

Abstract
Ikaros (also known as Lyf-1) was initially described as a lymphoid-specific transcription factor. Although Ikaros has been shown to regulate hematopoietic stem cell renewal, as well as the development and function of cells from multiple hematopoietic lineages, including the myeloid lineage, Ikaros has primarily been studied in context of lymphoid development and malignancy. This review focuses on the role of Ikaros in myeloid cells. We address the importance of post-transcriptional regulation of Ikaros function; the emerging role of Ikaros in myeloid malignancy; Ikaros as a regulator of myeloid differentiation and function; and the selective expression of Ikaros isoform-x in cells with myeloid potential. We highlight the challenges of dissecting Ikaros function in lineage commitment decisions among lymphoid-myeloid progenitors that have emerged as a major myeloid differentiation pathway in recent studies, which leads to reconstruction of the traditional map of murine and human hematopoiesis.
Keywords: Ikaros, Myeloid differentiation, Lineage commitment, Hematopoiesis, Post-transcriptional regulation, Post-translational regulation
OVERVIEW OF IKAROS FUNCTION
The Ikaros gene product is alternately spliced to produce multiple zinc finger proteins (Figure 1) that bind to promoter regions and regulate target gene expression[1,2]. Ikaros is essential for normal hematopoiesis[3] and has been implicated as a tumor suppressor[4,5]. Ikaros has been shown to both activate[6-9] and repress[10-14] gene expression and to participate in chromatin remodeling where it targets genes for epigenetic modifications and recruitment to pericentromeric heterochromatin[15]. Little is known about the mechanisms that regulate the expression of the Ikaros gene at the transcriptional level[16], although a variety of mechanisms for post-transcriptional regulation of Ikaros activity and/or protein levels have been identified.
Figure 1.
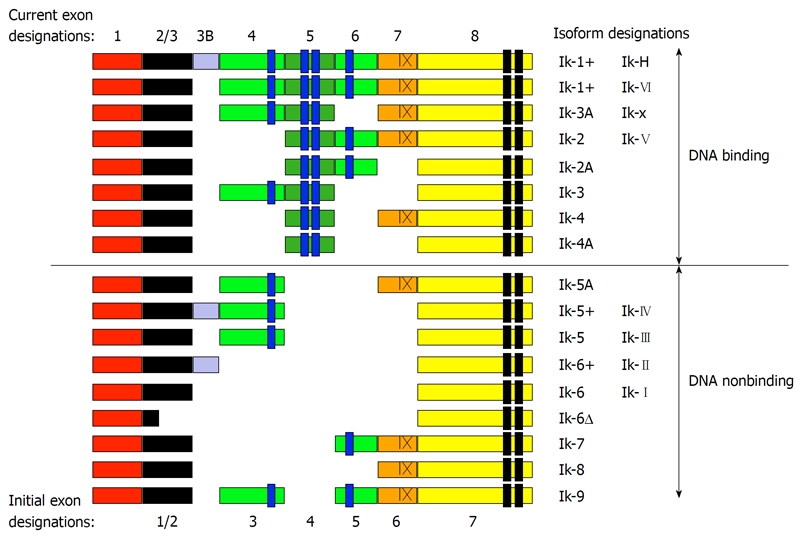
Ikaros isoforms. The Ikaros gene in mice (Ikzf1) and humans (IKZF1) includes eight coding exons (exons 2-8 and 3B) and one upstream exon that is not translated (Figure 1). The untranslated exon (shown in red) has not been identified in initial reports, and the alternate exon designations that have appeared in early reports are shown at the bottom of the figure. Exon 3B is currently not identified as an exon in Genbank. Splice forms that include exon 3B have been designated as “plus” forms and many such splice forms, in addition to the ones shown, have been identified at the protein and/or mRNA level (i.e. Ik-x+, Ik-2+, Ik-4+, Ik-7+, Ik-8+) in humans and mice. An alternate splice site gives rise to splice forms that lack the last 30 bases of exon 7 (indicated with an X). Such splice variants have been designated minus forms (e.g. Ik-1- and Ik-x-). The four N-terminal zinc fingers (shown in blue) contribute to DNA binding and the two C-terminal zinc fingers (shown in black) are responsible for dimerization[1,2,19,47-53].
Ikaros activity is dependent on its ability to bind DNA. Ikaros binds DNA via four N-terminal zinc fingers[1]. The loss of zinc fingers through alternate splicing or chromosomal deletion can result in Ikaros isoforms with reduced DNA binding affinity[1,2]. DNA-nonbinding Ikaros isoforms exert a dominant negative (DN) effect, inhibiting the ability of other Ikaros isoforms and Ikaros family members (e.g. Aiolos and Helios) to bind DNA[4]. Phosphorylation of Ikaros by casein kinase 2 (CK2)[17] has been shown to inhibit the ability of Ikaros to interact with DNA, including pericentromeric hetorochromatin[15], and to regulate Ikaros susceptibility to ubiquitin-mediated degradation[18]. Thus, in addition to transcriptional regulation of the Ikaros gene product, the level of Ikaros DNA binding activity can be regulated by at least four other mechanisms: (1) alternative splicing that results in altered Ikaros DNA binding; (2) DN Ikaros isoforms that inhibit the DNA-binding activity of other Ikaros isoforms; (3) phosphorylation of Ikaros that results in decreased DNA binding affinity; and (4) phosphorylation-mediated changes in the stability of Ikaros proteins.
Ikaros activity can be regulated by post-transcriptional events that are independent of its DNA binding affinity. In addition to changes in DNA binding affinity, alternative splicing gives rise to Ikaros isoforms with altered DNA binding specificity[1,2,19]. This provides a potential mechanism for fine tuning Ikaros targets in different cell types. Repression of target genes by Ikaros can be mediated by recruitment of histone deacetylase (HDAC)-containing complexes[20], as well as through the HDAC-independent mediator, CtBP[21]. Sumoylation of Ikaros has been shown to inhibit the ability of Ikaros to interact with both HDAC-dependent and HDAC-independent repressors of transcription[22]. The multiple mechanisms by which Ikaros activity can be regulated post-transcriptionally suggest that the complete picture of Ikaros function is likely to be complex and underscores the importance of Ikaros studies at the protein level.
IKAROS AS A REGULATOR OF MYELOID CELL DIFFERENTIATION AND FUNCTION
Ikaros was identified using strategies designed to detect transcription factors that regulate lymphoid genes and was initially described as a lymphoid-specific transcription factor[2,23,24]. Ikaros mutant mice are characterized by profound lymphoid defects and as a consequence Ikaros has primarily been studied in context of lymphoid development and function and lymphoid malignancy. The central role of Ikaros in the lymphoid lineages has largely overshadowed the more subtle, yet crucial roles that Ikaros plays in the myeloid lineage.
Defects in myelopoiesis are present in all of the Ikaros mutant mouse models that have been described to date. Homozygous IkarosDN mutants show a reduction in the number of myeloid lineage cells in the bone marrow. Terminal granulocyte differentiation is absent in homozygous IkarosDN[25] and Ikarosnull[3] mutants, in IKL/L mice that express very low levels of Ikaros[26], and in the plastic mutant mouse strain[27] that harbors a point mutation that prevents Ikaros from binding DNA.
A comprehensive analysis of neutrophil differentiation in the IKL/L mice showed defects in neutrophil survival and migration as well as a failure of immature granulocytes to upregulate Gr-1 (also a characteristic of other Ikaros mutants); a differentiation event that is preceded by high levels of Ikaros protein expression in wild-type mice[26]. Ikaros has also been shown to regulate the expression of inducible nitric oxide synthase downstream of lipopolysaccharide/interferon-γ stimulation in a macrophage cell line[28]. These data provide evidence that Ikaros regulates differentiation and immune function in the myeloid lineages as it does in the lymphoid lineages.
IKAROS AS A TUMOR SUPPRESSOR
Studies of Ikaros mutant mice together with clinical data provide compelling evidence that Ikaros acts as a tumor suppressor. The rapid development of T-cell lymphoma in mice that are heterozygous for a defect that produces DN Ikaros isoforms (IKDN) provided the first data to support a role for Ikaros in tumor suppression[4]. IKL/L mice that express low levels of Ikaros also develop T-cell lymphoma[11]. Similarly, multiple clinical studies have linked Ikaros mutations and deletions to human B-cell acute lymphoblastic leukemia and to a lesser extent T--cell acute lymphoblastic leukemia[29-39]. Ikaros has been shown to regulate expression of molecules that control cell cycle progression and cell survival[40,41], as well as hematopoietic differentiation; all of which are likely to contribute to the tumor suppressor activity of Ikaros. More recently, Ikaros defects have been linked to myeloproliferative neoplasms[42,43] and childhood acute myelogenous leukemia[40], providing evidence that Ikaros tumor suppressor activity extends to the myeloid lineage. In these cases, Ikaros activity is lost due to deletion of the Ikaros gene[42,43] or expression of DN Ikaros isoforms[40]. The mechanisms that regulate Ikaros tumor suppressor activity have not been defined[16]. However, the roles of CK2 in post-transcriptional regulation of Ikaros protein levels[18] and in functionally inactivating Ikaros[44] suggest that CK2 may be involved in regulating Ikaros tumor suppressor function. Overexpression of CK2 has been associated with myeloid malignancies[45,46]. Thus, overexpression of CK2 is a potential mechanism for the functional inactivation of Ikaros that leads to the loss of Ikaros tumor suppressor activity in myeloid leukemia.
EXPRESSION OF IKAROS ISOFORM-X IS ASSOCIATED WITH MYELOID POTENTIAL
It is important to note that Ikaros proteins are often divided into two broad categories, DNA-binding and DNA-nonbinding, and indeed this is a key distinction in studies of malignant hematopoiesis. However, the DNA-binding Ikaros is often treated as if it were one protein, although this is not the case. The Ikaros gene in mice (ikzf1) and humans (IKZF1) includes eight coding exons and one upstream exon that is not translated (Figure 1). Ikaros coding exons are alternately spliced to produce more than 10 DNA binding and nonbinding Ikaros isoforms[1] detectable by reverse transcription polymerase chain reaction (RT-PCR)[1,2,19,47-53]. The detection of Ikaros transcripts by RT-PCR correlates poorly with protein expression as measured by immunoblotting[54].
At the protein level, Ikaros expression in normal human bone marrow is dominated by the expression of Ikaros-x (Ik-x), followed by Ik-1, Ik-H, and Ik-2/3 (Ik-2 and Ik-3 are indistinguishable by molecular weight) with very little expression of DNA-nonbinding isoforms[54]. The expression pattern of Ikaros proteins is similar in murine hematopoietic cells, with the exception that expression of the Ik-H protein is largely absent in mice[19,54]. Although all of these isoforms bind DNA, they differentially incorporate exon 3B and/or the exons that encode the four N-terminal zinc fingers that contribute to DNA binding. The alternate use of these exons has been reported to fine tune the DNA binding specificity and/or affinity of Ikaros proteins[1,2,19]. Thus, differential expression of Ikaros isoforms is a potential mechanism for regulating the expression of Ikaros target genes.
The differential expression of Ikaros proteins observed in different hematopoietic lineages provides further evidence that alternative splicing is a mechanism for regulating Ikaros activity (Figure 2). B, natural killer (NK), activated T cells, and nucleated erythroid lineage cells express all of the major isoforms[54]. Surprisingly, Ikaros is largely absent in resting human T cells but is upregulated upon activation[19,54]. In contrast, Ik-x proteins in the hematopoietic system are detected exclusively in myeloid lineage cells or progenitors with myeloid differentiation potential. Human Lin- CD34+ hematopoietic stem cells (HSCs) express both Ik-x and Ik-1. When placed in cultures that selectively support lymphoid or myeloid differentiation, there is a loss of Ik-x under lymphoid conditions and an upregulation of Ik-x under myeloid conditions[54]. CD14+ monocytes express both Ik-1 and Ik-x while terminally differentiated granulocytes express only Ik-x[19,54]. These data identify the Ik-x isoform as a potential candidate for mediating myeloid lineage commitment decisions.
Figure 2.
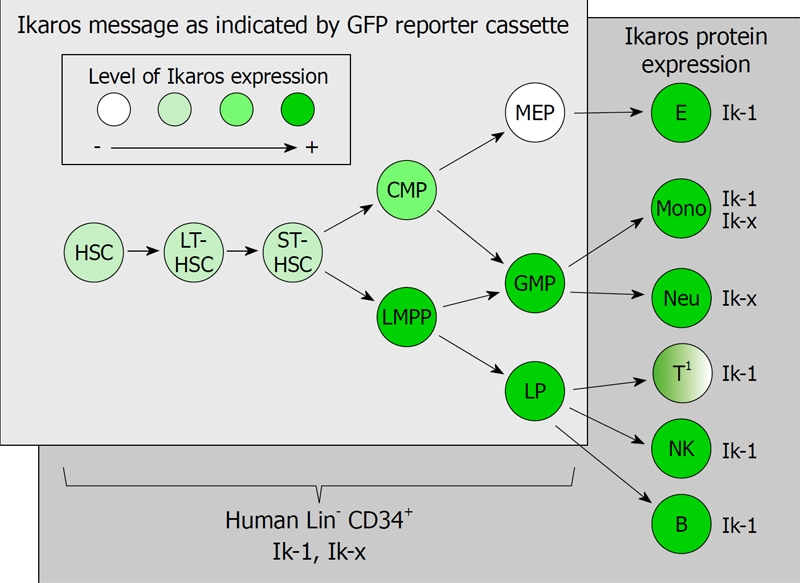
Overview of Ikaros expression in hematopoiesis. The relative expression levels of Ikaros mRNA in hematopoietic progenitors (light gray box in foreground) are indicated by levels of green shading. Progenitors from both the classic and the LMPP pathways are included. Relative Ikaros expression levels are from studies by Yoshida et al[63] that have used mice with an Ikaros reporter cassette [green fluorescent protein (GFP) down stream of an Ikaros promoter] as an indicator of Ikaros mRNA expression in early stages of murine hematopoiesis. Shown in the dark gray box in the background is the protein level expression of human Ikaros isoforms in differentiated hematopoietic cells and in Lin-CD34+ hematopoietic progenitors as described by Ronniet al[19] and Payne et al[54]. Human cells that express Ik-x and/or Ik-1 also express the plus form of these isoforms[19,54] (i.e. Ik-1+/Ik-H and Ik-x+, see Figure 1). 1Mature T cells express little Ikaros protein unless they are activated[19,54]. HSC: Hematopoietic stem cell; LT-HSC: Long-term-HSC; ST-HSC: Short-term-HSC; CMP: Common myeloid progenitor; GMP: Granulocyte/macrophage progenitor; LMPP: Progenitor with lymphoid and myeloid potential; MEP: Megakaryocyte/erythroid progenitor; LP: Lymphoid progenitor; NK: Natural killer.
Studies that examine Ikaros DNA-binding activity have largely focused on one isoform-Ik-1. Several factors have likely contributed to this. Ikaros studies have targeted the lymphoid lineages where Ik-x is not expressed and have been performed in mice, in which Ik-H is largely absent. If we are to obtain a clearer picture of Ikaros function in myelopoiesis, it will be important to widen our scope to include Ik-x; the most abundantly expressed Ikaros isoform. It will also be important to consider how the interplay between Ik-x and Ik-1 and/or other less abundant Ikaros isoforms might contribute to myeloid lineage commitment and granulocyte vs macrophage specification in human myeloid differentiation.
IKAROS AND MYELOID COMMITMENT IN THE CLASSIC AND LMPP HEMATOPOIETIC PATHWAYS
Protein studies are essential for elucidating the complete picture of Ikaros function as a mediator of hematopoietic lineage commitment. Nevertheless, transcriptional studies of normal and Ikaros null mice that express an Ikaros reporter cassette, coupled with bioinformatics approaches, are contributing to the outline of an important new pathway for myelopoiesis. Primitive multipotent HSCs give rise successively to long-term-HSCs and short-term (ST)-HSCs, which repopulate HSCs with multi-lineage potential, but increasingly limited self-renewal capacity[55] (Figure 2). According to the classic model of hematopoietic differentiation, the segregation of lymphoid and myelo-erythroid differentiation is an early event that occurs as ST-HSCs become either common myeloid progenitors (CMPs) or common lymphoid progenitors (CLPs). The CMPs then give rise to megakaryocyte/erythroid progenitors and granulocyte/macrophage progenitors (GMPs) and their lineage restricted progeny, while the CLPs generate all of the lymphoid lineages (T, B, NK and lymphoid dendritic cells). Evidence for the classic differentiation pathway has been reported in both mice and humans[56-59]. However, accumulating evidence suggests the existence of differentiation pathways that are distinct from the classic model of hematopoietic differentiation. Early branching of the megakaryocyte/erythroid lineages from progenitors with lymphoid and myeloid potential (LMPPs) have been demonstrated downstream of ST-HSCs in mice[60,61] and just recently in humans[62] (Figure 2). The extent to which the classic hematopoietic pathway or the more recently identified LMPP pathway contributes to normal myelopoiesis remains controversial. Whether a particular pathway predominates at a given point in ontogeny and/or whether one pathway overlays another throughout life is not yet clear.
Mice with an Ikaros reporter cassette produced by expressing the green fluorescent protein (GFP) under the control of an Ikaros promoter have been used to evaluate the relationship between Ikaros expression and hematopoietic lineage potential[63], thus shedding light on the role of Ikaros in lineage commitment decisions (Figure 2). Based on GFP expression and corresponding levels of Ikaros, early hematopoietic progenitors from these mice have been isolated, into Ikaros-, IkarosINT, and Ikaros+ populations. Ikaros- progenitors show expression of early erythroid-lineage genes (e.g. Gata1 or Gata2) but not myeloid promoting genes. Progenitors with intermediate levels of Ikaros have been shown to co-express early erythroid and myeloid genes, and consistent with the classic model of hematopoiesis, to contain functional CMPs with the capacity to generate myeloid and erythroid lineage cells[63].
Surprisingly, progenitors with high levels of Ikaros express early genes that promote early myeloid differentiation events (e.g. Gfi1b, Csf3r or Cebpa) as well as lymphoid promoting genes (e.g. Flt3, Rag1 or Il7r), but not erythroid-associated genes. Consistent with gene expression, functional assays have demonstrated that the progenitors that express the highest levels of Ikaros have both lymphoid and myeloid but not erythroid potential[63]. These data are consistent with the emerging picture of hematopoiesis in which the classic hematopoietic model co-exists with a hematopoietic program in which lymphoid and myeloid potential is segregated from erythroid differentiation early in hematopoiesis (Figure 2).
The role of Ikaros in the classic and LMPP hematopoietic pathways has been investigated using Ikaros null mice that are engineered to express the GFP Ikaros reporter cassette[63]. These mice, while lacking Ikaros, have expressed the GFP Ikaros reporter in a manner similar to that observed in wild-type mice. Analysis of (GFPINT) progenitors from wild-type mice and their Ikaros null counterparts show that the ability of the classic CMPs to produce myeloid-committed GMPs is substantially reduced in the absence of Ikaros. In contrast, LMPPs which express high levels of Ikaros in wild-type mice (GFP+) show increased ability to generate myeloid-committed cells in the Ikaros null mice[63].
These studies provide a glimpse of the complex role of Ikaros in myeloid differentiation, in which its impact has previously been obscured due to reciprocal effects on the CMPs and LMPPs. The above data suggest that low level Ikaros expression can promote myeloid instead of erythroid differentiation among CMPs with myelo-erythroid potential in the classic hematopoietic pathway. The role of Ikaros in lymphoid vs myeloid lineage commitment in LMPPs is less clear. Although Ikaros is required for the production of lymphoid lineage cells from LMPPs, this is not the case for myeloid cells. In fact, their numbers are increased with the loss of Ikaros. Thus Ikaros is clearly not a requirement for myeloid commitment among LMPPs.
The story of Ikaros in lineage commitment entered a new era when bioinformatics approaches made it possible to perform highly sensitive whole genome analysis of gene expression in small numbers of hematopoietic progenitors. Studies of the newly defined LMPP pathway[64] have shown that normal HSCs express not only genes that are associated with HSC function, such as self-renewal, but also low level expression of genes associated with early erythroid, myeloid and lymphoid differentiation. This expression is thought to prime progenitors for subsequent differentiation events that proceed in a stochastic manner. A comparison of normal progenitors in the LMPP pathway and their Ikaros null counterparts (identified using the GFP Ikaros cassette[63] as described above) gives important clues to Ikaros function in hematopoietic differentiation. Ikaros null mice fail to appropriately shut down expression of HSC-associated genes in downstream progenitors. These mice also fail to downregulate early erythroid-associated genes and to upregulate early lymphoid-associated genes. Dysregulated myeloid gene expression is observed in these mice, that is, genes such as Cfs1r, Cebpd and Id2 that are usually associated with late myeloid differentiation are upregulated in HSCs as well as in the LMPPs[64].
The failure of Ikaros null mice to suppress myeloid genes is consistent with the commonly accepted idea that Ikaros promotes lymphoid differentiation by inhibiting the expression of myeloid genes, and thereby, myeloid differentiation. However, models for lymphoid vs myeloid cell fate specification that account for the maintenance of myeloid potential in LMPPs and downstream GMPs that express high levels of the Ikaros gene product[63] have not been described. The lineage commitment studies discussed above were limited in that they examined Ikaros transcripts. The extent to which Ikaros transcripts correspond to Ikaros protein expression, and the particular isoforms expressed in normal hematopoietic progenitors, are hard to determine because of the technical limitations of obtaining adequate cell numbers. The status of Ikaros protein activity due to post-translational modifications is an important question to address because of the potential for phosphorylation and SUMOylation to affect Ikaros function. Given the high levels of Ikaros transcripts reported in the largely myeloid-committed GMPs[63], it seems likely that differential isoform expression or post-translational regulatory mechanisms will at least be a part of the Ikaros story in lymphoid vs myeloid lineage commitment.
CONCLUSION
Ikaros is a key regulator of normal and malignant hematopoiesis in multiple lineages, including the myeloid lineage. Currently, very little is known of the mechanisms that regulate Ikaros gene expression[16]. Information on the factors that control Ikaros expression is likely to provide important new insights. However, the fact that Ikaros transcripts are detected at similar levels in a wide range of progenitors and lineages, where Ikaros can have opposing effects, suggests that much of the Ikaros story may be found at the protein level. This could pose challenges for studies that involve hematopoietic progenitors or primary human samples in which cell numbers are often limiting. To get a complete picture of the roles that Ikaros plays in normal and malignant hematopoiesis, including myelopoiesis, it will be important to distinguish the parts played by the various Ikaros isoforms and to identify other cast members such as CK2, chromatin remodeling complexes, and other Ikaros family members that regulate and cooperate in Ikaros activities.
Footnotes
Peer reviewers: Jae Youl Cho, PhD, Associate Professor, College of Biomedical Science, Kangwon National University, 192-1 Hyoja-2-dong, Chuncheon 200-701, South Korea; Yi-Guo Zhang, PhD, MD, Professor, Laboratory of Cell Biochemistry and Gene Regulation, College of Bioengineering and Life Sciencces, University of Chongqing, Chongqing 400044, China
S- Editor Cheng JX L- Editor Kerr C E- Editor Zheng XM
References
- 1.Molnár A, Georgopoulos K. The Ikaros gene encodes a family of functionally diverse zinc finger DNA-binding proteins. Mol Cell Biol. 1994;14:8292–8303. doi: 10.1128/mcb.14.12.8292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hahm K, Ernst P, Lo K, Kim GS, Turck C, Smale ST. The lymphoid transcription factor LyF-1 is encoded by specific, alternatively spliced mRNAs derived from the Ikaros gene. Mol Cell Biol. 1994;14:7111–7123. doi: 10.1128/mcb.14.11.7111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wang JH, Nichogiannopoulou A, Wu L, Sun L, Sharpe AH, Bigby M, Georgopoulos K. Selective defects in the development of the fetal and adult lymphoid system in mice with an Ikaros null mutation. Immunity. 1996;5:537–549. doi: 10.1016/s1074-7613(00)80269-1. [DOI] [PubMed] [Google Scholar]
- 4.Winandy S, Wu P, Georgopoulos K. A dominant mutation in the Ikaros gene leads to rapid development of leukemia and lymphoma. Cell. 1995;83:289–299. doi: 10.1016/0092-8674(95)90170-1. [DOI] [PubMed] [Google Scholar]
- 5.Avitahl N, Winandy S, Friedrich C, Jones B, Ge Y, Georgopoulos K. Ikaros sets thresholds for T cell activation and regulates chromosome propagation. Immunity. 1999;10:333–343. doi: 10.1016/s1074-7613(00)80033-3. [DOI] [PubMed] [Google Scholar]
- 6.Harker N, Naito T, Cortes M, Hostert A, Hirschberg S, Tolaini M, Roderick K, Georgopoulos K, Kioussis D. The CD8alpha gene locus is regulated by the Ikaros family of proteins. Mol Cell. 2002;10:1403–1415. doi: 10.1016/s1097-2765(02)00711-6. [DOI] [PubMed] [Google Scholar]
- 7.Umetsu SE, Winandy S. Ikaros is a regulator of Il10 expression in CD4+ T cells. J Immunol. 2009;183:5518–5525. doi: 10.4049/jimmunol.0901284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wargnier A, Lafaurie C, Legros-Maïda S, Bourge JF, Sigaux F, Sasportes M, Paul P. Down-regulation of human granzyme B expression by glucocorticoids. Dexamethasone inhibits binding to the Ikaros and AP-1 regulatory elements of the granzyme B promoter. J Biol Chem. 1998;273:35326–35331. doi: 10.1074/jbc.273.52.35326. [DOI] [PubMed] [Google Scholar]
- 9.Ghanshani S, Wulff H, Miller MJ, Rohm H, Neben A, Gutman GA, Cahalan MD, Chandy KG. Up-regulation of the IKCa1 potassium channel during T-cell activation. Molecular mechanism and functional consequences. J Biol Chem. 2000;275:37137–37149. doi: 10.1074/jbc.M003941200. [DOI] [PubMed] [Google Scholar]
- 10.Kathrein KL, Chari S, Winandy S. Ikaros directly represses the notch target gene Hes1 in a leukemia T cell line: implications for CD4 regulation. J Biol Chem. 2008;283:10476–10484. doi: 10.1074/jbc.M709643200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Dumortier A, Jeannet R, Kirstetter P, Kleinmann E, Sellars M, dos Santos NR, Thibault C, Barths J, Ghysdael J, Punt JA, et al. Notch activation is an early and critical event during T-Cell leukemogenesis in Ikaros-deficient mice. Mol Cell Biol. 2006;26:209–220. doi: 10.1128/MCB.26.1.209-220.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Trinh LA, Ferrini R, Cobb BS, Weinmann AS, Hahm K, Ernst P, Garraway IP, Merkenschlager M, Smale ST. Down-regulation of TDT transcription in CD4(+)CD8(+) thymocytes by Ikaros proteins in direct competition with an Ets activator. Genes Dev. 2001;15:1817–1832. doi: 10.1101/gad.905601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dorsam G, Goetzl EJ. Vasoactive intestinal peptide receptor-1 (VPAC-1) is a novel gene target of the hemolymphopoietic transcription factor Ikaros. J Biol Chem. 2002;277:13488–13493. doi: 10.1074/jbc.M107922200. [DOI] [PubMed] [Google Scholar]
- 14.Sabbattini P, Lundgren M, Georgiou A, Chow C, Warnes G, Dillon N. Binding of Ikaros to the lambda5 promoter silences transcription through a mechanism that does not require heterochromatin formation. EMBO J. 2001;20:2812–2822. doi: 10.1093/emboj/20.11.2812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cobb BS, Morales-Alcelay S, Kleiger G, Brown KE, Fisher AG, Smale ST. Targeting of Ikaros to pericentromeric heterochromatin by direct DNA binding. Genes Dev. 2000;14:2146–2160. doi: 10.1101/gad.816400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liberg D, Smale ST, Merkenschlager M. Upstream of Ikaros. Trends Immunol. 2003;24:567–570. doi: 10.1016/j.it.2003.09.008. [DOI] [PubMed] [Google Scholar]
- 17.Gómez-del Arco P, Maki K, Georgopoulos K. Phosphorylation controls Ikaros's ability to negatively regulate the G(1)-S transition. Mol Cell Biol. 2004;24:2797–2807. doi: 10.1128/MCB.24.7.2797-2807.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Popescu M, Gurel Z, Ronni T, Song C, Hung KY, Payne KJ, Dovat S. Ikaros stability and pericentromeric localization are regulated by protein phosphatase 1. J Biol Chem. 2009;284:13869–13880. doi: 10.1074/jbc.M900209200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ronni T, Payne KJ, Ho S, Bradley MN, Dorsam G, Dovat S. Human Ikaros function in activated T cells is regulated by coordinated expression of its largest isoforms. J Biol Chem. 2007;282:2538–2547. doi: 10.1074/jbc.M605627200. [DOI] [PubMed] [Google Scholar]
- 20.Kim J, Sif S, Jones B, Jackson A, Koipally J, Heller E, Winandy S, Viel A, Sawyer A, Ikeda T, et al. Ikaros DNA-binding proteins direct formation of chromatin remodeling complexes in lymphocytes. Immunity. 1999;10:345–355. doi: 10.1016/s1074-7613(00)80034-5. [DOI] [PubMed] [Google Scholar]
- 21.Koipally J, Georgopoulos K. Ikaros interactions with CtBP reveal a repression mechanism that is independent of histone deacetylase activity. J Biol Chem. 2000;275:19594–19602. doi: 10.1074/jbc.M000254200. [DOI] [PubMed] [Google Scholar]
- 22.Gómez-del Arco P, Koipally J, Georgopoulos K. Ikaros SUMOylation: switching out of repression. Mol Cell Biol. 2005;25:2688–2697. doi: 10.1128/MCB.25.7.2688-2697.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lo K, Landau NR, Smale ST. LyF-1, a transcriptional regulator that interacts with a novel class of promoters for lymphocyte-specific genes. Mol Cell Biol. 1991;11:5229–5243. doi: 10.1128/mcb.11.10.5229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Georgopoulos K, Moore DD, Derfler B. Ikaros, an early lymphoid-specific transcription factor and a putative mediator for T cell commitment. Science. 1992;258:808–812. doi: 10.1126/science.1439790. [DOI] [PubMed] [Google Scholar]
- 25.Georgopoulos K, Bigby M, Wang JH, Molnar A, Wu P, Winandy S, Sharpe A. The Ikaros gene is required for the development of all lymphoid lineages. Cell. 1994;79:143–156. doi: 10.1016/0092-8674(94)90407-3. [DOI] [PubMed] [Google Scholar]
- 26.Dumortier A, Kirstetter P, Kastner P, Chan S. Ikaros regulates neutrophil differentiation. Blood. 2003;101:2219–2226. doi: 10.1182/blood-2002-05-1336. [DOI] [PubMed] [Google Scholar]
- 27.Papathanasiou P, Perkins AC, Cobb BS, Ferrini R, Sridharan R, Hoyne GF, Nelms KA, Smale ST, Goodnow CC. Widespread failure of hematolymphoid differentiation caused by a recessive niche-filling allele of the Ikaros transcription factor. Immunity. 2003;19:131–144. doi: 10.1016/s1074-7613(03)00168-7. [DOI] [PubMed] [Google Scholar]
- 28.Cho SJ, Huh JE, Song J, Rhee DK, Pyo S. Ikaros negatively regulates inducible nitric oxide synthase expression in macrophages: involvement of Ikaros phosphorylation by casein kinase 2. Cell Mol Life Sci. 2008;65:3290–3303. doi: 10.1007/s00018-008-8332-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Harvey RC, Mullighan CG, Chen IM, Wharton W, Mikhail FM, Carroll AJ, Kang H, Liu W, Dobbin KK, Smith MA, et al. Rearrangement of CRLF2 is associated with mutation of JAK kinases, alteration of IKZF1, Hispanic/Latino ethnicity, and a poor outcome in pediatric B-progenitor acute lymphoblastic leukemia. Blood. 2010;115:5312–5321. doi: 10.1182/blood-2009-09-245944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Karp JE, Merz WG, Dick JD, Saral R. Strategies to prevent or control infections after bone marrow transplants. Bone Marrow Transplant. 1991;8:1–6. [PubMed] [Google Scholar]
- 31.Kang H, Chen IM, Wilson CS, Bedrick EJ, Harvey RC, Atlas SR, Devidas M, Mullighan CG, Wang X, Murphy M, et al. Gene expression classifiers for relapse-free survival and minimal residual disease improve risk classification and outcome prediction in pediatric B-precursor acute lymphoblastic leukemia. Blood. 2010;115:1394–1405. doi: 10.1182/blood-2009-05-218560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Mullighan CG, Goorha S, Radtke I, Miller CB, Coustan-Smith E, Dalton JD, Girtman K, Mathew S, Ma J, Pounds SB, et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature. 2007;446:758–764. doi: 10.1038/nature05690. [DOI] [PubMed] [Google Scholar]
- 33.Mullighan CG, Miller CB, Radtke I, Phillips LA, Dalton J, Ma J, White D, Hughes TP, Le Beau MM, Pui CH, et al. BCR-ABL1 lymphoblastic leukaemia is characterized by the deletion of Ikaros. Nature. 2008;453:110–114. doi: 10.1038/nature06866. [DOI] [PubMed] [Google Scholar]
- 34.Marçais A, Jeannet R, Hernandez L, Soulier J, Sigaux F, Chan S, Kastner P. Genetic inactivation of Ikaros is a rare event in human T-ALL. Leuk Res. 2010;34:426–429. doi: 10.1016/j.leukres.2009.09.012. [DOI] [PubMed] [Google Scholar]
- 35.Den Boer ML, van Slegtenhorst M, De Menezes RX, Cheok MH, Buijs-Gladdines JG, Peters ST, Van Zutven LJ, Beverloo HB, Van der Spek PJ, Escherich G, et al. A subtype of childhood acute lymphoblastic leukaemia with poor treatment outcome: a genome-wide classification study. Lancet Oncol. 2009;10:125–134. doi: 10.1016/S1470-2045(08)70339-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jongmans MC, Kuiper RP, Carmichael CL, Wilkins EJ, Dors N, Carmagnac A, Schouten-van Meeteren AY, Li X, Stankovic M, Kamping E, et al. Novel RUNX1 mutations in familial platelet disorder with enhanced risk for acute myeloid leukemia: clues for improved identification of the FPD/AML syndrome. Leukemia. 2010;24:242–246. doi: 10.1038/leu.2009.210. [DOI] [PubMed] [Google Scholar]
- 37.Demarest RM, Ratti F, Capobianco AJ. It's T-ALL about Notch. Oncogene. 2008;27:5082–5091. doi: 10.1038/onc.2008.222. [DOI] [PubMed] [Google Scholar]
- 38.Greif PA, Tizazu B, Krause A, Kremmer E, Bohlander SK. The leukemogenic CALM/AF10 fusion protein alters the subcellular localization of the lymphoid regulator Ikaros. Oncogene. 2008;27:2886–2896. doi: 10.1038/sj.onc.1210945. [DOI] [PubMed] [Google Scholar]
- 39.Maser RS, Choudhury B, Campbell PJ, Feng B, Wong KK, Protopopov A, O'Neil J, Gutierrez A, Ivanova E, Perna I, et al. Chromosomally unstable mouse tumours have genomic alterations similar to diverse human cancers. Nature. 2007;447:966–971. doi: 10.1038/nature05886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yagi T, Hibi S, Takanashi M, Kano G, Tabata Y, Imamura T, Inaba T, Morimoto A, Todo S, Imashuku S. High frequency of Ikaros isoform 6 expression in acute myelomonocytic and monocytic leukemias: implications for up-regulation of the antiapoptotic protein Bcl-XL in leukemogenesis. Blood. 2002;99:1350–1355. doi: 10.1182/blood.v99.4.1350. [DOI] [PubMed] [Google Scholar]
- 41.Kano G, Morimoto A, Takanashi M, Hibi S, Sugimoto T, Inaba T, Yagi T, Imashuku S. Ikaros dominant negative isoform (Ik6) induces IL-3-independent survival of murine pro-B lymphocytes by activating JAK-STAT and up-regulating Bcl-xl levels. Leuk Lymphoma. 2008;49:965–973. doi: 10.1080/10428190801993462. [DOI] [PubMed] [Google Scholar]
- 42.Jäger R, Gisslinger H, Passamonti F, Rumi E, Berg T, Gisslinger B, Pietra D, Harutyunyan A, Klampfl T, Olcaydu D, et al. Deletions of the transcription factor Ikaros in myeloproliferative neoplasms. Leukemia. 2010;24:1290–1298. doi: 10.1038/leu.2010.99. [DOI] [PubMed] [Google Scholar]
- 43.Tefferi A. Novel mutations and their functional and clinical relevance in myeloproliferative neoplasms: JAK2, MPL, TET2, ASXL1, CBL, IDH and IKZF1. Leukemia. 2010;24:1128–1138. doi: 10.1038/leu.2010.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gurel Z, Ronni T, Ho S, Kuchar J, Payne KJ, Turk CW, Dovat S. Recruitment of ikaros to pericentromeric heterochromatin is regulated by phosphorylation. J Biol Chem. 2008;283:8291–8300. doi: 10.1074/jbc.M707906200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Phan-Dinh-Tuy F, Henry J, Boucheix C, Perrot JY, Rosenfeld C, Kahn A. Protein kinases in human leukemic cells. Am J Hematol. 1985;19:209–218. doi: 10.1002/ajh.2830190302. [DOI] [PubMed] [Google Scholar]
- 46.Kim JS, Eom JI, Cheong JW, Choi AJ, Lee JK, Yang WI, Min YH. Protein kinase CK2alpha as an unfavorable prognostic marker and novel therapeutic target in acute myeloid leukemia. Clin Cancer Res. 2007;13:1019–1028. doi: 10.1158/1078-0432.CCR-06-1602. [DOI] [PubMed] [Google Scholar]
- 47.Sun L, Liu A, Georgopoulos K. Zinc finger-mediated protein interactions modulate Ikaros activity, a molecular control of lymphocyte development. EMBO J. 1996;15:5358–5369. [PMC free article] [PubMed] [Google Scholar]
- 48.Klug CA, Morrison SJ, Masek M, Hahm K, Smale ST, Weissman IL. Hematopoietic stem cells and lymphoid progenitors express different Ikaros isoforms, and Ikaros is localized to heterochromatin in immature lymphocytes. Proc Natl Acad Sci U S A. 1998;95:657–662. doi: 10.1073/pnas.95.2.657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sun L, Heerema N, Crotty L, Wu X, Navara C, Vassilev A, Sensel M, Reaman GH, Uckun FM. Expression of dominant-negative and mutant isoforms of the antileukemic transcription factor Ikaros in infant acute lymphoblastic leukemia. Proc Natl Acad Sci U S A. 1999;96:680–685. doi: 10.1073/pnas.96.2.680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sun L, Crotty ML, Sensel M, Sather H, Navara C, Nachman J, Steinherz PG, Gaynon PS, Seibel N, Mao C, et al. Expression of dominant-negative Ikaros isoforms in T-cell acute lymphoblastic leukemia. Clin Cancer Res. 1999;5:2112–2120. [PubMed] [Google Scholar]
- 51.Payne KJ, Nicolas JH, Zhu JY, Barsky LW, Crooks GM. Cutting edge: predominant expression of a novel Ikaros isoform in normal human hemopoiesis. J Immunol. 2001;167:1867–1870. doi: 10.4049/jimmunol.167.4.1867. [DOI] [PubMed] [Google Scholar]
- 52.Beverly LJ, Capobianco AJ. Perturbation of Ikaros isoform selection by MLV integration is a cooperative event in Notch(IC)-induced T cell leukemogenesis. Cancer Cell. 2003;3:551–564. doi: 10.1016/s1535-6108(03)00137-5. [DOI] [PubMed] [Google Scholar]
- 53.Iacobucci I, Lonetti A, Cilloni D, Messa F, Ferrari A, Zuntini R, Ferrari S, Ottaviani E, Arruga F, Paolini S, et al. Identification of different Ikaros cDNA transcripts in Philadelphia-positive adult acute lymphoblastic leukemia by a high-throughput capillary electrophoresis sizing method. Haematologica. 2008;93:1814–1821. doi: 10.3324/haematol.13260. [DOI] [PubMed] [Google Scholar]
- 54.Payne KJ, Huang G, Sahakian E, Zhu JY, Barteneva NS, Barsky LW, Payne MA, Crooks GM. Ikaros isoform x is selectively expressed in myeloid differentiation. J Immunol. 2003;170:3091–3098. doi: 10.4049/jimmunol.170.6.3091. [DOI] [PubMed] [Google Scholar]
- 55.Morrison SJ, Wandycz AM, Hemmati HD, Wright DE, Weissman IL. Identification of a lineage of multipotent hematopoietic progenitors. Development. 1997;124:1929–1939. doi: 10.1242/dev.124.10.1929. [DOI] [PubMed] [Google Scholar]
- 56.Kondo M, Weissman IL, Akashi K. Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell. 1997;91:661–672. doi: 10.1016/s0092-8674(00)80453-5. [DOI] [PubMed] [Google Scholar]
- 57.Akashi K, Traver D, Miyamoto T, Weissman IL. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature. 2000;404:193–197. doi: 10.1038/35004599. [DOI] [PubMed] [Google Scholar]
- 58.Galy A, Travis M, Cen D, Chen B. Human T, B, natural killer, and dendritic cells arise from a common bone marrow progenitor cell subset. Immunity. 1995;3:459–473. doi: 10.1016/1074-7613(95)90175-2. [DOI] [PubMed] [Google Scholar]
- 59.Manz MG, Miyamoto T, Akashi K, Weissman IL. Prospective isolation of human clonogenic common myeloid progenitors. Proc Natl Acad Sci U S A. 2002;99:11872–11877. doi: 10.1073/pnas.172384399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Adolfsson J, Borge OJ, Bryder D, Theilgaard-Mönch K, Astrand-Grundström I, Sitnicka E, Sasaki Y, Jacobsen SE. Upregulation of Flt3 expression within the bone marrow Lin(-)Sca1(+)c-kit(+) stem cell compartment is accompanied by loss of self-renewal capacity. Immunity. 2001;15:659–669. doi: 10.1016/s1074-7613(01)00220-5. [DOI] [PubMed] [Google Scholar]
- 61.Adolfsson J, Månsson R, Buza-Vidas N, Hultquist A, Liuba K, Jensen CT, Bryder D, Yang L, Borge OJ, Thoren LA, et al. Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revised road map for adult blood lineage commitment. Cell. 2005;121:295–306. doi: 10.1016/j.cell.2005.02.013. [DOI] [PubMed] [Google Scholar]
- 62.Doulatov S, Notta F, Eppert K, Nguyen LT, Ohashi PS, Dick JE. Revised map of the human progenitor hierarchy shows the origin of macrophages and dendritic cells in early lymphoid development. Nat Immunol. 2010;11:585–593. doi: 10.1038/ni.1889. [DOI] [PubMed] [Google Scholar]
- 63.Yoshida T, Ng SY, Zuniga-Pflucker JC, Georgopoulos K. Early hematopoietic lineage restrictions directed by Ikaros. Nat Immunol. 2006;7:382–391. doi: 10.1038/ni1314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ng SY, Yoshida T, Zhang J, Georgopoulos K. Genome-wide lineage-specific transcriptional networks underscore Ikaros-dependent lymphoid priming in hematopoietic stem cells. Immunity. 2009;30:493–507. doi: 10.1016/j.immuni.2009.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]