

Abstract
The protective effect of KIOM-4, a mixture of plant extracts, was examined against streptozotocin (STZ)-induced mitochondrial oxidative stress in rat pancreatic β-cells (RINm5F). KIOM-4 scavenged superoxide and hydroxyl radicals generated by xanthine/xanthine oxidase and Fenton reaction (FeSO4/H2O2), respectively, in a cell-free chemical system. In addition, a marked increase in mitochondrial reactive oxygen species (ROS) was observed in STZ-induced diabetic cells; this increase was attenuated by KIOM-4 treatment. Mitochondrial manganese superoxide dismutase (Mn SOD) activity and protein expression were down-regulated by STZ treatment and up-regulated by KIOM-4 treatment. In addition, NF-E2 related factor 2 (Nrf2), a transcription factor for Mn SOD, was up-regulated by KIOM-4. KIOM-4 prevented STZ-induced mitochondrial lipid peroxidation, protein carbonyl and DNA modification. Moreover, KIOM-4 treatment restored the loss of mitochondrial membrane potential (Δψ) that was induced by STZ treatment, and inhibited the translocation of cytochrome c from the mitochondria to the cytosol. In addition, KIOM-4 treatment elevated the level of ATP, succinate dehydrogenase activity and insulin level, which were reduced by STZ treatment. These results suggest that KIOM-4 exhibits a protective effect through its antioxidant effect and the attenuation of mitochondrial dysfunction in STZ-induced diabetic cells.
1. Introduction
Mitochondria have gained importance in our understanding of diabetes because mitochondrial function is required for normal glucose-stimulated insulin release from pancreatic β-cells [1]. Mitochondria continuously generate superoxide radical as a byproduct of electron transport [2, 3]. The superoxide anion is quickly dismutated to hydrogen peroxide by mitochondrial manganese superoxide dismutase (Mn SOD) [4], and hydrogen peroxide is subsequently converted to water and oxygen by mitochondrial catalase and glutathione peroxidase [5]. Mitochondria not only produce reactive oxygen species (ROS) but are also the primary target of ROS attacks. Impaired mitochondrial function can lead to increased ROS generation and may increase oxidative stress if the antioxidant defense mechanisms of the cells are overwhelmed [6, 7]. Increased oxidative stress by mitochondrial dysfunction is considered a causal link between elevated glucose and the major biochemical pathways postulated to be involved in the pathogenesis of diabetes and diabetic complications [8, 9]. Streptozotocin (STZ) [N-(methyl nitro carbamoyl)-d-glucosamine] has been used to act as diabetogenic agent due to its ability to destruct pancreatic β-cells via the formation of ROS [10, 11]. It has been reported that production of mitochondrial ROS increased in STZ-treated rat, and mitochondrial lipid peroxidation was observed in pancreatic tissue, suggesting STZ-induced mitochondrial oxidative stress [12]. Thus, antioxidant therapy may be a promising therapeutic approach for controlling diabetes or diabetic complications.
KIOM-4 is a combination of extracts obtained from Magnolia officinalis, Pueraria lobata, Glycyrrhiza uralensis and Euphorbia pekinensis. M. officinalis exhibits antimutagenic, hepato-protective, neuro-protective, antiinflammatory and antimicrobial activities [13–18]. P. lobata has antimutagenic, antidiabetic and antioxidant effects [19–21]. G. uralensis has been documented as having detoxification, antioxidant, antiulcer, antiinflammatory, antiviral, antiatherogenic, anticarcinogenic effects and cytoprotective effect of hepatocyte against hepatotoxicity [22–24]. E. pekinensis has antiviral and cytotoxic activities [25, 26]. We recently demonstrated that KIOM-4 exhibits cytoprotective effects against STZ-induced oxidative stress damage in β-cells via the activation of catalase and heme oxygenase-1 [27, 28].
This study was undertaken to investigate the protective effect of KIOM-4 and its mechanism against STZ-induced oxidative mitochondrial damage in pancreatic β-cells.
2. Methods
2.1. Preparation of KIOM-4
The cortex of M. officinalis, and radixes of P. lobata, G. uralensis and E. pekinensis were collected from the Gamsuk province in China, and identified by Prof. J.H. Kim of the Division of Life Science, Daejeon University, Korea. All voucher specimens were deposited at the herbarium of the Department of Herbal Pharmaceutical Development, Korea Institute of Oriental Medicine (No. 1240, 2, 7 and 207, resp.). An equal amount of Magnoliae cortex, and radixes of Puerariae, Glycyrrhizae and Euphoriae was mixed, pulverized and extracted in 80% ethanol for one week at room temperature, concentrated using a rotary evaporator and lyophilized. The entire procedure was repeated four times. KIOM-4 was dissolved in dimethyl sulfoxide (DMSO), the final concentration of which did not exceed 0.1%.
2.2. Reagents
STZ was purchased from Calbiochem (San Diego, CA). Dihydrorhodamin 123 (DHR 123) and JC-1 (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-benzimidazolcarbocyanine iodide) were purchased from Molecular Probes (Eugene, OR). Cytochrome c (H-104), Nrf2 (C-20) antibodies were purchased from the Santa Cruz Biotechnology (Santa Cruz, CA). The Mn SOD polyclonal antibody was purchased from the Stressgen Corporation (Victoria, Canada). 5, 5-dimethyl-1-pyrroline-N-oxide (DMPO) and [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium] bromide (MTT) were purchased from the Sigma Chemical Company (St. Louis, MO).
2.3. Cell Culture and Treatments
RINm5F rat pancreatic β-cells were maintained at 37°C in an incubator with a humidified atmosphere of 5% CO2, and cultured in RPMI-1640 medium containing 10% heat-inactivated fetal calf serum, streptomycin (100 μg ml−1) and penicillin (100 units ml−1). Cells were seeded on to a culture plate at a concentration of 1 × 105 cells ml−1, and at 16 h after plating were treated with KIOM-4 at 50 μg ml−1. After 1 h, 10 mM STZ was added to the plate.
2.4. Detection of Superoxide Radical
Superoxide radicals were produced by reaction of the xanthine/xanthine oxidase system and reacted with spin trap DMPO. The DMPO–·OOH adducts were detected using electron spin resonance (ESR) spectroscopy [29]. The ESR spectrum was recorded 2.5 min after mixing in a phosphate buffer solution (pH 7.4) with 6 M DMPO 20 μl, xanthine oxidase (0.25 U ml−1) 20 μl, xanthine (5 mM) 20 μl and KIOM-4 (final 50 μg ml−1) 20 μl using JES-FA ESR spectrometer (JEOL, Tokyo, Japan).
2.5. Detection of Hydroxyl Radical
Hydroxyl radicals were generated by Fenton reaction. Hydroxyl radicals reacted with DMPO and the resultant DMPO–OH adducts were detected using an ESR spectrometer [29]. The ESR spectrum was recorded 2.5 min after mixing in a phosphate buffer solution (pH 7.4) with 0.2 ml of 0.3 M DMPO, 0.2 ml of 10 mM FeSO4, 0.2 ml of 10 mM H2O2 and KIOM-4 using ESR spectrometer.
2.6. Mitochondrial ROS Measurement
The RINm5F cells were seeded on to a 96-well plate at 2 × 104 cells/well. At 16 h after plating, the cells were treated with KIOM-4 at 50 μg ml−1, and 1 h later 10 mM STZ was added to the plate. The cells were incubated for an additional 30 min at 37°C. After addition of 20 μM of DHR 123 solution for 10 min, the fluorescence was detected using a Perkin Elmer LS-5B spectrofluorometer and flow cytometer (Becton Dickinson, Mountain View, CA). For image analysis of the generation of mitochondrial ROS, the cells were seeded on a cover-slip loaded six-well plate at 2 × 105 cells/well. At 16 h after plating, the cells were treated with KIOM-4, and 1 h later 10 mM STZ was added to the plate. After changing the media, 20 μM of DHR 123 was added to each well and the plate was incubated for an additional 30 min at 37°C. After washing with PBS, the stained cells were mounted onto a microscope slide in mounting medium (DAKO, Carpinteria, CA). Images were collected using the Laser Scanning Microscope 5 PASCAL program (Carl Zeiss, Jena, Germany) on a confocal microscope.
2.7. Cellular Mitochondrial Fractionation
Mitochondrial fractions were isolated by differential centrifugation using the mitochondria isolation kit (Active-Motif, Carlsbad, CA).
2.8. Measurement of Mn SOD Activity
The RINm5F cells were seeded on to a culture dish at a concentration of 1 × 105 cells ml−1, and at 16 h after plating were treated with KIOM-4 at 50 μg ml−1. After 1 h, 10 mM STZ was added to the plate, which was incubated for a further 24 h. The harvested cells were suspended in 10 mM phosphate buffer (pH 7.5) and then lysed on ice by sonicating twice for 15 s. Triton X-100 (1%) was then added to the lysates and incubated for 10 min on ice. The lysates were clarified, by centrifugation at 5000 g for 30 min at 4°C, and the protein content of the supernatant was determined. Fifty micrograms of protein was added to 500 mM phosphate buffer (pH 10.2), 1 mM potassium cyanide (inhibitor of Cu Zn SOD) and 1 mM epinephrine. Epinephrine rapidly undergoes auto-oxidation at pH 10 to produce adrenochrome, a pink-colored product, which was assayed at 480 nm using a UV/VIS spectrophotometer in the kinetic mode. Mn SOD inhibits the auto-oxidation of epinephrine. The rate of inhibition was monitored at 480 nm and the amount of enzyme required to produce 50% inhibition was defined as one unit of enzyme activity, and the Mn SOD activity was expressed as units/mg protein [30].
2.9. Western Blot
Aliquots of the lysates (40 μg of protein) were boiled for 5 min and electrophoresed on a 10% SDS-polyacrylamide gel. Blots in the gels were transferred onto nitrocellulose membranes (Bio-Rad, Hercules, CA). The nitrocellulose membrane was incubated with primary antibodies (1 : 1000) at 4°C overnight. Secondary antibody horseradish peroxidase conjugates (1 : 5000) (Pierce, Rockland, IL) were added, and incubated at room temperature for 1 h, and exposed to X-ray film. Protein bands were detected using an enhanced chemiluminescence western blotting detection kit (Amersham, Little Chalfont, Buckinghamshire, UK).
2.10. Lipid Peroxidation Assay
Lipid peroxidation was assayed by determination of 8-isoprostane levels [31] in the culture medium, by use of a commercial enzyme immunoassay kit (Cayman Chemical, Ann Arbor, MI) according to the manufacturer's instructions.
2.11. Protein Carbonyl Formation
The amount of protein carbonyl formation was determined using an Oxiselect protein carbonyl ELISA kit purchased from Cell Biolabs (San Diego, CA) according to the manufacturer's instructions.
2.12. 8-Hydroxyl-2′-deoxyguanosine Assay
The amount of 8-hydroxyl-2′-deoxyguanosine (8-OHdG) was determined using a Bioxytech 8-OHdG-ELISA Kit purchased from OXIS Health Products (Portland, OR) according to the manufacturer's instructions.
2.13. Mitochondrial Membrane Potential Analysis
Mitochondrial membrane potential (Δψ) was analyzed using JC-1, a lipophilic cationic fluorescence dye. Cells were harvested, and after changing the media, JC-1 was added to each well and incubated for an additional 30 min at 37°C. After washing with PBS, the stained cells were assayed using flow cytometer. For image analysis of mitochondrial Δψ, the stained cells were mounted onto microscope slide in mounting medium. Microscopic images were collected using the Laser Scanning Microscope 5 PASCAL program on confocal microscope [32].
2.14. Quantification of Cellular ATP Levels
The mitochondrial function was evaluated by measuring the cellular adenosine triphosphate (ATP) production in cells. The cells were harvested and washed twice with PBS. The harvested cells were then lysed on ice for 30 min in 200 μl of lysis buffer [25 mM Tris (pH 7.8), 270 mM sucrose, 1 mM EDTA] by sonicating three times for 15 s and centrifuged at 4°C for 10 min at 16 000 g. Supernatants were collected from the lysates and ATP content was assayed using a luciferase/luciferin ATP determination kit (Molecular Probes, Eugene, OR) [33].
2.15. Mitochondrial Succinate Dehydrogenase Activity
To evaluate mitochondrial metabolic activity, mitochondrial succinate dehydrogenase activity was estimated by the MTT assay [34]. The cells were treated with KIOM-4 at 50 μg ml−1. After 1 h, 10 mM of STZ, and the mixture was incubated for 24 h. Fifty microliters of the [3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium] bromide (MTT) stock solution (2 mg ml−1) was then added into each well to attain a total reaction volume of 200μl. After incubating for 4 h, the plate was centrifuged at 800 g for 5 min and the supernatants were aspirated. The formazan crystals in each well were dissolved in 150 μl of dimethylsulfoxide and read at A540 on a scanning multi-well spectrophotometer [35].
2.16. Measurements of Insulin Levels
The cells were treated with KIOM-4 at 50 μg ml−1, and 1 h later 10 mM STZ was added to the plate, and the mixture was incubated for 24 h. The amount of insulin was determined by using an ELISA rat specific insulin enzyme immunoassay kit (Spi-Bio, Massy, France).
2.17. Statistical Analysis
All the measurements were made in triplicate and all values are represented as the mean ± standard error of the mean (SEM). Data were subjected to an analysis of the variance (ANOVA) using the Tukey test to analyze the difference. A P-value of <.05 was considered significant.
3. Results
3.1. Radical Scavenging Activity of KIOM-4 in a Cell-Free System
Our previous result showed that the intracellular ROS scavenging activity of KIOM-4 was 48, 55 and 66% at concentrations of 10, 50 and 100 μg ml−1, respectively [24]. And KIOM-4 at concentrations of 100 μg ml−1 showed some cytotoxicity (data not shown). Therefore, we determined to choose 50 μg ml−1 as optimal dose for further study. The radical scavenging effects of KIOM-4 on superoxide radicals and hydroxyl radicals were measured. Superoxide radicals produced by the xanthine/xanthine oxidase system and the hydroxyl radicals generated by the Fenton reaction (FeSO4 + H2O2) in a cell-free system were detected by ESR spectrometry. The ESR results revealed no clear signal in the control and the 50 μg ml−1 of KIOM-4; however, the superoxide radical signal increased to 646 in the xanthine/xanthine oxidase system. KIOM-4 treatment decreased the superoxide radical signal to 209 (Figure 1(a)). In addition, the hydroxyl radical signal increased to 2809 in the FeSO4 + H2O2 system. KIOM-4 treatment decreased the hydroxyl radical signal to 386 (Figure 1(b)).
Figure 1.
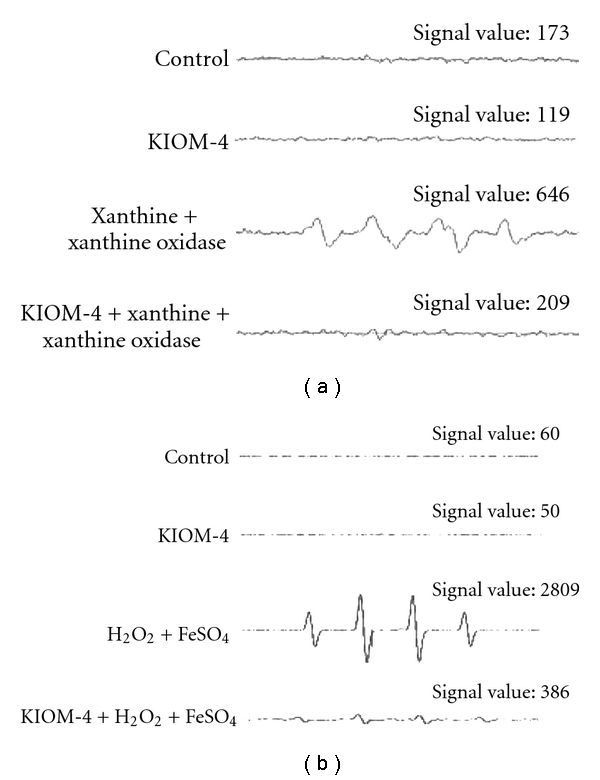
The effect of KIOM-4 on cell-free generation of superoxide and hydroxyl radicals. (a) Superoxide radicals were generated by xanthine/xanthine oxidase and reacted with DMPO; the resultant DMPO-OOH adducts were detected using an ESR spectrometer. (b) Hydroxyl radicals were generated by the Fenton reaction (H2O2/FeSO4) and reacted with DMPO; the resultant DMPO-OH adducts were detected using an ESR spectrometer.
3.2. Reduction of Mitochondrial ROS by KIOM-4 Treatment
The fluorescence dye DHR 123 was used to detect mitochondrial ROS in cells after STZ treatment. The fluorescence spectrometric data revealed that STZ treatment increased the level of mitochondrial ROS compared with control. However, treatment with KIOM-4 at 50 μg ml−1 attenuated the STZ-induced ROS increase (Figure 2(a)). In addition, flow cytometry revealed a fluorescence intensity of 106 for ROS in STZ-treated cells with KIOM-4 at 50 μg ml−1, compared with a fluorescence intensity of 533 in STZ-treated cells (Figure 2(b)). Confocal microscopy revealed that KIOM-4 reduced the red fluorescence intensity of STZ-induced mitochondrial ROS (Figure 2(c)). These data suggest that KIOM-4 had mitochondrial ROS scavenging properties.
Figure 2.
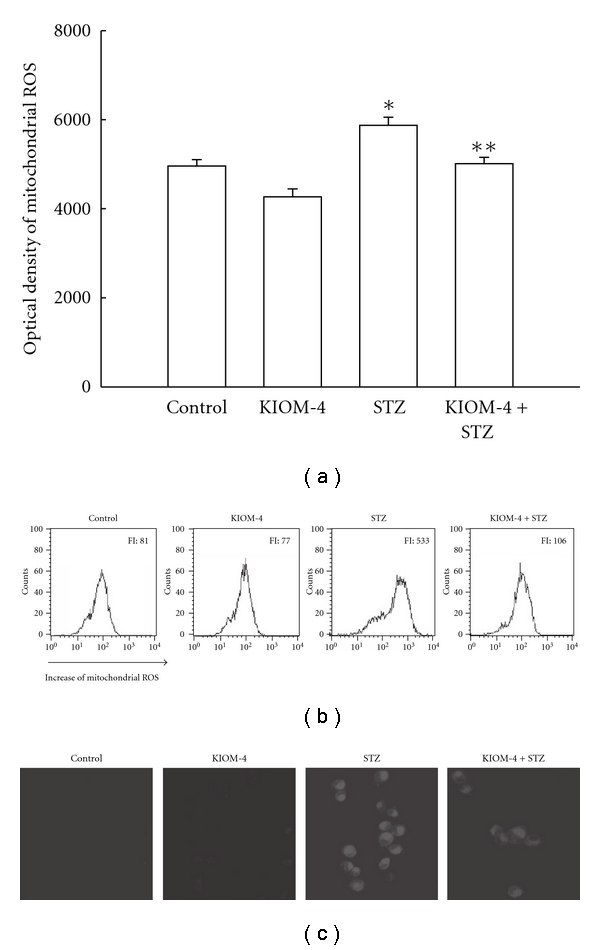
The effect of KIOM-4 on STZ-induced mitochondrial ROS generation. The cells were treated with KIOM-4 at 50 μg ml−1. After 1 h, 10 mM of STZ was added to the plate. After an additional 30 min, the mitochondrial ROS were detected by spectrofluorometry (a) and flow cytometry (b) after DHR 123 treatment. FI indicates the fluorescence intensity of DHR 123. (c) The representative confocal images illustrate the increase in red fluorescence intensity of DHR 123 produced by ROS in STZ-treated cells s compared with that in control and the lowered fluorescence intensity in STZ-treated cells with KIOM-4 (original magnification ×400). The measurements were made in triplicate and the values were expressed as means ± SEM. Asterisk represents significantly different from control cells (P < .05) and double asterisk represent significantly different from STZ-treated cells (P < .05).
3.3. Induction of Mn SOD and Its Transcription Factor by KIOM-4 Treatment
Mn SOD acts as a first defense system to protect mitochondria and other cellular components as it scavenges superoxide anion in the mitochondrial matrix [36]. As shown in Figure 3(a), Mn SOD activity was 30 U mg−1 protein with 50 μg ml−1 of KIOM-4, compared with 26 U mg−1 protein in the control. STZ treatment decreased the SOD activity to 12 U mg−1 protein; however, treatment with KIOM-4 increased this activity to 20 U mg−1 protein. In addition, western blot data revealed that STZ treatment decreased the expression of Mn SOD compared with control. KIOM-4 treatment at 50 μg ml−1 increased the Mn SOD level attenuated by STZ treatment (Figure 3(b)). Mn SOD has an antioxidant response element (ARE) sequence in its promoter region. Nrf2 is an important transcription factor that regulates ARE-driven Mn SOD expression. Nuclear Nrf2 expression was decreased by STZ treatment; however, KIOM-4 treatment cells increased the nuclear Nrf2 expression (Figure 3(c)). These data suggest that KIOM-4 exhibited induction of Mn SOD via activation of Nrf2.
Figure 3.
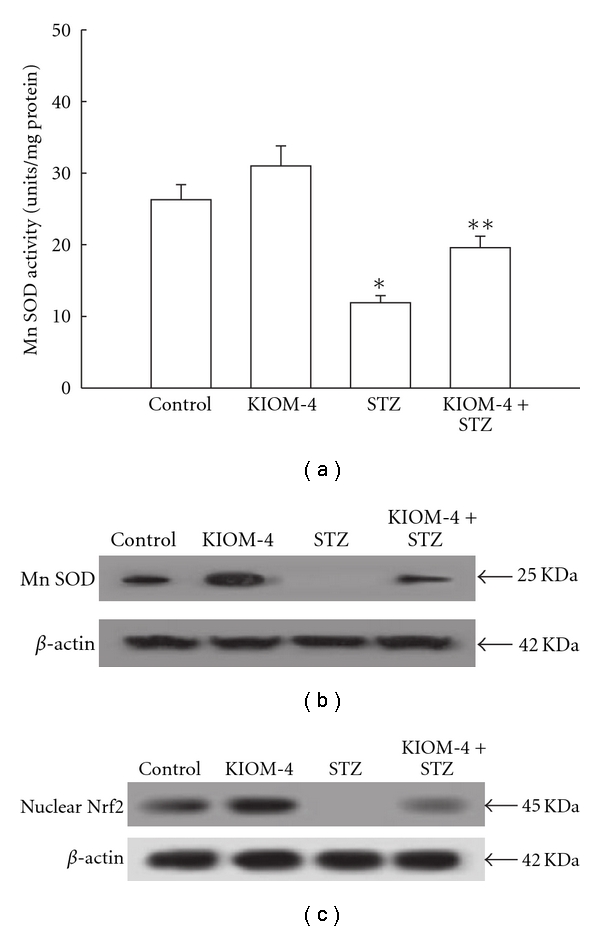
The effect of KIOM-4 on Mn SOD and its transcription factor. (a) The enzyme activities are expressed as average enzyme unit per mg protein ± SEM. Western blot analysis was performed using anti-Mn SOD (b) and Nrf2 (c) antibody. Asterisk represent significantly different from control cells (P < .05) and double asterisk represent significantly different from STZ-treated cells (P < .05).
3.4. Protection of Damaged Mitochondrial Components by KIOM-4 Treatment
The level of 8-isoprostan, a marker of lipid peroxidation, is increased to 230 pg ml−1 in cells exposed to STZ, compared with 159 pg ml−1 in control cells. KIOM-4, however, decreased this level to 161 pg ml−1 in STZ-treated cells (Figure 4(a)). The mitochondrial protein carbonyl content, which is marker of protein modification [37], increased significantly after STZ treatment, and KIOM-4 prevented the STZ-induced protein carbonyl formation (Figure 4(b)). STZ treatment increased the amount of 8-OHdG, which is marker of base modification in DNA [38], to 2006 pg ml−1 compared with 247 pg ml−1 in control cells, and KIOM-4 decreased 8-OHdG to 777 pg ml−1 in STZ-treated cells (Figure 4(c)). These data suggest that KIOM-4 provides protection against STZ-induced mitochondrial damages.
Figure 4.
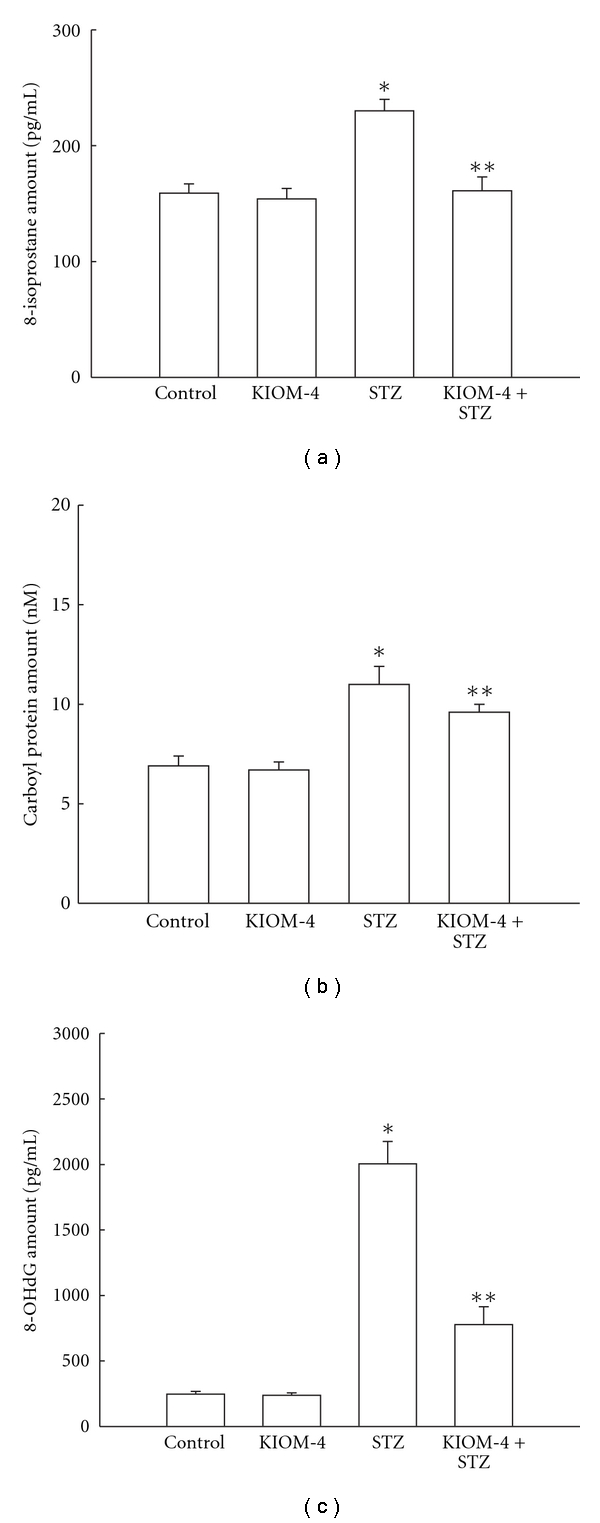
The effect of KIOM-4 on STZ-induced mitochondrial lipid, protein and DNA damage. (a) Lipid peroxidation was detected by measuring the amount of 8-isoprostane. (b) Protein oxidation was assayed by measuring carbonyl formation. (c) DNA damage was detected measuring the amount of 8-OHdG. Asterisk represent significantly different from control cells (P < .05) and double asterisk represent significantly different from STZ-treated cells (P < .05).
3.5. Recovery of Disrupted Mitochondrial Δψ and Its Related Protein by KIOM-4 Treatment
The mitochondrial Δψ, which is a marker of mitochondrial membrane integrity, was detected using flow cytometry and confocal microscopy after staining with the fluorescence dye JC-1. The flow cytometric data showed that the STZ treatment resulted in the loss of Δψ, as substantiated by an increase in fluorescence (FL-1) with JC-1. KIOM-4 treatment blocked the loss of Δψ in STZ-treated cells. The fluorescence intensity was 301 value in STZ-treated cells with 50 μg ml−1 of KIOM-4, compared with a fluorescence intensity of 586 in STZ-treated cells (Figure 5(a)). In addition, the confocal microscopy data showed that control cells and cells treated with KIOM-4 only exhibited strong red fluorescence in the mitochondria after JC-1 staining, indicating that mitochondrial Δψ was in the polarized state (Figure 5(b), left panel). However, STZ treatment resulted in decreased red fluorescence in the mitochondria and increased green fluorescence, suggesting that STZ treatment disrupted the mitochondrial Δψ to a depolarized state. KIOM-4 treatment decreased the green fluorescence in STZ-treated cells (Figure 5(b), right panel), indicating that KIOM-4 inhibited the loss of Δψ in response to STZ treatment. The pore opening induces the loss of Δψ, which in turn induces the release of cytochrome c from the mitochondria and most commonly leads to apoptotic cell death [39]. KIOM-4 inhibited the STZ-induced release of cytochrome c from the mitochondria into the cytosol (Figure 5(c)). These results suggest that KIOM-4 protected against STZ-damage to mitochondrial Δψ.
Figure 5.
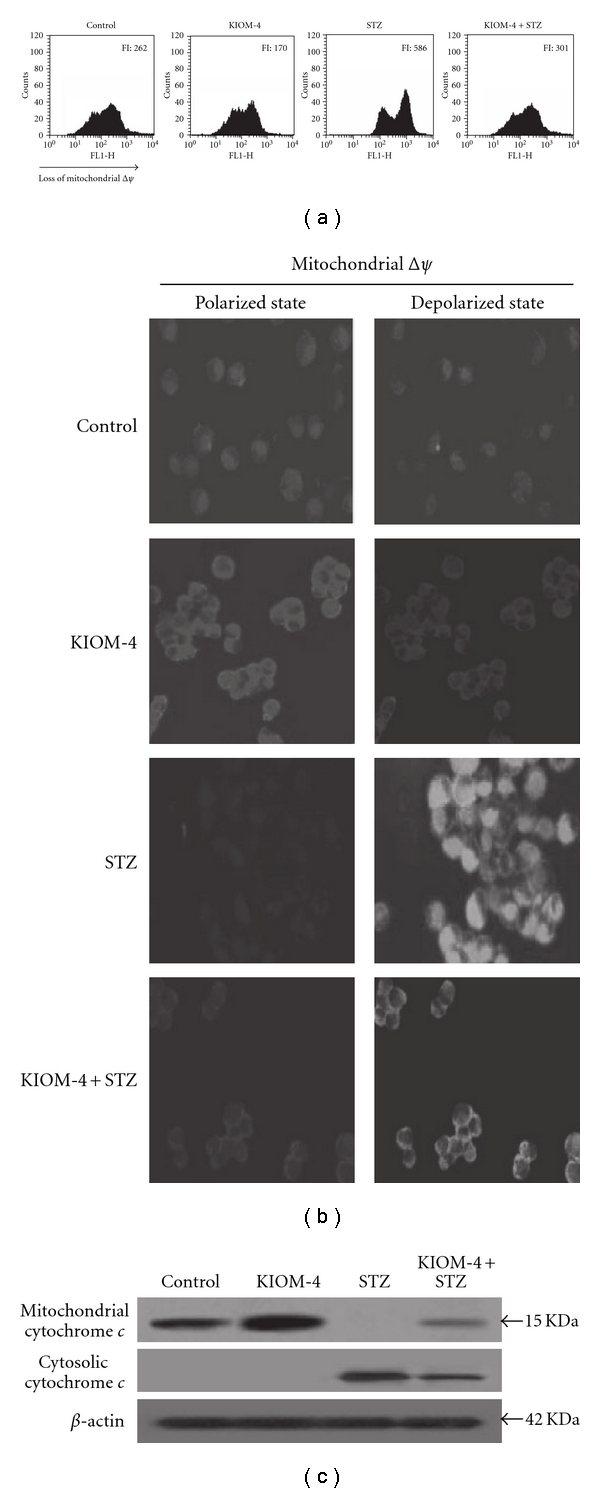
The effect of KIOM-4 on mitochondrial Δψ and its related proteins. The mitochondrial Δψ was analyzed using (a) flow cytometry and (b) confocal microscopy after staining cells with JC-1 dye. Western blot analysis was performed using anti-cytochrome c (c) antibody.
3.6. Enhancement of Decreased Intracellular ATP Level, Mitochondrial Enzymes, and Insulin Secretion by KIOM-4 Treatment
Mitochondrial injury is followed by the depletion of intracellular ATP and mitochondrial enzymes. As shown in Figure 6(a), STZ treatment reduced the ATP level compared with that in control cells; however, KIOM-4 treatment recovered the ATP level in STZ-treated cells. Mitochondrial succinate dehydrogenase activity was decreased in STZ-treated cells; however, KIOM-4 treatment of STZ-treated cells enhanced this activity (Figure 6(b)). Furthermore, STZ decreased the insulin level of RINm5F, which secretes insulin, however, KIOM-4 treatment of STZ-treated cells enhanced insulin secretion (Figure 6(c)). These results suggest that KIOM-4 attenuates mitochondrial dysfunction in STZ-induced diabetic cells.
Figure 6.
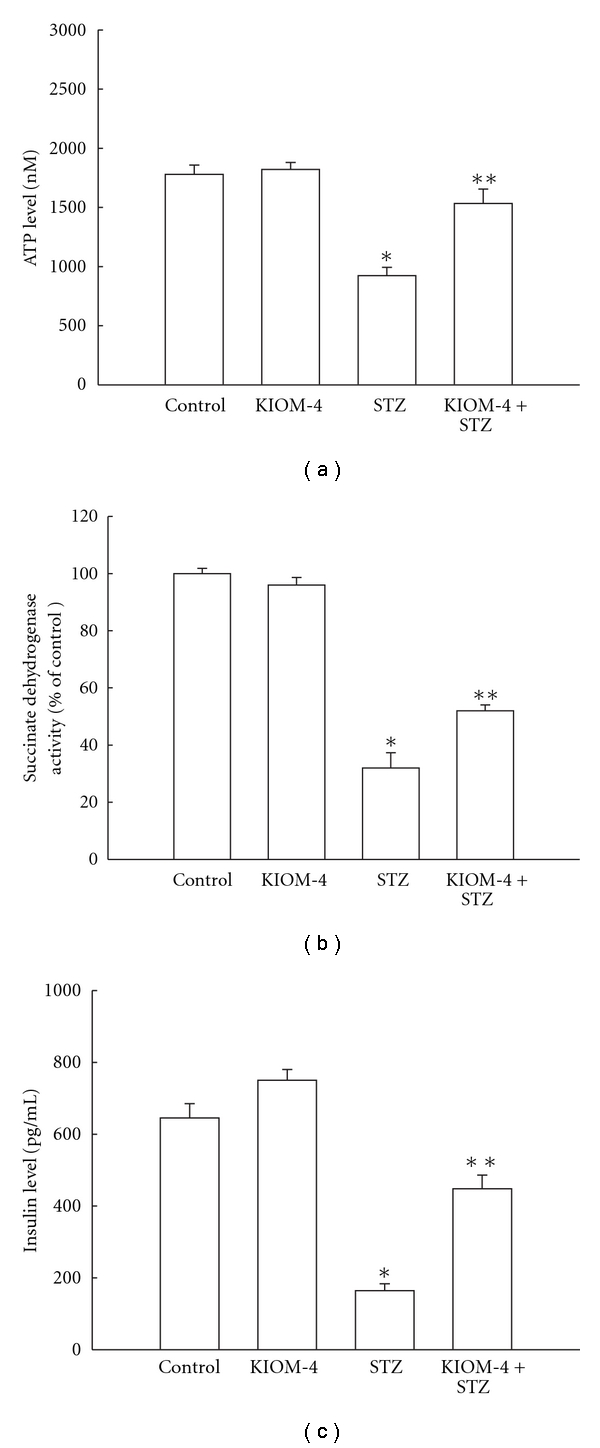
The effect of KIOM-4 on intracellular ATP level, mitochondrial enzyme, and insulin level. (a) ATP content was assayed using a luciferase/luciferin ATP determination kit. (b) Mitochondrial succinate dehydrogenase activity was estimated by the MTT assay. (c) The amount of insulin was determined by using an ELISA rat specific insulin enzyme immunoassay kit. Asterisk represent significantly different from control cells (P < .05) and double asterisk represent significantly different from STZ-treated cells (P < .05).
4. Discussion
Mitochondrial radical production and consequent oxidative damage contribute to the progressive and patho-physiological conditions of diabetes. It has been suggested that mitochondrial ROS induced by high glucose might cause the pathogenesis of diabetes mellitus and its complications through modification of various mitochondrial events [40]. Consequently, therapeutic strategies to decrease ROS production or to intercept these ROS should be explored [41]. Numerous studies have reported that antioxidant treatment, which targets oxidative stress, may help prevent or delay the development of diabetes and its complications [42, 43]. In this context, the possible anti-diabetic effects of KIOM-4 on the oxidative mitochondrial damage induced by STZ treatment in pancreatic β-cells were elucidated. In this study, KIOM-4 exhibited significant ROS radical scavenging activity against superoxide and hydroxyl radicals. Moreover, KIOM-4 attenuated the STZ-induced increase in mitochondrial ROS. Mn SOD is located in the mitochondrial matrix and is the primary SOD isoform that dismutates superoxide anions generated by the mitochondrial respiratory chain [44]. Pancreatic cells contain relatively low levels of antioxidant enzymes, making these cells more vulnerable to oxidative stress [42, 45]. A significant imbalance between ROS production and endogenous Mn SOD has been confirmed by the reduced activity and protein expression of Mn SOD in STZ-treated cells. In diabetes, decreased Mn SOD activity promotes the damage to cellular components such as lipid, protein and DNA [46]. Treatment with KIOM-4 restored the Mn SOD activity decreased by STZ. In addition, our previous study suggested that KIOM-4 exhibited cytoprotective effects against STZ-induced oxidative stress damage in β-cells via the activation of catalase and heme oxygenase-1 [27, 28]. Meanwhile, Nrf2 is able to activate the antioxidant-responsive element (ARE)-dependent gene expression in order to maintain cellular redox homeostasis [47]. Nrf2 is an important transcription factor that regulates ARE-driven expression of antioxidant genes, including Mn SOD. Nuclear Nrf2 expression was decreased by STZ treatment; however, KIOM-4 treatment recovered the nuclear Nrf2 expression. The components in the mitochondrial matrix are highly susceptible to an oxidative environment [48]. KIOM-4 prevented the STZ-induced mitochondrial lipid peroxidation, protein carbonyl and DNA modification. In general, mitochondrial DNA is more vulnerable to oxidative stress, and the subsequent damage is more extensive than that in nuclear DNA due to the lack of protective histones and low repair mechanisms [45, 46]. Elevated oxidative stress can increase the membrane permeability of the mitochondria by opening pores in the inner mitochondrial membrane, and can lead to the loss of Δψ [49]. The loss of mitochondrial Δψ can cause the release of cytochrome c and the activation of the apoptotic pathway [50]. Indeed, STZ treatment disrupted the mitochondrial Δψ and resulted in the release of mitochondrial cytochrome c to cytosol. However, KIOM-4 prevented the sequential mitochondrial damage process induced by STZ treatment. The elevated level of mitochondrial ROS generated by STZ acts as a powerful oxidant and causes damage to mitochondrial respiratory chain complexes. As mitochondria are the major producers of ATP, mitochondrial dysfunction also leads to reduced ATP levels [51]. Depletion of ATP content was observed in STZ-treated cells as a consequence of impaired mitochondrial respiratory chain activity. Succinate dehydrogenase is a mitochondrial TCA cycle enzyme and its activity is mainly regulated by ATP [52]. KIOM-4 treatment restored the ATP content and the succinate dehydrogenase activity, which were both reduced by STZ treatment. It has been reported that STZ treatment can increase mitochondrial ROS in rat pancreatic tissue, and the mitochondrial oxidative stress reduces insulin secretion by pancreatic β-cells [12, 53]. KIOM-4 significantly increased insulin content decreased by STZ treatment.
Hence, these results suggest that KIOM-4 protects against ROS-mediated mitochondrial dysfunction in diabetic pancreatic β-cells by scavenging ROS and inducing mitochondrial antioxidant enzymes (Figure 7).
Figure 7.
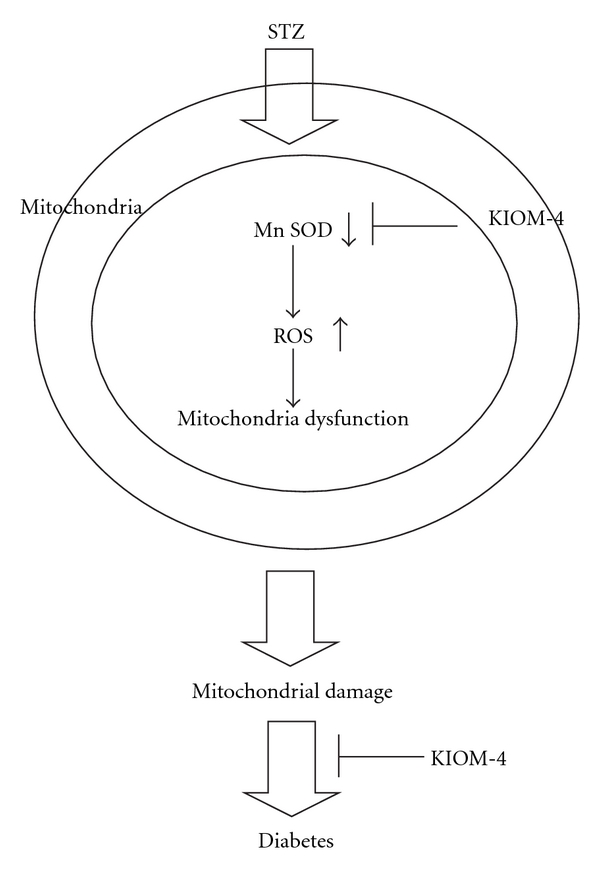
A proposed pathway for protective effect of KIOM-4 against STZ-induced mitochondrial oxidative stress in pancreatic β-cells via its antioxidant effects.
Funding
Korea Institute of Oriental Medicine (grant no. K09030).
Acknowledgment
K. A. Kang and J. S. Kim contribute equally to this work.
References
- 1.Wallace DC. Mitochondrial diseases in man and mouse. Science. 1999;283(5407):1482–1488. doi: 10.1126/science.283.5407.1482. [DOI] [PubMed] [Google Scholar]
- 2.Kristal BS, Jackson CT, Chung H-Y, Matsuda M, Nguyen HD, Yu BP. Defects at center P underlie diabetes-associated mitochondrial dysfunction. Free Radical Biology and Medicine. 1997;22(5):823–833. doi: 10.1016/s0891-5849(96)00428-5. [DOI] [PubMed] [Google Scholar]
- 3.Nishikawa T, Edelstein D, Du XL, et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycaemic damage. Nature. 2000;404(6779):787–790. doi: 10.1038/35008121. [DOI] [PubMed] [Google Scholar]
- 4.Moro MA, Almeida A, Bolaños JP, Lizasoain I. Mitochondrial respiratory chain and free radical generation in stroke. Free Radical Biology and Medicine. 2005;39(10):1291–1304. doi: 10.1016/j.freeradbiomed.2005.07.010. [DOI] [PubMed] [Google Scholar]
- 5.Singh P, Jain A, Kaur G. Impact of hypoglycemia and diabetes on CNS: correlation of mitochondrial oxidative stress with DNA damage. Molecular and Cellular Biochemistry. 2004;260(1):153–159. doi: 10.1023/b:mcbi.0000026067.08356.13. [DOI] [PubMed] [Google Scholar]
- 6.Ježek P, Hlavatá L. Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism. International Journal of Biochemistry and Cell Biology. 2005;37(12):2478–2503. doi: 10.1016/j.biocel.2005.05.013. [DOI] [PubMed] [Google Scholar]
- 7.Barker DJP, Osmond C. Infant mortality, childhood nutrition, and ischaemic heart disease in England and Wales. The Lancet. 1986;1(8489):1077–1081. doi: 10.1016/s0140-6736(86)91340-1. [DOI] [PubMed] [Google Scholar]
- 8.Nishikawa T, Edelstein D, Brownlee M. The missing link: a single unifying mechanism for diabetic complications. Kidney International. 2000;58(77):S26–S30. doi: 10.1046/j.1523-1755.2000.07705.x. [DOI] [PubMed] [Google Scholar]
- 9.Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414(6865):813–820. doi: 10.1038/414813a. [DOI] [PubMed] [Google Scholar]
- 10.Ohkuwa T, Sato Y, Naoi M. Hydroxyl radical formation in diabetic rats induced by streptozotocin. Life Sciences. 1995;56(21):1789–1798. doi: 10.1016/0024-3205(95)00150-5. [DOI] [PubMed] [Google Scholar]
- 11.Chen X, Touyz RM, Park JB, Schiffrin EL. Antioxidant effects of vitamins C and E are associated with altered activation of vascular NADPH oxidase and superoxide dismutase in stroke-prone SHR. Hypertension. 2001;38(3):606–611. doi: 10.1161/hy09t1.094005. [DOI] [PubMed] [Google Scholar]
- 12.Raza H, Prabu SK, Robin M-A, Avadhani NG. Elevated mitochondrial cytochrome P450 2E1 and glutathione S-transferase A4-4 in streptozotocin-induced diabetic rats tissue-specific variations and roles in oxidative stress. Diabetes. 2004;53(1):185–194. doi: 10.2337/diabetes.53.1.185. [DOI] [PubMed] [Google Scholar]
- 13.Saito J, Sakai Y, Nagase H. In vitro anti-mutagenic effect of magnolol against direct and indirect mutagens. Mutation Research. 2006;609(1):68–73. doi: 10.1016/j.mrgentox.2006.06.021. [DOI] [PubMed] [Google Scholar]
- 14.Park E-J, Kim S-Y, Zhao Y-Z, Sohn DH. Honokiol reduces oxidative stress, c-jun-NH2-terminal kinase phosphorylation and protects against glycochenodeoxycholic acid-induced apoptosis in primary cultured rat hepatocytes. Planta Medica. 2006;72(7):661–664. doi: 10.1055/s-2006-931571. [DOI] [PubMed] [Google Scholar]
- 15.Cao AH, Vo LT, King RG. Honokiol protects against carbon tetrachloride induced liver damage in the rat. Phytotherapy Research. 2005;19(11):932–937. doi: 10.1002/ptr.1757. [DOI] [PubMed] [Google Scholar]
- 16.Lin Y-R, Chen H-H, Ko C-H, Chan M-H. Neuroprotective activity of honokiol and magnolol in cerebellar granule cell damage. European Journal of Pharmacology. 2006;537(1–3):64–69. doi: 10.1016/j.ejphar.2006.03.035. [DOI] [PubMed] [Google Scholar]
- 17.Lee J, Jung E, Park J, et al. Anti-inflammatory effects of magnolol and honokiol are mediated through inhibition of the downstream pathway of MEKK-1 in NF-κB activation signaling. Planta Medica. 2005;71(4):338–343. doi: 10.1055/s-2005-864100. [DOI] [PubMed] [Google Scholar]
- 18.Ho KY, Tsai CC, Chen CP, Huang JS, Lin CC. Antimicrobial activity of honokiol and magnolol isolated from Magnolia officinalis. Phytotherapy Research. 2001;15(2):139–141. doi: 10.1002/ptr.736. [DOI] [PubMed] [Google Scholar]
- 19.Miyazawa M, Sakano K, Nakamura S-I, Kosaka H. Antimutagenic activity of isoflavone from Pueraria lobata . Journal of Agricultural and Food Chemistry. 2001;49(1):336–341. doi: 10.1021/jf000255w. [DOI] [PubMed] [Google Scholar]
- 20.Lee KT, Sohn IC, Kim DH, Choi JW, Kwon SH, Park HJ. Hypoglycemic and hypolipidemic effects of tectorigenin and kaikasaponin III in the streptozotocin-lnduced diabetic rat and their antioxidant activity in vitro . Archives of Pharmacal Research. 2000;23(5):461–466. doi: 10.1007/BF02976573. [DOI] [PubMed] [Google Scholar]
- 21.Lee K-T, Sohn I-C, Kim Y-K, et al. Tectorigenin, an isoflavone of Pueraria thunbergiana BENTH., induces differentiation and apoptosis in human promyelocytic leukemia HL-60 cells. Biological and Pharmaceutical Bulletin. 2001;24(10):1117–1121. doi: 10.1248/bpb.24.1117. [DOI] [PubMed] [Google Scholar]
- 22.Liao H, Banbury LK, Leach DN. Elucidation of Danzhixiaoyao Wan and its constituent herbs on antioxidant activity and inhibition of nitric oxide production. Evidence-Based Complementary and Alternative Medicine. 2007;4(4):425–430. doi: 10.1093/ecam/nel091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang ZY, Nixon DW. Licorice and cancer. Nutrition and Cancer. 2001;39(1):1–11. doi: 10.1207/S15327914nc391_1. [DOI] [PubMed] [Google Scholar]
- 24.Popovich DG, Yeo SY, Zhang W. Ginseng (Panax quinquefolius) and Licorice (Glycyrrhiza uralensis) root extract combinations increase hepatocarcinoma cell (Hep-G2) viability. doi: 10.1093/ecam/nep074. Evidence-Based Complementary and Alternative Medicine. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ahn MJ, Kim CY, Lee JS, et al. Inhibition of HIV-1 integrase by galloyl glucoses from Terminalia chebula and flavonol glycoside gallates from Euphorbia pekinensis . Planta Medica. 2002;68(5):457–459. doi: 10.1055/s-2002-32070. [DOI] [PubMed] [Google Scholar]
- 26.Kong L-Y, Li Y, Wu X-L, Min Z-D. Cytotoxic diterpenoids from Euphorbia pekinensis . Planta Medica. 2002;68(3):249–252. doi: 10.1055/s-2002-23132. [DOI] [PubMed] [Google Scholar]
- 27.Kang KA, Kim JS, Hyun JW. KIOM-4 protects RINm5F pancreatic β-cells against streptozotocin induced oxidative stress in vitro . Biotechnology and Bioprocess Engineering. 2008;13(2):150–157. [Google Scholar]
- 28.Kang KA, Kim JS, Zhang R, et al. Protective mechanism of KIOM-4 against streptozotocin induced diabetic cells: involvement of heme oxygenase-1. Biotechnology and Bioprocess Engineering. 2009;14(3):295–301. [Google Scholar]
- 29.Yoshimura Y, Inomata T, Nakazawa H, Kubo H, Yamaguchi F, Ariga T. Evaluation of free radical scavenging activities of antioxidants with an H2O2/NaOH/DMSO system by electron spin resonance. Journal of Agricultural and Food Chemistry. 1999;47(11):4653–4656. doi: 10.1021/jf990422w. [DOI] [PubMed] [Google Scholar]
- 30.Casano LM, Martin M, Sabater B. Sensitivity of superoxide dismutase transcript levels and activities to oxidative stress is lower in mature-senescent than in young barley leaves. Plant Physiology. 1994;106(3):1033–1039. doi: 10.1104/pp.106.3.1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Manzer R, Wang J, Nishina K, McConville G, Mason RJ. Alveolar epithelial cells secrete chemokines in response to IL-1β and lipopolysaccharide but not to ozone. American Journal of Respiratory Cell and Molecular Biology. 2006;34(2):158–166. doi: 10.1165/rcmb.2005-0205OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cossarizza A, Baccarani-Contri M, Kalashnikova G, Franceschi C. A new method for the cytofluorimetric analysis of mitochondrial membrane potential using the J-aggregate forming lipophilic cation 5,5’,6,6’-tetrachloro-1,1’,3,3’-tetraethylbenzimidazolcarbocyanine iodide (JC-1) Biochemical and Biophysical Research Communications. 1993;197(1):40–45. doi: 10.1006/bbrc.1993.2438. [DOI] [PubMed] [Google Scholar]
- 33.Connop BP, Thies RL, Beyreuther K, Ida N, Reiner PB. Novel effects of FCCP [carbonyl cyanide p-(trifluoromethoxy)phenylhydrazone] on amyloid precursor protein processing. Journal of Neurochemistry. 1999;72(4):1457–1465. doi: 10.1046/j.1471-4159.1999.721457.x. [DOI] [PubMed] [Google Scholar]
- 34.Wataha JC, Hanks CT, Craig RG. The in vitro effects of metal cations on eukaryotic cell metabolism. Journal of Biomedical Materials Research. 1991;25(9):1133–1149. doi: 10.1002/jbm.820250907. [DOI] [PubMed] [Google Scholar]
- 35.Carmichael J, DeGraff WG, Gazdar AF. Evaluation of a tetrazolium-based semiautomated colorimetric assay: assessment of chemosensitivity testing. Cancer Research. 1987;47(4):936–942. [PubMed] [Google Scholar]
- 36.Fridovich I. Superoxide radical and superoxide dismutases. Annual Review of Biochemistry. 1995;64:97–112. doi: 10.1146/annurev.bi.64.070195.000525. [DOI] [PubMed] [Google Scholar]
- 37.Levine RL, Garland D, Oliver CN, et al. Determination of carbonyl content in oxidatively modified proteins. Methods in Enzymology. 1990;186:464–478. doi: 10.1016/0076-6879(90)86141-h. [DOI] [PubMed] [Google Scholar]
- 38.Toraason M, Clark J, Dankovic D, et al. Oxidative stress and DNA damage in Fischer rats following acute exposure to trichloroethylene or perchloroethylene. Toxicology. 1999;138:43–53. doi: 10.1016/s0300-483x(99)00083-9. [DOI] [PubMed] [Google Scholar]
- 39.Cai J, Yang J, Jones DP. Mitochondrial control of apoptosis: the role of cytochrome c. Biochimica et Biophysica Acta. 1998;1366(1-2):139–149. doi: 10.1016/s0005-2728(98)00109-1. [DOI] [PubMed] [Google Scholar]
- 40.Nishikawa T, Araki E. Impact of mitochondrial ROS production in the pathogenesis of diabetes mellitus and its complications. Antioxidants & Redox Signaling. 2007;9:343–353. doi: 10.1089/ars.2006.1458. [DOI] [PubMed] [Google Scholar]
- 41.Green K, Brand MD, Murphy MP. Prevention of mitochondrial oxidative damage as a therapeutic strategy in diabetes. Diabetes. 2004;53(1):S110–S118. doi: 10.2337/diabetes.53.2007.s110. [DOI] [PubMed] [Google Scholar]
- 42.Jang YY, Song JH, Shin YK, Han ES, Lee CS. Protective effect of boldine on oxidative mitochondrial damage in streptozotocin-induced diabetic rats. Pharmacological Research. 2000;42(4):361–371. doi: 10.1006/phrs.2000.0705. [DOI] [PubMed] [Google Scholar]
- 43.Zhang L, Zalewski A, Liu Y, et al. Diabetes-induced oxidative stress and low-grade inflammation in porcine coronary arteries. Circulation. 2003;108(4):472–478. doi: 10.1161/01.CIR.0000080378.96063.23. [DOI] [PubMed] [Google Scholar]
- 44.Yen H-C, Oberley TD, Gairola CG, Szweda LI, St. Clair DK. Manganese superoxide dismutase protects mitochondrial complex I against adriamycin-induced cardiomyopathy in transgenic mice. Archives of Biochemistry and Biophysics. 1999;362(1):59–66. doi: 10.1006/abbi.1998.1011. [DOI] [PubMed] [Google Scholar]
- 45.Kakkar R, Mantha SV, Radhi J, Prasad K, Kalra J. Increased oxidative stress in rat liver and pancreas during progression of streptozotocin-induced diabetes. Clinical Science. 1998;94(6):623–632. doi: 10.1042/cs0940623. [DOI] [PubMed] [Google Scholar]
- 46.Adachi T, Ohta H, Hirano K, Hayashi K, Marklund SL. Non-enzymic glycation of human extracellular superoxide dismutase. Biochemical Journal. 1991;279(1):263–267. doi: 10.1042/bj2790263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Salvioli S, Sikora E, Cooper EL, Franceschi C. Curcumin in cell death processes: a challenge for CAM of age-related pathologies. Evidence-Based Complementary and Alternative Medicine. 2007;4(2):181–190. doi: 10.1093/ecam/nem043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Croteau DL, ap Rhys CMJ, Hudson EK, Dianov GL, Hansford RG, Bohr VA. An oxidative damage-specific endonuclease from rat liver mitochondria. Journal of Biological Chemistry. 1997;272(43):27338–27344. doi: 10.1074/jbc.272.43.27338. [DOI] [PubMed] [Google Scholar]
- 49.Cabreiro F, Picot CR, Perichon M, Castel J, Friguet B, Petropoulos I. Overexpression of mitochondrial methionine sulfoxide reductase B2 protects leukemia cells from oxidative stress-induced cell death and protein damage. Journal of Biological Chemistry. 2008;283(24):16673–16681. doi: 10.1074/jbc.M708580200. [DOI] [PubMed] [Google Scholar]
- 50.Yao K, Ye PP, Zhang L, Tan J, Tang XJ, Zhang YD. Epigallocatechin gallate protects against oxidative stress-induced mitochondria-dependent apoptosis in human lens epithelial cells. Molecular Vision. 2008;14:217–223. [PMC free article] [PubMed] [Google Scholar]
- 51.Singh KK. Mitochondria damage checkpoint in apoptosis and genome stability. FEMS Yeast Research. 2004;5(2):127–132. doi: 10.1016/j.femsyr.2004.04.008. [DOI] [PubMed] [Google Scholar]
- 52.Morita N, Iizuka K, Okita K, et al. Exposure to pressure stimulus enhances succinate dehydrogenase activity in L6 myoblasts. American Journal of Physiology. 2004;287(6):E1064–E1069. doi: 10.1152/ajpendo.00364.2003. [DOI] [PubMed] [Google Scholar]
- 53.Sakai K, Matsumoto K, Nishikawa T, et al. Mitochondrial reactive oxygen species reduce insulin secretion by pancreatic β-cells. Biochemical and Biophysical Research Communications. 2003;300(1):216–222. doi: 10.1016/s0006-291x(02)02832-2. [DOI] [PubMed] [Google Scholar]