1998.9109. [DOI] [PubMed] [Google Scholar]
- Centonze VE, Polinko E, Strome S, White JG. Free calcium levels following fertilization in mutants of C. elegans. Abstract International worm meeting. 1997.
- Mello C, Fire A. DNA Transformation. In Caenorhabditis elegans: Modern Biological Analysis of an Organism (Edited by HF Epstein and DC Shakes), Academic Press, New York, NY. 1995:452–462. [Google Scholar]
- Terasaki M. Redistribution of cytoplasmic components during germinal vesicle breakdown in starfish oocytes. J Cell Science. 1994;107:1797–1805. doi: 10.1242/jcs.107.7.1797. [DOI] [PubMed] [Google Scholar]
- Goldstein B, Hird SN. Specification of the anteroposterior axis in Caenorhabditis elegans. Development. 1996;121:1467–1474. doi: 10.1242/dev.122.5.1467. [DOI] [PubMed] [Google Scholar]
- Dal Santo P, Logan MA, Chisholm AD, Jorgensen EM. The inositol triphosphate receptor regulates a 50-second behavioral rhythm in C. elegans. Cell. 1999;98:757–767. doi: 10.1016/s0092-8674(00)81510-x. [DOI] [PubMed] [Google Scholar]
- Singson A. Every sperm is sacred: fertilization in Caenorhabditis elegans. Dev Biol. 2001;230:101–119. doi: 10.1006/dbio.2000.0118. [DOI] [PubMed] [Google Scholar]
- Sulston J, Hodgkin J. Methods. In The Nematode Caenorhabditis elegans (Edited by WB Wood) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 1988:587–606. [Google Scholar]
- Maduro M, Pilgrim D. Identification and cloning of unc-119, a gene expressed in the Caenorhabditis elegans nervous system. Genetics. 1995;141:977–988. doi: 10.1093/genetics/141.3.977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horvitz HR, Chalfie M, Trent C, Sulston JE, Evans PD. Serotonin and octopamine in the nematode Caenorhabditis elegans. Science. 1982;216:1012–1014. doi: 10.1126/science.6805073. [DOI] [PubMed] [Google Scholar]
- White J. Anatomy. In The Nematode Caenhorabditis elegans (Edited by WB Wood) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 1988:109–110. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Quicktime movie corresponding to Figure 3 (see Figure 3 legend for details). Frames are continuously presented; each frame represents 0.3 s.