

Abstract
Cardiac hypertrophy is a complex adaptive response to mechanical and neurohumoral stimuli and under continual stressor, it contributes to maladaptive responses, heart failure and death. Protein kinase C (PKC) and several other kinases play a role in the maladaptative cardiac responses, including cardiomyocyte hypertrophy, myocardial fibrosis and inflammation. Identifying specific therapies that regulate these kinases is a major focus of current research. PKC, a family of serine/threonine kinases, has emerged as potential mediators of hypertrophic stimuli associated with neurohumoral hyperactivity in heart failure. In this review, we describe the role of PKC isozymes that are involved in cardiac hypertrophy and heart failure.
Keywords: PKC signaling pathways, cardiac remodeling, heart failure
1. Introduction
Cardiac remodeling is a target organ response to cardiovascular diseases and is an independent risk factor for coronary heart disease, stroke, arrhythmias, heart failure, and cardiovascular morbidity and mortality [1–3]. The process of cardiac hypertrophy involves multiple progressive alterations of heart geometry mediated by neurohumoral stimulating stress (i.e. epinephrine, norepinephrine, angiotensin II and aldosterone). Various kinases have been described as candidate mediators of the cardiac biochemical stress and trophic response induced by activation of neurohormone receptors [4]. Subcellular changes associated with cardiac hypertrophy and remodeling may be beneficial in the short term, but are often maladaptative and lead to functional decompensation in the long term. Along with this, the potential molecular events underlying the transition from compensated cardiac hypertrophy to failure are still under investigation. Thus a number of studies have focused on identifying intracellular distal strategic nodes where signals converge and/or serve as multi-effector brakes to suppress or reverse hypertrophy inside the cardiac muscle cell, which would become attractive targets for heart failure pharmacological therapy. In this review, we focus on βIIPKC and εPKC isozymes as potential intracellular nodes that play critical roles in cardiac hypertrophy and failure.
2. PKC isozymes and their distribution in the heart
Identified in 1977 by Nishizuka and coworkers [5], protein kinase C (PKC) is a group of closely related phospholipid-dependent serine-threonine protein kinases, which are activated as a result of receptor-dependent activation of phospholipase C and the hydrolysis of membrane phosphoinositides [6]. The physiological importance of PKC is underscored by the existence of ten different isozymes. These enzymes are classified according to their structure and activation requirements into the following groups: classical or conventional PKCs (α, βI, βII, and γ), which are Ca2+-dependent and activated by binding to diacylglycerol (DAG) and phosphatidylserine (PS); novel PKCs (δ, ε, θ, and η), which are Ca2+-independent but are activated by DAG and PS; and the atypical PKCs (ζ, ι/λ), which are Ca2+ and DAG independent but are PS sensitive. For all PKC isozymes, translocation to membrane structures provides a mechanism to regulate access to substrate and has been taken as the hallmark of activation [7]. At least for some PKC isozymes, auto-phosphorylation on particular sites can also be used as a hallmark of PKC activation [8].
PKC is ubiquitously expressed in all tissues, whereas its distribution is tissue, species and time dependent. PKC isozymes α, δ, ε, η, and ζ have been identified in cultured cardiomyocytes [9, 10]. While the mouse myocardium expresses low levels of both βI and βIIPKC [11], abundant expression of these β PKCs in human and rat cardiomyocytes has been reported [12–14]. Immunoblotting analyses demonstrated the presence of PKC isozymes α, βI, βII, δ, ε, γ, μ, η, λ, and θ in human heart tissue [14]. Further species-specific differences in the expression of cardiac η, θ and εPKC were also reported [15]. Thus, caution must be taken when translating PKC findings from animal models, since expression patterns of specific PKC isozymes differs among species. Further, semi-quantitative assessment of PKC isozymes levels using specific antibodies might provide misleading information on the relative abundance of the PKC isozymes [16]. Using recombinant proteins as a standard, to provide absolute amounts of each PKC isozyme is essential, because of differences in relative immunoreactivity of the isozymes. Finally, temporal expression and activation of different PKC isozymes during ageing and their contribution to disease progression should be considered [17]. For example, during heart failure progression, εPKC and βIIPKC isozymes levels are increased at early (left ventricular hypertrophy) and late (cardiac dysfunction) stages, respectively [18].
3. PKC isozymes in cardiac hypertrophy and heart failure
Prolonged increase of workload triggers cardiac hypertrophy, an adaptative response to normalize wall stress and compensate for the increased neurohormonal stimuli and hemodynamic load. At a cellular level, cardiac myocytes assume a hypertrophic phenotype associated with reactivation of fetal gene programs and quantitative/qualitative changes in the contractile machinery, subcellular organelles, cellular signaling and myocardial metabolism [19, 20]. Several studies have addressed the intracellular mechanisms underlying cardiac hypertrophy and PKC isozymes emerged as potential mediators of hypertrophic stimuli [21, 22]. In fact, PKC activation with PMA (a non-selective PKC activator) causes cardiac myocyte hypertrophy, whereas inhibitory peptides of the cpKCs display an anti-hypertrophic effect [23]. Neurohormones- and mechanical stress-induced cardiac hypertrophic stimuli converge to PKC activation with increased PKC expression and activity in various in vivo models of cardiac hypertrophy [18, 24–26]. Therefore, experimental approaches using rat cultured myocytes, transgenic mice overexpressing PKC isozymes, and isozyme-selective agonist and antagonist peptides have been used to address the relative contribution of PKC isozymes to cardiac hypertrophy and heart failure. Among different PKC isozymes, εPKC and βIIPKC have been considered the main effectors. αPKC has also been involved in rat cardiomyocyte growth and cardiac dysfunction [27, 28].
εPKC is activated in response to hypertrophic stimuli in rat cultured cardiac myocyte and in vivo, and its overexpression in mice leads to cardiac hypertrophy associated with concentric remodeling and preserved cardiac contractility [18, 29]. Treatment with anti-sense εPKC decreased myotrophin-induced stimulation of protein synthesis in neonatal rat cardiomyocytes [30]. In fact, cardiac myocyte-restricted εPKC activation in transgenic mice expressing the εPKC-specific activator (ψεRACK) induces non-pathological hypertrophy, and inhibition of εPKC by a selective inhibitor, εV1 fragment, expressed in low levels in transgenic mice, results in a thin ventricular wall and myocyte hypertrophy [31, 32]. Additionally, high levels of εV1 expression lead to a lethal form of heart failure from dilated cardiomyopathy [31]. We have also demonstrated that εPKC activation during transition from compensated cardiac hypertrophy to heart failure increased mast cell degranulation-induced inflammatory responses, induced cardiac fibrosis and ventricular dysfunction, and significantly reduced animal survival, whereas sustained εPKC inhibition abrogated this pathological phenotype in a rat model [33, 34].
βPKC isozymes also play an important role in cardiac hypertrophy and failure. Even though βPKC is restricted to embryonic and neonatal cardiac myocytes, its expression is upregulated in adult cardiac myocytes under hypertrophic stimuli and in human heart failure [12, 14, 18, 24, 35]. Of interest, increased βIIPKC levels in cultured mouse cardiac myocytes are paralleled by increased ANF and βMHC, genes involved in the transition from cardiac maladaptative hypertrophy to heart failure in cardiac fetal reprogramming [35].
Targeted overexpression of βIIPKC in mouse cardiomyocytes results in left ventricular hypertrophy and fibrosis and oral treatment of the transgenic animal with LY333531, a βPKC inhibitor, prevents cardiac hypertrophy [35], supporting a direct relationship between βIIPKC and the pathological response. In contrast to these findings, another study using βPKC knockout mice showed no role for βPKC in heart failure progression. Thus, the role of βPKC in cardiac hypertrophy in genetically modified mice is controversial. More recently, hypertensive Dahl salt-sensitive rats treated with the βIIPKC-specific inhibitor peptide, but not the βIPKC specific inhibitor, greatly delayed the development of heart failure and improved survival [36]. Similar results were observed in myocardial infarction-induced heart failure in rats treated with βIIPKC-specific inhibitor peptide [36]. Finally, studies characterizing the level and activity of PKC isozymes in human heart failure found a significant increase in level and activation of βPKC [12, 14]. Together, these studies indicate that changes in βIIPKC correlate with human heart failure, suggesting that focusing on this PKC isozyme in considering therapeutic intervention is advisable. A summary of several studies using genetic and pharmacological approaches to determine the role of βIIPKC and εPKC isozymes in cardiac hypertrophy and heart failure is provided in Table 1.
Table 1.
The role of isozyme-specific βIIPKC and εPKC in cardiac remodeling and heart failure
| PKC isozyme | Cardiac phenotype | Model | Authors | Features |
|---|---|---|---|---|
| εPKC | Hypertrophy | Overexpression of constitutively active εPKC | Takeishi et al. [26] | Concentric cardiac hypertrophy |
| εPKC | Hypertrophy | Cardiac-specific expression of εPKC inhibitor, εV1 | Mochly-Rosen et al. [32] | Lethal dilated cardiomyopathy |
| εPKC | Hypertrophy | Cardiac-specific expression of εPKC activator, ψεRACK | Mochly-Rosen et al. [32] | Concentric cardiac hypertrophy |
| εPKC | Hypertrophy | Overexpression of constitutively active εPKC | Pass et al. [16] | Pathological cardiac hypertrophy |
| εPKC | Hypertrophy | Hypertensive rats-treated with εV1-2 (specific εPKC isozyme inhibitor) | Shizukuda et al. [62] | Attenuated isoproterenol-induced cell death |
| εPKC | Hypertrophy | Pressure-overload aortic banding εPKC knock-out mouse | Klein et al. [63] | Increased fibrosis |
| εPKC | Hypertrophy/heart failure | Pressure-overload heart failure rats | Takeishi et al. [64] | ACE inhibitor attenuates increased αPKC and εPKC translocation |
| εPKC/βII PKC | Hypertrophy/heart failure | Dahl Salt hypertensive rats | Koide et al. [65] | Increased cardiac levels of βIIPKC and εPKC |
| εPKC | Heart failure | Hypertensive rats-sustained treatment with εV1-2 (specific εPKC inhibitor) | Inagaki et al. [33] | Decreased cardiac fibrosis |
| βIIPKC | Heart failure | Heart failure rats-non-specific PKC inhibitor (LY333531) | Boyle et al. [46] | Decreased both fibrosis and TGFβ1 expression |
| βIIPKC | Heart failure | Cardiac-specific overexpression of βIIPKC | Wakasaki et al. [35] | Pathologic cardiac hypertrophy and fibrosis |
| βIIPKC | Heart failure | Dahl Salt hypertensive rats | Inagaki et al. [18] | Increased cardiac βIPKC, βIIPKC levels and translocation |
| βIIPKC | Heart failure | Human end-stage dilated or ischemic cardiomyopathy | Bowling et al. [12] | Increased cardiac βPKC activity |
| βIIPKC | Heart failure | Human end-stage dilated cardiomyopathy | Simonis et al. [14] | Increased cardiac βIIPKC levels |
| βIIPKC | Hypertrophy/heart Failure | Cardiac-specific βIIPKC overexpression | Bowman et al. [66] | Pathological cardiac hypertrophy |
| βIIPKC | Hypertrophy | Perfusion with angiotensin II-adult guinea pig heart (ex vivo) | Takeishi et al. [67] | Increased cardiac βIIPKC level |
| βIIPKC | Hypertrophy | Transgenic mice-active calciunerin overexpression | De Windt et al. [68] | Increased cardiac βPKC translocation |
| βIIPKC | Hypertrophy | Streptozotocin-induced diabetic rats | Inoguchi et al. [69] | Increased cardiac βIIPKC activity |
4. PKC targets in cardiac hypertrophy and heart failure
As discussed above, βIIPKC and εPKC isozymes are suited to operate as molecular switches at nodal points in signaling pathways leading to cardiac hypertrophy and heart failure, and downstream mediators of PKC effects have been identified. Pro-hypertrophic stimulation of adult cardiomyocyte cultures with endothelin-1 (ET-1), angiotensin II or phorbol myristate acetate (PMA) resulted in PKC-mediated phosphorylation with further activation of several pro-survival kinases, including mTOR and S6K1[37] (Fig. 1). Indeed, expression of dominant negative εPKC abrogated ET-1 stimulated mTOR and S6K1 phosphorylation, suggesting that εPKC activates mTOR and S6K1 leading to cardiomyocyte hypertrophy (Fig. 1).
Figure 1.

βIIPKC and εPKC isozyme signaling pathways and downstream targets in cardiac remodeling and heart failure.
Another kinase that has been implicated in PKC-mediated hypertrophic signaling is ERK, a MAPK involved in growth and cell survival (Fig. 1). Under hypertrophic stimuli, εPKC acts as an upstream regulator of the Ras/Raf/-ERK cascade [38] and mediates GPCR-dependent mobilization of transcription factors in cardiomyocyte cultures [39]. Indeed, using a constitutively-activated εPKC mutant, Heidkamp et al. [40] demonstrated that εPKC selectively activates ERK, resulting in cardiomyocyte remodeling. Of interest, using the same approach for δPKC, they observed that δPKC preferentially activates JNK and p38, which are implicated in stress-activated protein kinase cascades. Taken together these studies support the involvement of both εPKC and δPKC isozymes in MAPK cascade activation with distinctly different MAPK pathway downstream targets.
As described above, increased βPKC levels in rat primary cardiac myocytes under hypertrophic stimuli are paralleled by increased ANF expression, and the mechanism underlying this response seems to be related to GATA-4, a transcription factor that mediates βPKC activation of the ANF promoter in response to pro-hypertrophic Ang II, ultimately resulting in enhanced DNA binding activity [41]. In another study, Lim et al. showed that PKC-dependent TAK1 phosphorylation and ATF2 (activating transcription factor 2) activation is involved in TGF-β1-induced cardiac hypertrophy [42]. Indeed, non-selective PKC inhibition completely blocked TGF-β1-induced TAK1 kinase activity and subsequent downstream signaling, such as inhibition of β-MHC gene induction and ANF promoter activity. Lastly, protein kinase D, another downstream effector of PKC, directly phosphorylates HDAC5 and stimulates its nuclear export and cardiac hypertrophy, whereas a non-selective PKC inhibitor (GF 109203X) prevents nuclear export of HDAC5 in response to hypertrophic agonists [43]. According to these studies, neurohumoral activation of GPCR engages classical and novel PKC pathways towards cardiac hypertrophy-related gene reprogramming (Fig. 1). Therefore, reduced myofilament responsiveness to Ca2+ associated with a significant increase in troponin I phosphorylation levels have been seen in the myocardium of transgenic mice overexpressing βIIPKC. The depressed cardiomyocyte function improved after sequential perfusion of LY333531, a βPKC inhibitor. This study shows that βIIPKC-mediated phosphorylation of troponin I in vivo may decrease the Ca2+ responsiveness of myofilaments, and thus lead to cardiac myocyte dysfunction [44]. Depressed myofilament contractility-associated myocyte dysfunction was also observed in αPKC transgenic mice [28].
In addition to the effects of βIIPKC on cardiac hypertrophy-related gene reprogramming proteins and contractile myofilaments, our laboratory has recently identified new βIIPKC targets associated with protein quality control disruption in heart failure [45]. The cellular protein quality control (PQC) is set to detect, repair and dispose of cytotoxic damaged proteins by multilayered control mechanisms including chaperones, the ubiquitin-proteasome system and autophagy. We found that increased βIIPKC activity leads to proteasome dysfunction, the main effector of the ubiquitin-proteasome system, and contributes to heart failure development [45] (Ferreira et al in preparation). Therefore, blocking βIIPKC and proteasome interaction using a rationally designed peptide inhibitor for βIIPKC improves cardiac function and survival in both ischemic and hypertensive-induced heart failure in rats. These phenomena were mediated by increasing proteasomal activity and re-established cardiac protein quality control. A summary of βIIPKC and εPKC isozyme-mediated cellular responses in cardiac hypertrophy and heart failure is provided in Figure 2.
Figure 2.
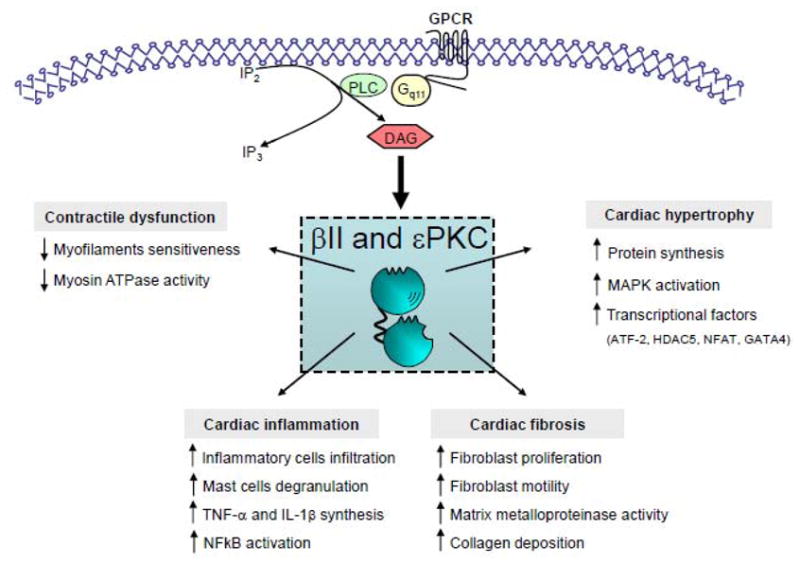
Schematic βIIPKC and εPKC isozyme-mediated cellular responses in cardiac remodeling and heart failure.
5. PKC inhibitors
Because specific PKC isozymes contribute to a wide variety of human diseases and sometimes exert even opposing effects in the same disease, the need to produce highly selective pharmacological PKC inhibitors is highlighted. A group of bisindolyl malelmide (BIM) compounds that are based on the scaffold of the nonspecific kinase inhibitor staurosporine have been a focus of active research. These compounds were reported to function as βPKC selective inhibitors and reverse cardiac remodeling and ventricular dysfunction [44, 46]. However, several staurosporine-originated PKC inhibitors have been tested in animal models and clinical trial and were found to have undesirable toxicity. Moreover, specific PKC inhibitors such as LV333531, BIM1and BMI2 inhibit the activity of several PKC isozymes as well as the activity of other kinases [47]. For example, the βPKC-selective inhibitor (LV333531) that is currently in clinical trial for diabetic retinopathy and diabetic macular edema has been reported to affect other PKC isozymes [48].
Our lab has taken another approach to generate PKC isozyme-selective pharmacological tools. We used a rational design to identity the interaction site between each PKC isozyme and its selective anchoring protein (RACK) and identified peptides corresponding to these sites that selectively interfere with the interaction. Importantly, we showed that inhibition of interaction of a particular isozyme with its RACK causes a selective loss of the function mediated by that isozyme without affecting the function of other PKC isozymes that are present in the same cells (reviewed elsewhere [49]). The peptides are delivered into cells and in vivo by cross-linking them to cell-permeating peptides, such as TAT47–57. These peptide inhibitors are highly selective and efficacious in treating animals used as models for different human diseases where specific PKC isozyme is activated [50–54]. Relevant to the topic of this review, selective inhibition of βIIPKC and εPKC with these peptide inhibitors significantly improved cardiac function and prolonged survival in different heart failure animal models [22, 33, 45, 53]. Therefore, although intermolecular proteins interactions are considered to be difficult to target for therapeutic, short peptides corresponding to sequences that mediate protein-protein interactions may offer a new class of drugs for treating a number of diseases including cardiovascular diseases.
6. Summary and perspectives
Taken together, studies using cultured cardiomyocytes, transgenic animals and selective pharmacological tools suggest that both βIIPKC and εPKC are key molecules involved in cardiac hypertrophy and heart failure. Avoiding pathological cardiac remodeling is a goal of heart failure therapy [55]. In this review, we describe a continuum of responses emanating from βIIPKC and εPKC isozymes that contribute to decompensated hypertrophy and heart failure (see Table 1 and Figure 1). Because specific modulators of PKC isozymes are already in clinical trials for a variety of diseases [56–59], it may now be possible to consider using βIIPKC and εPKC isozyme-specific inhibitors to treat human heart failure. Therefore, recent clinical trials suggest that systemic delivery of inhibitors of PKC isozymes is well tolerated [57, 58, 60]. Further, advances in drug delivery suggest that organ-selective delivery may also be another possibility in the near future, for example by delivering slow drug-releasing molecules to specific organs [61]. Therefore, clinical trials with βIIPKC and εPKC inhibitors to treat heart failure should be considered using either systemic or cardiac-specific drug delivery. Further in vivo studies using larger animal models will help to point out the relevance of βIIPKC and εPKC as therapeutic targets for the treatment of maladaptative cardiac remodeling and heart failure in humans.
Acknowledgments
Funding
This study was supported by National Institute of Health Grant HL076675 and HL52141 to DMR. JCF holds a post-doctoral fellowship from Fundação de Amparo a Pesquisa do Estado de São Paulo - Brasil (FAPESP 2009/03143-1). PCB holds a scholarship from Conselho Nacional de Pesquisa e Desenvolvimento, Brasil (CNPq BPQ 301519/2008-0).
Abbreviations
- ANF
atrial natriuretic factor
- ATF 2
activating transcription factor 2
- βMHC
β myosin heavy chain
- DAG
diacylglycerol
- ERK
extracellular signal-regulated kinase
- ET-1
endothelin-1
- GATA 4
cardiac-restricted zinc finger protein
- GPCR
G protein-coupled receptors
- HDAC
histone deacetylase
- JNK
Jun N-terminal kinase
- MAPK
mitogen-activated protein kinase
- mtor
mammalian target of rapamycin
- PKC
protein kinase C
- PMA
phorbol myristate acetate
- PS
phosphatidylserine
- Raf
Raf proto-oncogene serine/threonine-protein kinase
- Ras
small guanosine triphosphate hydrolase
- S6K1
p70 ribosomal protein S6 kinase 1
- TAK1
TGF-β activated kinase 1
- TGF-β
transforming growth factor β
Footnotes
Disclosures
DM-R is the founder of KAI Pharmaceuticals, Inc, a company that plans to bring PKC regulators to the clinic. However, none of the work described in this study is based on or supported by the company. Other authors have no disclosure.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Artham SM, Lavie CJ, Milani RV, Patel DA, Verma A, Ventura HO. Clinical impact of left ventricular hypertrophy and implications for regression. Prog Cardiovasc Dis. 2009;52:153–67. doi: 10.1016/j.pcad.2009.05.002. [DOI] [PubMed] [Google Scholar]
- 2.Levy D, Garrison RJ, Savage DD, Kannel WB, Castelli WP. Prognostic implications of echocardiographically determined left ventricular mass in the Framingham Heart Study. N Engl J Med. 1990;322:1561–6. doi: 10.1056/NEJM199005313222203. [DOI] [PubMed] [Google Scholar]
- 3.Messerli FH, Ventura HO, Elizardi DJ, Dunn FG, Frohlich ED. Hypertension and sudden death. Increased ventricular ectopic activity in left ventricular hypertrophy. Am J Med. 1984;77:18–22. doi: 10.1016/0002-9343(84)90430-3. [DOI] [PubMed] [Google Scholar]
- 4.Dorn GW, 2nd, Force T. Protein kinase cascades in the regulation of cardiac hypertrophy. J Clin Invest. 2005;115:527–37. doi: 10.1172/JCI24178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Inoue M, Kishimoto A, Takai Y, Nishizuka Y. Studies on a cyclic nucleotide-independent protein kinase and its proenzyme in mammalian tissues. II. Proenzyme and its activation by calcium-dependent protease from rat brain. J Biol Chem. 1977;252:7610–6. [PubMed] [Google Scholar]
- 6.Dempsey EC, Newton AC, Mochly-Rosen D, Fields AP, Reyland ME, Insel PA, et al. Protein kinase C isozymes and the regulation of diverse cell responses. Am J Physiol Lung Cell Mol Physiol. 2000;279:L429–38. doi: 10.1152/ajplung.2000.279.3.L429. [DOI] [PubMed] [Google Scholar]
- 7.Mochly-Rosen D, Gordon AS. Anchoring proteins for protein kinase C: a means for isozyme selectivity. FASEB J. 1998;12:35–42. [PubMed] [Google Scholar]
- 8.Rybin VO, Guo J, Harleton E, Feinmark SJ, Steinberg SF. Regulatory autophosphorylation sites on protein kinase C-delta at threonine-141 and threonine-295. Biochemistry. 2009;48:4642–51. doi: 10.1021/bi802171c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Erdbrugger W, Keffel J, Knocks M, Otto T, Philipp T, Michel MC. Protein kinase C isoenzymes in rat and human cardiovascular tissues. Br J Pharmacol. 1997;120:177–86. doi: 10.1038/sj.bjp.0700877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kohout TA, Rogers TB. Use of a PCR-based method to characterize protein kinase C isoform expression in cardiac cells. Am J Physiol. 1993;264:C1350–9. doi: 10.1152/ajpcell.1993.264.5.C1350. [DOI] [PubMed] [Google Scholar]
- 11.Schreiber KL, Paquet L, Allen BG, Rindt H. Protein kinase C isoform expression and activity in the mouse heart. Am J Physiol Heart Circ Physiol. 2001;281:H2062–71. doi: 10.1152/ajpheart.2001.281.5.H2062. [DOI] [PubMed] [Google Scholar]
- 12.Bowling N, Walsh RA, Song G, Estridge T, Sandusky GE, Fouts RL, et al. Increased protein kinase C activity and expression of Ca2+-sensitive isoforms in the failing human heart. Circulation. 1999;99:384–91. doi: 10.1161/01.cir.99.3.384. [DOI] [PubMed] [Google Scholar]
- 13.Shin HG, Barnett JV, Chang P, Reddy S, Drinkwater DC, Pierson RN, et al. Molecular heterogeneity of protein kinase C expression in human ventricle. Cardiovasc Res. 2000;48:285–99. doi: 10.1016/s0008-6363(00)00185-1. [DOI] [PubMed] [Google Scholar]
- 14.Simonis G, Briem SK, Schoen SP, Bock M, Marquetant R, Strasser RH. Protein kinase C in the human heart: differential regulation of the isoforms in aortic stenosis or dilated cardiomyopathy. Mol Cell Biochem. 2007;305:103–11. doi: 10.1007/s11010-007-9533-3. [DOI] [PubMed] [Google Scholar]
- 15.Rouet-Benzineb P, Mohammadi K, Perennec J, Poyard M, Bouanani Nel H, Crozatier B. Protein kinase C isoform expression in normal and failing rabbit hearts. Circ Res. 1996;79:153–61. doi: 10.1161/01.res.79.2.153. [DOI] [PubMed] [Google Scholar]
- 16.Pass JM, Gao J, Jones WK, Wead WB, Wu X, Zhang J, et al. Enhanced PKC beta II translocation and PKC beta II-RACK1 interactions in PKC epsilon-induced heart failure: a role for RACK1. Am J Physiol Heart Circ Physiol. 2001;281:H2500–10. doi: 10.1152/ajpheart.2001.281.6.H2500. [DOI] [PubMed] [Google Scholar]
- 17.Goldspink P, Ruch S, Los T, Buttrick P, Garcia J. Maladaptation of calcium homoeostasis in aging cardiac myocytes. Pflugers Arch. 2008;456:479–87. doi: 10.1007/s00424-007-0420-2. [DOI] [PubMed] [Google Scholar]
- 18.Inagaki K, Iwanaga Y, Sarai N, Onozawa Y, Takenaka H, Mochly-Rosen D, et al. Tissue angiotensin II during progression or ventricular hypertrophy to heart failure in hypertensive rats; differential effects on PKC epsilon and PKC beta. J Mol Cell Cardiol. 2002;34:1377–85. doi: 10.1006/jmcc.2002.2089. [DOI] [PubMed] [Google Scholar]
- 19.Bernardo BC, Weeks KL, Pretorius L, McMullen JR. Molecular distinction between physiological and pathological cardiac hypertrophy: Experimental findings and therapeutic strategies. Pharmacol Ther. 2010;128:191–27. doi: 10.1016/j.pharmthera.2010.04.005. [DOI] [PubMed] [Google Scholar]
- 20.Machackova J, Barta J, Dhalla NS. Myofibrillar remodeling in cardiac hypertrophy, heart failure and cardiomyopathies. Can J Cardiol. 2006;22:953–68. doi: 10.1016/s0828-282x(06)70315-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Churchill E, Budas G, Vallentin A, Koyanagi T, Mochly-Rosen D. PKC isozymes in chronic cardiac disease: possible therapeutic targets? Annu Rev Pharmacol Toxicol. 2008;48:569–99. doi: 10.1146/annurev.pharmtox.48.121806.154902. [DOI] [PubMed] [Google Scholar]
- 22.Palaniyandi SS, Sun L, Ferreira JC, Mochly-Rosen D. Protein kinase C in heart failure: a therapeutic target? Cardiovasc Res. 2009;82:229–39. doi: 10.1093/cvr/cvp001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stebbins EG, Mochly-Rosen D. Binding specificity for RACK1 resides in the V5 region of beta II protein kinase C. J Biol Chem. 2001;276:29644–50. doi: 10.1074/jbc.M101044200. [DOI] [PubMed] [Google Scholar]
- 24.Bogoyevitch MA, Parker PJ, Sugden PH. Characterization of protein kinase C isotype expression in adult rat heart. Protein kinase C-epsilon is a major isotype present, and it is activated by phorbol esters, epinephrine, and endothelin. Circ Res. 1993;72:757–67. doi: 10.1161/01.res.72.4.757. [DOI] [PubMed] [Google Scholar]
- 25.Pass JM, Zheng Y, Wead WB, Zhang J, Li RC, Bolli R, et al. PKCepsilon activation induces dichotomous cardiac phenotypes and modulates PKCepsilon-RACK interactions and RACK expression. Am J Physiol Heart Circ Physiol. 2001;280:H946–55. doi: 10.1152/ajpheart.2001.280.3.H946. [DOI] [PubMed] [Google Scholar]
- 26.Takeishi Y, Ping P, Bolli R, Kirkpatrick DL, Hoit BD, Walsh RA. Transgenic overexpression of constitutively active protein kinase C epsilon causes concentric cardiac hypertrophy. Circ Res. 2000;86:1218–23. doi: 10.1161/01.res.86.12.1218. [DOI] [PubMed] [Google Scholar]
- 27.Braz JC, Bueno OF, De Windt LJ, Molkentin JD. PKC alpha regulates the hypertrophic growth of cardiomyocytes through extracellular signal-regulated kinase1/2 (ERK1/2) J Cell Biol. 2002;156:905–19. doi: 10.1083/jcb.200108062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Braz JC, Gregory K, Pathak A, Zhao W, Sahin B, Klevitsky R, et al. PKC-alpha regulates cardiac contractility and propensity toward heart failure. Nat Med. 2004;10:248–54. doi: 10.1038/nm1000. [DOI] [PubMed] [Google Scholar]
- 29.Chen L, Hahn H, Wu G, Chen CH, Liron T, Schechtman D, et al. Opposing cardioprotective actions and parallel hypertrophic effects of delta PKC and epsilon PKC. Proc Natl Acad Sci U S A. 2001;98:11114–9. doi: 10.1073/pnas.191369098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sil P, Kandaswamy V, Sen S. Increased protein kinase C activity in myotrophin-induced myocyte growth. Circ Res. 1998;82:1173–88. doi: 10.1161/01.res.82.11.1173. [DOI] [PubMed] [Google Scholar]
- 31.Dorn GW, 2nd, Souroujon MC, Liron T, Chen CH, Gray MO, Zhou HZ, et al. Sustained in vivo cardiac protection by a rationally designed peptide that causes epsilon protein kinase C translocation. Proc Natl Acad Sci U S A. 1999;96:12798–803. doi: 10.1073/pnas.96.22.12798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mochly-Rosen D, Wu G, Hahn H, Osinska H, Liron T, Lorenz JN, et al. Cardiotrophic effects of protein kinase C epsilon: analysis by in vivo modulation of PKCepsilon translocation. Circ Res. 2000;86:1173–9. doi: 10.1161/01.res.86.11.1173. [DOI] [PubMed] [Google Scholar]
- 33.Inagaki K, Koyanagi T, Berry NC, Sun L, Mochly-Rosen D. Pharmacological inhibition of epsilon-protein kinase C attenuates cardiac fibrosis and dysfunction in hypertension-induced heart failure. Hypertension. 2008;51:1565–9. doi: 10.1161/HYPERTENSIONAHA.107.109637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Palaniyandi SS, Inagaki K, Mochly-Rosen D. Mast cells and epsilonPKC: a role in cardiac remodeling in hypertension-induced heart failure. J Mol Cell Cardiol. 2008;45:779–86. doi: 10.1016/j.yjmcc.2008.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wakasaki H, Koya D, Schoen FJ, Jirousek MR, Ways DK, Hoit BD, et al. Targeted overexpression of protein kinase C beta2 isoform in myocardium causes cardiomyopathy. Proc Natl Acad Sci U S A. 1997;94:9320–5. doi: 10.1073/pnas.94.17.9320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ferreira JC, Inagaki K, Fajardo G, Churchill E, Budas G, Disatnik M, et al. Pharmacological βIIPKC inhibition is cardioprotective in late-stage hypertrophy and end-stage heart failure in two rat models. Circulation. 2008;118:S535. [Google Scholar]
- 37.Moschella PC, Rao VU, McDermott PJ, Kuppuswamy D. Regulation of mTOR and S6K1 activation by the nPKC isoforms, PKCepsilon and PKCdelta, in adult cardiac muscle cells. J Mol Cell Cardiol. 2007;43:754–66. doi: 10.1016/j.yjmcc.2007.09.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Clerk A, Bogoyevitch MA, Anderson MB, Sugden PH. Differential activation of protein kinase C isoforms by endothelin-1 and phenylephrine and subsequent stimulation of p42 and p44 mitogen-activated protein kinases in ventricular myocytes cultured from neonatal rat hearts. J Biol Chem. 1994;269:32848–57. [PubMed] [Google Scholar]
- 39.Singal T, Dhalla NS, Tappia PS. Regulation of c-Fos and c-Jun gene expression by phospholipase C activity in adult cardiomyocytes. Mol Cell Biochem. 2009;327:229–39. doi: 10.1007/s11010-009-0061-1. [DOI] [PubMed] [Google Scholar]
- 40.Heidkamp MC, Bayer AL, Martin JL, Samarel AM. Differential activation of mitogen-activated protein kinase cascades and apoptosis by protein kinase C epsilon and delta in neonatal rat ventricular myocytes. Circ Res. 2001;89:882–90. doi: 10.1161/hh2201.099434. [DOI] [PubMed] [Google Scholar]
- 41.Wang J, Paradis P, Aries A, Komati H, Lefebvre C, Wang H, et al. Convergence of protein kinase C and JAK-STAT signaling on transcription factor GATA-4. Mol Cell Biol. 2005;25:9829–44. doi: 10.1128/MCB.25.22.9829-9844.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lim JY, Park SJ, Hwang HY, Park EJ, Nam JH, Kim J, et al. TGF-beta1 induces cardiac hypertrophic responses via PKC-dependent ATF-2 activation. J Mol Cell Cardiol. 2005;39:627–36. doi: 10.1016/j.yjmcc.2005.06.016. [DOI] [PubMed] [Google Scholar]
- 43.Vega RB, Harrison BC, Meadows E, Roberts CR, Papst PJ, Olson EN, et al. Protein kinases C and D mediate agonist-dependent cardiac hypertrophy through nuclear export of histone deacetylase 5. Mol Cell Biol. 2004;24:8374–85. doi: 10.1128/MCB.24.19.8374-8385.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Takeishi Y, Chu G, Kirkpatrick DM, Li Z, Wakasaki H, Kranias EG, et al. In vivo phosphorylation of cardiac troponin I by protein kinase Cbeta2 decreases cardiomyocyte calcium responsiveness and contractility in transgenic mouse hearts. J Clin Invest. 1998;102:72–8. doi: 10.1172/JCI2709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ferreira JC, Boer BN, Grimberg M, Brum PC, Mochly-Rosen D. Protein quality control disruption by PKCβII in heart failure. ISHR North American Section Meeting; 2009. [Google Scholar]
- 46.Boyle AJ, Kelly DJ, Zhang Y, Cox AJ, Gow RM, Way K, et al. Inhibition of protein kinase C reduces left ventricular fibrosis and dysfunction following myocardial infarction. J Mol Cell Cardiol. 2005;39:213–21. doi: 10.1016/j.yjmcc.2005.03.008. [DOI] [PubMed] [Google Scholar]
- 47.Komander D, Kular GS, Schuttelkopf AW, Deak M, Prakash KR, Bain J, et al. Interactions of LY333531 and other bisindolyl maleimide inhibitors with PDK1. Structure. 2004;12:215–26. doi: 10.1016/j.str.2004.01.005. [DOI] [PubMed] [Google Scholar]
- 48.Graff JR, McNulty AM, Hanna KR, Konicek BW, Lynch RL, Bailey SN, et al. The protein kinase Cbeta-selective inhibitor, Enzastaurin ( LY317615.HCl), suppresses signaling through the AKT pathway, induces apoptosis, and suppresses growth of human colon cancer and glioblastoma xenografts. Cancer Res. 2005;65:7462–9. doi: 10.1158/0008-5472.CAN-05-0071. [DOI] [PubMed] [Google Scholar]
- 49.Souroujon MC, Mochly-Rosen D. Peptide modulators of protein-protein interactions in intracellular signaling. Nat Biotechnol. 1998;16:919–24. doi: 10.1038/nbt1098-919. [DOI] [PubMed] [Google Scholar]
- 50.Chen CH, Budas GR, Churchill EN, Disatnik MH, Hurley TD, Mochly-Rosen D. Activation of aldehyde dehydrogenase-2 reduces ischemic damage to the heart. Science. 2008;321:1493–5. doi: 10.1126/science.1158554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Churchill EN, Ferreira JC, Brum PC, Szweda LI, Mochly-Rosen D. Ischaemic preconditioning improves proteasomal activity and increases the degradation of deltaPKC during reperfusion. Cardiovasc Res. 2010;85:385–94. doi: 10.1093/cvr/cvp334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kim J, Choi YL, Vallentin A, Hunrichs BS, Hellerstein MK, Peehl DM, et al. Centrosomal PKCbetaII and pericentrin are critical for human prostate cancer growth and angiogenesis. Cancer Res. 2008;68:6831–9. doi: 10.1158/0008-5472.CAN-07-6195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Koyanagi T, Noguchi K, Ootani A, Inagaki K, Robbins RC, Mochly-Rosen D. Pharmacological inhibition of epsilon PKC suppresses chronic inflammation in murine cardiac transplantation model. J Mol Cell Cardiol. 2007;43:517–22. doi: 10.1016/j.yjmcc.2007.06.003. [DOI] [PubMed] [Google Scholar]
- 54.Qi X, Inagaki K, Sobel RA, Mochly-Rosen D. Sustained pharmacological inhibition of deltaPKC protects against hypertensive encephalopathy through prevention of blood-brain barrier breakdown in rats. J Clin Invest. 2008;118:173–82. doi: 10.1172/JCI32636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cohn JN, Ferrari R, Sharpe N. Cardiac remodeling--concepts and clinical implications: a consensus paper from an international forum on cardiac remodeling. Behalf of an International Forum on Cardiac Remodeling. J Am Coll Cardiol. 2000;35:569–82. doi: 10.1016/s0735-1097(99)00630-0. [DOI] [PubMed] [Google Scholar]
- 56.Advani R, Peethambaram P, Lum BL, Fisher GA, Hartmann L, Long HJ, et al. A Phase II trial of aprinocarsen, an antisense oligonucleotide inhibitor of protein kinase C alpha, administered as a 21-day infusion to patients with advanced ovarian carcinoma. Cancer. 2004;100:321–6. doi: 10.1002/cncr.11909. [DOI] [PubMed] [Google Scholar]
- 57.Aiello LP, Davis MD, Girach A, Kles KA, Milton RC, Sheetz MJ, et al. Effect of ruboxistaurin on visual loss in patients with diabetic retinopathy. Ophthalmology. 2006;113:2221–30. doi: 10.1016/j.ophtha.2006.07.032. [DOI] [PubMed] [Google Scholar]
- 58.Bates E, Bode C, Costa M, Gibson CM, Granger C, Green C, et al. Intracoronary KAI-9803 as an adjunct to primary percutaneous coronary intervention for acute ST-segment elevation myocardial infarction. Circulation. 2008;117:886–96. doi: 10.1161/CIRCULATIONAHA.107.759167. [DOI] [PubMed] [Google Scholar]
- 59.Grossman SA, Alavi JB, Supko JG, Carson KA, Priet R, Dorr FA, et al. Efficacy and toxicity of the antisense oligonucleotide aprinocarsen directed against protein kinase C-alpha delivered as a 21-day continuous intravenous infusion in patients with recurrent high-grade astrocytomas. Neuro Oncol. 2005;7:32–40. doi: 10.1215/S1152851703000353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Effect of ruboxistaurin in patients with diabetic macular edema: thirty-month results of the randomized PKC-DMES clinical trial. Arch Ophthalmol. 2007;125:318–24. doi: 10.1001/archopht.125.3.318. [DOI] [PubMed] [Google Scholar]
- 61.Mayer CR, Bekeredjian R. Ultrasonic gene and drug delivery to the cardiovascular system. Adv Drug Deliv Rev. 2008;60:1177–92. doi: 10.1016/j.addr.2008.03.004. [DOI] [PubMed] [Google Scholar]
- 62.Shizukuda Y, Buttrick PM. Protein kinase C(epsilon) modulates apoptosis induced by beta -adrenergic stimulation in adult rat ventricular myocytes via extracellular signal-regulated kinase (ERK) activity. J Mol Cell Cardiol. 2001;33:1791–803. doi: 10.1006/jmcc.2001.1442. [DOI] [PubMed] [Google Scholar]
- 63.Klein G, Schaefer A, Hilfiker-Kleiner D, Oppermann D, Shukla P, Quint A, et al. Increased collagen deposition and diastolic dysfunction but preserved myocardial hypertrophy after pressure overload in mice lacking PKCepsilon. Circ Res. 2005;96:748–55. doi: 10.1161/01.RES.0000161999.86198.1e. [DOI] [PubMed] [Google Scholar]
- 64.Takeishi Y, Bhagwat A, Ball NA, Kirkpatrick DL, Periasamy M, Walsh RA. Effect of angiotensin-converting enzyme inhibition on protein kinase C and SR proteins in heart failure. Am J Physiol. 1999;276:H53–62. doi: 10.1152/ajpheart.1999.276.1.H53. [DOI] [PubMed] [Google Scholar]
- 65.Koide Y, Tamura K, Suzuki A, Kitamura K, Yokoyama K, Hashimoto T, et al. Differential induction of protein kinase C isoforms at the cardiac hypertrophy stage and congestive heart failure stage in Dahl salt-sensitive rats. Hypertens Res. 2003;26:421–6. doi: 10.1291/hypres.26.421. [DOI] [PubMed] [Google Scholar]
- 66.Bowman JC, Steinberg SF, Jiang T, Geenen DL, Fishman GI, Buttrick PM. Expression of protein kinase C beta in the heart causes hypertrophy in adult mice and sudden death in neonates. J Clin Invest. 1997;100:2189–95. doi: 10.1172/JCI119755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Takeishi Y, Jalili T, Ball NA, Walsh RA. Responses of cardiac protein kinase C isoforms to distinct pathological stimuli are differentially regulated. Circ Res. 1999;85:264–71. doi: 10.1161/01.res.85.3.264. [DOI] [PubMed] [Google Scholar]
- 68.De Windt LJ, Lim HW, Haq S, Force T, Molkentin JD. Calcineurin promotes protein kinase C and c-Jun NH2-terminal kinase activation in the heart. Cross-talk between cardiac hypertrophic signaling pathways. J Biol Chem. 2000;275:13571–9. doi: 10.1074/jbc.275.18.13571. [DOI] [PubMed] [Google Scholar]
- 69.Inoguchi T, Battan R, Handler E, Sportsman JR, Heath W, King GL. Preferential elevation of protein kinase C isoform beta II and diacylglycerol levels in the aorta and heart of diabetic rats: differential reversibility to glycemic control by islet cell transplantation. Proc Natl Acad Sci U S A. 1992;89:11059–63. doi: 10.1073/pnas.89.22.11059. [DOI] [PMC free article] [PubMed] [Google Scholar]