

Abstract
Background
The gene encoding the inorganic pyrophosphatase (PPase) of the intracellular pathogen Legionella pneumophila is induced during intracellular infection, but is constitutively expressed in Eschericia coli. The causative agent of tuberculosis, Mycobacterium tuberculosis, contains a well conserved copy of PPase. We sought to determine if expression of the M. tuberculosis PPase is regulated by the intracellular environment.
Results
A strain of Mycobacterium bovis bacille Calmette-Guérin (BCG) was constructed in which the Aequoria victoria green fluorescent protein (GFP) is controlled by the promoter of the M. tuberculosis ppa gene. After prolonged exposure of the recombinant BCG strain within murine bone-marrow-derived macrophages, there was no observed increased activity of the ppa promoter. Furthermore, there was no change in promoter activity after exposure to various stress stimuli such as reduced pH, osmotic shock, nutrient limitation or oxidative stress.
Conclusions
These results suggest that macrophage induction of ppa is not a general phenomenon among intracellular pathogens.
Background
Pathogenic bacteria such as Mycobacterium tuberculosis respond to conditions within the host by coordinate regulation of gene expression. The resulting production of specific gene products permits persistence and multiplication resulting in the manifestation of disease. Studies of facultative intracellular pathogens, who typically survive and replicate within phagocytic cells of the host anti-microbial defence system, indicate that the major genes expressed in vivo are in response to conditions encountered within the host cell [1]. These include genes required for the acquisition of metals, DNA repair, thermotolerance, osmotic tolerance and acid tolerance. Genes encoding proteins of an identical or similar function are often expressed in vivo by numerous bacteria, suggesting that responses to certain conditions are conserved among pathogens [1, 2].
The microbial inorganic pyrophosphatase (PPase) plays an important role in macromolecular biosynthesis and is essential for the viability of Escherichia coli and yeast [3, 4]. The protein appears to be ubiquitous and is highly conserved amongst differing species. In Legionella pneumophila, the ppa gene is induced during intracellular infection of U937 macrophage-like cells [5] and represents the first example of a regulated ppa gene in response to environmental stimuli. In order to determine if induction of ppa is a general phenomenon among intracelluar pathogens, we have analysed the activity of the M. tuberculosis ppa promoter upon exposure to the intracellular environment.
Results
Identification of a M. tuberculosis inorganic pyrophosphatase (PPase)
Analysis of the M. tuberculosis genome revealed an open reading frame (Rv3628) that was highly similar to L. pneumophila PPase [5]. The open reading frame consisted of 486 bp and was predicted to encode a protein that was 44% identical to L. pneumophila PPase [5] and 38% identical to E. coli PPase [6] (figure 1). Many of the residues that have been shown to be crucial for the structural and catalytic activity of the enzyme in E. coli are conserved in both M. tuberculosis and L. pneumophila [5, 7, 8, 9, 10, 11]. These data suggest that the product of Rv3628 is a PPase.
Figure 1.

Comparison of the PPase proteins of E. coli (E. coli), L. pneumophila (L. mon) and M. tuberculosis (M. tub). Identical amino acids shared between any 2 proteins are shaded.
The ppa promoter region was amplified from M. tuberculosis genomic DNA and cloned into the vector pJFX2 [12]. The resulting vector, pJIN6, contains the ppa promoter controlling a strongly fluorescent version of the Aequoria victoria green fluorescent protein (GFP) [13]. Transformation of Mycobacterium bovis bacille Calmette-Guérin (BCG) with pJIN6 resulted in fluorescent bacterial colonies as assessed by fluorescence microscopy (data not shown).
Activity of the M. tuberculosis ppa promoter within murine macrophages
In order to determine if the ppa promoter displayed enhanced activity within host cells, murine bone-marrow-derived macrophages were infected with BCG/pJIN6, and the level of fluorescence of recovered bacteria compared by flow cytometry after 0 or 6 days of infection. Bacteria displayed equivalent levels of fluorescence at both time points, suggesting that there was no increased activity of the ppa promoter after exposure to the intracellular environment (figure 2a). Analysis of GFP levels after 3 days of infection also showed no change in ppa promoter activity (data not shown). This was in contrast to BCG harbouring pJIN10, in which GFP is controlled by the M. tuberculosis sigE promoter, known to be active within macrophages [14]. BCG containing pJIN10 displayed approximately 2 times the level of fluorescence after 6 days of infection compared to the initial inoculum (figure 2b). Due to the relative stability of the GFP used in this study [15] it is unlikely that transient induction of the ppa promoter was not detected due to rapid breakdown of the reporter protein.
Figure 2.
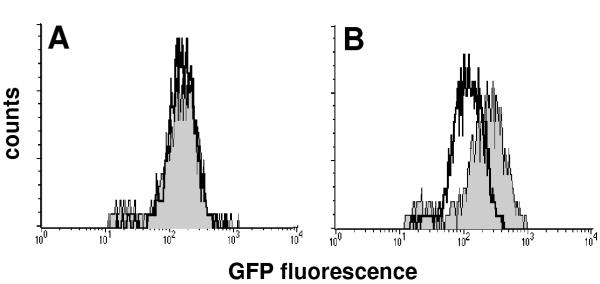
Activity of the M. tuberculosis ppa promoter in macrophages. Murine bone marrow-derived macrophages were infected with A) BCG/pJIN6 (gfp controlled by ppa promoter) or B) BCG/pJIN10 (gfp controlled by sigE promoter). After 6 days, macrophages were lysed and the fluorescence of recovered bacteria assessed by flow cytometry (grey histogram) and compared to the fluorescence of the initial inoculum (white histogram).
Effect of stress stimuli on M. tuberculosis ppa promoter activity
While the ppa promoter of L. pneumophila is induced within the intracellular environment, it is not induced after exposure to stress stimuli such as acid shock, osmotic shock or oxidative stress [5]. BCG/pJIN6 (ppa promoter) displayed no change in fluorescence after exposure to H2O2 (oxidative stress), high NaCl concentrations (osmotic shock), deprivation of iron by 2,2'-dipridyl treatment (nutrient limitation) or a reduction in pH (acid shock) (figure 3). The conditions used did permit analysis of in vitro stress as the sigE promoter of BCG/pJIN10 displayed increased activity due to acid shock, nutrient limitation and osmotic shock (figure 3). These data suggest that expression of the M. tuberculosis ppa is not regulated by the stress stimuli tested here.
Figure 3.
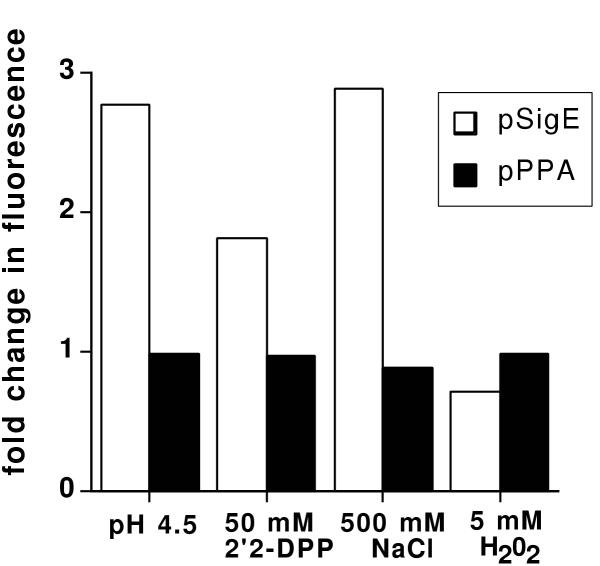
Effect of stress stimuli on M. tuberculosis ppa promoter activity. BCG harbouring pJIN6 (gfp controlled by ppa promoter; hatched bars) or pJIN10 (gfp controlled by sigE promoter; white bars) were grown for 24 hours in 7H9 medium of pH 4.5 or containing either 50 mM 2,2'-dipyridyl, 500 mM NaCl or 5 mM H2O2. The fold change in fluorescence represents the fluorescence value of bacteria after 24 hour exposure to stress conditions divided by the fluorescence level of the same bacteria grown in the absence of stress stimuli.
Discussion
Bacterial pathogens regulate gene expression in response to environmental stimuli encountered within the host and often contain related genes whose expression are influenced by the macrophage environment [1, 2]. In this report, we have investigated the intracellular expression of the M. tuberculosis ppa gene, whose counterpart in L. pneumophila is induced within the macrophage. While M. tuberculosis contains a well conserved homologue of the L. pneumophila PPase, we observed no increased activity of the M. tuberculosis ppa promoter within macrophages or by exposure to stress stimuli in vitro. This implies that the M. tuberculosis ppa promoter is not responsive to any specific intracellular triggers that influence the L. pneumophila counterpart. This may in part be due to differences in the intracellular microenvironment encountered by these different pathogens [16]. Alternatively, induction of the L. pneumophila promoter may occur independently of any intracellular signal and could be a by-product of an increased requirement for PPase in vivo, possibly a consequence of the greater growth rate of L. pneumophila within macrophages compared to in vitro-grown bacteria [17]. The results of this study suggest that the M. tuberculosis ppa is more characteristic of the constitutively expressed ppa of E. coli [18] rather than the in vivo-induced homologue of L. pneumophila, implying that induction of ppa does not appear to be an event common to intracellular pathogens.
Materials and Methods
Bacterial strains and growth conditions
Escherichia coli DH5α was grown routinely on liquid or solid Luria-Bertani medium. M. tuberculosis 103 (isolated directly from an tuberculosis patient; laboratory collection) and M. bovis BCG Pasteur were grown in liquid Middlebrook 7H9 medium (Difco laboratories, Detroit, USA) supplemented with ADC enrichment (Difco) or solid Middlebrook 7H10 medium (Difco) supplemented with OADC enrichment (Difco). When required, the antibiotic kanamycin was added at a concentration of 25 μg ml-1 for both E. coli and mycobacteria. For the analysis of stress induced responses, 7H9 medium was supplemented with either 500 mM NaCl (Sigma Chemical Co. St Louis, USA), 5 mM H2O2 (Sigma) or 50 mM 2,2'-dipyridyl (Aldrich Chemical Co. Milwauke, USA).
Vector construction
The ppa gene was located on the M. tuberculosis genome ([19] www.sanger.ac.uk/Projects/M_tuberculosis/) and 500 bp upstream of the initiation codon amplified from M. tuberculosis genomic DNA using the primers PPA.for (5'-GTCGGAGTACTAAACGCCGAAGCGT) and PPA.rev (5'-GAATTGGATCCGTCGGCTCCTTCAG). The product was cloned into the E. coli/mycobacterial shuttle vector pJFX2 [12] to yield plasmid pJIN6. The sigE promoter was amplified from the M. tuberculosis genome using primers SIGE.for (CCGCAAGTACTCGGCGACGTAATCT) and SIGE.rev (CGAGGGGATCCATGGGAATTACCGT) and cloned into pJFX2 to yield plasmid pJIN10. Preparation of competent cells and electroporation of mycobacteria was carried out as described previously [20].
Macrophage preparation and infection
Murine bone-marrow-derived macrophages were prepared as described previously [12]. Macrophage monolayers were infected with bacteria at a multiplicity of infection (MOI) of 1:1. After 4 hours of infection, extracellular bacteria were removed by washing 4 times with PBS, and incubation continued at 37°C in 5% CO2. After 6 days of infection, BCG infected-macrophages were washed 3 times in PBS, centrifuged and lysed in water plus 0.1% Tween 80. Recovered bacteria were analysed directly by flow cytometry. For the estimation of initial levels of fluorescence, bacteria were added to macrophage monolayers and immediately recovered and treated as above for flow cytometric analysis.
Flow cytometric analysis of fluorescent bacteria
Bacteria were analysed with a FACScan (Becton-Dickinson Immunocytometry Systems, Franklin Lake, USA). After growth in 7H9 media or macrophages, BCG cells were resuspended in 1 ml of PBS (approximately 1 x 106 bacteria), and a total of 2000 bacteria were analysed as a function of side scatter and GFP fluorescence. Quantitation of fluorescence levels was determined by the use of the Lysis II program (Becton-Dickinson).
Acknowledgments
Acknowledgments
We thank Danielle Ensergueix for macrophage preparation. J.A.T. was the recipient of an Institut Pasteur Cantarini Fellowship. This work was supported by the European Community (grant BMH4 CT92167) and a Pasteur Merieux Connaught Research and Development grant.
Contributor Information
James A Triccas, Email: J.Triccas@centenary.usyd.edu.au.
Brigitte Gicquel, Email: bgicquel@pasteur.fr.
References
- Heithoff DM, Conner CP, Mahan MJ. Dissecting the biology of a pathogen during infection. Trends Microbiol. 1997;5:509–513. doi: 10.1016/S0966-842X(97)01153-0. [DOI] [PubMed] [Google Scholar]
- Triccas JA, Gicquel B. Life on the inside: probing Mycobacterium tuberculosis gene expression during infection. Immunol Cell Biol. 2000;78:311–317. doi: 10.1046/j.1440-1711.2000.00934.x. [DOI] [PubMed] [Google Scholar]
- Chen J, Brevet A, Fromant M, Leveque F, Schmitter JM, Blanquet S, Plateau P. Pyrophosphatase is essential for growth of Escherichia coli. J Bacteriol. 1990;172:5686–5689. doi: 10.1128/jb.172.10.5686-5689.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundin M, Baltscheffsky H, Ronne H. Yeast PPA2 gene encodes a mitochondrial inorganic pyrophosphatase that is essential for mitochondrial function. J Biol Chem. 1991;266:12168–12172. [PubMed] [Google Scholar]
- Kwaik YA. Induced expression of the Legionella pneumophila gene encoding a 20-kilodalton protein during intracellular inection. Infect Immun. 1998;66:203–212. doi: 10.1128/iai.66.1.203-212.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lahti R, Pitkaranta T, Valve E, Ilta I, Kukko-Kalske E, Heinonen J. Cloning and characterization of the gene encoding inorganic pyrophosphatase of Escherichia coli K-12. J Bacteriol. 1988;170:5901–5907. doi: 10.1128/jb.170.12.5901-5907.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baykov AA, Dudarenkov VYW, Kapyla J, Salminen T, Hyytia T, Kasho VN, Husgafvel S, Cooperman BS, Goldman A, Lahti R. Dissociation of hexameric Escherichia coli inorganic pyrophosphatase into trimers on His-136-->Gln or His-140-->Gln substitution and its effect on enzyme catalytic properties. J Biol Chem. 1995;270:30804–30812. doi: 10.1074/jbc.270.51.30804. [DOI] [PubMed] [Google Scholar]
- Kankare J, Neal GS, Salminen T, Glumoff T, Glumhoff T, Cooperman BS, Lahti R, Goldman A. The structure of Escherichia coli soluble inorganic pyrophosphatase at 2.7 A. Protein Eng. 1994;7:823–830. doi: 10.1093/protein/7.7.823. [DOI] [PubMed] [Google Scholar]
- Kankare J, Salminen T, Lahti R, Cooperman BS, Baykov AA, Goldman A. Crystallographic identification of metal-binding sites in Escherichia coli inorganic pyrophosphatase. Biochemistry. 1996;35:4670–4677. doi: 10.1021/bi952637e. [DOI] [PubMed] [Google Scholar]
- Lahti R, Pohjanoksa K, Pitkaranta T, Heikinheimo P, Salminen T, Meyer P, Heinonen J. A site-directed mutagenesis study on Escherichia coli inorganic pyrophosphatase. Glutamic acid-98 and lysine-104 are important for structural integrity, whereas aspartic acids-97 and -102 are essential for catalytic activity. Biochemistry. 1990;29:5761–5766. doi: 10.1021/bi00476a017. [DOI] [PubMed] [Google Scholar]
- Lahti R, Salminen T, Latonen S, Heikinheimo P, Pohjanoksa K, Heinonen J. Genetic engineering of Escherichia coli inorganic pyrophosphatase. Tyr55 and Tyr141 are important for the structural integrity. Eur J Biochem. 1991;198:293–297. doi: 10.1111/j.1432-1033.1991.tb16015.x. [DOI] [PubMed] [Google Scholar]
- Triccas JA, Berthet FX, Pelicic V, Gicquel B. Use of fluorescence induction and sucrose counterselection to identify Mycobacterium tuberculosis genes expressed within host cells. Microbiology. 1999;145:2923–2930. doi: 10.1099/00221287-145-10-2923. [DOI] [PubMed] [Google Scholar]
- Cormack BP, Valdivia RH, Falkow S. FACS-optimized mutants of the green fluorescent protein (GFP). Gene. 1996;173:33–38. doi: 10.1016/0378-1119(95)00685-0. [DOI] [PubMed] [Google Scholar]
- Graham JE, Clark-Curtiss JE. Identification of Mycobacterium tuberculosis RNAs synthesized in response to phagocytosis by human macrophages by selective capture of transcribed sequences (SCOTS): Proc Natl Acad Sci USA. 1999;96:11554–11559. doi: 10.1073/pnas.96.20.11554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andersen JB, Sternberg C, Poulsen LK, Bjorn SP, Givskov M, Molin S. New unstable variants of green fluorescent protein for studies of transient gene expression in bacteria. Appl Environ Microbiol. 1998;64:2240–2246. doi: 10.1128/aem.64.6.2240-2246.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwak AK, Harb OS. Phenotypic modulation by intracellular bacterial pathogens. Electrophoresis. 1999;20:2248–2258. doi: 10.1002/(SICI)1522-2683(19990801)20:11<2248::AID-ELPS2248>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- Horwitz MA, Silverstein SC. Legionnaires' disease bacterium (Legionella pneumophila) multiplies intracellularly in human monocytes. J Clin Invest. 1980;66:441–450. doi: 10.1172/JCI109874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burton PM, Hall DC, Josse J. Constitutive inorganic pyrophosphatase of Escherichia coli. IV. Chemical studies of protein structure: J Biol Chem. 1970;245:4346–4352. [PubMed] [Google Scholar]
- Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, Gordon SV, Eiglmeier K, Gas S, Barry CE, et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature. 1998;393:537–544. doi: 10.1038/31159. [DOI] [PubMed] [Google Scholar]
- Pelicic V, Jackson M, Reyrat JM, Jacobs WR, Jr, Gicquel B, Guilhot C. Efficient allelic exchange and transposon mutagenesis in Mycobacterium tuberculosis. Proc Natl Acad Sci USA. 1997;94:10955–10960. doi: 10.1073/pnas.94.20.10955. [DOI] [PMC free article] [PubMed] [Google Scholar]