

Abstract
Background
Many bacteria swim by rotating helical flagellar filaments [1]. Waterbury et al. [15] discovered an exception, strains of the cyanobacterium Synechococcus that swim without flagella or visible changes in shape. Other species of cyanobacteria glide on surfaces [2,7]. The hypothesis that Synechococcus might swim using traveling surface waves [6,13] prompted this investigation.
Results
Using quick-freeze electron microscopy, we have identified a crystalline surface layer that encloses the outer membrane of the motile strain Synechococcus sp. WH8113, the components of which are arranged in a rhomboid lattice. Spicules emerge in profusion from the layer and extend up to 150 nm into the surrounding fluid. These spicules also send extensions inwards to the inner cell membrane where motility is powered by an ion-motive force [17].
Conclusion
The envelope structure of Synechococcus sp. WH8113 provides new constraints on its motile mechanism. The spicules are well positioned to transduce energy at the cell membrane into mechanical work at the cell surface. One model is that an unidentified motor embedded in the cell membrane utilizes the spicules as oars to generate a traveling wave external to the surface layer in the manner of ciliated eukaryotes.
Results and discussion
The cell envelope of Synechococcus sp. WH8113 has multiple layers (Fig. 1). Proceeding from inside, the cell envelope consists of cell membrane (∼ 10 nm thick), peptidoglycan layer (∼ 15 nm thick), outer membrane (∼ 10 nm thick), and surface layer (∼ 35 nm thick). The surface layer evident in Fig. 1 is not evident in conventional, chemically-fixed preparations (data not shown, see Ref. 16). Chemical fixation appears to cause detachment of the surface layer from the outer membrane whereas cryopreservation and freeze-substitution (data not shown) retain this layer.
Figure 1.

Synechococcus (strain WH8113) Cross fracture revealing concentric layers of cell envelope. The inset corresponds to the outlined section of cell envelope comprising cell membrane (CM), peptidoglycan layer (P), outer membrane (OM), and surface layer (S). A thylakoid layer (T) is also indicated. Scale bar, 200 nm.
Convex fractures of the inner cell membrane exposing its inner leaflet (Fig. 2a) as well as concave fractures exposing its outer leaflet (Fig. 2b) are densely covered by ball-shaped intramembrane particles that likely represent a panoply of membrane proteins and channels. These particles range in diameter from 5 nm to 13 nm. There is no obvious pattern in their distribution on either face, although a subset may represent the roots of the spicules described below. Pits complementary to the intramembrane particles on both convex and concave fractures of the cell membrane were infrequent [12].
Figure 2.

(a) Synechococcus (strain WH8113). Membrane fracture uncovering the inner cell membrane (CMi) showing a dense distribution of intramembrane particles. Fracture plane then crosses the surface layer (S) into the surrounding medium showing spicules (arrowhead) that extend from the cell surface. Arrow shows fiber extending from outer membrane to cell membrane. Scale bar, 100 nm. (b) Synechococcus (strain WH8113) Complementary fracture plane showing the outer leaflet of the cell membrane (CMo) which has fewer intramembrane particles than the inner leaflet. The fracture then crosses to the outer leaflet of the outer membrane (OMo), and then turns to fracture across the surface layer (S). Scale bar, 100 nm.
Convex fractures of the outer membrane exposing its inner leaflet (Fig. 3) are covered by pits. The pits range in diameter from 3 to 18 nm. The smaller pits occur in greater density than the larger pits. There is no obvious pattern in the distribution of pits. Concave fractures of the outer membrane exposing its outer leaflet appear rough and uneven, lacking clear structures like particles or pits (Fig. 2b).
Figure 3.

Synechococcus (strain WH8113). (OMi) and a patch of the inner leaflet of cell membrane (CMi) where the outer membrane has been pulled away. Typical of such outer membrane fractures, a profusion of spicules lie about the perimeter or lie flat along the outer membrane surface. Consideration of the fracture process explains the disposition of these spicules. If the spicules are tightly rooted to their bases, and the bond energy of their composition exceeds that of the interaction between their surfaces and the ambient ice, then the spicules would be pulled out of the frozen material that is cleaved away by the knife fracture, and would then fall onto the newly exposed surface thereby demonstrating their original continuity with it. Differences in angles of cleavage may explain the relative sparseness of the spicules on the top of the exposed surface compared to the edges. Scale bar, 100 nm.
A surface layer uniformly covers the outer membrane. In cross section, the surface layer appears as cylindrical pillars of diameter ∼ 8 nm that extend radially ∼ 18 nm from the outer leaflet of the outer membrane (Figs. 1, 2). The pillars are separated by gaps of ∼ 4 nm that are revealed by freeze-etch, indicating that they were formerly filled with water. Beyond ∼ 18 nm, the adjacent pillars connect laterally forming a fused region ∼ 17 nm thick. In cross sections of the cell envelope, this region sometimes appears as a congealed layer (see Fig. 1) but sometimes appears as a row of balls (see upper-left quadrant of Fig. 2a). This variability may reflect cleavages along different axes of the surface layer, or simply different degrees of plastic deformation and damage during fracture. The surface layer is a crystalline lattice, as is evident in aerial views. Such views reveal balls packed in a rhomboid organization with spacing ∼ 12 nm and obtuse angle ∼ 110° (Fig. 4a, 4b). These balls have the same size and spacing as those balls identified in cross-sections.
Figure 4.

(a) Synechococcus (strain WH8113) Crystalline outer surface of the surface layer (S) revealed where ambient ice is broken away. Arrowhead indicates fiber of 5 nm thickness that arises from cell membrane and extends to higher layers. Scale bar, 50 nm. (b) Synechococcus (strain WH8113) Patch of crystalline outer surface near fracture plane revealed by etching ambient ice. Since the bacterial surface is perpendicular to the line of sight, we used this image to measure the lattice arrangement: rhomboid with spacing ∼ 12 nm and obtuse angle ∼ 110°. Scale bar, 50 nm.
Spicules arise in profusion over the surface of the bacterium and extend up to 150 nm into the surrounding medium. The smallest observed separations between adjacent spicules are 12 nm and 24 nm, which corresponds to the spacing of the surface lattice. Spicules have uniform thickness of ∼ 5 nm along their length. Spicules are flexible; we have measured curvatures with radii as small as 30 nm.
It is difficult to trace the contour of a spicule through the entire cell envelope. Convex fractures of the outer membrane are characterized by a quantity of spicules projecting toward outer layers (Fig. 3). We frequently find spicules protruding outward between adjacent balls in cross-sections of the surface layer (arrow, Fig. 2a; arrowhead, Fig. 4a). We also frequently find fibers of 5 nm thickness that arise from the cell membrane before entering higher layers that could represent the lower stems of spicules (arrow, Fig 2a). These facts taken together suggest that the spicules extend outward through channels in the surface layer and extend inward through underlying layers to contact the cell membrane.
Pitta et al. [11] found that depriving Synechococcus of calcium inhibited motility and that the addition of calcium chelators EDTA or EGTA in excess of the initial concentration of calcium irreversibly arrested motility. We found that resuspension of Synechococcus in calcium-free seawater supplemented with 10 mM EDTA severely disrupted both the surface layer and internal thylakoid membranes. The surface layer lost its regularity, and would sometimes fall off entirely. Thylakoid layers that are ordinarily tightly apposed (see Fig. 1) separate by as much as 50 nm. Such severe damage (data not shown) could explain the abolition of motility by any of multiple causes, including the removal of the protein SwmA, an outer membrane protein which has been implicated in motility. Similar EDTA treatment is used to isolate SwmA from motile Synechococcus [3].
In the absence of flagella, the cell surface of Synechococcus sp. WH8113 must generate thrust. Theoretical work suggests that a cyanobacterium could swim at the observed speeds by oscillating its surface in the manner of a traveling wave [6,13]. Whereas motility is powered by an ion-motive force across the cell membrane [17] and the protein SwmA that is required for propulsion is associated with the outer membrane [3], the traveling wave must be formed at the very exterior of the cell body, outside the crystalline surface layer. Since the spicules contact the cell and outer membranes and protrude into the surrounding medium, they could be utilized to transmit motions in the cell membrane or outer membrane into the surrounding medium. If motors concealed in the cell membrane or outer membrane are capable of contracting and expanding in a regular manner (perhaps in analogy to the motor protein embedded in the basolateral membrane of cochlear outer hair cells that contract and expand to set the length of the cells [5]), these oscillations would be transduced into a rowing motion of the spicules. In this case, the crystalline surface layer could serve as oarlocks for the spicules, converting smaller motions at the bases of spicules into larger motions at the tips.
The motile mechanism of Synechococcus sp. WH8113 may be related to that of bacteria that glide over surfaces. Type IV pili (Tfp) have been implicated in the gliding movement of a species of the cyanobacterium Synechocystis [2], social gliding of Myxococcus, and twitching motility of Pseudomonas aeruginosa [14]. The spicules of Synechococcus sp. WH8113 could be related to Tfp, but comparison reveals differences. Tfp are generally much longer (1-2 μm), and stiffer than the spicules of Synechococcus. Tfp also tend to form bundles, playing roles in bacterial aggregation or attachment to surfaces. The spicules of Synechococcus do not tend to attach to one another, nor do cell bodies of Synechococcus tend to aggregate.
The motility apparatus of Synechococcus sp. WH8113 may be related to that of filamentous gliding cyanobacteria [6]. The cell envelopes of the gliding Oscillatoriaceae cyanobacterial species do have crystalline surface layers that resemble the outermost layer of Synechococcus, but the crystalline surface layer of Synechococcus is considerably thicker (∼ 35 nm thick compared to ∼ 20 nm thick). Structures like the spicules of Synechococcus have not been described in Oscillatoriaceae. Arrangements of helical fibers with diameters of 8-12 nm lie flat along the crystalline surface layer of Oscillatoriaceae. No such fibers are evident on Synechococcus. The surface of Oscillatoriaceae continuously extrudes slime while moving, whereas the surface of Synechococcus does not.
Synechococcus sp. WH8113 is but one of a large number of closely related species of Synechococcus, both motile and non-motile, that have been isolated. Comparative analysis of a selection of species could illuminate the motile mechanism of the cell envelope and the role of the structures described here.
Inactivation of the abundant outer membrane protein SwmA abolishes swimming motility [3]. However, when either motile wild-type or immotile SwmA cells collide with and become attached to a cover slip, cells will rotate about the point of attachment at rates ∼ 1 Hz [4]. This suggests that part, but not all, of the motility apparatus is disrupted in the absence of SwmA.
We uncovered no clear structural flaw in the mutant SwmA that would conclusively explain its motility phenotype or specify a role in motility for the structures described here. No images of spicules extending into the surrounding medium analogous to Fig. 2a have been obtained for SwmA. However, fibers crossing from the cell membrane to higher layers that we interpret as the lower stems of spicules are evident for SwmA, and spicules are evident on some convex fractures of the outer membrane of SwmA (data not shown). It remains possible that the cell envelope of the SwmA mutant has subtle flaws consistent with a partial disturbance of its motility apparatus.
Conclusions
The organelle that drives the motility of swimming cyanobacteria has not yet been identified. The spicules identified here could comprise part of the motility apparatus as they extend to both the site of energy transduction (the cell membrane) and the site of mechanical work (external to the surface layer). In any case, our elucidation of the surface structure (summarized in Fig. 5) provides new constraints on theoretical models for motility.
Figure 5.
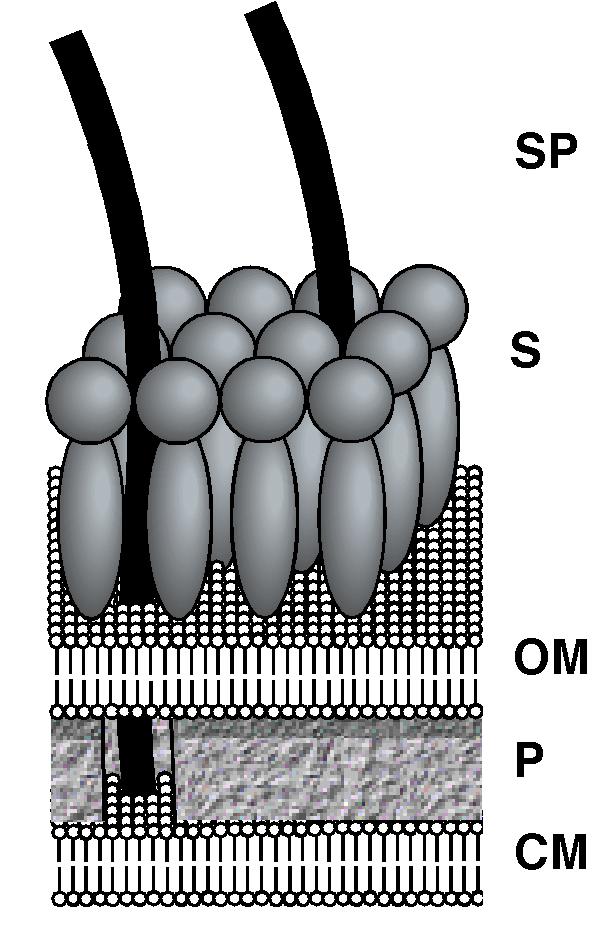
Diagram imaging the present knowledge of the cell envelope structure of Synechococcus. Present observations taken together suggest that the spicules (SP) extend through the surface layer (S) and outer membrane (OM) to contact the cell membrane (CM) (as shown in the cutaway of the peptidoglycan layer (P)).
Materials & Methods
Bacterial strains, culture conditions
The Synechococcus strain analyzed here was the motile wild-type strain WH8113 [15]. The laboratory of J. Waterbury provided the cultures of WH8113. Culture conditions are described elsewhere [16,17].
Sample preparation and viewing
The quick-freeze method used here is described elsewhere [8,9]. In summary, a few microliters of cell suspension were spread on a layer of gelatin mounted on a freezing stage that was then slammed against a super-cooled copper block. For freeze-fracture/freeze-etch preparations, quick-frozen specimens were then knife fractured at nominally -108°C in a Balzer's 301 freeze-fracture apparatus equipped with a high-speed rotary stage. Fractures were within a few micrometers of the frozen surface. The fractured samples were lightly etched for 4 min at nominally -108°C. The high concentration of salt in the artificial sea water used as culture medium precluded deeper etching. The etched surface was rotary shadowed with platinum, and then rotary coated with a carbon backing. Replicas were cleaned in commercial bleach, rinsed in distilled water, and picked up on 400-mesh grids.
For freeze-substitution, quick-frozen specimens were freeze substituted in 4% OsO4 in acetone at -80°C using standard procedures [10]. Additional staining was provided by immersion in 0.1% HfCl in acetone for 4 hr. Embedding was in araldite and sections were stained with uranyl acetate and lead citrate.
Samples were viewed in a JEOL 200CX electron microscope. Regions of the specimen with negligible ice crystal damage were chosen for analysis. Electron micrographs were scanned directly using an Agfa DuoScan scanner, and images were analyzed using stereo images generated in Adobe Photoshop.
Acknowledgments
Acknowledgements
We are indebted to J. Waterbury, F. Valois, and B. Brahamsha for kindly providing bacterial cultures and for helpful discussions. ADTS was supported by the Rowland Institute for Science and is an Amgen Fellow of the Life Sciences Research Foundation. ADTS dedicates his share of this work to his graduate advisor, Howard C. Berg.
Contributor Information
Aravinthan DT Samuel, Email: asamuel@fas.harvard.edu.
Jennifer D Petersen, Email: petersej@ninds.nih.gov.
Thomas S Reese, Email: tsr@codon.nih.gov.
References
- Berg HC, Anderson RA. Bacteria swim by rotating their flagellar filaments. Nature (London) 1973;245:380–384. doi: 10.1038/245380a0. [DOI] [PubMed] [Google Scholar]
- Bhaya D, Watanabe N, Ogawa T, Grossman AR. The role of an alternative sigma factor in motility and pilus formation in the cyanobacterium Synchecocystis sp. strain PCC6803. Proc Natl Acad Sci USA. 1999;96:3188–3193. doi: 10.1073/pnas.96.6.3188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brahamsha B. An abundant cell-surface polypeptide is required for swimming by the nonflagellated marine cyanobacterium Synechococcus . Proc Natl Acad Sci USA. 1996;93:6504–6509. doi: 10.1073/pnas.93.13.6504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brahamsha B. Non-flagellar swimming in marine Synechococcus. J Molec Microbiol Biotechnol. 1999;1:59–62. [PubMed] [Google Scholar]
- Dallos P, Evans BN. High-frequency motility of outer hair cells and the cochlear amplifier. Science. 1995;267:2006–2009. doi: 10.1126/science.7701325. [DOI] [PubMed] [Google Scholar]
- Ehlers KM, Samuel ADT, Berg HC, Montgomery R. Do cyanobacteria swim using traveling surface waves? Proc Natl Acad Sci USA. 1996;93:8340–8343. doi: 10.1073/pnas.93.16.8340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoiczyk E, Baumeister W. Envelope structure of four gliding filamentous cyanobacteria. J Bacteriol. 1995;177:2387–2395. doi: 10.1128/jb.177.9.2387-2395.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khan S, Dapice M, Reese TS. Effects of mot gene expression on the structure of the flagellar motor. J Mol Biol. 1988;202:575–584. doi: 10.1016/0022-2836(88)90287-2. [DOI] [PubMed] [Google Scholar]
- Khan S, Khan IH, Reese TS. New structural features of the flagellar base in Salmonella typhimurium revealed by rapid-freeze electron microscopy. J Bacteriol. 1991;173:2888–2896. doi: 10.1128/jb.173.9.2888-2896.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ornberg RL, Reese TS. Beginning of exocytosis captured by rapid-freezing of Limulus amebocytes. J Cell Biol. 1981;90:40–54. doi: 10.1083/jcb.90.1.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitta TP, Sherwood EE, Kobel AM, Berg HC. Calcium is required for swimming by the nonflagellated cyanobacterium Synechococcus strain WH8113. J Bacteriol. 1997;179:2524–2528. doi: 10.1128/jb.179.8.2524-2528.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rash JE, Giddings FD. Counting and measuring IMPs and pits: why accurate counts are exceedingly rare. J Electron Microsc Techn. 1989;13:204–215. doi: 10.1002/jemt.1060130307. [DOI] [PubMed] [Google Scholar]
- Stone HA, Samuel ADT. Propulsion of microorganisms by surface distortions. Phys Rev Lett. 1996;77:4102–4104. doi: 10.1103/PhysRevLett.77.4102. [DOI] [PubMed] [Google Scholar]
- Wall D, Kaiser D. Type IV pili and cell motility. Mol Microbiol. 1999;32:1–10. doi: 10.1046/j.1365-2958.1999.01339.x. [DOI] [PubMed] [Google Scholar]
- Waterbury JB, Willey JM, Franks DG, Valois FW, Watson SW. A cyanobacterium capable of swimming motility. Science. 1985;230:74–76. doi: 10.1126/science.230.4721.74. [DOI] [PubMed] [Google Scholar]
- Willey JM. Ph.D. Thesis. Massachusetts Institute of Technology and Woods Hole Oceanographic Institution. Characterization of swimming motility in a marine unicellular cyanobacterium. 1988.
- Willey JM, Waterbury JB, Greenberg EP. Sodium-coupled motility in a swimming cyanobacterium. J Bacteriol. 1987;169:3429–3434. doi: 10.1128/jb.169.8.3429-3434.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]