

Abstract
Current levels of endangerment and historical trends of species and habitats are the main criteria used to direct conservation efforts globally. Estimates of future declines, which might indicate different priorities than past declines, have been limited by the lack of appropriate data and models. Given that much of conservation is about anticipating and responding to future threats, our inability to look forward at a global scale has been a major constraint on effective action. Here, we assess the geography and extent of projected future changes in suitable habitat for terrestrial mammals within their present ranges. We used a global earth-system model, IMAGE, coupled with fine-scale habitat suitability models and parametrized according to four global scenarios of human development. We identified the most affected countries by 2050 for each scenario, assuming that no additional conservation actions other than those described in the scenarios take place. We found that, with some exceptions, most of the countries with the largest predicted losses of suitable habitat for mammals are in Africa and the Americas. African and North American countries were also predicted to host the most species with large proportional global declines. Most of the countries we identified as future hotspots of terrestrial mammal loss have little or no overlap with the present global conservation priorities, thus confirming the need for forward-looking analyses in conservation priority setting. The expected growth in human populations and consumption in hotspots of future mammal loss mean that local conservation actions such as protected areas might not be sufficient to mitigate losses. Other policies, directed towards the root causes of biodiversity loss, are required, both in Africa and other parts of the world.
Keywords: mammals, habitat loss, human development scenarios, Millennium Ecosystem Assessment, country prioritization
1. Introduction
Since the 1500s, 76 species and seven subspecies of mammals have gone extinct and another two are only extant in captivity. The hotspots of extinctions during this period have been Australia (because of direct killing, invasive rats, foxes, cats, habitat loss), the Caribbean (invasive rats and mongoose, direct killing) and South-Pacific islands (direct killing, invasive rats and snakes) [1]. Despite some conservation successes [2], most species are still declining, including a further 29 that may already be extinct such as the Christmas Island Pipistrelle Pipistrellus murrayi, the Kouprey Bos sauveli and the Baiji dolphin Lipotes vexillifer [3].
Twenty-five per cent (n = 1144) of all mammals for which there is sufficient information for an assessment of conservation status are threatened with extinction. The largest concentration of threatened terrestrial species is in South and Southeast Asia, the tropical Andes in South America, the Cameroonian Highlands and Albertine Rift in Africa, and the Western Ghats in India. All these regions combine high species richness, high numbers of range-restricted species [3] and high human pressure [4]. Threatened marine species are concentrated in the North Atlantic, the North Pacific and Southeast Asia, and these are also areas of concentration of range-restricted species [3] and high human impact [5].
Worldwide, the main threats to mammals are habitat loss and degradation (affecting 40% of all mammals) and harvesting (hunting or gathering for food, medicine and materials, affecting 19%). Among the drivers of habitat loss for mammals, agriculture and pastoralism are the most important, together affecting 40 per cent of terrestrial mammals (n = 2089) [1].
Recently, agriculture and grazing have expanded almost exclusively in the tropics [6]. Between 1980 and 2000, cattle pastureland increased by approximately 35 million ha in South America and approximately 7 million ha in Central America [7]. Cropland area increased by approximately 5 million ha in South America, further fragmenting and reducing the natural habitats of the Llanos of Venezuela, the Atlantic forest of Brazil, the Cerrado and the Amazon. In Southeast Asia, most agricultural expansion during the same period has been for tree plantations, which increased from roughly 11 to 17.4 million ha [7]. Oil palm Elaeis guineensis plantations increased tenfold from 0.2 to 2.7 million ha in Borneo alone. These plantations pose a serious threat to many endangered species such as the Bornean Orangutan Pongo pygmaeus, with remaining populations occurring mostly outside protected areas in lowland areas of high suitability for oil palm [1]. In the period 1980–2000, cropland area increased by approximately 50 and 25 per cent in East Africa and West Africa, respectively.
Globally, between 1995 and 2007, agricultural land increased by 400 million ha in developing countries but decreased by 412 million ha in developed countries [7]. The vast majority of this new agricultural land has come at the expense of native vegetation, particularly primary forest where clearing gives the added benefit of timber products [6]. Worryingly, this agricultural expansion is expected to continue in the future. Demand for agricultural products is predicted to increase by up to 50 per cent by 2050, with most expansion in tropical countries [7].
An assessment of the projected impacts of agricultural expansion on mammals is of utmost urgency to facilitate pre-emptive and effective conservation actions. Here, we estimate the impact of future scenarios of expanding agricultural land on the world's terrestrial mammals. We couple fine-scale, species-specific suitability models for terrestrial mammals with fine-scale projections of land use according to four global scenarios of socio-economic development. We highlight the countries in which the largest global losses of mammal distributions are predicted to occur between 2000 and 2050 and those countries predicted to host the species most in need of protection during this period. We define these countries as the future hotspots of global mammal loss (using loss of habitat as a proxy for species decline and potential extinction), recognizing that these future losses are likely to add to (rather than replace) those in areas currently concentrating high numbers of threatened species [3].
2. Methods
(a). Habitat suitability and land use change models
We projected the habitat suitability models described by Rondinini et al. [8] for 5086 species of terrestrial mammals onto four scenarios of human development from the Millennium Ecosystem Assessment [9]. Two of the scenarios, TechnoGarden and Adapting Mosaic, assume that countries generally take a proactive approach to environmental challenges, with environmental policies implemented to preserve ecosystem services and biodiversity. The other two scenarios, Order from Strength and Global Orchestration, assume that countries will generally react to environmental challenges, with policies implemented only when ecosystem degradation negatively affects human wellbeing. TechnoGarden and Global Orchestration envisage a world with global coordination of economic and environmental policies and sharing and advancement of ideas and technology. However, in the former, coordination emphasizes the environment, while in the latter, it emphasizes the economy. In contrast, Order from Strength envisages countries acting in isolation, trade barriers increasing and global institutions weakening or dissolving. Adapting Mosaic follows the same assumptions as Order from Strength initially, then converges towards TechnoGarden around the second half of the twenty-first century.
For each scenario, we obtained spatially explicit projections of agriculture and pasture land at 6′ resolution (approx. 10 km at the equator) globally at decadal intervals from 2000 to 2050 using the GLOBIO/HYDE land use change model [10,11]. We used the year 2000 as a baseline because data for 2010 were provided as projections for the Millennium Ecosystem Assessment scenarios. The scenarios are derived from quantitative, spatially explicit models of patterns and trends in human population growth, consumption, production and productivity at 30′ resolution from the integrated assessment model IMAGE [12] used for the Millennium Ecosystem Assessment [13]. For each decade, the GLOBIO/HYDE model uses the distribution and extent of crop and pasture in 18 macro-regions estimated by IMAGE to calculate the fraction of different land cover types (GLC2000; [14]) within 6′ cells using the algorithm described in Klein Goldewijk et al. ([15], see electronic supplementary material for details on the model).
We adapted the habitat suitability scores for terrestrial mammals from Rondinini et al. [8] based on the GLOBCOVER v. 2.1 classification [16] to the classification of the Global Land Cover 2000 model [14] used in the land use projections. Both legends are based on the Food and Agriculture Organization (FAO) Land Cover Classification System [17], which facilitated the building of a correspondence table (electronic supplementary material, table S1) based on descriptions of land cover classes. When one GLC2000 class corresponded to multiple GLOBCOVER classes, we averaged the suitability scores and rounded the value to the closest integer (0,1,2). We considered only GLC2000 cover types of high suitability for species (primary habitat for the species). We excluded medium suitability habitat (suitability score of 1), where the species can be found but not live permanently [8], to avoid overestimating loss of habitat to expanding land uses. We estimated the amount of suitable area for each species in each 6′ cell by multiplying the area occupied by land cover types suitable for the species (from Rondinini et al. [8]) by the proportion of the cell within the species' altitudinal range extracted from the International Union for Conservation of Nature (IUCN) database [1]. We used 1″ Shuttle Radar Topography Mission (SRTM) elevation to measure this proportion [18]. Suitable habitat and suitable elevation were spatially correlated, so we might have underestimated the amount of suitable habitat by multiplying these factors. However, this was the only feasible method because we had no spatially explicit data on land cover types within 6′ cells. For each species, at intervals of 10 years starting from 2000 (reference year) and ending in 2050, we calculated the total area of suitable habitat (in km2) within its range. We assumed species ranges to remain fixed until 2050, so assessed losses and gains of suitable habitat only within present ranges [1]. This might have underestimated both losses of habitat (range contractions) and gains (range expansions).
(b). Loss measures
For each scenario, we aggregated the measures of loss of species' habitat by country. We intersected our gridded projections of suitable habitat with the boundaries of 206 countries and overseas territories from the VMap0 data [19]. Countries and territories too small to overlay with our 6′ grid are in the electronic supplementary material, table S2. We then calculated three different measures of concern or priority for future mammal conservation: species richness weighted by global loss; species richness weighted by national contribution to global loss; and richness of species with large global declines.
(i). Species richness weighted by global loss
For each species we measured the fraction of the global range in 2000 predicted to be lost by 2050 (relative global loss). We used this as a species weighting and summed these weightings for all species predicted to lose habitat by 2050, and excluded species that gained habitat, to obtain a weighted richness of declining species for each country.
Countries have high values for this weighted richness if they are rich in species incurring large proportional losses of habitat within their global ranges, even if predicted to lose little or no habitat within the countries' borders. Because the measure involves species richness, it is sensitive to country size.
(ii). Species richness weighted by national contribution to global loss
We measured for each species and each country the fraction of the global loss of suitable habitat by 2050 occurring within the country's borders. We multiplied this fraction by the percentage global loss of the species to emphasize species predicted to be of future global concern. The resulting weighting for species s, ws was therefore
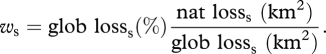 |
The resulting measure of weighted species richness indicated which countries contributed most to the global loss of suitable habitat for the species they host. In addition to species richness being correlated with size of country, the weighting itself is sensitive to country size because larger countries encompass larger proportions of the global ranges of many species (the fraction of national and global losses can approach or reach 1). Also for this measure, we excluded species predicted to gain habitat, as these are not of conservation concern.
(iii). Richness of species with large global declines
For each scenario, we also mapped the number of species in each country with large projected global declines (>30%). This threshold of loss followed criterion A3 of the IUCN Red List for declaring a species as Vulnerable [20]. However, according to this IUCN criterion, the projected future loss of habitat for a species must be expected to occur within 10 years or three generations from the time of listing, whichever is the longer. Our timeline of 50 years will therefore classify as vulnerable more species than the IUCN criterion, especially among short-lived mammals.
(c). Analyses
For each of the two measures of weighted species richness at the country level, we initially obtained four values, one for each scenario of global change. To assess the extent to which national or global losses varied across human development scenarios, we calculated for each country the variance across scenarios of each weighted richness measure. We report the top 10 countries with the largest variance values for both measures of weighted richness.
We also calculated a single value of each weighted richness measure for each country by averaging the country values across the four scenarios. This is mathematically equivalent to averaging the weights (species global and national losses) across scenarios and summing the mean weights within countries.
For our third measure, we aggregated the number of species with large projected global declines across the four scenarios in two ways. First, we created a worst-case outcome in which a species was accounted for if it was predicted to lose at least 30 per cent of its suitable habitat globally in any of the four scenarios. This involved the unrealistic assumption that the outcome for each species in each country will arise from the combination of all and only the negative attributes of each of the four scenarios. This would require habitat losses within single countries to result from land-use changes predicted in different scenarios that are mutually inconsistent, such as extensive pressure for both meat and vegetable production. This provided an upper bound on the number of species with large projected declines. For our second method of aggregation, we created a best-case outcome in which a species was accounted for only if it lost at least 30 per cent of its suitable habitat globally in all four scenarios. This provided a lower bound to the number of species with large projected declines. It carried the unrealistic assumption that the outcome for each species in each country will arise from the combination of all and only the positive attributes of each of the four scenarios. This is unrealistic because some factors positively affecting species persistence in one scenario can conflict with positive factors in other scenarios. For example, the increase in productivity predicted for some countries by TechnoGarden and Global Orchestration, arising from improved technology, is unlikely to be accompanied by low per capita consumption driven by the extreme poverty envisaged in the same countries by Order from Strength. To reiterate, while both the worst-case and the best-case outcomes are based on unrealistic assumptions, the rationale for them is to provide bounds around the number of species with large projected global declines rather than accurate predictions.
3. Results
(a). Richness weighted by global loss
The country with the highest richness weighted by projected range-wide losses is Mexico, followed by Democratic Republic of Congo, Tanzania, Kenya, South Africa, several other Sub-Saharan countries, Brazil and USA. These countries are not necessarily predicted to incur large losses of habitat for mammals because these losses could occur anywhere within the ranges of species they host. In fact, although most of the top 15 countries ranked by these measures are very large, in average across species predicted to have large global declines (more than 30% decline; n = 351), 69 per cent of the loss of habitat is predicted to occur outside the borders of each individual country.
Some African species with very large relative global losses across different scenarios are the Pardine Genet Genetta pardina (minimum and maximum losses between Millennium Ecosystem Assessment scenarios of 45–63%), Maxwell's Duiker Philantomba maxwelli (58–62%), Malawi Galago Galagoides nyasae (63–74% of its very restricted range lost), Southern Talapoin Monkey Miopithecus talapoin (49–60%), Pouched Gerbil Desmodilliscus braueri (89–97%) and Matthey's Mouse Mus matthei (82–90%). In North America, examples of species projected to have significant losses of habitat are the Mexican Spiny Pocket Mouse Liomys irroratus (in Mexico and USA, 41–87%), the San Cristobal Shrew Sorex stizodon (in Mexico only, 77–84%) and the Swift Fox Vulpes velox (in USA only, 37–53%).
Brazil does not have any species with notably high global losses, but this country hosts approximately 550 species that would lose some habitat globally (average projected change in habitat across the four scenarios)—more than any other country (electronic supplementary material, figure S1).
The 10 countries with the largest variance in weighted richness among scenarios are Democratic Republic of Congo, Cameroon, Mexico, Congo, Cote d'Ivoire, Guinea, Nigeria, Ghana Central African Republic and Liberia.
For African countries, Order from Strength predicts far worse habitat losses than the other three scenarios. Adapting Mosaic is the most favourable or perhaps more appropriately, the least worst scenario for African mammals, having lower but still significant losses of suitable habitat for most species. For Mexico, the best scenarios are TechnoGarden and Global Orchestration with very similar predicted losses, while the worst is Order from Strength.
(b). Richness weighted by national contribution to global loss
The countries with the largest richness weighted by national contribution to projected global loss are Mexico, Brazil, USA, Democratic Republic of Congo, Tanzania, Ethiopia, India, Angola, China and Madagascar. These are all large countries with high levels of endemism.
Examples of species with large national and global losses are the Tumbalà Climbing Rat Tylomys tumbalensis (24–53% of its global decline in Mexico), the Red-nosed Tree Rat Phyllomys brasiliensis (31–47% of global decline in Brazil) and the Angolan Long-Eared Bat (11–83% of global decline in Congo). In USA, large losses are predicted for two endemic canids, the Red Wolf Canis rufus (global decline of 10–51%) and the Swift Fox Vulpes velox (global decline of 42–53%), while in Tanzania, the Mountain Dwarf Galago, Galagoides orinus is predicted to lose 14–41% of suitable habitat. An example for India is the Kashmir Muskdeer, Moschus cupreus; losses in India contributed 31 per cent of the global decline of this endangered species, with overall global decline varying between scenarios from 37 to 56 per cent. Finally, the projected 42–46% global decline of habitat for the Mountain Nihala Tragelaphus buxtoni is predicted to occur completely within Ethiopia's borders.
The 10 countries with largest variance among scenarios of richness weighted by national contribution to global loss are Democratic Republic of Congo, Mexico, Cameroon, Brazil, United States, Nigeria, Liberia, Congo, Tanzania and Russia.
The best scenario for the USA is Global Orchestration and the worst is Order from Strength. For Brazil, the best is Adapting Mosaic and the worst is Order from Strength. For Russia, the best is Global Orchestration and the worst TechnoGarden.
(c). Richness of species with large global declines
In the worst-case outcome for species with large global declines (at least 30% of suitable habitat lost between 2000 and 2050 in at least one scenario), Democratic Republic of Congo takes the first place with 132 such species (figure 1a). Mexico is second with 103 species followed by Angola, Cameroon and Nigeria each with 100 species. In the best-case outcome for species with large global declines (at least 30% lost in all four scenarios; figure 1b), South Africa takes first place with 18 species, followed by USA with 11 and Namibia with 10. For the worst-case outcome, 28 countries have at least 50 species with large global declines and 63 have at least 10 species. For the best-case outcome, only three countries (USA, South Africa and Namibia) have at least 10 species with large global declines.
Figure 1.

Global patterns of projected mammal loss in relation to global Biodiversity Hotspots [21] (hatched). (a) Worst-case outcome for number of mammal species in each country with large projected global declines (losing at least 30% of suitable habitat globally by 2050 in any Millennium Ecosystem Assessment scenario). (b) Best-case outcome for number of mammal species in each country with large projected global declines (losing at least 30% of suitable habitat globally by 2050 in all four Millennium Ecosystem Assessment scenarios). (c) Mammal richness weighted by average global loss (GL) across the four scenarios. (d) Mammal richness weighted by average national contribution to global loss (NCGL) across the four scenarios. Legend categories use natural breaks adjusted to the closest integer with ArcGis 10 [22].
(d). Overlap between current global priorities and future hotspots of loss for mammals
There is little overlap between the regions predicted, according to any criteria, to be future hotspots for terrestrial mammal loss and the current global conservation priorities exemplified by the Biodiversity Hotspots [23] (figure 1a–d). Overlaps are confined to the Eastern Afromontane hotspot, the Brazilian Cerrado, the Madrean Pine-Oak woodlands in Mexico, the Cape Floristic Region in South Africa and the Western Ghats in India.
4. Discussion
(a). Patterns of global and national losses in relation to scenarios of the Millennium Ecosystem Assessment
Our models show that Mexico is the country with the highest weighted richness of declining terrestrial mammals, accounting for both global and national loss weightings. Mexico is also among the countries with the most species suffering large habitat declines by 2050. Large increases in food production and consumption are predicted in Mexico, especially from 2040, driven by accelerated growth of population and consumption. This is expected to require less land conversion in globalized scenarios than in regionalized ones because of the improved productivity in globalized scenarios from innovative agricultural practices and technological improvements [24].
Our models predict many African countries to rank among the top 10 in terms of national and global losses. Under Order from Strength, the African continent is expected to triple its 1995 population by 2050 [24]. Africa is also the only continent predicted by all scenarios to have a monotonic increase in human population until 2100 [24]. All scenarios predict economic improvement in Africa with steady increase in average income and household consumption. However, in Order from Strength, the increased consumption is predicted to outstrip productivity improvement and adoption of sustainable agricultural practices which will be hampered by low technology uptake, insufficient financial capital and limited attention to environmental issues [9]. These combined effects result in a predicted increase in grazing and cropping land of 71 and 56 per cent, respectively, across Africa in the Order from Strength scenario between 2000 and 2050, with consequent severe declines of mammals.
Brazil is the only South American country among the top 10 for any of our measures of mammal decline. Large expansions of cattle grazing, food crops and biofuel plantations in Brazil are predicted by the IMAGE set of models for all scenarios [24]. This agricultural expansion is predicted mainly in the Cerrado and the Atlantic forest, two ecoregions already severely impacted [21].
The USA is also among the countries with large global and national declines. In this country, regionalized scenarios predict increase in food crops and grazing areas to offset the reduced import of agricultural products. TechnoGarden, in contrast, predicts increases in biofuel plantations to become a key driver of habitat loss for mammals in the USA. The USA ranks seventh globally for number of endemic mammals, which explains its high values of richness weighted by national contribution to global loss.
Some countries show large variations in predicted habitat declines among scenarios, reflecting idiosyncratic effects in particular regions. TechnoGarden, for example, gives the worst projections of loss for Russia and other countries in central Asia. This scenario involves smaller reductions than others in food crop production in this region because of smaller population reductions in ex-USSR countries [24]. In addition, land-use models for TechnoGarden show meat consumption being replaced by vegetables and grazing land being replaced by cropland. This reduces the overall impact on mammals in areas with intensive man-made pastures or industrial livestock production, such as western Europe and the USA. However, TechnoGarden increases the pressure on mammals in central Asia where low-impact pastoralism on natural grasslands is expected to be replaced by cropland. Large losses of habitat under TechnoGarden are also driven by the projected expansion of biofuel plantations, particularly in the USA, central and Southeast Asia and South America.
Order from Strength has the most severe impacts on mammals in most countries. Mammals in developing countries are affected in this scenario by unchecked population growth and consumption, and by the dominance of economic security over biodiversity and ecosystem services [24]. In developed countries, the market fragmentation of Order from Strength expands food crops and pasture in regions, such as the USA and western Europe, where farmland would otherwise be abandoned [24].
(b). Our results relative to other global assessments
Previous studies have estimated the biodiversity impacts of human development scenarios. Jetz et al. [25] used the predictions of land use and land cover change (with climate change) for the Millennium Ecosystem Assessment scenarios at 30′ resolution to predict impacts on birds. With coarser resolution, there is higher potential for overestimates in changes of suitable habitat. This can artificially increase proportional losses for small-ranged species. Therefore, the coarser resolution of their study, their accounting for climate change (see §5) and their earlier baseline (1985 as opposed to 2000 in this study) all contribute to explaining their higher average estimates of loss per species by 2050 compared with our simulations (21–26% in Jetz et al. and 2.3–5.8% here). However, the spatial pattern of highest proportional losses is very similar, which is to be expected given the similar underlying data. In their study, however, the Himalayan region follows central Africa in terms of numbers of species losing large proportions of suitable habitat. In our study, Bhutan and Nepal do not rank among the countries with the most species having at least 30 per cent loss (figure 1). This is because the Himalaya is richer in range-restricted birds [26] than range-restricted mammals [3].
In another study, Giam et al. [27] ranked countries first by number of endemic plant species corrected by country area and then by the expected proportion of natural vegetation subject to land use or land cover change. They combined these rankings to measure future endangerment of plant species based on the assumption that endemics will be more threatened by future changes. Not surprisingly, countries with high plant endemism, such as Papua New Guinea, New Caledonia, Indonesia and Madagascar, figure prominently in their study but not in ours. Beyond differences in taxa, the different results depend also on our more direct measure of threat, based on spatially explicit and species-specific impacts instead of the intersection of country-level endemism and habitat loss. Our spatial explicitness is important because many endemic species might not be affected by loss of habitat and many non-endemic species might lose large amounts of habitat nationally and globally.
(c). Current and future international conservation priorities
Some existing conservation priorities such as Biodiversity Hotspots [21] and Crisis Ecoregions [28] have been based on rates of past conversion of natural habitat. However, consistent with other recent studies on other taxa [29], we show that predicted future hotspots of biodiversity loss according to the Millennium Ecosystem Assessment scenarios do not align with present hotspots (figure 1). This may be partially owing to the use of different taxa (i.e. plants rather than mammals, in the case of Biodiversity Hotspots), but it is also caused by a poor overlap between present and future projected patterns of habitat loss. This is reflected on the scarce concordance of areas rich in mammal species threatened by habitat loss now [3, fig. 2b] and in the future (this study). Therefore, reactive approaches to conservation—those focusing on regions with high past and present biodiversity loss—while important to prevent imminent extinctions, are unlikely to mitigate these projected losses. Additionally, many countries identified here as priorities for terrestrial mammals are poorly protected and poorly represented in other global conservation priority schemes [30], including the Global 200 Ecoregions [31], High-Biodiversity Wilderness Areas [23], the Last of the Wild [4] and Endemic Bird Areas [32]. We do not suggest that our rankings and maps should directly guide future conservation investments. Instead, we join Lee & Jetz [29] in recommending that projected future threats to biodiversity should be accounted for in conservation priorities.
Figure 2.

Scatter plot of countries in relation to our two weighted richness measures. The x-axis shows richness weighted by national contribution to global loss from 2000 to 2050, averaged across the four scenarios. The y-axis shows richness weighted by global loss from 2000 to 2050, averaged across the four scenarios. Country labels and names are in the electronic supplementary material.
Future threats can be accounted for in different ways. A risk-averse (proactive) strategy would protect globally imperilled species in countries with lower pressure on mammal habitat, thereby maximizing the likelihood of success (figure 2, top-left sector). A more risk-prone strategy would mitigate future losses in countries with high predicted losses of habitat and harbouring species with large expected global losses (figure 2, top-right sector). No single prioritization strategy will work well in all circumstances, and different contexts require different strategies. A mix of approaches is necessary because countries with many endemic species are towards the right-hand side of the graph and will not benefit from a risk-averse strategy. When options are available to protect species in countries with different levels of predicted loss, the choice between proactive or reactive interventions will depend also on socio-political factors such as existing conservation initiatives [33], costs of protection [34], investment opportunities [35], governance [36] and the kinds of threats faced by species (see last paragraph).
(d). National conservation priorities and reporting
Our measures have some affinity with the Red List Index (RLI) that has been proposed for monitoring trends of taxonomic groups globally or nationally [37,38] and adopted by the Convention on Biological Diversity as one measure to assess progress towards the 2010 targets [39]. The RLI is a compound measure synthesizing the genuine changes (those not resulting from improved knowledge or taxonomic changes) in the Red List status of all species in a taxon. A disadvantage of the RLI is that it can track only changes in species status large enough to trigger down-listing or up-listing. Our measures are instead continuous and can be complementary to the RLI. Being based on the global status of each species, the RLI is also geographically coarse. Fine-scale monitoring is possible through national RLIs, but these are only possible for the very few countries having red lists available at two points in time. The African continent, which figures prominently in our study, has the lowest number of national red lists in the world. Only 10 countries out of 53 have compiled red lists within the last 10 years for at least one taxon. Only three countries have a mammal red list [40].
We suggest that, by exploring a country's national loss measure and identifying which species most contribute to its score, it is possible to identify priority species and areas for conservation, monitoring and assessment. While we have summarized our results at the country level, the underlying analyses have a resolution of about 10 km and can be further improved by incorporating more ecological and socio-economic information (see below) to derive spatially explicit prioritization maps within countries.
5. Limitations
Although our approach has merits, our study also faced data limitations that call for refinements. Our study would have benefited from incorporating other threats to mammals such as direct killing and invasive species that are important in Asia [41], Australia and the Pacific [42]. Accounting for these factors might have changed the results proposed here, but we are not aware of any extensive projections of future patterns of these drivers of mammal decline. Moreover, countries in the Amazon and the Congo Basin, which we have highlighted here, have very high hunting pressures, second only to Southeast Asia [43]. They are likely to retain this primacy, given their increasing population densities in rural areas and their reliance on bush meat.
There are many uncertainties involved in projecting future global agricultural land cover, the major ones related to the assumptions about socio-political, economic, demographic and technological changes which are addressed by exploring multiple development scenarios. These scenarios are not meant to be accurate predictions of the future but rather explorations of the consequences of different development pathways. There are further uncertainties in the downscaling of the 30′ land-use change model. This process necessitated simplifications so that the criteria used to allocate regional conversion to crops and pasture locally were general enough to be valid globally. The model has been validated against the current global distribution of cropland and pasture, showing a good concordance [15], but its ability to predict land-use change has not been explored.
Our estimates of habitat loss are likely to be too small in some countries, because we did not incorporate projections of logging and other forestry activities for the four development scenarios. In IMAGE and GLOBIO, the changes in forestry are not spatially explicit, being more or less randomly applied within macro-regions, and were therefore not suitable for our analyses. This exclusion of forestry activities explains why countries like Indonesia, Malaysia and Papua New Guinea are among the top priorities for mammals in other studies [44] but are not highlighted here.
The mammal suitability models accurately predicted approximately 80 ± 16.8% of known species occurrences for a sample of species (n = 263), and reduced false presences compared with using the species range for 92 per cent of these species [8]. However, the model accuracy is unknown for most species, suggesting that our results need to be taken cautiously. In addition, because of the many-to-one relationship between GLOBCOVER classes and the GLC2000 classes used by GLOBIO, the original scores by Rondinini et al. were averaged in some instances. In 5 per cent of cases, the averaging involved different scores (e.g. suitable and unsuitable habitats). This happened mainly in the category ‘pasture and rangelands’, which is not present in GLOBCOVER or in GLC2000 but was introduced in the GLOBIO land-use change model. Future versions of these models will have to assign a specific suitability score to this land-use category.
We did not incorporate climate change effects on species distributions except for the modest indirect effects of climate on suitability for agriculture, which were modelled in IMAGE and reflected in the land-use change model used here (electronic supplementary material). At the time of writing, we did not have species-specific models of climate change impacts on mammals. The relative contribution and the synergistic effects of climate and land-use change on mammal distributions are of key importance in devising future conservation strategies [45] and will be the focus of our future research.
Finally, we did not account for isolation and fragmentation effects on mammals. Different spatial patterns of habitat loss and different histories of landscape conversion will have different impacts on biodiversity [46] and could potentially alter the ranking of countries presented here. However, these effects are landscape- and species-specific and cannot presently be incorporated into our analyses.
(a). Challenges in avoiding predicted losses
Technological improvements to increase productivity will be important but might not be sufficient to offset the increasing demand for agricultural products [13]. Estimates of future increases in productivity were part of the IMAGE scenarios. TechnoGarden involved optimistic assumptions about development and transfer of advanced agricultural technologies to developing countries. Yet, even in this scenario, African mammals pay a high cost for increased pastureland and cropland. Alternatively, in the fragmented world of Order from Strength, technological improvements are slow and technology transfer is limited, resulting in less land spared from production and worse outcomes for mammals. International trade and resource extraction will also determine outcomes for mammals. A global economy with little environmental responsibility, like the one assumed in Global Orchestration, envisages developing countries providing the labour and natural resources to shore up the prosperity of developed countries. This shift in agricultural land from developed to developing countries might come at a high environmental cost for developing regions, especially in the tropics (this study and [47]).
A policy of expanding protected areas will not avert the pressures on mammals from the growing demand for agricultural products, because protected areas often displace land-use change to unprotected areas [48]. Relief from pressures on natural habitat and species will rely mainly on structural changes in production and consumption [25,49]. Regional and global studies have demonstrated that protected areas are necessary but insufficient to prevent future loss of biodiversity in developing landscapes and regions [49,50]. Additionally, protected areas and other local conservation actions are at risk of failure in unstable and corrupt countries [35,51,52]. While socio-economic stability and corruption need to be taken into account for effective conservation of mammals [44,53], this does not imply that organizations should abandon these countries. On the contrary, conservation efforts need to be expanded in countries with the highest needs and the lowest means to undertake effective conservation [29]. We argue that conservation efforts should be integrated with development strategies. For this to happen, conservation strategies need to be applied with the support of local communities by promoting activities that address both development and biodiversity conservation goals such as certified community timber enterprises and nature-based tourism (reviewed in [54]). Conservation will succeed in the future battlegrounds of biodiversity loss only if there is a serious global effort to enforce compliance with environmental rules, promote the use of technological improvements to increase productivity, stabilize human populations, encourage responsible consumption patterns, reduce losses of agricultural products before consumption, improve forest management and limit the impacts of climate change.
Acknowledgements
The authors acknowledge Ana Rodrigues, Anni Arponen, April Reside, Tien Ming Lee, Tom Brewer, Jorge Alvarez-Romero, Natalie Ban, Tom Brooks and two anonymous reviewers for helpful comments to previous drafts of this manuscript. P.V. was supported by an IDP Australia postgraduate mobility scholarship and by a JCU postgraduate scholarship. R.L.P. was supported by the Australian Research Council.
References
- 1.IUCN 2010. IUCN Red List of threatened species, version 2010.4. See http://www.iucnredlist.org (accessed 25 November 2010)
- 2.Hoffmann M., et al. 2010. The impact of conservation on the status of the world's vertebrates. Science 330, 1503–1509 10.1126/science.1194442 (doi:10.1126/science.1194442) [DOI] [PubMed] [Google Scholar]
- 3.Schipper J., et al. 2008. The status of the world's land and marine mammals: diversity, threat, and knowledge. Science 322, 225–230 10.1126/science.1165115 (doi:10.1126/science.1165115) [DOI] [PubMed] [Google Scholar]
- 4.Sanderson E. W., Jaiteh M., Levy M. A., Redford K. H., Wannebo A. V., Woolmer G. 2002. The human footprint and the last of the wild. BioScience 52, 891–904 10.1641/0006-3568(2002)052[0891:THFATL]2.0.CO;2 (doi:10.1641/0006-3568(2002)052[0891:THFATL]2.0.CO;2) [DOI] [Google Scholar]
- 5.Halpern B., et al. 2008. A global map of human impact on marine ecosystems. Science 319, 948–952 10.1126/science.1149345 (doi:10.1126/science.1149345) [DOI] [PubMed] [Google Scholar]
- 6.Gibbs H., Ruesch A., Achard F., Clayton M., Holmgren P., Ramankutty N., Foley J. 2010. Tropical forests were the primary sources of new agricultural land in the 1980s and 1990s. Proc. Natl Acad. Sci. USA 107, 16 732–16 737 10.1073/pnas.0910275107 (doi:10.1073/pnas.0910275107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.FAOSTAT. 2009. FAO Statistical Databases 2009. See http://faostat.fao.org/
- 8.Rondinini C., et al. 2011. Global habitat suitability models of terrestrial mammals. Phil. Trans. R. Soc. B 366, 2633–2641 (doi:10.1098/rstb.2011.0113) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Millennium Ecosystem Assessment 2005. Ecosystems and human well-being: four scenarios, pp. 224–293 Washington, DC: Island Press [Google Scholar]
- 10.PBL Nederland Environmental Assessment Agency. 2010 GLOBIO 3.0. See www.globio.info. (accessed 1 July 2010)
- 11.Alkemade R., van Oorschot M., Miles L., Nellemann C., Bakkenes M., ten Brink B. 2009. GLOBIO3: a framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 12, 374–390 10.1007/s10021-009-9229-5 (doi:10.1007/s10021-009-9229-5) [DOI] [Google Scholar]
- 12.Bouwman A., Kram T., Klein Goldewijk K. 2007. Integrated modelling of global environmental change: an overview of IMAGE 2.4. Contract No.: Report no. 500110002 Netherlands Environmental Assessment Agency, Bilthoven, The Netherlands [Google Scholar]
- 13.Millennium Ecosystem Assessment 2005. Ecosystems and human well-being: synthesis. Washington, DC: Island Press [Google Scholar]
- 14.Bartholomé E., Belward A. 2005. GLC2000: a new approach to global land cover mapping from Earth observation data. Int. J. Remote Sens. 26, 1959–1977 10.1080/01431160412331291297 (doi:10.1080/01431160412331291297) [DOI] [Google Scholar]
- 15.Klein Goldewijk K., Van Drecht G., Bouwman A. F. 2007. Mapping contemporary global cropland and grassland distributions on a 5 × 5 minute resolution. J. Land Use Sci. 2, 167–190 10.1080/17474230701622940 (doi:10.1080/17474230701622940) [DOI] [Google Scholar]
- 16.European Space Agency 2008. GlobCover Land Cover v2 2008 database. Paris, France: European Space Agency GlobCover Project; See http://ionia1.esrin.esa.int/index.asp [Google Scholar]
- 17.di Greggio A., Jansen L. J. M. & cartographers 2000. Land Cover Classification System (LCCS): classification concepts and user manual. Rome, Italy: FAO; See http://www.fao.org/docrep/003/x0596e/x0596e00.HTM [Google Scholar]
- 18.United States Geological Survey 2006. Shuttle Radar Topography Mission 3 arc second version 2.0. See http://www.landcover.org/data/srtm (accessed 26 February 2010)
- 19.National Imagery and Mapping Agency 1997. Digital chart of the world (VMAP0). See http://gis-lab.info/qa/vmap0-eng.html (accessed 10 October 2010)
- 20.IUCN 2001. IUCN Red List Categories and Criteria, Version 3.1, p. ii + 30 pp Gland/Cambridge, Switzerland/UK: IUCN Species Survival Commission [Google Scholar]
- 21.Myers N., Mittermeier R. A., Mittermeier C. G., da Fonseca G. A. B., Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 10.1038/35002501 (doi:10.1038/35002501) [DOI] [PubMed] [Google Scholar]
- 22.ESRI (Environmental Systems Research Institute) 2008. ArcGIS 9.3.1. Redlands, CA: ESRI [Google Scholar]
- 23.Mittermeier R. A., Myers N., Thomsen J. B., da Fonseca G. A. B., Olivieri S. 1998. Biodiversity hotspots and major tropical wilderness areas: approaches to setting conservation priorities. Conserv. Biol. 12, 516–520 10.1046/j.1523-1739.1998.012003516.x (doi:10.1046/j.1523-1739.1998.012003516.x) [DOI] [Google Scholar]
- 24.Millennium Ecosystem Assessment 2005. Ecosystems and human well-being: changes in ecosystem services and their drivers across the scenarios, pp. 297–373 Washington, DC: Island Press [Google Scholar]
- 25.Jetz W., Wilcove D., Dobson A. 2007. Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol. 5, 1211–1219 10.1371/journal.pbio.0050157 (doi:10.1371/journal.pbio.0050157) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Grenyer R., et al. 2006. Global distribution and conservation of rare and threatened vertebrates. Nature 444, 93–96 See http://www.nature.com/nature/journal/v444/n7115/suppinfo/nature05237_S1.html 10.1038/nature05237 (doi:10.1038/nature05237) [DOI] [PubMed] [Google Scholar]
- 27.Giam X., Bradshaw C. J. A., Tan H. T. W., Sodhi N. S. 2010. Future habitat loss and the conservation of plant biodiversity. Biol. Conserv. 143, 1594–1602 10.1016/j.biocon.2010.04.019 (doi:10.1016/j.biocon.2010.04.019) [DOI] [Google Scholar]
- 28.Hoekstra J. M., Boucher T. M., Ricketts T. H., Roberts C. 2005. Confronting a biome crisis: global disparities of habitat loss and protection. Ecol. Lett. 8, 23–29 10.1111/j.1461-0248.2004.00686.x (doi:10.1111/j.1461-0248.2004.00686.x) [DOI] [Google Scholar]
- 29.Lee T., Jetz W. 2008. Future battlegrounds for conservation under global change. Proc. R. Soc. B 275, 1261–1270 10.1098/rspb.2007.1732 (doi:10.1098/rspb.2007.1732) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Brooks T. M., Mittermeier R. A., da Fonseca G. A. B., Gerlach J., Hoffmann M., Lamoreux J. F., Mittermeier C. G., Pilgrim J. D., Rodrigues A. S. L. 2006. Global biodiversity conservation priorities. Science 313, 58–61 10.1126/science.1127609 (doi:10.1126/science.1127609) [DOI] [PubMed] [Google Scholar]
- 31.Olson D. M., Dinerstein E. 1998. The Global 200: a representation approach to conserving the earth's most biologically valuable ecoregions. Conserv. Biol. 12, 502–515 10.1046/j.1523-1739.1998.012003502.x (doi:10.1046/j.1523-1739.1998.012003502.x) [DOI] [Google Scholar]
- 32.Stattersfield A., Crosby M., Long A., Wege D. 1998. Endemic bird areas of the world: priorities for biodiversity conservation (birdlife conservation series) (no. 7). Cambridge, UK: BirdLife International [Google Scholar]
- 33.Bode M., Probert W., Turner W. R., Wilson K. A., Venter O. 2010. Conservation planning with multiple organizations and objectives (Planificación de la Conservación con Múltiples Organizaciones y Objetivos). Conserv. Biol. 25, 295–304 10.1111/j.1523-1739.2010.01610.x (doi:10.1111/j.1523-1739.2010.01610.x) [DOI] [PubMed] [Google Scholar]
- 34.Naidoo R., Balmford A., Ferraro P. J., Polasky S., Ricketts T. H., Rouget M. 2006. Integrating economic costs into conservation planning. Trends Ecol. Evol. 21, 681–687 10.1016/j.tree.2006.10.003 (doi:10.1016/j.tree.2006.10.003) [DOI] [PubMed] [Google Scholar]
- 35.McBride M. F., Wilson K. A., Bode M., Possingham H. P. 2007. Incorporating the effects of socioeconomic uncertainty into priority setting for conservation investment. Conserv. Biol. 21, 1463–1474 10.1111/j.1523-1739.2007.00832.x (doi:10.1111/j.1523-1739.2007.00832.x) [DOI] [PubMed] [Google Scholar]
- 36.Smith R. J., Muir R. D. J., Walpole M. J., Balmford A., Leader-Williams N. 2003. Governance and the loss of biodiversity. Nature 426, 67–70 10.1038/nature02025 (doi:10.1038/nature02025) [DOI] [PubMed] [Google Scholar]
- 37.Butchart S., Stattersfield A., Bennun L., Shutes S., Akçakaya H., Baillie J., Stuart S., Hilton-Taylor C., Mace G. 2004. Measuring global trends in the status of biodiversity: Red List Indices for birds. PLoS Biol. 2, 2294–2304 10.1371/journal.pbio.0020383 (doi:10.1371/journal.pbio.0020383) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Butchart S. H. M., et al. 2007. Improvements to the Red List Index. PLoS ONE 2, e140. 10.1371/journal.pone.0000140 (doi:10.1371/journal.pone.0000140) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.United Nations Environment Programme. 2006. Decisions adopted by the Conference of Parties to the Convention on Biological Diversity at its eighth meeting (UNEP/CBD/COP/8/31/Annex 1), decision VIII/15. [Google Scholar]
- 40.Zamin T. J., Baillie J. E. M., Miller R. M., Rodríguez J. P., Ardid A. N. A., Collen B. E. N. 2010. National Red Listing beyond the 2010 target. Conserv. Biol. 24, 1012–1020 10.1111/j.1523-1739.2010.01492.x (doi:10.1111/j.1523-1739.2010.01492.x) [DOI] [PubMed] [Google Scholar]
- 41.Sodhi N. S., Koh L. P., Brook B. W., Ng P. K. L. 2004. Southeast Asian biodiversity: an impending disaster. Trends Ecol. Evol. 19, 654–660 10.1016/j.tree.2004.09.006 (doi:10.1016/j.tree.2004.09.006) [DOI] [PubMed] [Google Scholar]
- 42.Hoffmann M., Belant J. L., Chanson J. S., Cox N. A., Lamoreux J., Rodrigues A. S. L., Schipper J., Stuart S. N. 2011. The changing fates of the world's mammals. Phil. Trans. R. Soc. B 366, 2598–2610 (doi:10.1098/rstb.2011.0116) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fa J., Peres C., Meeuwig J. 2002. Bushmeat exploitation in tropical forests: an intercontinental comparison. Conserv. Biol. 16, 232–237 10.1046/j.1523-1739.2002.00275.x (doi:10.1046/j.1523-1739.2002.00275.x) [DOI] [PubMed] [Google Scholar]
- 44.Wilson K. A., Evans M. C., Di Marco M., Green D. C., Boitani L., Possingham H. P., Chiozza F., Rondinini C. 2011. Prioritizing conservation investments for mammal species globally. Phil. Trans. R. Soc. B 366, 2670–2680 10.1098/rstb.2011.0108 (doi:10.1098/rstb.2011.0108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Maiorano L., Falcucci A., Zimmermann N. E., Psomas A., Pottier J., Baisero D., Rondinini C., Guisan A., Boitani L. 2011. The future of terrestrial mammals in the Mediterranean basin under climate change. Phil. Trans. R. Soc. B 366, 2681–2692 10.1098/rstb.2011.0121 (10.1098/rstb.2011.0121) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Crooks K. R., Burdett C. L., Theobald D. M., Rondinini C., Boitani L. 2011. Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Phil. Trans. R. Soc. B 366, 2642–2651 10.1098/rstb.2011.0120 (doi:10.1098/rstb.2011.0120) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.McKinney L., Kick E., Fulkerson G. 2010. World system, anthropogenic, and ecological threats to bird and mammal species: a structural equation analysis of biodiversity loss. Org. Environ. 23, 3. 10.1177/1086026609358965 (doi:10.1177/1086026609358965) [DOI] [Google Scholar]
- 48.Ewers R. M., Rodrigues A. S. L. 2008. Estimates of reserve effectiveness are confounded by leakage. Trends Ecol. Evol. 23, 113–116 10.1016/j.tree.2007.11.008 (doi:10.1016/j.tree.2007.11.008) [DOI] [PubMed] [Google Scholar]
- 49.Alkemade R., et al. 2010. Rethinking global biodiversity strategies: exploring structural changes in production and consumption to reduce biodiversity loss. The Hague/Bilthoven, The Netherlands: Netherlands Environmental Assessment Agency (PBL) [Google Scholar]
- 50.Soares-Filho B., et al. 2006. Modelling conservation in the Amazon basin. Nature 440, 520–523 10.1038/nature04389 (doi:10.1038/nature04389) [DOI] [PubMed] [Google Scholar]
- 51.Laurance W. F. 2004. The perils of payoff: corruption as a threat to global biodiversity. Trends Ecol. Evol. 19, 399–401 10.1016/j.tree.2004.06.001 (doi:10.1016/j.tree.2004.06.001) [DOI] [PubMed] [Google Scholar]
- 52.Smith R. J., Walpole M. J. 2005. Should conservationists pay more attention to corruption? Oryx 39, 251–256 10.1017/S0030605305000608 (doi:10.1017/S0030605305000608) [DOI] [Google Scholar]
- 53.Eklund J., Arponen A., Visconti P., Cabeza M. 2011. Governance factors in the identification of global conservation priorities for mammals. Phil. Trans. R. Soc. B 366, 2661–2669 10.1098/rstb.2011.0114 (doi:10.1098/rstb.2011.0114) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.IIED, UNEP-WCMC, African Wildlife Foundation 2010. Linking biodiversity conservation and poverty alleviation: a state of knowledge review. CBD Technical series 55.