

Abstract
It is generally accepted that the combination of both Plasmodium falciparum parasite and human host factors is involved in the pathogenesis of complicated severe malaria, including cerebral malaria (CM). Among parasite products, the malarial pigment haemozoin (HZ) has been shown to impair the functions of mononuclear and endothelial cells. Different CM models were associated with enhanced levels of matrix metalloproteinases (MMPs), a family of proteolytic enzymes able to disrupt subendothelial basement membrane and tight junctions and shed, activate, or inactivate cytokines, chemokines, and other MMPs through cleavage from their precursors. Among MMPs, a good candidate for targeted therapy might be MMP-9, whose mRNA and protein expression enhancement as well as direct proenzyme activation by HZ have been recently investigated in a series of studies by our group and others. In the present paper the role of HZ and MMP-9 in complicated malaria, as well as their interactions, will be discussed.
1. Introduction
Among protozoan parasites of the genus Plasmodium, P. falciparum is the most deadly agent of human malaria, causing a broad spectrum of clinical manifestations ranging from asymptomatic to severe multiorgan disease. Despite recent major efforts by the research community, malaria remains one of the major diseases in poor areas, including Sub-Saharan Africa and South-East Asia. It is associated with several million clinical cases per year and leads annually to over one million deaths [1, 2]. The pathophysiology of severe malaria complications is not well understood. In some cases, including cerebral malaria (CM), renal failure, lung pathology and malaria during pregnancy, it appears associated with cytoadherence and sequestration of P. falciparum-parasitized red blood cells (pRBCs) to vascular endothelium, leading to microcirculatory obstruction, tissue hypoxia, and metabolic disturbances [3–5]. In some cases additional leukocyte extravasation has been reported [6, 7].
As described in the following sections, either pRBCs or parasite products such as haemozoin (HZ, malarial pigment), a lipid-enriched ferriprotoporphyrin IX crystal derived from haemoglobin catabolism by the parasite [8], can modulate the functions of mononuclear and endothelial cells and promote the production of proinflammatory molecules and other soluble factors, including matrix metalloproteinases (MMPs). MMPs are a well-known family of proteolytic enzymes able to disrupt subendothelial basement membranes [9, 10], to destroy tight junctions [11], and to shed, activate, or inactivate cytokines, chemokines, and other MMPs through cleavage from their precursors [12–14]. In the last decade, growing evidence on involvement of MMPs in falciparum malaria became available: human postmortem studies showed enhanced protein levels of MMP-1 in brains of CM patients [15], whereas MMP-8 was found increased in plasma of severe malaria patients [16]; additionally, activation of the human MMP-9 gene by P. falciparum has been reported in microarray studies on whole blood from children with malaria [17]. Interestingly, a role for MMPs during malaria is suggested also by evidence from nonhuman models of CM: in the brain of mice infected by P. Berghei ANKA, the etiological agent of murine CM, increased MMP-2, MMP-7, and MMP-9 levels, and pro-MMP-9 activation were found [18, 19].
The present paper will explore the effects of HZ on functions of mononuclear and endothelial cells, focusing on regulation of human MMP-9, which at present among the malaria-related MMPs is the most studied and could be a potential target for adjunctive therapy of complicated severe malaria.
2. Effects of HZ on Human Mononuclear and Endothelial Cells
HZ is a birefringent crystalline material made of Fe3+-Protoporphyrin IX dimers that derives from the degradation of haemoglobin by intraerythrocytic Plasmodium [8]. Unpurified HZ contains also unspecifically attached polyunsaturated fatty acids (PUFAs) such as arachidonic and linolenic acids, originating from membranes of Plasmodium digestive vacuoles [20]. The presence in unpurified HZ of large quantities of ferric haem with small amounts of free iron makes HZ a generator of oxidative radicals capable of forming lipoperoxides or other breakdown products from PUFAs (quasilipoxygenase activity) [21]. Analysis of the lipid fraction isolated from native HZ showed large amounts of hydroxyeicosatetraenoic acids (HETEs), hydroxyoctadecadienoic acids (HODEs), and the terminal aldehyde 4-hydroxynonenal (4-HNE) [22].
HZ is avidly phagocytosed by human monocytes, and the fatal risk for patients with severe falciparum malaria has been reported to be directly proportional to levels of HZ-containing monocytes in peripheral blood [25]. After phagocytosis of HZ, several monocyte functions are dramatically impaired, as shown in Figure 1. The effects of HZ on human monocytes are either inhibiting or stimulating. A detrimental case of functional inhibition led by HZ phagocytosis is represented by immunodepression. As evidenced from in vitro studies, HZ-fed human monocytes showed impairment of repeated phagocytosis [26], bacterial killing abilities [27], and oxidative burst [28]. Additionally HZ-laden monocytes did not respond to IFN-gamma stimulation and failed in MHC Class II expression, with following disturbances in antigen presentation [29]. These effects appear to be dependent on the lipid moiety of HZ, and low doses of 4-HNE have been reported to mimic HZ action by inhibiting the IFN-gamma-mediated MHC Class II expression [30]. On the contrary, the inhibitory role of HZ on monocyte differentiation/maturation to dendritic cells is less clear, since similar in vitro studies by different authors led to opposite results, showing either inhibition [31] or enhancement [32, 33] of differentiation/maturation to dendritic cells. HZ appears also to be related indirectly to anaemia, as HZ-fed monocytes are not able to coordinate the maturation of erythroid precursors; also in this case a role for 4-HNE in HZ-dependent inhibition of erythropoiesis has been proposed [34]. Regarding the stimulating effects of malarial pigment, HZ-laden monocytes have been shown to overproduce a large series of proinflammatory molecules, including cytokines (TNF, IL-1 beta, and IL-1RA), chemokines (IL-8/CXCL8, ENA-78/CXCL5, GROα, GROβ, GROγ, MIP-1α, MIP-1β, and MCP-1) and the cyto/chemokine-related proteolytic enzyme MMP-9 [23, 35–38]. The lipid moiety of HZ appears to be responsible for these effects, and a major role for 15-HETE has been proposed. Due to the important direct connections to MMP-9, the mechanisms underlying the HZ-triggered pro-inflammatory response will be deeply discussed in a following dedicated section. Curiously, although HZ compromises a large number of monocyte functions, leading to rapid inflammatory response, it has been recently shown that HZ-laden monocytes do not undergo apoptosis. To explain such an apparent dichotomy, a role for heat shock protein-27 (HSP-27), a chemokine-dependent molecule with antiapoptotic properties, has been recently proposed by our group. Indeed, in two models in vitro we showed HZ-dependent upregulation of HSP-27 either in immunopurified monocytes from peripheral blood or in the THP-1 monocytic cell line [38, 39].
Figure 1.
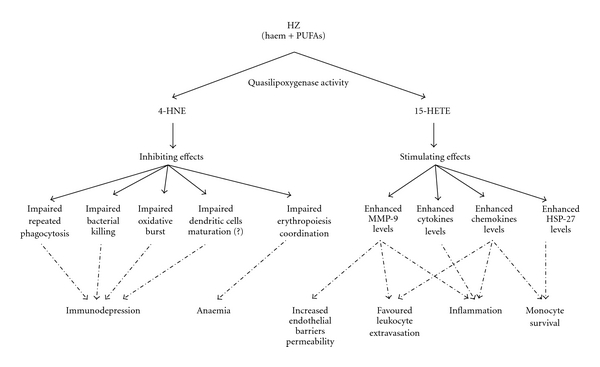
Effects of phagocytosis of HZ on human monocyte functions.
Anyway, phagocytic cells such as monocytes are not the only human cells which appear to be influenced by HZ. Few data are also available on direct alterations of the human endothelial cell phenotype by HZ. Such a change might be dramatically detrimental during CM, since endothelial cells are a crucial component of the blood-brain barrier (BBB). At present, HZ has been reported to enhance either basal or cytokine-induced levels of adhesion molecules [40, 41] and to inhibit the release of the vasoconstrictor mediator ET-1 [42–44]. Additionally, recent evidence from human microvascular endothelial cells (HMECs) as well as human umbilical vein endothelial cells (HUVECs) showed enhancement or de novo induction of protein expression and activity of several MMPs, including MMP-1, MMP-3, and MMP-9. This topic will be deepened in the following sections.
3. Involvement of MMP-9 in Complicated Severe Malaria
MMP-9 is produced by several cells, including mononuclear and endothelial cells.
Structurally, MMP-9 shares with the other members of the MMP family the following conserved domains: a prodomain, an active domain, a Zn2+-binding domain and a carboxyterminal haemopexin domain (which is absent in MMP-7 and MMP-26). In addition MMP-9, as well as the other gelatinase MMP-2, has a gelatin-binding fibronectin domain, composed of three fibronectin repeats, inserted between the active-site domain and the Zn2+-binding domain, plus an additional collagen type V domain in a particular hinge region. The Zn2+-binding domain, which contains three histidines responsible for the coordination of the catalytic Zn2+ ion, forms the active site with the active domain and is required for the enzymatic activity. The fourth ligand of the Zn2+ ion is a cysteine of the prodomain. This propeptide can be removed by proteolysis or distorted by substrate binding. The haemopexin domain plays a role in the substrate specificity and is important for the binding of the tissue inhibitors of metalloproteinases (TIMPs). The fibronectin repeats are related to gelatin, laminin, and collagens I/IV binding [9, 10, 45].
In the last decade, few evidence of direct involvement of MMP-9 in severe malaria has been reported. Activation of the human MMP-9 gene by P. falciparum has been shown in microarray studies on whole blood from children with severe malaria [17], whereas MMP-9 increases in cerebrospinal fluid levels were not apparent, possibly because of the enzyme's tight association with the extracellular matrix [46]. At present no studies of MMP-9 in human postmortem brain tissues have been reported, but MMP-9 levels have been shown to be strongly upregulated in C57BL/6 mice brain infected with P. berghei ANKA, a murine model of CM sharing similar characteristics with human CM. Elevated MMP-9 levels were selective for the central nervous system, where they were found associated with the vasculature and parenchyma whereas immunohistochemistry showed that higher amounts of MMP-9 were produced by cells of monocytic lineage (CD11b+) [18].
Activation of pro-MMP-9 by removal of the propeptide may occur through several proteolytic mechanisms. It is well known that it can be processed and further activated through a proteolytic cascade involving plasminogen, MMP-3 and MMP-1 [13, 47–49]. Interestingly, an accumulation of urokinase-type plasminogen activator receptor (uPAR) and MMP-1 has been described in brain of patients who died with CM [15, 50]. Furthermore, increased levels of MMP-3 in brain and spleen were observed in the murine model of CM previously cited [18]. Alternative mechanisms for pro-MMP-9 activation involve MMP-2 [51] and MMP-7 [52]. These molecules have not been studied yet in human malaria. However, in CM murine models, MMP-2 mRNA and protein were upregulated in brain [18] and MMP-7 mRNA was increased in liver but not in brain [18], whereas MMP-7 protein was increased in brain [19].
After secretion and activation, MMP-9 activity can still be regulated by degradation or inhibition. MMP-9 is inhibited not specifically by alpha2-macroglobulin and specifically by TIMPs, TIMP-1 showing the highest affinity for MMP-9 [53]. Inhibition of the activated MMP-9 occurs through interaction between the N-terminal domains of TIMP-1 and the active site of the enzyme [54]. The C-terminal part of MMP-9 is also involved, and seems to be responsible for high-affinity interaction with TIMP-1 [55]. Recently, Dietmann et al. measured serum levels of several TIMPs and MMPs in patients with severe malaria [16] and found that elevated TIMP-1 was associated with signs and symptoms of severe malaria; in the same study the authors detected increased levels of MMP-8, but not of MMP-9. MMP-9 is also inhibited by a large number of synthetic compounds [56–58]. Interestingly, antimalarial artemisinin- and artemisinin-derived drugs also showed anti-MMP-9 properties [59–62].
Functionally, MMP-9 is able to process a large pattern of molecules, including matrix proteins, inflammatory factors, surface molecules, and intercellular junctions [12, 14]. Among these substrates, several have been associated to severe malaria. Table 1 lists the principal MMP-9 substrates which might be relevant for malaria studies. The indicated molecules are either soluble factors (cytokines, chemokines) or membrane-associated proteins (pro-TNF, cell surface proteins, and intercellular junctions). All these molecules appear to be crucial for the development of complications of severe malaria, and their overbalanced regulation is often associated to CM. This is characterized by cytoadherence and sequestering of pRBCs to brain endothelial cells through ICAM-1, followed by an elevated inflammatory response, with high production of TNF and IL-1 beta, as well as several chemokines, which might recruit mononuclear cells and favour monocyte extravasation through the blood-brain barrier (BBB), which appears leaky due to the disruption of tight junctions ZO-1, claudin-5, and occludin [6, 7, 83–90]. Thus, in order to better understand and eventually prevent the mechanisms underlying CM, in the future it will be certainly intriguing to investigate the effects of MMP-9 on these molecules in complicated severe malaria models. Interestingly, the gene induction as well as the direct activation of MMP-9 by HZ has been demonstrated in vitro either in mononuclear or on endothelial cells, and an active role for HZ-dependent enhancement of soluble TNF production by human monocytes has already been demonstrated [23], as it will be discussed in the following section.
Table 1.
Substrates of MMP-9 that might be critical to complicated severe malaria.
| MMP-9 substrate | Functional classification | Effect of MMP-9 proteolytic activity | References |
|---|---|---|---|
| Pro-TNF | Cytokine proform | Shedding | [63] |
| Pro IL-1beta | Cytokine proform | Activation | [64] |
| IL-1beta | Cytokine | Degradation | [65] |
| Pro-TGF-beta | Cytokine proform | Activation | [66] |
| GRO-alpha/CXCL1 | Chemokine | Degradation | [67] |
| PF-4/CXCL4 | Chemokine | Degradation | [67] |
| ENA-78/CXCL5 | Chemokine | Transient potentiation, further inhibition | [68] |
| GCP-2/CXCL6 | Chemokine | No effects | [68] |
| NAP-2/CXCL7 | Chemokine | Degradation | [67] |
| IL-8/CXCL8 | Chemokine | Potentiation | [67] |
| MIG/CXCL9 | Chemokine | Inhibition | [69] |
| IP-10/CXCL10 | Chemokine | Degradation | [69] |
| SDF-1/CXCL12 | Chemokine | Inhibition | [70] |
| ICAM-1 | Cell surface protein | Inhibition | [71] |
| IL-2R-alpha | Cell surface protein | Inhibition | [72] |
| Occludin | Intercellular junction | Degradation | [73] |
| ZO-1 | Intercellular junction | Degradation | [74] |
| ZO-2 | Intercellular junction | Degradation (hypothesis) | [75] |
| Claudin-1 | Intercellular junction | Degradation | [76] |
| Claudin-2 | Intercellular junction | Degradation (hypothesis) | [77] |
| Claudin-4 | Intercellular junction | Activation (hypothesis) | [78] |
| Claudin-5 | Intercellular junction | Degradation (to be further investigated) | [79] |
| Syndecan-1 | Structural protein | Shedding | [80] |
| MBP | Structural protein | Degradation | [81] |
| Beta-dystroglycan | Structural protein | Degradation | [82] |
4. Interactions between HZ and MMP-9
The relationship between falciparum HZ and human MMP-9, as well as several related molecules, has been investigated in-depth in a series of recent works. In an elegant work Geurts et al. demonstrated that pro-MMP-9 binds directly to the beta-haematin core of HZ through its haemopexin domain, resulting in priming of the activation of the zymogen by other MMPs [91]. In human monocytes HZ has been reported to influence the MMP-9 gene and protein expression, as well as following protein release. Experiments by Real Time RT-PCR have shown that HZ and HZ-containing trophozoites enhanced the mRNA expression of MMP-9 [23]. HZ-dependent MMP-9 gene induction was also confirmed by additional microarray [92] and macroarray [38] studies. As a consequence, pro-MMP-9 protein expression was found to be increased [23]. The degranulation of gelatinase granules with MMP-9, lysozyme, and TIMP-1 was also promoted, as demonstrated by data showing HZ/trophozoite-dependent enhanced release of active MMP-9 [23], lysozyme [93, 94], and TIMP-1 [94] in monocyte cell supernatants. As a result, the total gelatinolytic activity of monocytes was increased by HZ [94] and cells were able to migrate through extracellular matrix [23].
The production of several soluble factors related to MMP-9 has also been studied. HZ and trophozoites were shown to stimulate the release of TNF [23] and IL-1 beta [37] by human monocytes. These cytokines, which can modulate the MMP-9 gene expression [95, 96], have been shown to play a crucial role in HZ-dependent enhancement, since the use of anti-TNF and anti-IL-1beta antibodies abrogated the previous HZ effects on pro-MMP-9 expression and active MMP-9 release [23, 37]. Anti-TNF antibodies also blocked the HZ-dependent release of active lysozyme [93]. Interestingly, pro-TNF and pro-IL-1 beta are also two molecular substrates of MMP-9 (see Table 1); as a consequence of cleavage by MMP-9, the soluble TNF is shed from its membrane-bound precursor [63], whereas IL-1 beta is directly activated after removal of the propeptide [64]. The use of a synthetic inhibitor of MMP-9 was shown to abrogate the HZ-dependent enhancement of TNF release by human monocytes [23]. Such an evidence demonstrated that during malaria a pathological autoenhancing loop between MMP-9 and TNF is likely, and the inhibition of MMP-9 could be particularly important in order to avoid detrimental inflammatory effects during complicated malaria, CM above all. On the other hand, the MMP-9 inhibitor did not alter the HZ-enhanced release of IL-1 beta [37], suggesting that in HZ-fed monocytes the production of IL-1 beta precedes the MMP-9 enhancement. This hypothesis was confirmed by recent time-dependent macroarray studies on expression of inflammatory genes, showing that praecox IL-1 beta mRNA expression was already present 2 h after exposure of human monocytes to HZ; therefore, IL-1beta was rapidly endorsed by additional transcription of other inflammatory genes, including MMP-9, TNF, IL-1RA, and several chemokines (IL-8/CXCL8, ENA-78/CXCL5, GRO-alpha/CXCL1, GRObeta, GRO-gamma, MIP-1 alpha, MIP-1 beta, and MCP-1) [38]. Moreover, crossed experiments with anti-TNF and anti-IL-1beta blocking antibodies suggested that the production of IL-1 beta was earlier than TNF production [37].
Recently, the effects of HZ on MMP-9 regulation have been also studied in endothelial cells. MMP-9 protein expression and release was shown to be dose-dependently induced de novo by HZ in HMECs and to enhance total gelatinolytic activity; as a consequence, cells displayed morphological changes, showing elongated shape and increased ability to form microtubule-like structures through extracellular matrix. In the same work, the enhanced expression of MMP-1 and MMP-3, two enzymes causally connected in the cascade of activation of pro-MMP-9, was also described [97]. Additional data for endothelial cells from large calibre vessels are described in Figure 2, showing that HZ and HZ-containing trophozoites were able to enhance basal levels of activated MMP-9 released in cell supernatants from HUVEC cultures.
Figure 2.
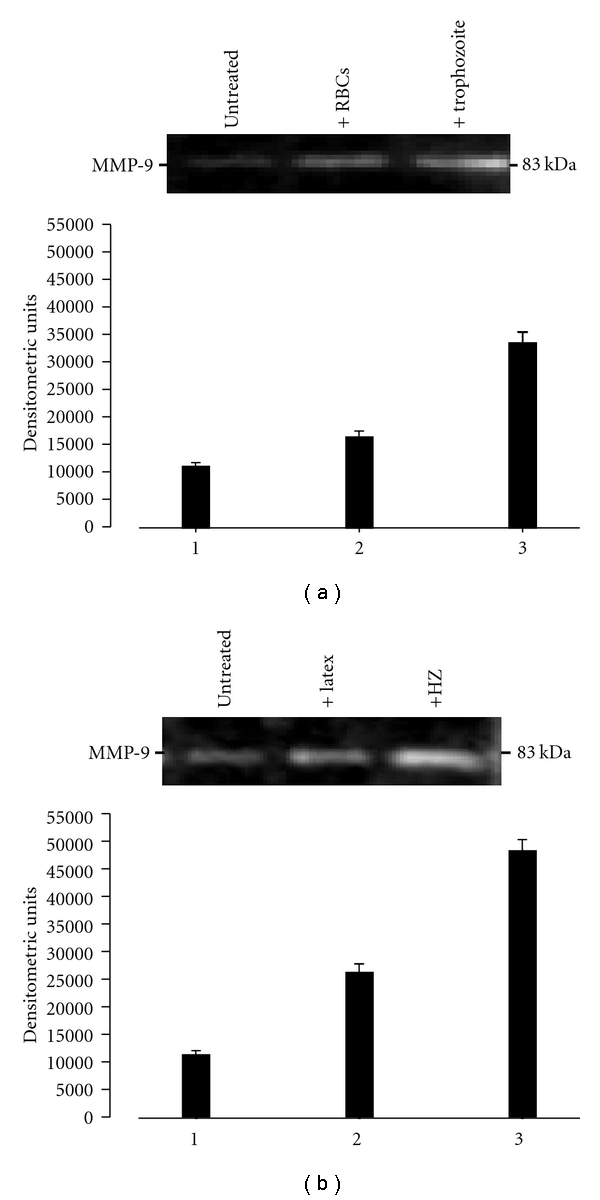
HZ-containing pRBCs (trophozoites) and free HZ enhance MMP-9 levels in HUVECs. Untreated cells (column/lane 1, both panels), cells treated with uninfected RBCs or latex as control stimuli (column/lane 2, (a) and (b) resp.), and cells treated with HZ-containing-trophozoites or HZ (column/lane 3, (a) and (b), resp.) were incubated for 48 h. At the end of the treatment, MMP-9 levels were measured in HUVEC supernatants by SDS-PAGE gelatin zymography and densitometric analysis through a computerized densitometer as previously described [23, 24]. Column data (lower panels) are mean values of arbitrary densitometric units + SEM of three independent experiments; gel data (upper panels) are from one representative experiment; the 83-kDa negative bands in the gel indicate MMP-9 levels. Data were analyzed for significance by Student's t-test and all differences were significant ((a) 1 versus 2: P < .02; 1 versus 3: P < .001; 2 versus 3: P < .002. (b) 1 versus 2: P < .001; 1 versus 3: P < .0001; 2 versus 3: P < .001).
Further studies performed either on mononuclear or endothelial cells were aimed to clarify what component (haem or lipid moiety) was responsible for the effects of HZ on MMP-9 and related soluble factors described above. In human monocytes the use of delipidized native or lipid-free synthetic HZ did not reproduce the enhancing effects of HZ on MMP-9 levels [37], gelatinolytic activity [94], lysozyme release [94], and production of TNF [98], IL-1 beta [37], and several chemokines [38], suggesting a major role for the lipid moiety of HZ. Further investigation suggested that a valid candidate responsible for HZ stimulatory effects could be 15-HETE, since micromolar doses of this compound mimicked HZ effects on all previous parameters [37, 38, 94, 98]. A role for the lipid moiety of HZ has been also proposed to explain the HZ effects on HMECs, since delipidized and synthetic HZ did not affect MMP-9, MMP-3, and MMP-1 protein expression [97].
Finally, a mechanism involving NF-kappaB activation has been recently proposed to explain how HZ and 15-HETE might promote the production of MMP-9 and related molecules in human monocytes. After phagocytosis of HZ and trophozoites, as well as stimulation with 15-HETE, the phosphorylation and degradation of cytosolic IkappaB-alpha protein and the following nuclear translocation of the NF-kappaB complex were promoted either in short-term (2 h after phagocytosis) or long-term (24 h after phagocytosis) studies. These events appeared directly related to HZ-dependent enhancement of MMP-9 release and production of TNF and IL-1 beta, since the use of NF-kappaB inhibitors showing antimalarial properties (quercetin, artemisinin, and parthenolide) abrogated the effects of both HZ and 15-HETE [61]. Additionally, the same inhibitors also abrogated the 15-HETE enhancing effects on total gelatinolytic and lysozyme activity [94]. In another study using a monocytic cell line (THP-1), an extract from a plant with antimalarial properties (Punica granatum) inhibited HZ-dependent enhancement of MMP-9 expression and secretion by blocking the NF-kappaB-driven transcription [99]. The previous literature data have shown that HETEs are peroxisome proliferator-activated receptor-gamma ligands inhibiting the production of inflammatory cytokines and suppressing the NF-kappaB system. However, alternative mechanisms which might lead to HETEs-driven NF-kappaB activation have also been documented. A first alternative activation of NF-kappaB is based on HETE-mediated activation of protein kinase C [100–102] which in turn activates IKK [103], the kinase responsible for IkappaB-alpha phosphorylation and degradation [104], and subsequent nuclear translocation of NF-kappaB. Transient activation of protein kinase C by HZ [105] may thus explain the first NF-kappaB activation peak revealed 2 h after HZ phagocytosis. A second alternative activation of NF-kappaB [106] was found to involve MAP kinases, ERK, and p38 cascade. The p38 cascade was also proposed for HETEs-dependent modulation of TNF [107] and MMP-9 [108]. Interestingly, ERK-1/2 phosphorylation by HZ has been documented in a murine system [109]. A third alternative activation of NF-kappaB may result from the direct oxidation and inactivation of IkappaB by lipid hydroperoxides [110]. Figure 3 summarizes the putative mechanisms through which HZ and HETEs might activate the NF-kappaB transcription system and promote the following enhancement of gene induction of MMP-9 and related cytokines.
Figure 3.
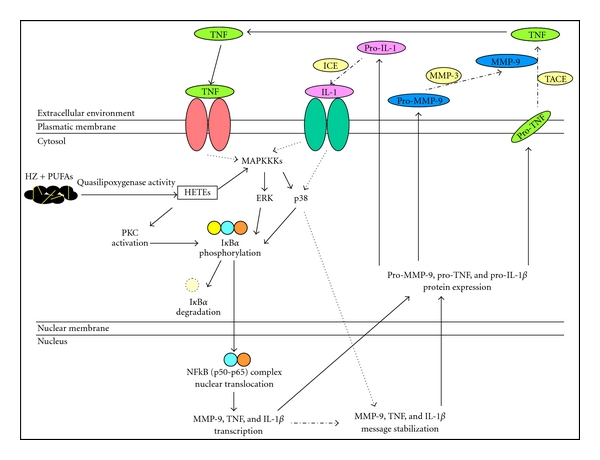
Putative mechanisms through which HZ and 15-HETE might promote NF-kappaB-controlled gene expression of MMP-9 and related molecules.
5. Conclusions
Evidence of the involvement of MMPs in complicated severe malaria has been emerging during the last decade, and HZ-dependent induction of MMP-9 expression and activity has been demonstrated in mononuclear or endothelial cells. Since several MMP inhibitors are already available as potential therapeutic tools in other pathologies [55–57], MMPs should be taken in account as potential new targets for an innovative and adjunctive therapy for severe malaria.
Acknowledgments
Thanks are due to Dr. Simona Cardaropoli for her help with HUVEC cultures and to Dr. Nicoletta Basilico, Professor Donatella Taramelli, Dr. Philippe Van den Steen and Professor Ghislain Opdenakker for their precious suggestions and stimulating discussions. This study was supported by University of Torino Intramural Funds to GG, by grants from Compagnia di San Paolo to MP in the context of the Italian Malaria Network and by kind supplemental donation from Mrs. Franca Squazza to MP. The authors have no conflicting financial interests.
Abbreviations
- HZ:
Haemozoin
- MMP:
Matrix metalloproteinase
- CM:
Cerebral malaria
- HMEC:
Human microvascular endothelial cell
- HUVEC:
Human umbilical vein endothelial cell
- BBB:
Blood-brain barrier
- MHC:
Major histocompatibility complex
- PUFA:
Polyunsaturated fatty acid
- HETE:
Hydroxyeicosatetraenoic acid
- HODE:
Hydroxyoctadecadienoic acid
- HNE:
Hydroxynonenal
- TIMP:
Tissue inhibitor of metalloproteinase
- TNF:
Tumor necrosis factor
- IL:
Interleukin
- ICAM:
Intercellular adhesion molecule
- uPAR:
Urokinase-type plasminogen activator receptor
- TGF:
Transforming growth factor
- ZO:
Zonula occludens
- MBP:
Mannose binding protein
- GRO:
Growth-regulated oncogene
- PF:
Platele factor
- ENA:
Epithelial neutrophil activating peptide
- MIP:
Macrophage inflammatory protein
- MCP:
Monocyte chemoattractant protein
- GCP:
Granulocyte chemotactic protein
- NAP:
Neutrophil attractant-activation protein
- MIG:
Monokine induced by interferon-gamma
- IP:
Interferon-gamma-inducible protein
- SDF:
Stromal derived factor
- ET:
Endothelin
- HSP:
Heat shock protein.
References
- 1.Murphy SC, Breman JG. GAPS in the childhood malaria burden in Africa: cerebral malaria, neurological sequelae, anemia, respiratory distress, hypoglycemia, and complications of pregnancy. American Journal of Tropical Medicine and Hygiene. 2001;64(1-2):57–67. doi: 10.4269/ajtmh.2001.64.57. [DOI] [PubMed] [Google Scholar]
- 2.Khadjavi A, Giribaldi G, Prato M. From control to eradication of malaria: the end of being stuck in second gear? Asian Pacific Journal of Tropical Medicine. 2010;3(5):412–420. [Google Scholar]
- 3.Brown H, Rogerson S, Taylor T, et al. Blood-brain barrier function in cerebral malaria in Malawian children. American Journal of Tropical Medicine and Hygiene. 2001;64(3-4):207–213. doi: 10.4269/ajtmh.2001.64.207. [DOI] [PubMed] [Google Scholar]
- 4.Andrews KT, Lanzer M. Maternal malaria: Plasmodium falciparum sequestration in the placenta. Parasitology Research. 2002;88(8):715–723. doi: 10.1007/s00436-002-0624-5. [DOI] [PubMed] [Google Scholar]
- 5.Beeson JG, Reeder JC, Rogerson SJ, Brown GV. Parasite adhesion and immune evasion in placental malaria. Trends in Parasitology. 2001;17(7):331–337. doi: 10.1016/s1471-4922(01)01917-1. [DOI] [PubMed] [Google Scholar]
- 6.Patnaik JK, Das BS, Mishra SK, Mohanty S, Satpathy SK, Mohanty D. Vascular clogging, mononuclear cell margination, and enhanced vascular permeability in the pathogenesis of human cerebral malaria. American Journal of Tropical Medicine and Hygiene. 1994;51(5):642–647. [PubMed] [Google Scholar]
- 7.Grau GE, Mackenzie CD, Carr RA, et al. Platelet accumulation in brain microvessels in fatal pediatric cerebral malaria. Journal of Infectious Diseases. 2003;187(3):461–466. doi: 10.1086/367960. [DOI] [PubMed] [Google Scholar]
- 8.Slater AFG, Swiggard WJ, Orton BR, et al. An iron-carboxylate bond links the heme units of malaria pigment. Proceedings of the National Academy of Sciences of the United States of America. 1991;88(2):325–329. doi: 10.1073/pnas.88.2.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nagaset H, Woessner JF. Matrix metalloproteinases. Journal of Biological Chemistry. 1999;274(31):21491–21494. doi: 10.1074/jbc.274.31.21491. [DOI] [PubMed] [Google Scholar]
- 10.Opdenakker G, Van den Steen PE, Dubois B, et al. Gelatinase B functions as regulator and effector in leukocyte biology. Journal of Leukocyte Biology. 2001;69(6):851–859. [PubMed] [Google Scholar]
- 11.Rosenberg GA, Yang Y. Vasogenic edema due to tight junction disruption by matrix metalloproteinases in cerebral ischemia. Neurosurgical focus. 2007;22(5):p. E4. doi: 10.3171/foc.2007.22.5.5. [DOI] [PubMed] [Google Scholar]
- 12.Cauwe B, Van den Steen PE, Opdenakker G. The biochemical, biological, and pathological kaleidoscope of cell surface substrates processed by matrix metalloproteinases. Critical Reviews in Biochemistry and Molecular Biology. 2007;42(3):113–185. doi: 10.1080/10409230701340019. [DOI] [PubMed] [Google Scholar]
- 13.Cuzner ML, Opdenakker G. Plasminogen activators and matrix metalloproteases, mediators of extracellular proteolysis in inflammatory demyelination of the central nervous system. Journal of Neuroimmunology. 1999;94(1-2):1–14. doi: 10.1016/s0165-5728(98)00241-0. [DOI] [PubMed] [Google Scholar]
- 14.Van Lint P, Libert C. Chemokine and cytokine processing by matrix metalloproteinases and its effect on leukocyte migration and inflammation. Journal of Leukocyte Biology. 2007;82(6):1375–1381. doi: 10.1189/jlb.0607338. [DOI] [PubMed] [Google Scholar]
- 15.Deininger MH, Winkler S, Kremsner PG, Meyermann R, Schluesener HJ. Angiogenic proteins in brains of patients who died with cerebral malaria. Journal of Neuroimmunology. 2003;142(1-2):101–111. doi: 10.1016/s0165-5728(03)00250-9. [DOI] [PubMed] [Google Scholar]
- 16.Dietmann A, Helbok R, Lackner P, et al. Matrix metalloproteinases and their tissue inhibitors (TIMPs) in Plasmodium falciparum malaria: serum levels of TIMP-1 are associated with disease severity. Journal of Infectious Diseases. 2008;197(11):1614–1620. doi: 10.1086/587943. [DOI] [PubMed] [Google Scholar]
- 17.Griffiths MJ, Shafi MJ, Popper SJ, et al. Genomewide analysis of the host response to malaria in Kenyan children. Journal of Infectious Diseases. 2005;191(10):1599–1611. doi: 10.1086/429297. [DOI] [PubMed] [Google Scholar]
- 18.Van den Steen PE, Van Aelst I, Starckx S, Maskos K, Opdenakker G, Pagenstecher A. Matrix metalloproteinases, tissue inhibitors of MMPs and TACE in experimental cerebral malaria. Laboratory Investigation. 2006;86(9):873–888. doi: 10.1038/labinvest.3700454. [DOI] [PubMed] [Google Scholar]
- 19.Szklarczyk A, Stins M, Milward EA, et al. Glial activation and matrix metalloproteinase release in cerebral malaria. Journal of neurovirology. 2007;13(1):2–10. doi: 10.1080/13550280701258084. [DOI] [PubMed] [Google Scholar]
- 20.Olliaro PL, Goldberg DE. The Plasmodium digestive vacuole: metabolic headquarters and choice drug target. Parasitology Today. 1995;11(8):294–297. doi: 10.1016/0169-4758(95)80042-5. [DOI] [PubMed] [Google Scholar]
- 21.Kuhn H, Hache A, Sklenar H. A structural model for the interaction of haem with unsaturated fatty acids explaining its quasi-lipoxygenase activity. Quantum chemical calculations. Biomedica Biochimica Acta. 1983;42(11-12):S175–S176. [PubMed] [Google Scholar]
- 22.Schwarzer E, Kühn H, Valente E, Arese P. Malaria-parasitized erythrocytes and hemozoin nonenzymatically generate large amounts of hydroxy fatty acids that inhibit monocyte functions. Blood. 2003;101(2):722–728. doi: 10.1182/blood-2002-03-0979. [DOI] [PubMed] [Google Scholar]
- 23.Prato M, Giribaldi G, Polimeni M, Gallo V, Arese P. Phagocytosis of hemozoin enhances matrix metalloproteinase-9 activity and TNF-α production in human monocytes: role of matrix metalloproteinases in the pathogenesis of falciparum malaria. Journal of Immunology. 2005;175(10):6436–6442. doi: 10.4049/jimmunol.175.10.6436. [DOI] [PubMed] [Google Scholar]
- 24.Mitola S, Strasly M, Prato M, Ghia P, Bussolino F. IL-12 regulates an endothelial cell-lymphocyte network: effect on metalloproteinase-9 production. Journal of Immunology. 2003;171(7):3725–3733. doi: 10.4049/jimmunol.171.7.3725. [DOI] [PubMed] [Google Scholar]
- 25.Phu NH, Day N, Diep PT, Ferguson DJP, White NJ. Intraleucocytic malaria pigment and prognosis in severe malaria. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1995;89(2):200–204. doi: 10.1016/0035-9203(95)90496-4. [DOI] [PubMed] [Google Scholar]
- 26.Schwarzer E, Bellomo G, Giribaldi G, Ulliers D, Arese P. Phagocytosis of malarial pigment haemozoin by human monocytes: a confocal microscopy study. Parasitology. 2001;123(2):125–131. doi: 10.1017/s0031182001008216. [DOI] [PubMed] [Google Scholar]
- 27.Fiori PL, Rappelli P, Mirkarimi SN, Ginsburg H, Cappuccinelli P, Turrini F. Reduced microbicidal and anti-tumour activities of human monocytes after ingestion of Plasmodium falciparum-infected red blood cells. Parasite Immunology. 1993;15(12):647–655. doi: 10.1111/j.1365-3024.1993.tb00579.x. [DOI] [PubMed] [Google Scholar]
- 28.Schwarzer E, Turrini F, Ulliers D, Giribaldi G, Ginsburg H, Arese P. Impairment of macrophage functions after ingestion of Plasmodium falciparum-infected erythrocytes or isolated malarial pigment. Journal of Experimental Medicine. 1992;176(4):1033–1041. doi: 10.1084/jem.176.4.1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schwarzer E, Alessio M, Ulliers D, Arese P. Phagocytosis of the Malarial Pigment, Hemozoin, Impairs Expression of Major Histocompatibility Complex Class II Antigen, CD54, and CD11c in Human Monocytes. Infection and Immunity. 1998;66(4):1601–1606. doi: 10.1128/iai.66.4.1601-1606.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Schwarzer E, Skorokhod OA, Barrera V, Arese P. Hemozoin and the human monocyte–a brief review of their interactions. Parassitologia. 2008;50(1-2):143–145. [PubMed] [Google Scholar]
- 31.Skorokhod OA, Alessio M, Mordmüller B, Arese P, Schwarzer E. Hemozoin (malarial pigment) inhibits differentiation and maturation of human monocyte-derived dendritic cells: a peroxisome proliferator-activated receptor-γ-mediated effect. Journal of Immunology. 2004;173(6):4066–4074. doi: 10.4049/jimmunol.173.6.4066. [DOI] [PubMed] [Google Scholar]
- 32.Coban C, Ishii KJ, Sullivan DJ, Kumar N. Purified malaria pigment (hemozoin) enhances dendritic cell maturation and modulates the isotype of antibodies induced by a DNA vaccine. Infection and Immunity. 2002;70(7):3939–3943. doi: 10.1128/IAI.70.7.3939-3943.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Coban C, Ishii KJ, Kawai T, et al. Toll-like receptor 9 mediates innate immune activation by the malaria pigment hemozoin. Journal of Experimental Medicine. 2005;201(1):19–25. doi: 10.1084/jem.20041836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Giribaldi G, Ulliers D, Schwarzer E, Roberts I, Piacibello W, Arese P. Hemozoin- and 4-hydroxynonenal-mediated inhibition of erythropoiesis. Possible role in malarial dyserythropoiesis and anemia. Haematologica. 2004;89(4):492–493. [PubMed] [Google Scholar]
- 35.Pichyangkul S, Saengkrai P, Webster HK. Plasmodium falciparum pigment induces monocytes to release high levels of tumor necrosis factor-α and interleukin-1β . American Journal of Tropical Medicine and Hygiene. 1994;51(4):430–435. [PubMed] [Google Scholar]
- 36.Sherry BA, Alava G, Tracey KJ, Martiney J, Cerami A, Slater AFG. Malaria-specific metabolite hemozoin mediates the release of several potent endogenous pyrogens (TNF, MIP-1α, and MIP-1β) in vitro, and altered thermoregulation in vivo. Journal of Inflammation. 1995;45(2):85–96. [PubMed] [Google Scholar]
- 37.Prato M, Gallo V, Giribaldi G, Arese P. Phagocytosis of haemozoin (malarial pigment) enhances metalloproteinase-9 activity in human adherent monocytes: role of IL-1beta and 15-HETE. Malaria Journal. 2008;7:p. 157. doi: 10.1186/1475-2875-7-157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Giribaldi G, Prato M, Ulliers D, et al. Involvement of inflammatory chemokines in survival of human monocytes fed with malarial pigment. Infection and Immunity. 2010;78(11):4912–4921. doi: 10.1128/IAI.00455-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Prato M, Gallo V, Valente E, Khadjavi A, Mandili G, Giribaldi G. Malarial pigment enhances Heat Shock Protein-27 in THP-1 cells: new perspectives for in vitro studies on monocyte apoptosis prevention. Asian Pacific Journal of Tropical Medicine. 2010;3(12):934–938. [Google Scholar]
- 40.Tripathi AK, Sullivan DJ, Stins MF. Plasmodium falciparum-infected erythrocytes increase intercellular adhesion molecule 1 expression on brain endothelium through NF-κB. Infection and Immunity. 2006;74(6):3262–3270. doi: 10.1128/IAI.01625-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Taramelli D, Basilico N, De Palma AM, et al. The effect of synthetic malaria pigment (β-haematin) on adhesion molecule expression and interleukin-6 production by human endothelial cells. Transactions of the Royal Society of Tropical Medicine and Hygiene. 1998;92(1):57–62. doi: 10.1016/s0035-9203(98)90954-3. [DOI] [PubMed] [Google Scholar]
- 42.Basilico N, Speciale L, Parapini S, Ferrante P, Taramelli D. Endothelin-1 production by a microvascular endothelial cell line treated with Plasmodium falciparum parasitized red blood cells. Clinical Science. 2002;103(supplement 48):464S–466S. doi: 10.1042/CS103S464S. [DOI] [PubMed] [Google Scholar]
- 43.Basilico N, Mondani M, Parapini S, Speciale L, Ferrante P, Taramelli D. Plasmodium falciparum parasitized red blood cells modulate the production of endothelin-1 by human endothelial cells. Minerva Medica. 2004;95(2):153–158. [PubMed] [Google Scholar]
- 44.Basilico N, Parapini S, Sisto F, et al. The lipid moiety of haemozoin (Malaria Pigment) and P. falciparum parasitised red blood cells bind synthetic and native endothelin-1. Journal of biomedicine & biotechnology. 2010;2010 doi: 10.1155/2010/854927. Article ID 854927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Van den Steen PE, Dubois B, Nelissen I, Rudd PM, Dwek RA, Opdenakker G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9) Critical Reviews in Biochemistry and Molecular Biology. 2002;37(6):375–536. doi: 10.1080/10409230290771546. [DOI] [PubMed] [Google Scholar]
- 46.Brown HC, Chau TTH, Mai NTH, et al. Blood-brain barrier function in cerebral malaria and CNS infections in Vietnam. Neurology. 2000;55(1):104–111. doi: 10.1212/wnl.55.1.104. [DOI] [PubMed] [Google Scholar]
- 47.Ogata Y, Enghild JJ, Nagase H. Matrix metalloproteinase 3 (stromelysin) activates the precursor for the human matrix metalloproteinase 9. Journal of Biological Chemistry. 1992;267(6):3581–3584. [PubMed] [Google Scholar]
- 48.Suzuki KO, Enghild JJ, Morodomi T, Salvesen G, Nagase H. Mechanisms of activation of tissue procollagenase by matrix metalloproteinase 3 (stromelysin) Biochemistry. 1990;29(44):10261–10270. doi: 10.1021/bi00496a016. [DOI] [PubMed] [Google Scholar]
- 49.Hahn-Dantona E, Ramos-DeSimone N, Sipley J, Nagase H, French DL, Quigley JP. Activation of ProMMP-9 by a plasmin/MMP3 cascade in a tumor cell model. Regulation by tissue inhibitors of metalloproteinases. Annals of the New York Academy of Sciences. 1999;878:372–387. doi: 10.1111/j.1749-6632.1999.tb07696.x. [DOI] [PubMed] [Google Scholar]
- 50.Fauser S, Deininger MH, Kremsner PG, et al. Lesion associated expression of urokinase-type plasminogen activator receptor (uPAR, CD87) in human cerebral malaria. Journal of Neuroimmunology. 2000;111(1-2):234–240. doi: 10.1016/s0165-5728(00)00368-4. [DOI] [PubMed] [Google Scholar]
- 51.Toth M, Chvyrkova I, Bernardo MM, Hernandez-Barrantes S, Fridman R. Pro-MMP-9 activation by the MT1-MMP/MMP-2 axis and MMP-3: role of TIMP-2 and plasma membranes. Biochemical and Biophysical Research Communications. 2003;308(2):386–395. doi: 10.1016/s0006-291x(03)01405-0. [DOI] [PubMed] [Google Scholar]
- 52.Wang FQ, So J, Reierstad S, Fishman DA. Matrilysin (MMP-7) promotes invasion of ovarian cancer cells by activation of progelatinase. International Journal of Cancer. 2005;114(1):19–31. doi: 10.1002/ijc.20697. [DOI] [PubMed] [Google Scholar]
- 53.Visse R, Nagase H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circulation Research. 2003;92(8):827–839. doi: 10.1161/01.RES.0000070112.80711.3D. [DOI] [PubMed] [Google Scholar]
- 54.Murphy G, Houbrechts A, Cockett MI, Williamson RA, O’Shea M, Docherty AJP. The N-terminal domain of tissue inhibitor of metalloproteinases retains metalloproteinase inhibitory activity. Biochemistry. 1991;30(33):8097–8102. doi: 10.1021/bi00247a001. [DOI] [PubMed] [Google Scholar]
- 55.O’Connell JP, Willenbrock F, Docherty AJP, Eaton D, Murphy G. Analysis of the role of the COOH-terminal domain in the activation, proteolytic activity, and tissue inhibitor of metalloproteinase interactions of gelatinase B. Journal of Biological Chemistry. 1994;269(21):14967–14973. [PubMed] [Google Scholar]
- 56.Levin JI. The design and synthesis of aryl hydroxamic acid inhibitors of MMPs and TACE. Current Topics in Medicinal Chemistry. 2004;4(12):1289–1310. doi: 10.2174/1568026043387935. [DOI] [PubMed] [Google Scholar]
- 57.Overall CM, López-Otín C. Strategies for MMP inhibition in cancer: innovations for the post-trial era. Nature Reviews Cancer. 2002;2(9):657–672. doi: 10.1038/nrc884. [DOI] [PubMed] [Google Scholar]
- 58.Hu J, Van den Steen PE, Sang QXA, Opdenakker G. Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nature Reviews Drug Discovery. 2007;6(6):480–498. doi: 10.1038/nrd2308. [DOI] [PubMed] [Google Scholar]
- 59.Wartenberg M, Wolf S, Budde P, et al. The antimalaria agent artemisinin exerts antiangiogenic effects in mouse embryonic stem cell-derived embryoid bodies. Laboratory Investigation. 2003;83(11):1647–1655. doi: 10.1097/01.lab.0000098424.38003.ff. [DOI] [PubMed] [Google Scholar]
- 60.Hwang YP, Yun HJ, Kim HG, Han EH, Lee GW, Jeong HG. Suppression of PMA-induced tumor cell invasion by dihydroartemisinin via inhibition of PKCα/Raf/MAPKs and NF-κB/AP-1-dependent mechanisms. Biochemical Pharmacology. 2010;79(12):1714–1726. doi: 10.1016/j.bcp.2010.02.003. [DOI] [PubMed] [Google Scholar]
- 61.Prato M, Gallo V, Giribaldi G, Aldieri E, Arese P. Role of the NF-κB transcription pathway in the haemozoin- and 15-HETE-mediated activation of matrix metalloproteinase-9 in human adherent monocytes. Cellular Microbiology. 2010;12(12):1780–1791. doi: 10.1111/j.1462-5822.2010.01508.x. [DOI] [PubMed] [Google Scholar]
- 62.Wang Y, Huang Z-Q, Wang C-Q, et al. Artemisinin inhibits extracellular matrix metalloproteinase inducer (EMMPRIN) and matrix metalloproteinase-9 expression via a protein kinase Cδ/p38/extracellular signal-regulated kinase pathway in phorbol myristate acetate-induced THP-1 macrophages. Clinical and Experimental Pharmacology and Physiology. 2011;38(1):11–18. doi: 10.1111/j.1440-1681.2010.05454.x. [DOI] [PubMed] [Google Scholar]
- 63.Gearing AJH, Beckett P, Christodoulou M, et al. Matrix metalloproteinases and processing of pro-TNF-α . Journal of Leukocyte Biology. 1995;57(5):774–777. doi: 10.1002/jlb.57.5.774. [DOI] [PubMed] [Google Scholar]
- 64.Schönbeck U, Mach F, Libby P. Generation of biologically active IL-1β by matrix metalloproteinases: a novel caspase-1-independent pathway of IL-1β processing. Journal of Immunology. 1998;161(7):3340–3346. [PubMed] [Google Scholar]
- 65.Ito A, Mukaiyama A, Itoh Y, et al. Degradation of interleukin 1β by matrix metalloproteinases. Journal of Biological Chemistry. 1996;271(25):14657–14660. doi: 10.1074/jbc.271.25.14657. [DOI] [PubMed] [Google Scholar]
- 66.Yu Q, Stamenkovic I. Cell surface-localized matrix metalloproteinase-9 proteolytically activates TGF-β and promotes tumor invasion and angiogenesis. Genes and Development. 2000;14(2):163–176. [PMC free article] [PubMed] [Google Scholar]
- 67.Van den Steen PE, Proost P, Wuyts A, Van Damme JO, Opdenakker G. Neutrophil gelatinase B potentiates interleukin-8 tenfold by aminoterminal processing, whereas it degrades CTAP-III, PF-4, and GRO-α and leaves RANTES and MCP-2 intact. Blood. 2000;96(8):2673–2681. [PubMed] [Google Scholar]
- 68.Van den Steen PE, Wuyts A, Husson SJ, Proost P, Van Damme JO, Opdenakker G. Gelatinase B/MMP-9 and neutrophil collagenase/MMP-8 process the chemokines human GCP-2/CXCL6, ENA-78/CXCL5 and mouse GCP-2/LIX and modulate their physiological activities. European Journal of Biochemistry. 2003;270(18):3739–3749. doi: 10.1046/j.1432-1033.2003.03760.x. [DOI] [PubMed] [Google Scholar]
- 69.Van den Steen PE, Husson SJ, Proost P, Van Damme JO, Opdenakker G. Carboxyterminal cleavage of the chemokines MIG and IP-10 by gelatinase B and neutrophil collagenase. Biochemical and Biophysical Research Communications. 2003;310(3):889–896. doi: 10.1016/j.bbrc.2003.09.098. [DOI] [PubMed] [Google Scholar]
- 70.McQuibban GA, Butler GS, Gong JH, et al. Matrix metalloproteinase activity inactivates the CXC chemokine stromal cell-derived factor-1. Journal of Biological Chemistry. 2001;276(47):43503–43508. doi: 10.1074/jbc.M107736200. [DOI] [PubMed] [Google Scholar]
- 71.Fiore E, Fusco C, Romero P, Stamenkovic I. Matrix metalloproteinase 9 (MMP-9/gelatinase B) proteolytically cleaves ICAM-1 and participates in tumor cell resistance to natural killer cell-mediated cytotoxicity. Oncogene. 2002;21(34):5213–5223. doi: 10.1038/sj.onc.1205684. [DOI] [PubMed] [Google Scholar]
- 72.Sheu BC, Hsu SM, Ho HN, Lien HC, Huang SC, Lin RH. A novel role of metalloproteinase in cancer-mediated immunosuppression. Cancer Research. 2001;61(1):237–242. [PubMed] [Google Scholar]
- 73.Giebel SJ, Menicucci G, McGuire PG, Das A. Matrix metalloproteinases in early diabetic retinopathy and their role in alternation of the blood-retinal barrier. Laboratory Investigation. 2005;85(5):597–607. doi: 10.1038/labinvest.3700251. [DOI] [PubMed] [Google Scholar]
- 74.Asahi M, Wang X, Mori T, et al. Effects of matrix metalloproteinase-9 gene knock-out on the proteolysis of blood-brain barrier and white matter components after cerebral ischemia. Journal of Neuroscience. 2001;21(19):7724–7732. doi: 10.1523/JNEUROSCI.21-19-07724.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Beauchesne É, Desjardins P, Hazell AS, Butterworth RF. Altered expression of tight junction proteins and matrix metalloproteinases in thiamine-deficient mouse brain. Neurochemistry International. 2009;55(5):275–281. doi: 10.1016/j.neuint.2009.03.014. [DOI] [PubMed] [Google Scholar]
- 76.Ichiyasu H, McCormack JM, McCarthy KM, Dombkowski D, Preffer FI, Schneeberger EE. Matrix metalloproteinase-9-deficient dendritic cells have impaired migration through tracheal epithelial tight junctions. American Journal of Respiratory Cell and Molecular Biology. 2004;30(6):761–770. doi: 10.1165/rcmb.2003-0370OC. [DOI] [PubMed] [Google Scholar]
- 77.Angelow S, Zeni P, Höhn B, Galla HJ. Phorbol ester induced short- and long-term permeabilization of the blood-CSF barrier in vitro. Brain Research. 2005;1063(2):168–179. doi: 10.1016/j.brainres.2005.09.058. [DOI] [PubMed] [Google Scholar]
- 78.Takehara M, Nishimura T, Mima S, Hoshino T, Mizushima T. Effect of claudin expression on paracellular permeability, migration and invasion of colonic cancer cells. Biological and Pharmaceutical Bulletin. 2009;32(5):825–831. doi: 10.1248/bpb.32.825. [DOI] [PubMed] [Google Scholar]
- 79.Yang YI, Estrada EY, Thompson JF, Liu W, Rosenberg GA. Matrix metalloproteinase-mediated disruption of tight junction proteins in cerebral vessels is reversed by synthetic matrix metalloproteinase inhibitor in focal ischemia in rat. Journal of Cerebral Blood Flow and Metabolism. 2007;27(4):697–709. doi: 10.1038/sj.jcbfm.9600375. [DOI] [PubMed] [Google Scholar]
- 80.Brule S, Charnaux N, Sutton A, et al. The shedding of syndecan-4 and syndecan-1 from HeLa cells and human primary macrophages is accelerated by SDF-1/CXCL12 and mediated by the matrix metalloproteinase-9. Glycobiology. 2006;16(6):488–501. doi: 10.1093/glycob/cwj098. [DOI] [PubMed] [Google Scholar]
- 81.Proost P, Van Damme J, Opdenakker G. Leukocyte gelatinase B cleavage releases encephalitogens from human myelin basic protein. Biochemical and Biophysical Research Communications. 1993;192(3):1175–1181. doi: 10.1006/bbrc.1993.1540. [DOI] [PubMed] [Google Scholar]
- 82.Agrawal S, Anderson P, Durbeej M, et al. Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. Journal of Experimental Medicine. 2006;203(4):1007–1016. doi: 10.1084/jem.20051342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Adams S, Brown H, Turner G. Breaking down the blood-brain barrier: signaling a path to cerebral malaria? Trends in Parasitology. 2002;18(8):360–366. doi: 10.1016/s1471-4922(02)02353-x. [DOI] [PubMed] [Google Scholar]
- 84.Brown H, Turner G, Rogerson S, et al. Cytokine expression in the brain in human cerebral malaria. Journal of Infectious Diseases. 1999;180(5):1742–1746. doi: 10.1086/315078. [DOI] [PubMed] [Google Scholar]
- 85.Dietrich JB. The adhesion molecule ICAM-1 and its regulation in relation with the blood-brain barrier. Journal of Neuroimmunology. 2002;128(1-2):58–68. doi: 10.1016/s0165-5728(02)00114-5. [DOI] [PubMed] [Google Scholar]
- 86.Turner G. Cerebral malaria. Brain Pathology. 1997;7(1):569–582. doi: 10.1111/j.1750-3639.1997.tb01075.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Coltel N, Combes V, Hunt NH, Grau GE. Cerebral malaria—a neurovascular pathology with many riddles still to be solved. Current Neurovascular Research. 2004;1(2):91–110. doi: 10.2174/1567202043480116. [DOI] [PubMed] [Google Scholar]
- 88.Medana IM, Turner GDH. Human cerebral malaria and the blood-brain barrier. International Journal for Parasitology. 2006;36(5):555–568. doi: 10.1016/j.ijpara.2006.02.004. [DOI] [PubMed] [Google Scholar]
- 89.Brown H, Hien TT, Day N, et al. Evidence of blood-brain barrier dysfunction in human cerebral malaria. Neuropathology and Applied Neurobiology. 1999;25(4):331–340. doi: 10.1046/j.1365-2990.1999.00188.x. [DOI] [PubMed] [Google Scholar]
- 90.Gillrie MR, Krishnegowda G, Lee K, et al. Src-family kinase-dependent disruption of endothelial barrier function by Plasmodium falciparum merozoite proteins. Blood. 2007;110(9):3426–3435. doi: 10.1182/blood-2007-04-084582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Geurts N, Martens E, Van Aelst I, Proost P, Opdenakker G, Van den Steen PE. β-hematin interaction with the hemopexin domain of gelatinase B/MMP-9 provokes autocatalytic processing of the propeptide, thereby priming activation by MMP-3. Biochemistry. 2008;47(8):2689–2699. doi: 10.1021/bi702260q. [DOI] [PubMed] [Google Scholar]
- 92.Schrimpe AC, Wright DW. Comparative analysis of gene expression changes mediated by individual constituents of hemozoin. Chemical Research in Toxicology. 2009;22(3):433–445. doi: 10.1021/tx8002752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Prato M, Giribaldi G, Arese P. Hemozoin triggers tumor necrosis factor alpha-mediated release of lysozyme by human adherent monocytes: new evidences on leukocyte degranulation in P. falciparum malaria. Asian Pacific Journal of Tropical Medicine. 2009;2(3):35–40. [Google Scholar]
- 94.Prato M, Valente E, Ulliers D, Khadjavi A, Giribaldi G. Quercetin, artemisinin and parthenolide abrogate the haemozoin- and 15-HETE-enhanced activity of enzymes released from gelatinase granules by human monocytes. In: Proceedings of the Antimal Annual Conference; March 2011; London, UK. [Google Scholar]
- 95.Bond M, Fabunmi RP, Baker AH, Newby AC. Synergistic upregulation of metalloproteinase-9 by growth factors and inflammatory cytokines: an absolute requirement for transcription factor NF-κB. FEBS Letters. 1998;435(1):29–34. doi: 10.1016/s0014-5793(98)01034-5. [DOI] [PubMed] [Google Scholar]
- 96.Sasaki K, Hattori T, Fujisawa T, Takahashi K, Inoue H, Takigawa M. Nitric oxide mediates interleukin-1-induced gene expression of matrix metalloproteinases and basic fibroblast growth factor in cultured rabbit articular chondrocytes. Journal of Biochemistry. 1998;123(3):431–439. doi: 10.1093/oxfordjournals.jbchem.a021955. [DOI] [PubMed] [Google Scholar]
- 97.Prato M, D'Alessandro S, Van den Steen PE, et al. Natural haemozoin modulates matrix metalloproteinases and induces morphological changes in human microvascular endothelium. doi: 10.1111/j.1462-5822.2011.01620.x. Cellular Microbiology. In press. [DOI] [PubMed] [Google Scholar]
- 98.Prato M, Gallo V, Arese P. Higher production of tumor necrosis factor alpha in hemozoin-fed-human adherent monocytes is dependent on lipidic component of malarial pigment: new evidences on cytokine regulation in Plasmodium falciparum malaria. Asian Pacific Journal of Tropical Medicine. 2010;3(2):85–89. [Google Scholar]
- 99.Dell'Agli M, Galli GV, Bulgari M, et al. Ellagitannins of the fruit rind of pomegranate (Punica granatum) antagonize in vitro the host inflammatory response mechanisms involved in the onset of malaria. Malaria Journal. 2010;9(1) doi: 10.1186/1475-2875-9-208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Sharma D, Ottino P, Bazan NG, Bazan HE. Epidermal and hepatocyte growth factors, but not keratinocyte growth factor, modulate protein kinase Calpha translocation to the plasma membrane through 15(S)-hydroxyeicosatetraenoic acid synthesis. The Journal of Biological Chemistry. 2005;280(9):7917–7924. doi: 10.1074/jbc.M408852200. [DOI] [PubMed] [Google Scholar]
- 101.Wyke SM, Khal J, Tisdale MJ. Signalling pathways in the induction of proteasome expression by proteolysis-inducing factor in murine myotubes. Cellular Signalling. 2005;17(1):67–75. doi: 10.1016/j.cellsig.2004.05.015. [DOI] [PubMed] [Google Scholar]
- 102.Chen FL, Wang XZ, Li JY, Yu JP, Huang CY, Chen ZX. 12-Lipoxygenase induces apoptosis of human gastric cancer AGS cells via the ERK1/2 signal pathway. Digestive Diseases and Sciences. 2008;53(1):181–187. doi: 10.1007/s10620-007-9841-1. [DOI] [PubMed] [Google Scholar]
- 103.Smith HJ, Wyke SH, Tisdale MJ. Role of protein kinase C and NF-κB in proteolysis-inducing factor-induced proteasome expression in CC myotubes. British Journal of Cancer. 2004;90(9):1850–1857. doi: 10.1038/sj.bjc.6601767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Verma IM, Stevenson J. IkappaB kinase: beginning, not the end. Proceedings of the National Academy of Science of United States of America. 1997;94(22):11758–11760. doi: 10.1073/pnas.94.22.11758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Schwarzer E, Turrini F, Giribaldi G, Cappadoro M, Arese P. Phagocytosis of P. falciparum malarial pigment hemozoin by human monocytes inactivates monocyte protein kinase C. Biochimica et Biophysica Acta. 1993;1181(1):51–54. doi: 10.1016/0925-4439(93)90089-j. [DOI] [PubMed] [Google Scholar]
- 106.Di Mari JF, Saada JI, Mifflin RC, Valentich JD, Powell DW. HETEs enhance IL-1-mediated COX-2 expression via augmentation of message stability in human colonic myofibroblasts. American Journal of Physiology. 2007;293(4):G719–G728. doi: 10.1152/ajpgi.00117.2007. [DOI] [PubMed] [Google Scholar]
- 107.Wen Y, Gu J, Chakrabarti SK, et al. The role of 12/15-lipoxygenase in the expression of interleukin-6 and tumor necrosis factor-α in macrophages. Endocrinology. 2007;148(3):1313–1322. doi: 10.1210/en.2006-0665. [DOI] [PubMed] [Google Scholar]
- 108.Nguyen J, Gogusev J, Knapnougel P, Bauvois B. Protein tyrosine kinase and p38 MAP kinase pathways are involved in stimulation of matrix metalloproteinase-9 by TNF-α in human monocytes. Immunology Letters. 2006;106(1):34–41. doi: 10.1016/j.imlet.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 109.Jaramillo M, Godbout M, Olivier M. Hemozoin induces macrophage chemokine expression through oxidative stress-dependent and -independent mechanisms. Journal of Immunology. 2005;174(1):475–484. doi: 10.4049/jimmunol.174.1.475. [DOI] [PubMed] [Google Scholar]
- 110.Flohé L, Brigelius-Flohé R, Saliou C, Traber MG, Packer L. Redox regulation of NF-kappa B activation. Free Radical Biology and Medicine. 1997;22(6):1115–1126. doi: 10.1016/s0891-5849(96)00501-1. [DOI] [PubMed] [Google Scholar]