

Abstract
The neurobiological bases of social learning, by which an animal can ‘exploit the expertise of others' and avoid the disadvantages of individual learning, are only partially understood. We examined the involvement of the dopaminergic system in social learning by administering a dopamine D1-type receptor antagonist, SCH23390 (0.01, 0.05, and 0.1 mg/kg), or a D2-type receptor antagonist, raclopride (0.1, 0.3, and 0.6 mg/kg), to adult female mice prior to socially learning a food preference. We found that while SCH23390 dose-dependently inhibited social learning without affecting feeding behavior or the ability of mice to discriminate between differently flavored diets, raclopride had the opposite effects, inhibiting feeding but leaving social learning unaffected. We showed that food odor, alone or in a social context, was insufficient to induce a food preference, proving the specifically social nature of this paradigm. The estrous cycle also affected social learning, with mice in proestrus expressing the socially acquired food preference longer than estrous and diestrous mice. This suggests gonadal hormone involvement, which is consistent with known estrogenic regulation of female social behavior and estrogen receptor involvement in social learning. Furthermore, a detailed ethological analysis of the social interactions during which social learning occurs showed raclopride- and estrous phase-induced changes in agonistic behavior, which were not directly related to effects on social learning. Overall, these results suggest a differential involvement of the D1-type and D2-type receptors in the regulation of social learning, feeding, and agonistic behaviors that are likely mediated by different underlying states.
Keywords: social transmission, social learning, agonistic behavior, feeding, dopamine receptor, estrous cycle
INTRODUCTION
Social learning, an adaptive, biologically significant type of learning in which an animal's behavior is influenced by observation or interaction with another (Heyes, 1994), allows animals to avoid potentially costly trial-and-error learning (Galef and Laland, 2005). Social learning has been described in numerous wild and laboratory animal species (Choleris and Kavaliers, 1999; Galef and Laland, 2005), and is often tested with the social transmission of food preferences (STFP) paradigm (Galef et al, 1984). In this paradigm, social learning is observed when, during a brief interaction with a conspecific who has just eaten a novel diet, an ‘observer' (OBS) can acquire olfactory information about food eaten by a conspecific ‘demonstrator' (DEM) by sniffing the DEM's mouth area (Galef et al, 1988). Specific neurons in the olfactory system (Munger et al, 2010) detect carbon disulfide, a semiochemical product of digestion needed for the social learning of a food preference (Galef et al, 1988; Heyes and Durlach, 1990), so investigation of the DEM's mouth area is critical for STFP in rats (Galef and Stein, 1985) and mice (Valsecchi and Galef, 1989).
Rather than just socially increasing the salience of the food-related stimulus, the OBS may develop an actual preference for its DEM's food (Heyes and Durlach, 1990), as the OBS eat more of the DEM food in a choice paradigm, and display hedonic responses to the DEM food in the taste reactivity paradigm (Galef et al, 1997). Classical appetitive conditioning predicts that social learning involves a positive and rewarding state associated with the social interaction, and that a non-rewarding social context would produce less or no learning. This is consistent with the elimination of STFP in strange gerbils, who display heightened aggression (Valsecchi et al, 1996), and STFP's reinstatement by aggression-reducing anxiolytic drug chlordiazepoxide (Choleris et al, 1998).
Dopamine (DA) has been implicated in various types of learning, likely through actions on synaptic plasticity (Calabresi et al, 2007) and/or motivation (Wise, 2006). Acting through its D1-type (D1 and D5) and D2-type (D2, D3, and D4) receptors, DA mediates learning about novel flavors and spontaneous flavor preferences (Rusk and Cooper, 1988, 1989; Sills et al, 1998), such that the two DA receptor (DAR) types are similarly involved in the former, but affect the latter differently (Rusk and Cooper, 1988, 1989, 1994; Yu et al, 2000a, 2000b; Baker et al, 2003; Cooper and Al-Naser, 2006).
The dopaminergic system mediates several motivated behaviors (Berridge, 2007). For example, optimal levels of DA function are needed for normal feeding behavior (Wise, 2006) and normal social behavior. Systemic administration of both DA agonists and antagonists can reduce feeding (Bednar et al, 1995; Szczypka et al, 1999), and DAR and DA-deficient mice work less for and eat less of a palatable or preferred food (Szczypka et al, 1999; Wise, 2006), while DA-overexpressing mice show increased consumption of and responding for a rewarding, palatable food (Pecina et al, 2003; Cagniard et al, 2006). DA and DAR are also involved in social behavior, from affiliative social bonding (Wang et al, 1999; Gingrich et al, 2000; Curtis et al, 2006; Aragona et al, 2006) to aggressive behavior (Cabib et al, 2000), and both DAR types affect locomotor and social aspects of social reactivity (Puglisi-Allegra and Cabib, 1988; Gendreau et al, 1997, 2000; Gariepy et al, 1998).
DA's involvement in feeding behavior, expression of spontaneous food preferences, development and maintenance of acquired flavor preferences, and aspects of social behavior make the dopaminergic system a good candidate for involvement in STFP. Indeed, OBS mice in which the DA transporter (DAT) has been eliminated (‘knocked out,' KO) preferred the food not eaten by their DEM, suggesting a reversal of social learning (Rodriguiz et al, 2004). In the present study, we administered D1-type or D2-type DAR antagonists prior to STFP. A detailed ethological analysis, performed on the social interactions during which STFP occurs, allowed us to assess effects of the two drugs on social and non-social aspects of the mice's behavior (Clipperton et al, 2008); effects on feeding were also analyzed. Due to our recent finding that estrogen receptor (ER)-α blocked and ER-β prolonged the preference for the demonstrated food in female mice in STFP (Clipperton et al, 2008), and DA's known estrous-cycle-related natural fluctuations (Thompson and Moss, 1997), the mice's estrous cycle was monitored and its involvement in STFP was assessed. Separate control experiments also assessed the specificity of STFP and any drug effects.
MATERIALS AND METHODS
Animals
Experimentally naive, 3- to 4-month-old, CD-1 female mice (Charles Rivers, QC, Canada) were housed in same-sex pairs in clear polyethylene cages (26 × 16 × 12 cm3), provided with corncob bedding, environmental enrichment (plastic containers), paper nesting material, and food (Purina rodent chow) and tap water ad libitum, and allowed at least 1 week to adjust to the colony room before testing. The colony room was kept at 21±1°C, under a 12 : 12-h light/dark cycle (lights on at 2000 hours). One mouse per dyad was ear punched for permanent identification. After being OBS in the STFP, several non-drug-treated mice were re-utilized as DEM. This research was conducted in accordance with the regulations of the Canadian Council on Animal Care and approved by the Institutional Animal Care and Use Committee of the University of Guelph.
Materials
In the STFP, DEM feeding occurred in clear polyethylene cages (26 × 16 × 12 cm3), with a 5-cm high, 7.5-cm wide cylindrical jar (Dyets, Bethlehem, PA) fitted with a stainless steel ring (2.5 cm diameter) and sleeve collar to prevent spillage. For olfactory exposure to the diets in the individual learning and social context paradigms, 5.5 cm mason jars (6.5 cm diameter), which had wire mesh (0.5 cm grid) fitted into their screw-on lids were used. Binary choice tests in the STFP, individual learning, and social context paradigms occurred in 31 × 23 × 24 cm3 DietMax clear acrylic cages (Accuscan Instruments, Columbus, OH) with stainless steel grid floors. One side of each cage held two Plexiglas tunnels (5.5 × 3.75 × 3 cm3) leading to two 6 × 4 × 2.5 cm3 feeding trays, each on a scale (AND, Bradford, MA) connected to a DietMax analyzer and Dell computer that stored each scale's reading every 1.5 s. Water bottles were located equidistantly between the two feeding stations. In the flavor recognition paradigm, commercial cotton swabs placed in a bent 17 × 2 cm2 piece of wire mesh (from commercial fine-mesh stainless steel colanders) were used. At the end of each experiment, all equipments were washed with unscented soap (Alconox) and baking soda, then paper and air dried.
Flavored Diets
Ground rodent chow was mixed with either 1% ground cinnamon (CIN diet; McCormick Ground Cinnamon, McCormick Canada, London, Canada) or 2% ground cocoa (COC diet; Fry's Premium Cocoa, Cadbury, Mississauga, Canada). Prior unpublished studies established that the two flavored diets had equivalent metabolic and physical features and were equally palatable to other female CD-1 mice from Charles Rivers.
Drugs
The DA D1-type antagonist SCH23390 hydrochloride (Hyttel, 1983) and the DA D2-type antagonist raclopride tartarate (Hall et al, 1988) were obtained from Sigma-Aldrich (Oakville, ON, Canada) and dissolved in saline solution (0.9% NaCl).
Experimental Procedures
STFP paradigm
Two pharmacological studies used 300 mice that were pair caged for a minimum of 3 days before testing, then weighed, transferred to the experimental room, food deprived, and left undisturbed for 12–14 h. One mouse from each dyad was designated DEM and the other OBS. Early in the dark active phase, DEM were placed for 1 h in clean cages with one jar containing either the COC or the CIN diet, and then DEM were returned to home cages and left to interact, undisturbed, with their respective OBS for 30 min. Immediately after social interaction, OBS were moved to testing cages and given a 24 h choice between the COC and CIN diets, during which both diets were continuously available. In two separate experiments, OBS received a 10 ml/kg intraperitoneal injection of either SCH23390 (0.01, 0.05, or 0.1 mg/kg) or raclopride (0.1, 0.3, or 0.6 mg/kg) 15 or 20 min, respectively, prior to the interaction with their DEM during which social learning occurs. In each experiment, two additional groups of OBS were either injected with saline solution or received no injection (untreated group). In both experiments, only those OBS whose DEM's food consumption was at least 0.1 g were kept in the data pool (final group sample sizes are in Figures 1 and 2). All social interactions were videotaped (8 mm Sony Handycam) for behavioral analysis. A trained investigator unaware of the OBS's treatment scored the videotapes using specific software (The Observer, Noldus, The Netherlands). The behavioral elements collected from the OBS, described in Supplementary Table S1 in Supplementary Information, included measures of social investigation (active sniffing of DEM), horizontal and vertical activity (not in relation to the DEM), anxiety-related behaviors (stretched approaches; Blanchard et al, 1993; Choleris et al, 2003; Clipperton et al, 2008), and other active (eg, digging in bedding) and non-active behaviors (eg, sitting). Variables calculated from groups of behavioral elements (Supplementary Table S2 in Supplementary Information) provided general assessment of mice's activity, social and aggressive motivation, and dominance status.
Figure 1.
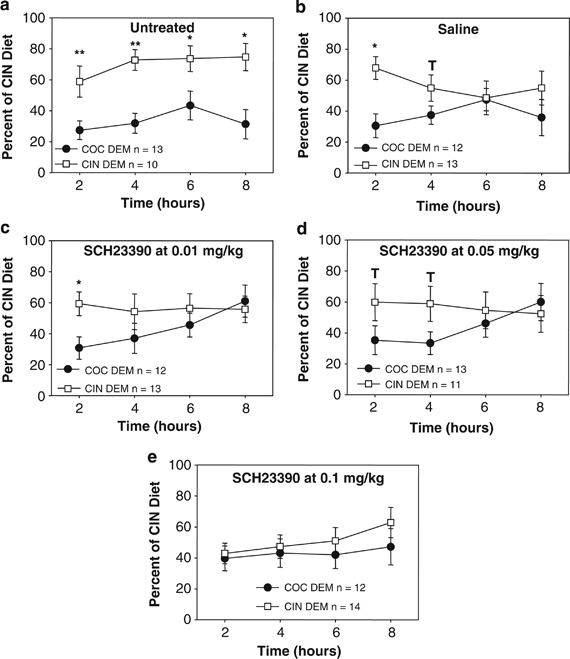
Percent of cinnamon (CIN) diet consumed over the total food intake by the observer female mice across all phases of the estrous cycle that had received either no treatment (a), saline vehicle (b), or the D1-type antagonist SCH23390 at 0.01 mg/kg (c), 0.05 mg/kg (d), or 0.1 mg/kg (e) 15 min prior to a 30-min social interaction with a demonstrator (DEM) female mouse that had just consumed either a 2% cocoa (COC, black circle) or a 1% CIN (white square) flavored diet. CIN preferences are shown at 2, 4, 6, and 8 h from the beginning of testing. **p<0.001; *p<0.01; p<0.05; T=0.05 < p<0.1 in the comparison between the COC DEM and CIN DEM observers.
Figure 2.

Percent of cinnamon (CIN) diet consumed over the total food intake by the observer female mice across all phases of the estrous cycle that had received either no treatment (a), saline solution (b), or the D2-type antagonist raclopride at 0.1 mg/kg (c), 0.3 mg/kg (d) or 0.6 mg/kg (e) 20 min prior to a 30-min social interaction with a demonstrator (DEM) female mouse that had just consumed either a 2% cocoa (COC, black circle) or a 1% CIN (white square) flavored diet. CIN preferences are shown at 2, 4, 6, and 8 h from the beginning of testing. **p<0.001; *p<0.01; p<0.05 in the comparison between the COC DEM and CIN DEM observers.
Control study paradigms
Social Context Learning This paradigm was as the STFP described above, except that the DEM ate unflavored ground rodent chow, and jars containing either COC (n=10) or CIN (n=12) diets, covered with wire mesh to prevent consumption, were placed in the cages during the social interaction. Twenty-two dyads of untreated mice were tested in this paradigm, in which OBS were exposed to the COC or CIN diet in a social context, except that diet odor was not carried by the DEM. Behavioral data were collected from the videos of six randomly chosen mice.
Individual Learning Thirty-one single-housed, untreated females were tested as described above, except that their 30 min exposure to a jar containing either COC (n=15) or CIN (n=16) flavored diet took place without social interaction. Behavioral data were collected from the videos of six randomly chosen mice.
Flavor Recognition Twenty-three single-housed mice received five consecutive tests in home cage. In the first four habituation tests, a cotton swab dipped in tap water and one of the two flavored diets was covered with fine wire mesh and lowered into the cage through gaps in the lid. In the fifth (dishabituation) test, the swab carried the other odor. The ‘meshed' swab was held with a commercial plastic clothespin on top of the cage's lid and left in place for 5 min. Half of the mice received CIN during the four habituation tests and COC in the dishabituation test, while the other had the reverse. Fifteen minutes prior to testing, mice received a 10-ml/kg intraperitoneal injection of either 0.1 mg/kg SCH23390 (n=10), saline solution (n=8), or no injection (n=5). The drug dose was chosen because it affected social learning. A trained investigator, 2 m away from the cage, used two stopwatches to measure latency and duration of mouse investigation of the swab.
Estrous phase determination
One hour prior to the learning experiments and immediately after the flavor recognition paradigm, vaginal smears were taken from OBS and DEM mice. Methylene blue- or giemsa-stained (Sigma-Aldrich) microscope slides were analyzed under a microscope ( × 100). Proestrus consisted of a predominance of nucleated epithelial cells, with few leukocytes or cornified epithelial cells; estrus was predominantly cornified epithelial cells, with few faded nucleated epithelial cells; diestrus consisted of a predominance of leukocytes, with occasional cornified or nucleated epithelial cells (Clipperton et al, 2008).
Statistical analyses
Consumption of each diet in the choice tests was calculated at 2 h intervals. OBS choice data were expressed as arcsine transformed CIN preference ratios—the amount of CIN diet eaten divided by the total food consumed (CIN/(COC+CIN)). Both the CIN preference ratio and the total amount of food eaten by each OBS (COC+CIN) at each 2 h interval were analyzed with mixed-design analyses of variance (ANOVAs). In the two STFP studies, main factors were DEM's food (COC and CIN), drug treatment (untreated, saline, 0.01, 0.05, or 0.1 mg/kg of SCH23390, or 0.1, 0.3, or 0.6 mg/kg of raclopride), and estrous phase of OBS and DEM (proestrus, estrus, and diestrus). CIN preference and total food consumption data from the individual and social context learning paradigms were also analyzed with mixed-model ANOVAs with pre-exposed diets (COC and CIN) and estrous phase as main factors. In all learning paradigms, time (the 2 h intervals) was the repeated measure. Data from the flavor recognition paradigm were also analyzed with a mixed-model ANOVA, with habituation flavor (COC and CIN), treatment (0.1 mg/kg SCH23390, saline, untreated), and estrous phase as main factors and test (1–5) as the repeated measure.
An additional analysis was run on the pooled data of the untreated groups from the two pharmacological studies. A mixed-design ANOVA assessed the effects of demonstrated food and OBS and DEM estrous phase on the CIN preference ratio of the OBS mice in the social learning paradigm.
Duration and frequency of the various individual and grouped behaviors collected from the social interactions were expressed in 5 min intervals over the 30-min social interaction. Behavior data were ln transformed when needed to achieve normality and analyzed with mixed-design ANOVAs similar to those used for the CIN preference scores. When normality could not be achieved, data were analyzed with the Kruskal–Wallis and Mann–Whitney U non-parametric tests. Group mean comparisons were planned a priori in all ANOVA models. Because analyses of the duration, frequency, and latency of the various behaviors provided mostly consistent results, only the durations are reported below; other analyses are reported only when meaningfully different. Throughout, all reported drug effects are in comparison with saline-treated mice.
Main models used α=0.05 as the criterion for significance and the Greenhouse–Geisser correction for repeated measures was applied. Selected contrasts were planned a priori in the ANOVA model and protected against type I error for multiple comparisons with appropriate adjustments of α-levels based on differences in variance and sample size. Analyses were performed with Statview (SAS Institute, Cary, NC) and SuperANOVA (Abacus Concepts, Berkeley, CA) statistical packages.
RESULTS
Social Learning Study: Effects of the D1-Type Antagonist, SCH23390, on Social Learning and Feeding
The ANOVA run on the CIN preference ratios revealed main effects of treatment (F(4, 21)=4.24, p<0.025) and OBS estrous phase (F(2, 12)=6.20, p<0.016; see Figure 1). Mean comparisons revealed no further effect of the OBS estrous phase, likely because of the small number of mice at any given phase for each treatment group. Mean comparisons did show that SCH23390 blocked STFP independently of the phase of the estrous cycle (Figure 1a–e). In the first 2 h interval, the food eaten by their DEM significantly affected the OBS' CIN preferences in the untreated (F(1, 22)=6.80, p<0.010), saline injected (F(1, 23)=9.74, p<0.0023) and 0.01 mg/kg SCH23390-treated mice (F(1, 24)=5.25, p<0.024). In the 0.05-mg/kg SCH23390 group, this only approached statistical significance (F(1, 23)=3.43, p<0.067), while the OBS that had received 0.1 mg/kg SCH23390 showed no significant preference for their DEM food (F(1, 25)=0.79, ns). At 4 h of testing, only the untreated group still showed a significant preference for their DEM food (F(1, 22)=10.12, p<0.0019), with the saline and SCH23390 0.05 mg/kg treated mice only approaching statistical significance (saline: F(1, 223)=3.13, p<0.080; 0.05 mg/kg SCH23390: F(1, 23)=3.25, p<0.074). Only the untreated group still showed a significant effect of the demonstrated food at 6 h (F(1, 22)=5.24, p<0.024) and 8 h (F(1, 18)=5.41, p<0.022) of testing. At all other time intervals, there was no effect of the DEM food on the CIN preferences of the OBS of any of the groups.
The ANOVA run on the food intake of the OBS showed no effects of treatment (Figure 4a), but revealed a significant main effect of OBS estrous phase (F(2, 75)=4.25, p<0.018; data not shown) as well as significant OBS estrous phase × treatment (F(8, 75)=2.75, p<0.017) and OBS estrous phase × time interactions (F(22, 825)=2.12, p<0.002). Mean comparisons revealed no further significances of the OBS estrous phase in relation to the various groups of treatment, but did show an effect of the OBS estrous phase on OBS food intake, independent of treatment. In the first 2 h of testing, which is when all mice ate the most food (Figure 3a), diestrous mice ate significantly more than mice in estrus (F(1, 103)=4.98, p<0.028) and proestrus (F(1, 85)=4.15, p<0.045). At 4 h of testing, mice in estrus ate significantly less than those in diestrus (F(1, 103)=15.37, p<0.0002) and proestrus (F(1, 107)=5.23, p<0.025). There were no effects of the OBS estrous phase on the amount of food eaten at the other time intervals except for the last 2 h interval (from 22 to 24 h), when the next-day dark active phase of the mice began and their feeding behavior resumed. At this point, again, diestrous mice ate more than proestrous (F(1, 85)=14.71, p<0.0003) and estrous mice (F(1, 103)=3.58, p<0.062).
Figure 3.

Percent of cinnamon (CIN) diet consumed over the total food intake by the observer female mice that had received no treatment, combined from the two pharmacological studies, and were in proestrus (a), estrus (b), or diestrus (c) on the day they had a 30-min social interaction with a demonstrator (DEM) female mouse that had just consumed either a 2% cocoa (COC, black circle) or a 1% CIN (white square) flavored diet. CIN preferences are shown at 2, 4, 6, and 8 h from the beginning of testing. **p<0.001; *p<0.01; p<0.05; T=0.05 < p<0.1 in the comparison between the COC DEM and CIN DEM observers.
Social Learning Study: Effects of the D2-Like Antagonist, Raclopride, on Social Learning and Feeding
The ANOVA run on the CIN preference ratios revealed a significant treatment × DEM food × time interaction (F(33, 143)=1.818, p<0.0089) and a trend for a treatment × DEM food interaction (F(3, 143)=3.21, p<0.059). Mean comparisons (Figure 2) revealed that while the DEM food significantly affected the initial (2 h) preference of the OBS for CIN (untreated: F(1, 23)=19.32, p<0.0001; saline: F(1, 23)=6.04, p<0.015; raclopride 0.1 mg/kg: F(1, 23)=19.72, p<0.0001; raclopride 0.3 mg/kg: F(1, 23)=8.45, p<0.0044; raclopride 0.6 mg/kg: F(1, 23)=8.91, p<0.0035), at 4 h, only the untreated group still showed a significant effect of DEM food (F(1, 23)=3.92, p<0.050). Thus, while raclopride at all doses did not affect social learning, merely receiving a saline injection did reduce the duration of the STFP expression in female mice.
The ANOVA run on the food intake of the OBS revealed significant main effects of treatment (F(4, 737)=6.11, p<0.0003) and time (F(11, 737)=59.03, p<0.0001) as well as a significant treatment × time interaction (F(44, 737)=2.27, p<0.0001; see Figure 4b). Two hours after the beginning of testing, the OBS treated with all three doses of raclopride ate significantly less than OBS that had received saline (0.1 mg/kg: F(1, 47)=26.48, p<0.0001; 0.3 mg/kg: F(1, 47)=31.37, p<0.0001; 0.6 mg/kg: F(1, 47)=10.86, p<0.0014). Beyond the 2 h time point, there were no effects of raclopride on food intake. In this study, there were also no effects of the phase of the estrous cycle on the mice's feeding behavior, and the saline and untreated groups did not differ in their food consumptions.
Figure 4.

Total food intake (in grams) by the observer mice in the initial 8 h of a 24-h test with intakes being reported over 2 h intervals. (a) Observers that had received no treatment (black circle), saline solution (white circle), SCH23390 at 0.01 mg/kg (black square), 0.05 mg/kg (white square), or 0.1 mg/kg (black triangle) 45 min prior to testing. (b) Observers that had received no treatment (black circle), saline solution (white circle), raclopride at 0.1 mg/kg (black square), 0.3 mg/kg (white square), or 0.6 mg/kg (black triangle) 50 min prior to testing. (c) Observers that had received no treatment, combined from the two pharmacological studies, and were in proestrus (black circle), estrus (white square), or diestrus (black triangle) on the day that testing begun. **p<0.001 in the comparison between the raclopride-treated mice (at all three doses) and the saline-treated group.
Social Learning Studies in Untreated Mice: Effects of the Estrous Cycle on Social Learning and Feeding
The ANOVA run on the preference for the CIN food in the untreated mice from both the SCH23390 and raclopride experiments revealed a significant interaction between phase of the estrous cycle of the OBS and food preference over time (F(2, 22)=2.706, p<0.0015; see Figure 3). In the proestrous females (Figure 3a), the food eaten by the DEM affected the OBS' CIN preference for a total of 8 h (2 h: F(1, 14)=10.53, p<0.0059; 4 h: F(1, 14)=5.06, p<0.041; 8 h: F(1, 14)=7.71, p<0.018). The effect of the food eaten by the DEM on the estrous females' CIN preference (Figure 3b) was only marginally significant and lasted for 4 h (2 h: F(1, 10)=4.58, p<0.058; 4 h: F(1, 10)=4.42, p<0.061), half as long as it lasted in the proestrous OBS. In the diestrous females (Figure 3c), the effect of the DEM food on the CIN preference was highly significant at 2 h (F(1, 7)=13.16, p<0.0084), and still approached significance 6 h after the beginning of test (F(1, 7)=4.2, p<0.080). The total intake of the mice was not affected by their stage of the estrous cycle (Figure 4c).
Social Learning Study: Effects of the D1-Type Antagonist, SCH23390, the D2-Like Antagonist, Raclopride, and the Phase of the Estrous Cycle on Behavior During the Social Interactions
The results of these analyses are reported in detail in the online Supplementary Material. Briefly, we found that, while both drugs reduced the overall activity of the mice during the social interactions, there were differences in the type of behaviors affected. SCH23390 affected predominantly non-social aspects of the mice's behavior (Supplementary Figure S1 in Supplementary Information), while raclopride affected both social and non-social aspects of behavior (Supplementary Figure S2 in Supplementary Information). SCH23390 reduced total activity without affecting overall time engaged in social behavior, compensating for an increase in the time the mice spent in social inactivity with a reduction in active social investigation of the body of the DEM. Thus, it appears that the overall motivation to interact with a familiar conspecific was unaffected by SCH23390 and that its inhibitory effects on social learning may be directly on learning mechanisms, rather than on social motivation. Raclopride, instead, reduced both social investigatory (oronasal and body investigation) and agonistic behaviors initiated by the treated mouse in a manner that resulted in an overall reduction of the OBS's dominance over the DEM (Figure 5b; Supplementary Figure S2 in Supplementary Information). Thus, raclopride may have affected the specific motivation underlying social agonistic behavior over and above its nonspecific effects on activity and without affecting social learning.
Figure 5.

Oronasal investigation (in seconds) of the demonstrator by the observer female mice in all phases of the estrous cycle during the 30-min social interaction with their demonstrator. In (a), observer mice had received either no treatment (n=23) or an intraperitoneal injection of either saline vehicle (n=25) or the D1-type antagonist SCH23390 at 0.01 mg/kg (n=25), 0.05 mg/kg (n=24), and 0.1 mg/kg (n=26). In (b), observer mice had received either no treatment (n=8) or an intraperitoneal injection of either saline vehicle (n=20) or the D2-type antagonist raclopride at 0.1 mg/kg (n=19), 0.3 mg/kg (n=20), and 0.6 mg/kg (n=19). In (c) the pooled untreated observer mice from (a) and (b) were either in diestrus (n=6), proestrus (n=13), or estrus (n=10) on the day of testing. **p<0.001; *p<0.01; p<0.05; in comparison to the saline-treated control group.
The behavior during the social interactions in the pooled untreated groups from the two social learning studies was affected by the estrous phase of both mice. We found that overall levels of activity were lower when either the OBS or the DEM were in diestrus (Supplementary Figure S3A in Supplementary Information). This effect was predominantly reflected in a reduction in non-social active behaviors (horizontal and vertical exploration) and a parallel increase in non-social inactivity (solitary inactivity). While social behavior, overall, was not affected by the estrous phase (Supplementary Figure S3C in Supplementary Information), agonistic behavior was selectively affected: diestrous OBS showed a reduced dominance score (Supplementary Figure S3B in Supplementary Information) due to increased amounts of agonistic behavior received from the DEM.
Control Studies
In the social context and individual learning studies, the analyses performed on the CIN preference ratios showed no statistically significant effects, indicating that the food choices of the mice were not affected by prior olfactory exposure to one of the two flavored diets alone or in the context of a social interaction with a conspecific that has not consumed a flavored diet (Figure 6a and b). As well, the phase of the estrous cycle of OBS and DEM did not affect the mice food choices in these control studies. The food-deprived mice in both experiments showed high levels of investigation of the meshed top of the food-containing jar (individual learning experiment mean=516 s, SE=100.6; social context experiment mean=579 s, SE=53.9). In the social context study, this was greater than the time the mice spent engaged in social interactions (mean=225 s, SE=76.7), including oronasal investigation (mean=78 s, SE=20.3). Social and flavor investigations were comparable to those observed in the social learning paradigms, suggesting that the lack of food preference learning in these two control studies was not due to reduced social motivation or lower exposure to the food odor.
Figure 6.

Percent of cinnamon (CIN) diet consumed over the total food intake by the female mice that had received no treatment and were exposed to the odor of either a 2% cocoa (COC, black circle) or a 1% CIN (white square) flavored diet for 30 min either individually (a), or while interacting with another female mouse that had just consumed an unflavored diet (b). CIN preferences are shown over an 8 h test at 2 h intervals. In (c), the time (in seconds) that the mice spent investigating the extreme of a cotton swab covered with either a 2% cocoa (COC) or a 1% CIN flavored diet over five tests (T1–T5) with 5 min exposures. Fifteen minutes prior to T1, the mice either received no treatment (white circles), an intraperitoneal injection of saline solution (white triangles), or the D1-type antagonist SCH23390 at 0.1 mg/kg (black square). Over T1–T4, the mice could habituate to one of the two flavored diets. At T5, the mice received the flavored diet to which they had not habituated. *p in all three groups, significantly different from both T1 (habituation) and T5 (dishabituation); p<0.01 for the saline and SCH23390 at 0.1 mg/kg groups. p<0.05 for the untreated group.
In the flavor recognition study, the ANOVA run on the time spent investigating the flavored swab revealed a statistical trend for the repeated measure time (F(4, 36)=2.28, p<0.080). Planned mean comparisons showed that in all three groups of treatment, the mice had a normal habituation–dishabituation response (Figure 6c). At test 4, they all investigated the swab less than at test 1 (untreated: F(1, 9)=5.59, p<0.02; saline: F(1, 15)=7.10, p<0.01; 0.1 mg/kg SCH23390: F(1, 19)=11.34, p<0.002), indicating that they all habituated to the repeatedly presented flavor. Mice of all three groups at test 5 showed greater swab investigation than at test 4 (untreated: F(1, 9)=4.91, p<0.04; saline: F(1, 15)=14.49, p<0.0007; 0.1 mg/kg SCH23390: F(1, 19)=14.92, p<0.0004), revealing that they all had a dishabituation response when the swab carried a novel flavor. This indicated that the dose of SCH23390 that blocked the STFP did not impair the mice's capabilities to discriminate between the two flavored diets (COC or CIN). Treatment, phase of the estrous cycle, and habituated flavor did not affect the mice's habituation–dishabituation responses, with neither the factors in the main model nor the planned mean comparisons revealing any statistically significant results.
DISCUSSION
In the present study with female mice, we found that while blocking the DA D1 receptor with SCH23390 dose-dependently inhibited social learning (Figure 1) without affecting feeding behavior per se (Figure 4a), blocking the DA D2 receptor with raclopride had the opposite effects: it did not affect social learning (Figure 2), while it inhibited feeding behavior (Figure 4b). This suggests that the STFP is mediated by the D1-type, while food consumption is mediated by the D2-type receptors. Conversely, aspects of social interactions that are related to dominance hierarchies and agonistic behaviors between familiar female mice appear to be mediated more by D2-type than D1-type receptors.
The estrous cycle also affected social learning, as mice in proestrus expressed the socially acquired food preference for a longer time than mice in estrus and diestrus (Figure 3). As well, mice in the non-reproductive phase (diestrus) were less active and performed less agonistic behavior. The ethological analysis of the social interactions (Figure 5; Supplementary Figure S1–S3 in Supplementary Information) and the results of targeted control studies (Figure 6) further demonstrated the exquisitely social nature of the STFP learning paradigm in mice. These control assessments also allowed the possibility that the observed drug effects were either due to sensory impairments (Figure 6c), or were due solely to effects on the overall activity levels of the mice (Supplementary Figures S1–S3 in Supplementary Information), to be ruled out.
Effects of the Dopaminergic D1-Type and D2-Type Antagonists SCH23390 and Raclopride
The differential effects of SCH23390 (inhibitory) and raclopride (no effect) on the STFP are in agreement with D1 and D2 receptors having opposing roles in appetitive conditioning (Eyny and Horvitz, 2003) and in line with results showing a role for the D1 (Beninger and Miller, 1998; Smith-Roe and Kelley, 2000; Azzara et al, 2001; Baldwin et al, 2002; Eyny and Horvitz, 2003; Touzani et al, 2008) but not D2 (Bushnell and Levin, 1993) receptor in appetitive learning in non-social learning paradigms. D2-type receptors can act in direct antagonism to the D1-type receptors in learning mechanisms, with D1 activating and D2 inhibiting the regulatory DA and cAMP-regulated phosphoprotein of 32 kDa (DARPP-32; Calabresi et al, 2000; Otani et al, 2003; Calabresi et al, 2007; Nishi et al, 1997; Svenningsson et al, 2004) in glutamate-dependent long-term potentiation of synapses (Nishi et al, 1997; Otani et al, 2003; Svenningsson et al, 2004; Lemon and Manahan-Vaughan, 2006). A predominance of D2 over D1 effects may explain why increased extracellular DA in DATKO mice reversed the STFP (Rodriguiz et al, 2004). It is possible that had we used higher doses or a more prolonged treatment with raclopride, we would have observed a similar reversal too. Short-latency and short-lasting phase bursts of activation of ventral tegmental area DA projections to the hippocampus (El-Ghundi et al, 1999, 2003; Li et al, 2003; Moncada and Viola, 2007) are believed to foster learning by highlighting novel aspects of a stimulus and/or a response that is contingently or subsequently paired with a reward (Schultz, 2006). In this view, our findings on the STFP for a novel food item are in agreement with results showing that SCH23390 could block novelty seeking in rats (Peters et al, 2007) and the impaired reversal learning of D1KO mice in both spatial and food rewarded learning (El-Ghundi et al, 1999, 2003). Overall, our results with SCH23390 on a hippocampus-dependent learning paradigm (Alvarez et al, 2002; Countryman et al, 2005) support the idea that blocking the D1 receptor impairs the mice's abilities to process novel information, especially when attempting to alter established learned behavior (El-Ghundi et al, 2007).
The findings that food preference was not developed following exposure to the odor of the food alone (Figure 6a, individual learning study), on a powdered gauze-and-cotton-batten surrogate (Valsecchi and Galef, 1989), or during a social interaction with a conspecific that had not eaten the novel diet (Figure 6b, social context study), suggest that, like in rats (Galef et al, 1988), even in mice the food odor must be carried by the DEM. The inhibitory effects of SCH23390 on social learning (Figure 1) cannot be attributed to an effect of the drug on oronasal investigation (Figure 5a), as SCH23390 affected this behavior in a manner that was not consistent with its effects on social learning: the dose that blocked social learning (0.1 mg/kg) did not affect oronasal investigation, while the dose that reduced this behavior (0.01 mg/kg) did not affect social learning. Interestingly, raclopride, which left social learning unaffected, did reduce oronasal investigation. Hence, even a reduced socially mediated exposure to the novel food is sufficient for the transfer of information. Consistently, we have demonstrated that oronasal investigation is not correlated with the strength of the preference for the DEM food expressed by the OBS (Clipperton et al, 2008). This suggests that the total amount of exposure to the food eaten by the DEM does not directly and linearly determine socially acquired food preferences in a proportional manner (ie, the more the oronasal investigation, the stronger the preference). Rather, there may exist a minimal amount of oronasal investigation that is necessary and sufficient for the STFP to occur in mice.
Conceivably, the impairing effect of SCH23390 on social learning could have been mediated by a sensory impairment. The literature on DA involvement in odor discrimination shows mixed results (Doty et al, 1998; Yue et al, 2004; Pavlis et al, 2006; Smith et al, 1998; Rodriguiz et al, 2004; Kruzich and Grandy, 2004), possibly because of different olfaction tests used (eg, detection vs discrimination), different ways the DA receptors were manipulated (single or repeated drug administrations, chronic gene KO), or even to species differences (rats vs mice) in the DA system. In the present study, SCH23390 effects on STFP cannot be directly attributed to deficits in flavor discrimination because the same dose that blocked the STFP did not impair the discrimination of the COC and CIN diets in the habituation/dishabituation paradigm (Figure 6c, flavor recognition study).
In the present study, the involvement of the DA system in food preferences and in total intake were pharmacologically dissociable. Blocking the D1-type receptors inhibited only the socially acquired food preference, while blocking the D2-type receptors inhibited only total intake (the combined consumption of the COC and CIN diets; Figure 4). The STFP in rats (Galef et al, 1988) and possibly also in mice (Bean et al, 1988) involves the formation of an association between the odor of the novel food consumed by the DEM with that of carbon disulfide present in the DEM's breath which is detected by specific neurons in the olfactory system (Munger et al, 2010). This odor–odor association is reminiscent of the flavor–flavor association investigations (Yu et al, 2000a, 2000b; Baker et al, 2003), where a novel flavor was paired with a preferred sweet solution. The ensuing conditioned preference could be blocked with both SCH233390 and raclopride (Yu et al, 2000b; Baker et al, 2003), but only when fructose, not sucrose, was used as the unconditioned stimulus (Baker et al, 2003), with SCH23390 being somewhat more effective than raclopride. The different results of our study may reflect a species difference (rats vs mice), methodological differences (sweet palatable solutions vs regular rodent chow), differences in the learning paradigms used (eg, high number of training session vs a single training/social interaction), or inherent differences in DA involvement in individually and socially acquired flavor preferences. The findings that when a less palatable sweet solution (fructose) was used, raclopride was less effective than SCH23390 at reducing flavor preferences (Baker et al, 2003) and that D1-type, but not of a D2-type agonist enhanced the consumption of a preferred food (Cooper and Al-Naser, 2006), are closer to our results and suggests that the use of palatable vs regular food influences the way DA antagonists affect learned food preferences. This may be due to an interaction between the involvement of DA in spontaneous and acquired food preference. Furthermore, SCH23390, but not raclopride, blocked the acquisition of flavor preferences in a flavor-nutrient-conditioning paradigm (Azzara et al, 2001). This result is more similar to ours, suggesting that the post-ingestive consequences of eating may contribute to the expression of the STFP.
The effects of the two antagonists on feeding behavior per se (the combined consumption of COC and CIN diets) also suggest a differential involvement of the D1-type and D2-type receptors. We found that while SCH23390 had no effects on the OBS's total food consumption (Figure 4a), raclopride at all doses significantly reduced it (Figure 4b). That SCH23390 did not affect feeding per se supports the notion that it affected social learning directly, rather than through a generalized inhibition of the primary food motivation of the mice. This is in agreement with others' results showing that D1-type antagonists or genetic disruption of D1 functions disrupt the mice's motivation to work for a palatable food reward, but not their home-cage feeding on rodent chow or sucrose (Salamone et al, 1997, 2001, 2007). This is reflected in nucleus accumbens DA release increasing in response to acute and intermittent sucrose access, but not when the palatable diet was not novel (Bassareo and Di Chiara, 1997, 1999) or available ad libitum (Rada et al, 2005). As well, DA release did not occur in rats that had ad libitum or intermittent access to the less palatable rodent chow (Rada et al, 2005). These studies suggest a limited involvement of DA in the ingestion of rodent chow as opposed to more palatable diets.
The raclopride-induced reduction in chow consumption observed in the present study (Figure 4b) is consistent with the results of others that showed the involvement of D2-type receptors in food seeking behavior and in the ingestion of rodent chow (Duarte et al, 2003) as well as with findings showing a link between D2-type receptor polymorphisms and vulnerability to anorexia nervosa (Bergen, 2005), overeating, and obesity (Levitan et al, 2006; Davis et al, 2008) in humans. It could be argued that the decreased food consumption observed in the raclopride-treated mice in the present study (Figure 4b) may be secondary to the observed reduction in activity (Supplementary Figure S2A in Supplementary Information). This would be in agreement with previous findings where feeding and motor functions of D2, but not of D1 (El-Ghundi et al, 2003), were difficult to distinguish (Caine et al, 2002; Yu et al, 2000a). However, in the present study, raclopride effects on activity were comparable to, if not lower than, those of SCH23390 (Supplementary Figures S1A and S2A in Supplementary Information), which did not cause any reductions in food intake. This suggests that the treated mice were able to reach and consume the food, and that raclopride may have affected feeding specifically. Overall, the different effects of the D1-type and the D2-type antagonists support the idea that optimal levels of dopaminergic activation are needed for normal feeding behavior (Wise, 2006). This would also explain why systemic administration of both DA agonists and DA antagonists reduced feeding (Bednar et al, 1995; Szczypka et al, 1999).
The reduction in activity caused by both drugs in the present study (Supplementary Figures S1A and S2A in Supplementary Information) was expected, in view of the known involvement of DA in the motor system (Boutrel, 2008). However, the effect on activity was relatively small and it did not extend to the commonly reported increases in stereotypical behavior and self-grooming (Cooper and Al-Naser, 2006). Rather, the low doses used revealed specific effects on either social learning (SCH23390) or on other aspects of social behavior (raclopride). Thus, the low doses used, together with a complete ethological assessment of the mice's interactions, have allowed us to assess specific and differential effects of SCH23390 and raclopride on aspects of behavior, including social learning, social behavior, and feeding, rather than a generalized behavioral disruption.
Effects of the Estrous Cycle
Estrogens have been shown to enhance the activity of the DA system, by both increasing DA release (McDermott et al, 1994; Liu and Xie, 2004), and decreasing DA reuptake by DAT (Karakaya et al, 2007). In the present study, the estrous cycle of the OBS did not interact with SCH23390 to affect social learning. This is likely due to the low number of mice in each of the phases of the estrous cycle in each treatment group. However, in the combined untreated mice from the two pharmacological experiments, the expression of the learned food preference (Figure 3) lasted twice as long in OBS in proestrus (8 h) as in diestrus (4–6 h) and estrus (2–4 h), despite similar levels of oronasal investigation during the social interaction, when the food preference was acquired (Figure 5c). Our results are consistent with those reported by Sanchez-Andrade et al (2005) showing that, when tested 24 h after interacting with a DEM, only OBS that were in proestrus on the day of the interaction showed a socially acquired food preference. This suggests that proestrous levels of gonadal hormones, estrogen and possibly even progesterone (Walmer et al, 1992), facilitate social learning, while their absence inhibits it. Accordingly, in ovariectomized OBS, the socially acquired food preference lasted only 2 h (Clipperton et al, 2008), while high-estrogen postpartum rats showed a stronger preference for the DEM food than virgin females (Fleming et al, 1994). Overall, these results strongly suggest that further studies need to investigate the interplay of DA and gonadal hormones in the regulation of social learning.
Differently from others' results (reviewed in Asarian and Geary, 2006; Fessler, 2003), we did not observe any effects of the phase of the estrous cycle on total food consumption (Figure 4c). This may be due to the fact that the mice in the present study were deprived of food for 12–14 h prior to testing, and the consequent increase in the motivation to eat may have masked any effects of the estrous cycle. Consistently, during the first 2 h of testing, the mice of all groups consumed almost twice as much food as in the following 2 h feeding intervals (Figure 4c). Alternatively, gonadal hormones may affect food preferences more than feeding behavior per se. Accordingly, estrous cycle or estrogenic drug effects on feeding are often observed when a preferred novel food item is provided to the animals (eg, when sucrose is added to either rodent chow or water; Reddy and Kulkarni, 1999; Fudge et al, 2006, but see Eckel et al, 2000). This would explain why we observed an effect of the ovarian cycle on the socially acquired food preferences, but not on feeding behavior per se.
Note on the Effects of the Control Saline Injection
In both social learning studies, we observed a few differences between the untreated and the saline-injected groups. The OBS that had received saline showed a reduction in the duration of the food preference for their DEM food (Figures 1a and 2a) in both experiments. Furthermore, in the SCH23390 social learning experiment, the saline-injected mice showed increased frequency of horizontal activity and decreased frequency of oronasal investigation and agonistic behaviors delivered by the OBS, resulting in a lower dominance score. The reduced duration of the socially acquired food preference may be ascribed to stress-induced impairment in either learning mechanisms (McEwen and Wingfield, 2003) or motivational aspects of learning (via effects on dominance). As the frequency, but not the duration of oronasal investigation, was affected by the saline injection, the total exposure to the food-related cue in the mouth of the DEM was unaffected. This and the lack of correlation between the duration of a socially acquired food preference and oronasal investigation (Clipperton et al, 2008) suggest that DEM food exposure does not explain the shorter preference in the saline-treated mice. Similarly, a dominance explanation seems unlikely in that we previously found no correlation between the dominance score and social learning (see discussion in Clipperton et al, 2008). The increased frequency in horizontal activity and shifting between behaviors may be ascribed to arousal induced by the stress of the injection (Pfaff et al, 2008). Finding that saline injection can affect stress and anxiety-like behavior is neither surprising nor new (Lapin, 1995). However, in the present study, these effects were not major and do not directly explain our drug treatment results, because these were assessed in comparison with the saline-injected groups.
Conclusions and Outlook
Our results can have implications for the understanding of the impaired social skills and limited control over aggressive behavior shown by individuals affected by neuropsychiatric disorders of the DA system, such as Parkinson's disease, schizophrenia, and attention deficit hyperreactivity disorder (Eyny and Horvitz, 2003). Our results suggest that disruption in DA functioning may cause an altered perception of stimuli of social origin, which may reflect in inappropriate responses in the social domain. Accordingly, in humans, striatal D2-type levels predict socially desirable responding (Miczek et al, 1994; Steinert et al, 1999; Wright and Klee, 2001; Viggiano et al, 2003).
A better understanding of the involvement of estrogens and their receptors in aspects of social cognition may inform the development of ER-specific hormone replacement therapy (HRT) for the aging women of the ‘baby boom' generation. This may allow the fostering of positive effects, while avoiding negative side effects, of estradiol-based HRTs, including increased risk of breast cancer, cardiovascular disease, and cognitive impairment (Shumaker et al, 2003; Choleris et al, 2008).
Last, we wish to emphasize the importance of performing comprehensive ethological analyses on behavior (Blanchard et al, 1993; Clipperton et al, 2008; Clipperton-Allen et al, 2010, 2011), not to indiscriminately increase the number of data points one collects, but rather, to gain a better understanding of effects and ‘side effects' of experimental manipulations. As well, the complexity of social behavior can only be fully understood when different aspects of it are assessed to include various expressions of agonistic and affiliative behaviors, as well as social recognition and social learning (reviewed in Choleris et al, 2009).
Acknowledgments
We thank Nara Anderson, Rachel Barbero, Rochelle Colling, and Julie Irwin for their help with data collection. We also thank Dr Martin Kavaliers and three anonymous reviewers for very helpful advice on an early version of the article.
The authors declare that this work was funded by NSERC Grant 400212 to EC and a NSERC graduate scholarship to AEC-A. The authors declare that, except for income received from their primary employer, no financial support or compensation has been received from any individual or corporate entity over the past 3 years for research or professional service and there are no personal financial holdings that could be perceived as constituting a potential conflict of interest.
Footnotes
Supplementary Information accompanies the paper on the Neuropsychopharmacology website (http://www.nature.com/npp)
Supplementary Material
References
- Alvarez P, Wendelken L, Eichenbaum H. Hippocampal formation lesions impair performance in an odor-odor association task independently of spatial context. Neurobiol Learn Mem. 2002;78:470–476. doi: 10.1006/nlme.2002.4068. [DOI] [PubMed] [Google Scholar]
- Aragona BJ, Liu Y, Yu YJ, Curtis JT, Detwiler JM, Insel TR, et al. Nucleus accumbens dopamine differentially mediates the formation and maintenance of monogamous pair bonds. Nat Neurosci. 2006;9:133–139. doi: 10.1038/nn1613. [DOI] [PubMed] [Google Scholar]
- Asarian L, Geary N. Modulation of appetite by gonadal steroid hormones. Philos Trans R Soc Lond B Biol Sci. 2006;361:1251–1263. doi: 10.1098/rstb.2006.1860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azzara AV, Bodnar RJ, Delamater AR, Sclafani A. D1 but not D2 dopamine receptor antagonism blocks the acquisition of a flavor preference conditioned by intragastric carbohydrate infusions. Pharmacol Biochem Behav. 2001;68:709–720. doi: 10.1016/s0091-3057(01)00484-1. [DOI] [PubMed] [Google Scholar]
- Baker RM, Shah MJ, Sclafani A, Bodnar RJ. Dopamine D1 and D2 antagonists reduce the acquisition and expression of flavor-preferences conditioned by fructose in rats. Pharmacol Biochem Behav. 2003;75:55–65. doi: 10.1016/s0091-3057(03)00039-x. [DOI] [PubMed] [Google Scholar]
- Baldwin AE, Sadeghian K, Kelley AE. Appetitive instrumental learning requires coincident activation of NMDA and dopamine D1 receptors within the medial prefrontal cortex. J Neurosci. 2002;22:1063–1071. doi: 10.1523/JNEUROSCI.22-03-01063.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Differential influence of associative and nonassociative learning mechanisms on the responsiveness of prefrontal and accumbal dopamine transmission to food stimuli in rats fed ad libitum. J Neurosci. 1997;17:851–861. doi: 10.1523/JNEUROSCI.17-02-00851.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Modulation of feeding-induced activation of mesolimbic dopamine transmission by appetitive stimuli and its relation to motivational state. Eur J Neurosci. 1999;11:4389–4397. doi: 10.1046/j.1460-9568.1999.00843.x. [DOI] [PubMed] [Google Scholar]
- Bean NJ, Galef BG, Mason JR. The effect of carbon-disulfide on food-consumption by house mice. J Wildl Manage. 1988;52:502–507. [Google Scholar]
- Bednar I, Carrer H, Qureshi GA, Sodersten P. Dopamine D-1 or D-2 antagonists enhance inhibition of consummatory ingestive behavior by Cck-8. Am J Physiol Regul Integr Comp Physiol. 1995;38:R896–R903. doi: 10.1152/ajpregu.1995.269.4.R896. [DOI] [PubMed] [Google Scholar]
- Beninger RJ, Miller R. Dopamine D1-like receptors and reward-related incentive learning. Neurosci Biobehav Rev. 1998;22:335–345. doi: 10.1016/s0149-7634(97)00019-5. [DOI] [PubMed] [Google Scholar]
- Bergen AW. DRD2 and anorexia nervosa. Biol Psychiatry. 2005;57:212S. [Google Scholar]
- Berridge KC. The debate over dopamine's role in reward: the case for incentive salience. Psychopharmacology (Berl) 2007;191:391–431. doi: 10.1007/s00213-006-0578-x. [DOI] [PubMed] [Google Scholar]
- Blanchard RJ, Yudko EB, Rodgers RJ, Blanchard DC. Defense system psychopharmacology: an ethological approach to the pharmacology of fear and anxiety. Behav Brain Res. 1993;58:155–165. doi: 10.1016/0166-4328(93)90100-5. [DOI] [PubMed] [Google Scholar]
- Boutrel B. A neuropeptide-centric view of psychostimulant addiction. Br J Pharmacol. 2008;154:343–357. doi: 10.1038/bjp.2008.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bushnell PJ, Levin ED. Effects of dopaminergic drugs on working and reference memory in rats. Pharmacol Biochem Behav. 1993;45:765–776. doi: 10.1016/0091-3057(93)90119-e. [DOI] [PubMed] [Google Scholar]
- Cabib S, D'Amato FR, Puglisi-Allegra S, Maestripieri D. Behavioral and mesocorticolimbic dopamine responses to non aggressive social interactions depend on previous social experiences and on the opponent's sex. Behav Brain Res. 2000;112:13–22. doi: 10.1016/s0166-4328(00)00157-1. [DOI] [PubMed] [Google Scholar]
- Cagniard B, Balsam PD, Brunner D, Zhuang XX. Mice with chronically elevated dopamine exhibit enhanced motivation, but not learning, for a food reward. Neuropsychopharmacology. 2006;31:1362–1370. doi: 10.1038/sj.npp.1300966. [DOI] [PubMed] [Google Scholar]
- Caine SB, Negus SS, Mello NK, Patel S, Bristow L, Kulagowski J, et al. Role of dopamine D2-like receptors in cocaine self-administration: studies with D2 receptor mutant mice and novel D2 receptor antagonists. J Neurosci. 2002;22:2977–2988. doi: 10.1523/JNEUROSCI.22-07-02977.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calabresi P, Gubellini P, Centonze D, Picconi B, Bernardi G, Chergui K, et al. Dopamine and cAMP-regulated phosphoprotein 32 kDa controls both striatal long-term depression and long-term potentiation, opposing forms of synaptic plasticity. J Neurosci. 2000;20:8443–8451. doi: 10.1523/JNEUROSCI.20-22-08443.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calabresi P, Picconi B, Tozzi A, Di Filippo M. Dopamine-mediated regulation of corticostriatal synaptic plasticity. Trends Neurosci. 2007;30:211–219. doi: 10.1016/j.tins.2007.03.001. [DOI] [PubMed] [Google Scholar]
- Choleris E, Gustafsson JA, Korach KS, Muglia LJ, Pfaff DW, Ogawa S. An estrogen-dependent four-gene micronet regulating social recognition: a study with oxytocin and estrogen receptor-alpha and -beta knockout mice. Proc Natl Acad Sci USA. 2003;100:6192–6197. doi: 10.1073/pnas.0631699100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choleris E, Kavaliers M. Social learning in animals: sex differences and neurobiological analysis. Pharmacol Biochem Behav. 1999;64:767–776. doi: 10.1016/s0091-3057(99)00141-0. [DOI] [PubMed] [Google Scholar]
- Choleris E, Valsecchi P, Wang Y, Ferrari P, Kavaliers M, Mainardi M. Social learning of a food preference in male and female Mongolian gerbils is facilitated by the anxiolytic, chlordiazepoxide. Pharmacol Biochem Behav. 1998;60:575–584. doi: 10.1016/s0091-3057(98)00005-7. [DOI] [PubMed] [Google Scholar]
- Choleris E, Clipperton AE, Phan A, Kavaliers M. Estrogen receptor β agonists in neurobehavioral investigations. Curr Opin Investig Drugs. 2008;9:760–773. [PubMed] [Google Scholar]
- Choleris E, Clipperton Allen AE, Phan A, Kavaliers M. Neuroendocrinology of social information processing in rats and mice. Front Neuroendocrinol. 2009;30:442–459. doi: 10.1016/j.yfrne.2009.05.003. [DOI] [PubMed] [Google Scholar]
- Clipperton AE, Spinato JM, Chernets C, Pfaff DW, Choleris E. Differential effects of estrogen receptor alpha and beta specific agonists on social learning of food preferences in female mice. Neuropsychopharmacology. 2008;33:2362–2375. doi: 10.1038/sj.npp.1301625. [DOI] [PubMed] [Google Scholar]
- Clipperton-Allen AE, Almey A, Melichercik A, Choleris E.2011Effects of an estrogen receptor alpha agonist on agonistic behavior in gonadally intact and gonadectomized male and female mice Psychoneuroendocrinology(in press) doi: 10.1016/j.psyneuen.2010.12.010 [DOI] [PubMed]
- Clipperton-Allen AE, Cragg CL, Wood AJ, Pfaff DW, Choleris E. Agonistic behavior in males and females: effects of an estrogen receptor beta agonist in gonadectomized and gonadally intact mice. Psychoneuroendocrinology. 2010;35:1008–1022. doi: 10.1016/j.psyneuen.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper SJ, Al-Naser HA. Dopaminergic control of food choice: contrasting effects of SKF 38393 and quinpirole on high-palatability food preference in the rat. Neuropharmacology. 2006;50:953–963. doi: 10.1016/j.neuropharm.2006.01.006. [DOI] [PubMed] [Google Scholar]
- Countryman RA, Kaban NL, Colombo PJ. Hippocampal c-fos is necessary for long-term memory of a socially transmitted food preference. Neurobiol Learn Mem. 2005;84:175–183. doi: 10.1016/j.nlm.2005.07.005. [DOI] [PubMed] [Google Scholar]
- Curtis JT, Aragona BJ, Wang ZX. Dopamine and monogamy. Brain Res. 2006;1126:76–90. doi: 10.1016/j.brainres.2006.07.126. [DOI] [PubMed] [Google Scholar]
- Davis C, Levitan RD, Kaplan AS, Carter J, Reid C, Curtis C, et al. Reward sensitivity and the D2 dopamine receptor gene: a case-control study of binge eating disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2008;32:620–628. doi: 10.1016/j.pnpbp.2007.09.024. [DOI] [PubMed] [Google Scholar]
- Doty RL, Li C, Bagla R, Huang W, Pfeiffer C, Brosvic GM, et al. SKF 38393 enhances odor detection performance. Psychopharmacology. 1998;136:75–82. doi: 10.1007/s002130050541. [DOI] [PubMed] [Google Scholar]
- Duarte C, Biala G, Le Bihan C, Hamon M, Thiebot MH. Respective roles of dopamine D-2 and D-3 receptors in food-seeking behaviour in rats. Psychopharmacology. 2003;166:19–32. doi: 10.1007/s00213-002-1310-0. [DOI] [PubMed] [Google Scholar]
- Eckel LA, Houpt TA, Geary N. Spontaneous meal patterns in female rats with and without access to running wheels. Physiol Behav. 2000;70:397–405. doi: 10.1016/s0031-9384(00)00278-x. [DOI] [PubMed] [Google Scholar]
- El-Ghundi M, Fletcher PJ, Drago J, Sibley DR, O'Dowd BF, George SR. Spatial learning deficit in dopamine D-1 receptor knockout mice. Eur J Pharmacol. 1999;383:95–106. doi: 10.1016/s0014-2999(99)00573-7. [DOI] [PubMed] [Google Scholar]
- El-Ghundi M, O'Dowd BF, Erclik M, George SR. Attenuation of sucrose reinforcement in dopamine D-1 receptor deficient mice. Eur J Neurosci. 2003;17:851–862. doi: 10.1046/j.1460-9568.2003.02496.x. [DOI] [PubMed] [Google Scholar]
- El-Ghundi M, O'Dowd BF, George SR. Insights into the role of dopamine receptor systems in learning and memory. Rev Neurosci. 2007;18:37–66. doi: 10.1515/revneuro.2007.18.1.37. [DOI] [PubMed] [Google Scholar]
- Eyny YS, Horvitz JC. Opposing roles of D-1 and D-2 receptors in appetitive conditioning. J Neurosci. 2003;23:1584–1587. doi: 10.1523/JNEUROSCI.23-05-01584.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fessler DMT. No time to eat: an adaptionist account of periovulatory behavioral changes. Q Rev Biol. 2003;78:3–21. doi: 10.1086/367579. [DOI] [PubMed] [Google Scholar]
- Fleming AS, Kuchera C, Lee A, Winocur G. Olfactory-based social-learning varies as a function of parity in female rats. Psychobiology. 1994;22:37–43. [Google Scholar]
- Fudge MA, Kavaliers M, Ossenkopp K-P. Comparing the effects of selective estrogen receptor modulators tamoxifen and raloxifene on sucrose drinking and gustatory conditioning (CTA) in male rats. Abstr Soc Neurosci. 2006;661:27. [Google Scholar]
- Galef BG, Kennett DJ, Wigmore SW. Transfer of information concerning distant foods in rats—a robust phenomenon. Anim Learn Behav. 1984;12:292–296. [Google Scholar]
- Galef BG, Laland KN. Social learning in animals: empirical studies and theoretical models. Bioscience. 2005;55:489–499. [Google Scholar]
- Galef BG, Mason JR, Preti G, Bean NJ. Carbon-disulfide—a semiochemical mediating socially-induced diet choice in rats. Physiol Behav. 1988;42:119–124. doi: 10.1016/0031-9384(88)90285-5. [DOI] [PubMed] [Google Scholar]
- Galef BG, Stein M. Demonstrator influence on observer diet preference: analyses of critical social interactions and olfactory signals. Anim Learn Behav. 1985;13:31–38. [Google Scholar]
- Galef BG, Whiskin EE, Bielavska E. Interaction with demonstrator rats changes observer rats' affective responses to flavors. J Comp Psychol. 1997;111:393–398. doi: 10.1037/0735-7036.111.4.393. [DOI] [PubMed] [Google Scholar]
- Gariepy JL, Gendreau PL, Cairns RB, Lewis MH. D-1 dopamine receptors and the reversal of isolation-induced behaviors in mice. Behav Brain Res. 1998;95:103–111. doi: 10.1016/s0166-4328(97)00215-5. [DOI] [PubMed] [Google Scholar]
- Gendreau PL, Petitto JM, Gariepy JL, Lewis MH. D-1 dopamine receptor mediation of social and nonsocial emotional reactivity in mice: effects of housing and strain difference in motor activity. Behav Neurosci. 1997;111:424–434. doi: 10.1037//0735-7044.111.2.424. [DOI] [PubMed] [Google Scholar]
- Gendreau PL, Petitto JM, Petrova A, Gariepy JL, Lewis MH. D-3 and D-2 dopamine receptor agonists differentially modulate isolation-induced social-emotional reactivity in mice. Behav Brain Res. 2000;114:107–117. doi: 10.1016/s0166-4328(00)00193-5. [DOI] [PubMed] [Google Scholar]
- Gingrich B, Liu Y, Cascio C, Wang ZX, Insel TR. Dopamine D2 receptors in the nucleus accumbens are important for social attachment in female prairie voles (Microtus ochrogaster) Behav Neurosci. 2000;114:173–183. doi: 10.1037//0735-7044.114.1.173. [DOI] [PubMed] [Google Scholar]
- Hall H, Köhler C, Gawell L, Farde L, Sedvall G. Raclopride, a new selective ligand for the dopamine-D2 receptors. Prog Neuropsychopharmacol Biol Psychiatry. 1988;12:559–568. doi: 10.1016/0278-5846(88)90001-2. [DOI] [PubMed] [Google Scholar]
- Heyes CM. Social learning in animals: categories and mechanism. Biol Rev. 1994;69:207–231. doi: 10.1111/j.1469-185x.1994.tb01506.x. [DOI] [PubMed] [Google Scholar]
- Heyes CM, Durlach PJ. Social blockade of taste aversion learning is Norway rats (Rattus norvegicus): is it a social phenomenon. J Comp Psychol. 1990;104:82–87. doi: 10.1037/0735-7036.104.1.82. [DOI] [PubMed] [Google Scholar]
- Hyttel J. SCH 23390-The first selective dopamine D-1 antagonist. Eur J Pharmacol. 1983;91:153–154. doi: 10.1016/0014-2999(83)90381-3. [DOI] [PubMed] [Google Scholar]
- Karakaya S, Kipp M, Beyer C. Oestrogen regulates the expression and function of dopamine transporters in astrocytes of the nigrostriatal system. J Neuroendocrinol. 2007;19:682–690. doi: 10.1111/j.1365-2826.2007.01575.x. [DOI] [PubMed] [Google Scholar]
- Kruzich PJ, Grandy DK. Dopamine D-2 receptors mediate two-odor discrimination and reversal learning in C57BL/6. BMC Neurosci. 2004;5:12. doi: 10.1186/1471-2202-5-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapin IP. Only controls: effect of handling, sham injection, and intraperitoneal injection of saline on behavior of mice in an elevated plus-maze. J Pharmacol Toxicol Methods. 1995;34:73–77. doi: 10.1016/1056-8719(95)00025-d. [DOI] [PubMed] [Google Scholar]
- Lemon N, Manahan-Vaughan D. Dopamine D-1/D-5 receptors gate the acquisition of novel information through hippocampal long-term potentiation and long-term depression. J Neurosci. 2006;26:7723–7729. doi: 10.1523/JNEUROSCI.1454-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levitan RD, Masellis M, Lam RW, Kaplan AS, Davis C, Tharmalingam S, et al. A birth-season/DRD4 gene interaction predicts weight gain and obesity in women with seasonal affective disorder: a seasonal thrifty phenotype hypothesis. Neuropsychopharmacology. 2006;31:2498–2503. doi: 10.1038/sj.npp.1301121. [DOI] [PubMed] [Google Scholar]
- Li SM, Cullen WK, Anwyl R, Rowan MJ. Dopamine-dependent facilitation of LTP induction in hippocampal CA1 by exposure to spatial novelty. Nat Neurosci. 2003;6:526–531. doi: 10.1038/nn1049. [DOI] [PubMed] [Google Scholar]
- Liu B, Xie JX. Increased dopamine release in vivo by estradiol benzoate from the central amygdaloid nucleus of Parkinson's disease model rats. J Neurochem. 2004;90:654–658. doi: 10.1111/j.1471-4159.2004.02518.x. [DOI] [PubMed] [Google Scholar]
- McDermott JL, Liu BJ, Dluzen DE. Differences and effects of estrogen on dopamine and dopac release from the striatum of male and female CD-1 mice. Exp Neurol. 1994;125:306–311. doi: 10.1006/exnr.1994.1034. [DOI] [PubMed] [Google Scholar]
- McEwen BS, Wingfield JC. The concept of allostasis in biology and biomedicine. Horm Behav. 2003;43:2–15. doi: 10.1016/s0018-506x(02)00024-7. [DOI] [PubMed] [Google Scholar]
- Miczek KA, Weerts E, Haney M, Tidey J. Neurobiological mechanisms controlling aggression—preclinical developments for pharmacotherapeutic interventions. Neurosci Biobehav Rev. 1994;18:97–110. doi: 10.1016/0149-7634(94)90040-x. [DOI] [PubMed] [Google Scholar]
- Moncada D, Viola H. Induction of long-term memory by exposure to novelty requires protein synthesis: evidence for a behavioral tagging. J Neurosci. 2007;27:7476–7481. doi: 10.1523/JNEUROSCI.1083-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munger SD, Leinders-Zufall T, McDougall LM, Cockerham RE, Schmid A, Wandernoth P, et al. An olfactory subsystem that detets carbon disulfide and mediates food-related scial learning. Curr Biol. 2010;20:1438–1444. doi: 10.1016/j.cub.2010.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishi A, Snyder GL, Greengard P. Bidirectional regulation of DARPP-32 phosphorylation by dopamine. J Neurosci. 1997;17:8147–8155. doi: 10.1523/JNEUROSCI.17-21-08147.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otani S, Daniel H, Roisin MP, Crepel F. Dopaminergic modulation of long-term synaptic plasticity in rat prefrontal neurons. Cereb Cortex. 2003;13:1251–1256. doi: 10.1093/cercor/bhg092. [DOI] [PubMed] [Google Scholar]
- Pavlis M, Feretti C, Levy A, Gupta N, Linster C. (L)-DOPA improves odor discrimination learning in rats. Physiol Behav. 2006;87:109–113. doi: 10.1016/j.physbeh.2005.09.011. [DOI] [PubMed] [Google Scholar]
- Pecina S, Cagniard B, Berridge KC, Aldridge JW, Zhuang XX. Hyperdopaminergic mutant mice have higher ‘wanting' but not ‘liking' for sweet rewards. J Neurosci. 2003;23:9395–9402. doi: 10.1523/JNEUROSCI.23-28-09395.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters JR, Vallie B, Difronzo M, Donaldson ST. Role of dopamine D1 receptors in novelty seeking in adult female long-evans rats. Brain Res Bull. 2007;74:232–236. doi: 10.1016/j.brainresbull.2007.06.016. [DOI] [PubMed] [Google Scholar]
- Pfaff DW, Kieffer BL, Swanson LW. Mechanisms for the regulation of state changes in the central nervous system—an introduction. Ann N Y Acad Sci. 2008;1129:1–7. doi: 10.1196/annals.1417.031. [DOI] [PubMed] [Google Scholar]
- Puglisi-Allegra S, Cabib S. Pharmacological evidence for the role of D2 dopamine receptors in the defensive behavior of the mouse. Behav Neural Biol. 1988;50:98–111. doi: 10.1016/s0163-1047(88)90804-7. [DOI] [PubMed] [Google Scholar]
- Rada P, Avena NM, Hoebel BG. Daily bingeing on sugar repeatedly releases dopamine in the accumbans shell. Neuroscience. 2005;134:737–744. doi: 10.1016/j.neuroscience.2005.04.043. [DOI] [PubMed] [Google Scholar]
- Reddy DS, Kulkarni SK. Sex and estrous cycle-dependent changes in neurosteroid and benzodiazepine effects on food consumption and plus-maze learning behaviors in rats. Pharmacol Biochem Behav. 1999;62:53–60. doi: 10.1016/s0091-3057(98)00126-9. [DOI] [PubMed] [Google Scholar]
- Rodriguiz RM, Chu R, Caron MG, Wetsel WC. Aberrant responses in social interaction of dopamine transporter knockout mice. Behav Brain Res. 2004;148:185–198. doi: 10.1016/s0166-4328(03)00187-6. [DOI] [PubMed] [Google Scholar]
- Rusk IN, Cooper SJ. Parametric studies of selective D1 or D2 antagonists—effects on appetitive and feeding-behavior. Behav Pharmacol. 1994;5:615–622. doi: 10.1097/00008877-199410000-00007. [DOI] [PubMed] [Google Scholar]
- Rusk IN, Cooper SJ. Profile of the selective dopamine D-2 receptor agonist N-0437—its effects on palatability-induced and deprivation-induced feeding, and operant responding for food. Physiol Behav. 1988;44:545–553. doi: 10.1016/0031-9384(88)90317-4. [DOI] [PubMed] [Google Scholar]
- Rusk IN, Cooper SJ. The selective dopamine-D1 receptor agonist Sk-And-F 38393—its effects on palatability-induced and deprivation-induced feeding, and operant responding for food. Pharmacol Biochem Behav. 1989;34:17–22. doi: 10.1016/0091-3057(89)90346-8. [DOI] [PubMed] [Google Scholar]
- Salamone JD. Functions of mesolimbic dopamine: changing concepts and shifting paradigms. Psychopharmacology. 2007;191:389. doi: 10.1007/s00213-006-0623-9. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Cousins MS, Snyder BJ. Behavioral functions of nucleus accumbens dopamine: empirical and conceptual problems with the anhedonia hypothesis. Neurosci Biobehav Rev. 1997;21:341–359. doi: 10.1016/s0149-7634(96)00017-6. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Wisniecki A, Carlson BB, Correa M. Nucleus accumbens dopamine depletions make animals highly sensitive to high fixed ratio requirements but do not impair primary food reinforcement. Neuroscience. 2001;105:863–870. doi: 10.1016/s0306-4522(01)00249-4. [DOI] [PubMed] [Google Scholar]
- Sanchez-Andrade G, James BM, Kendrick KM. Neural encoding of olfactory recognition memory. J Reprod Dev. 2005;51:547–558. doi: 10.1262/jrd.17031. [DOI] [PubMed] [Google Scholar]
- Schultz W. Behavioral theories and the neurophysiology of reward. Annu Rev Psychol. 2006;57:87–115. doi: 10.1146/annurev.psych.56.091103.070229. [DOI] [PubMed] [Google Scholar]
- Shumaker SA, Legault C, Rapp SR, Thal L, Wallace RB, Ockene JK, et al. Estrogen plus progestin and the incidence of dementia and mild cognitive impairment in postmenopausal women—The Women's Health Initiative Memory Study: a randomized control trial. JAMA. 2003;289:2651–2662. doi: 10.1001/jama.289.20.2651. [DOI] [PubMed] [Google Scholar]
- Sills TL, Onalaja AO, Crawley JN. Mesolimbic dopaminergic mechanisms underlying individual differences in sugar consumption and amphetamine hyperlocomotion in Wistar rats. Eur J Neurosci. 1998;10:1895–1902. doi: 10.1046/j.1460-9568.1998.00201.x. [DOI] [PubMed] [Google Scholar]
- Smith DR, Striplin CD, Geller AM, Mailman RB, Drago J, Lawler CP, et al. Behavioural assessment of mice lacking D-1A dopamine receptors. Neuroscience. 1998;86:135–146. doi: 10.1016/s0306-4522(97)00608-8. [DOI] [PubMed] [Google Scholar]
- Smith-Roe SL, Kelley AE. Coincident activation of NMDA and dopamine D-1 receptors within the nucleus accumbens core is required for appetitive instrumental learning. J Neurosci. 2000;20:7737–7742. doi: 10.1523/JNEUROSCI.20-20-07737.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinert T, Wiebe C, Gebhardt RP. Aggressive behavior against self and others among first-admission patients with schizophrenia. Psychiatr Serv. 1999;50:85–90. doi: 10.1176/ps.50.1.85. [DOI] [PubMed] [Google Scholar]
- Svenningsson P, Nishi A, Fisone G, Girault JA, Nairn AC, Greengard P. DARPP-32: an integrator of neurotransmission. Annu Rev Pharmacol Toxicol. 2004;44:269–296. doi: 10.1146/annurev.pharmtox.44.101802.121415. [DOI] [PubMed] [Google Scholar]
- Szczypka MS, Rainey MA, Kim DS, Alaynick WA, Marck BT, Matsumoto AM, et al. Feeding behavior in dopamine-deficient mice. Proc Natl Acad Sci USA. 1999;96:12138–12143. doi: 10.1073/pnas.96.21.12138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson TL, Moss RL. Modulation of mesolimbic dopaminergic activity over the rat estrous cycle. Neurosci Lett. 1997;229:145–148. doi: 10.1016/s0304-3940(97)00450-3. [DOI] [PubMed] [Google Scholar]
- Touzani K, Bodnar R, Sclafani A. Activation of dopamine D1-like receptors in nucleus accumbens is critical for the acquisition, but not the expression, of nutrient-conditioned flavor preferences in rats. Eur J Neurosci. 2008;27:1525–1533. doi: 10.1111/j.1460-9568.2008.06127.x. [DOI] [PubMed] [Google Scholar]
- Valsecchi P, Choleris E, Moles A, Guo C, Mainardi M. Kinship and familiarity as factors affecting social transfer of food preferences in adult Mongolian gerbils (Meriones unguiculatus) J Comp Psychol. 1996;110:243–251. doi: 10.1037/0735-7036.110.3.243. [DOI] [PubMed] [Google Scholar]
- Valsecchi P, Galef BG. Social influences on the food preferences of house mice (Mus musculus) Intl J Comp Psychol. 1989;2:245–256. [Google Scholar]
- Viggiano D, Ruocco LA, Sadile AG. Dopamine phenotype and behaviour in animal models: in relation to attention deficit hyperactivity disorder. Neurosci Biobehav Rev. 2003;27:623–637. doi: 10.1016/j.neubiorev.2003.08.006. [DOI] [PubMed] [Google Scholar]
- Walmer DK, Wrona MA, Hughes CL, Nelson KG. Lactoferrin expression in the mouse reproductive-tract during the natural estrous cycle—correlation with circulating estradiol and progesterone. Endocrinology. 1992;131:1458–1466. doi: 10.1210/endo.131.3.1505477. [DOI] [PubMed] [Google Scholar]
- Wang ZX, Yu GZ, Cascio C, Liu Y, Gingrich B, Insel TR. Dopamine D2 receptor-mediated regulation of partner preferences in female prairie voles (Microtus ochrogaster): a mechanism for pair bonding. Behav Neurosci. 1999;113:602–611. doi: 10.1037//0735-7044.113.3.602. [DOI] [PubMed] [Google Scholar]
- Wise RA. Role of brain dopamine in food reward and reinforcement. Philos Trans R Soc Lond B Biol Sci. 2006;361:1149–1158. doi: 10.1098/rstb.2006.1854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright S, Klee P. Violent crime, aggression and amphetamine: what are the implications for drug treatment services. Drugs: Educ Prev Policy. 2001;8:73–90. [Google Scholar]
- Yu WZ, Silva RM, Sclafani A, Delamater AR, Bodnar RJ. Pharmacology of flavor preference conditioning in sham-feeding rats: effects of dopamine receptor antagonists. Pharmacol Biochem Behav. 2000a;65:635–647. doi: 10.1016/s0091-3057(99)00239-7. [DOI] [PubMed] [Google Scholar]
- Yu WZ, Silva RM, Sclafani A, Delamater AR, Bodnar RJ. Role of D(1) and D(2) dopamine receptors in the acquisition and expression of flavor-preference conditioning in sham-feeding rats. Pharmacol Biochem Behav. 2000b;67:537–544. doi: 10.1016/s0091-3057(00)00396-8. [DOI] [PubMed] [Google Scholar]
- Yue EL, Cleland TA, Pavlis M, Linster C. Opposing effects of D-1 and D-2 receptor activation on odor discrimination learning. Behav Neurosci. 2004;118:184–190. doi: 10.1037/0735-7044.118.1.184. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.