

Research highlights
▸ Social behavior becomes more flexible across development ▸ The orbitofrontal and ventrolateral prefrontal cortices contribute to social flexibility ▸ The maturation of the ventral prefrontal cortex enables inhibitory control, complex rule use, and computation of value.
Keywords: Childhood, Adolescence, Emotion, Affiliative
Abstract
Over the last several years a number of studies in both humans and animals have suggested that the orbitofrontal and ventrolateral prefrontal cortices play an important role in generating flexible behavior. We suggest that input from these brain regions contribute to three functions involved in generating flexible behavior within social contexts: valuation, inhibition, and rule use. Recent studies have also demonstrated that the prefrontal cortex undergoes a prolonged course of maturation that extends well after puberty. Here, we review evidence that the prolonged development of these prefrontal regions parallels a slowly emerging ability for flexible social behavior. We also speculate on the possibility that sensitive periods for organizing social behavior may be embedded within this developmental time-fame. Finally, we discuss the role of prefrontal cortex in adolescent mood and anxiety disorders, particularly as orbitofrontal and ventrolateral prefrontal cortices are engaged in a social context.
1. Introduction
One of the most challenging calculations that social animals have to make is how (or whether) to interact with another individual. In contrast to many inanimate objects, whose properties and value are relatively constant across all representations of the class (e.g., a banana is a banana), the value of social stimuli are orders of magnitude more variable from one individual to the next. This value depends on many factors that are intrinsic to the individual. For example, factors such as relative dominance status, the group or subgroup to which an individual belongs, and past experience can all critically impact the relative value attributed to another person (or animal). Furthermore, even within an organism, social signals are constantly changing. For example, the same individual can convey both solicitous and threatening signals, can provide access to different resources at different points in time, and can be in various phases of the sexual reproductive cycle. In addition, all of the information about the relative value of another person must be integrated with the internal states of the evaluating individual in generating social behaviors (e.g., engaging someone in a conversation, asking someone out on a date), which are also under a constant state of flux. Finally, if a decision is made to engage with another organism, the manner in which the other individual is best approached may also vary depending on a number of factors such as dominance, gender, and context. Thus, given that social contexts and internal states are ever changing, dynamic, and complex for organisms like humans, a great deal of flexibility is required for engaging in competent social behavior.
In this review, we argue that flexibility in social behavior is a critical component of social interaction, and like other forms of cognitive flexibility, this is largely carried out by circuits located in prefrontal cortex (PFC). While the PFC mediates flexible behavior in a variety of domains besides social interaction, flexibility is a particularly important aspect of smooth and competent social behavior; thus, the social domain is the primary focus of this review. Social flexibility differs mainly from other forms of flexibility (such as motor inhibition) not in the PFC per se, but in the regions with which PFC interfaces. As we propose in greater detail below, social behavior is the culmination of input from dedicated sensory and perceptual systems (e.g., fusiform face area and superior temporal sulcus) that is integrated with input from more general regions dedicated to affect (e.g., amygdala and ventral striatum), and executive function (e.g., prefrontal cortex) (Nelson et al., 2005). Here, we focus specifically on three executive functions that have been attributed to the ventral prefrontal cortex (VPFC): valuation; inhibition; and rule acquisition. We targeted these specific functions based on findings from behavioral, lesion, and neuroimaging studies that implicate these functions with VPFC engagement in social contexts.
As such, the purpose for this review is threefold. First, we aim to provide a brief overview of the literature which relates valuation, inhibition, and rule use to the VPFC; second, we seek to demonstrate how these relatively abstract executive functions might participate in social flexibility; and third, we propose that the developmental course of these functions influences the maturation of flexible social engagement. Our ultimate goal is to provide a framework for the interpretation and prediction of neurobiological and behavioral manifestations of the maturation of social behavior. Finally, we end with a discussion of the role of VPFC engagement in adolescent mood and anxiety disorders, particularly in response to social context.
While some conditions involving limited flexibility in social-emotional functioning, such as autism, are clearly relevant to the construct of social flexibility, they will not be a focus of this review. Certainly, a number of cognitive functions likely underlie social flexibility such as perceiving social others, having theory of mind, being able to self reference, and monitoring expectations. These aspects of flexibility implicate a distributed network of regions outside of the VPFC, including the paracingulate cortex, dorsal medial prefrontal cortex, dorsal anterior cingulate cortex, superior temporal sulcus, tempo-parietal junction, and temporal poles. While we acknowledge the importance of these neural networks for social flexibility, we do not focus on these specific cognitive processes in this review. Rather, we concentrate here on the ventral regions of the PFC and distinguish three cognitive regulatory processes – valuation, inhibition, and rule use – that are linked to emotion in social settings. Readers are referred elsewhere for excellent empirical evidence and theoretical discussions of the role of the dorsal and medial PFC regions in developmental social cognitive neuroscience writings (Amodio and Frith, 2006, Blakemore, 2008, Gallagher and Frith, 2003, Moriguchi et al., 2007, Pelphrey et al., 2004, Pfeifer et al., 2007, Wang et al., 2006).
2. Anatomical portrait of the VPFC
Within the VPFC, the orbitofrontal cortex (OFC) occupies the ventral floor of the prefrontal cortex and receives input from the mediodorsal thalamus among many other regions (Kringelbach and Rolls, 2004, Price, 2007). Based on detailed anatomical studies of both human and nonhuman primates, Price has broadly construed the OFC as comprising two distinct functional networks, the orbital and the medial networks. The orbital network is primarily located in the lateral and caudal areas of the OFC, whereas the medial network is mainly situated in the medial portion of the OFC. The lateral OFC network as designated by Price is densely innervated by sensory systems, whereas the medial network is interconnected with limbic and other emotion-related regions such as amygdala, ventral striatum, anterior cingulate and periaqueductal gray (Barbas, 2007, Carmichael and Price, 1994, Kringelbach and Rolls, 2004, Price, 2007). Although Price considers the limbic network part of the medial OFC, this region also includes extreme lateral and caudal regions such as Brodmann area 47 (and Walkers area 12 in the monkey) in this designation. Others have restricted the designation of OFC to medial regions and excluded the caudolateral region (Blair, 2007, Izquierdo et al., 2005, Murray et al., 2007). We refer here to the OFC as including the medial portion of the OFC. We also subsume what Price refers to as lateral or sensory OFC (including BA 47) as part of the ventrolateral prefrontal cortex (vlPFC), which includes both the lateral portions of the orbital gyrus and the inferior frontal gyrus. Like the OFC, the vlPFC is bi-directionally innervated by limbic structures such as the amygdala, but also has strong connections to other areas such as the inferior temporal cortex, hippocampal complex, and dorsolateral PFC (Petrides, 2005, Sakagami and Pan, 2007). We will consider both OFC and vlPFC together as the VPFC (Fig. 1).
Fig. 1.
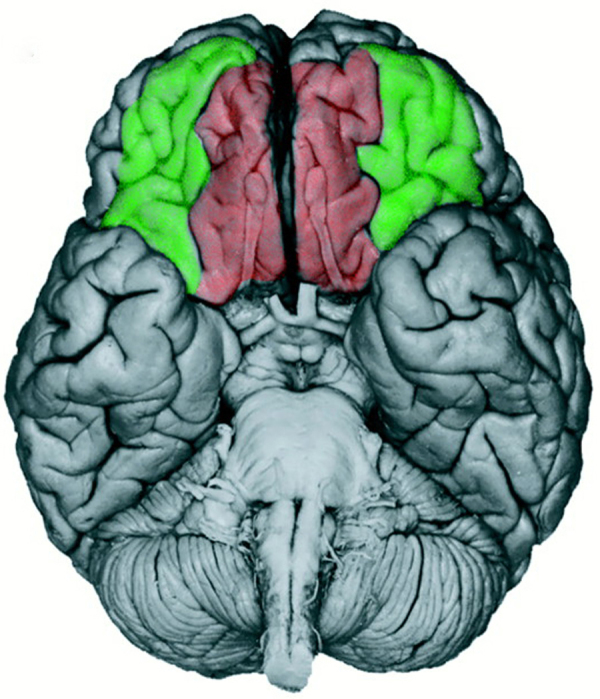
Schematic depiction of the ventrolateral prefrontal cortex (vlPFC in green) and orbitofrontal cortex (OFC in red) regions of the VPFC that contribute to the development of social flexibility. Inhibitory control and rule use are controlled by the vlPFC, whereas computation of expected value of social stimuli is performed by the OFC. All three of these functions mature slowly across development.
From Davidson et al. (2000). Reprinted with permission from AAAS.
3. Linking VPFC to social flexibility
After a long period of neglect, the functional role of the VPFC has received a fair amount of attention in recent years (Dolan, 2007, Murray et al., 2007). While the VPFC has a well-established role in integrating response to emotional stimuli with behavior, the exact role the VPFC plays in this integrative process is still a matter of some debate. We highlight here the main functions of the OFC and vlPFC within the VPFC that may be most relevant to guiding flexibility in social behaviors.
Both the OFC and vlPFC play a critical role in the generation of appropriately flexible patterns of behavior required to display fluid and competent social interaction under varying conditions. This is particularly the case for the complex social behavior of humans and many nonhuman primates (Brothers et al., 1990, Dunbar and Shultz, 2007). We propose that there are three specific aspects of social flexibility that are subserved by the VPFC, including: (1) emotional value computation; (2) inhibitory control of behavior; and (3) acquisition and generation of rule use under different contexts. By emotional value computation, we refer to the process of generating an expectation of the emotional reward or punishment that will be produced by a specific stimulus. This expectation is generated from both past experience and the inherent biological value that has been imbued on a stimulus (e.g., food and reproduction) by evolution (Murray and Wise, 2010). Since the expected emotional values of stimuli are constantly changing, valuation is constantly being computed and updated. By inhibitory control of behavior, we refer to the capacity to inhibit response tendencies that are irrelevant or in direct contrast to one's larger goals. In the course of engaging with the world, many salient and provocative stimuli are encountered that are either a distraction or potentially even counter-productive to a larger ultimate goal. Inhibitory control of behavior is the process of repressing these tendencies (Casey et al., 2008, Dillon and Pizzagalli, 2007). By acquisition and generation of rule use, we refer to learning the contingencies of behavior in different contexts. As discussed in the opening paragraph, social stimuli and social contexts are quite varied and each context can have a unique set of behavioral contingencies (Bunge, 2004).
The OFC appears to be particularly important in generating current value computations while the vlPFC is likely to be important for inhibitory control and rule acquisition. This dissociation is derived both from differential patterns of activation observed in functional neuroimaging tasks and from behavioral dissociations from lesion studies. For example, in a recent study, rhesus monkeys with orbital PFC lesions showed impairments in modulating their behavior as reward value fluctuated, but monkeys with lateral PFC lesions only displayed aberrant behavior if the fluctuating reward size was combined with a delay (Simmons et al., 2010). These results suggest a specific role for OFC in establishing context-specific parameters for value computation and for vlPFC in shifting or inhibiting behavior tendencies across domains when integrating multiple sources of information. Likewise, in a study of marmosets, Rygula et al. (2010) reported deficits in stimulus reversal learning following medial OFC lesions. Deficits were also observed following vlPFC lesions but only when new stimuli were used in the association, while reversals using existing associations were unaffected. This pattern is consistent with a role for OFC in value expectation and for vlPFC in new rule acquisition because of new stimulus sets. Similar regional PFC dissociations have also been reported in lesion studies examining human social behavior. Humans with lesions encompassing vlPFC report having fewer stereotypes about gender than do people with OFC lesions suggesting deficits in the application of socially constructed rules following vlPFC lesions (Gozzi et al., 2009).
Many neuroimaging studies have found that both OFC and vlPFC are readily engaged by social stimuli, particularly those that are emotionally salient (Guyer et al., 2008a, Kalin et al., 2007, Kim et al., 2007, Monk et al., 2008, Nitschke et al., 2004, O’Doherty et al., 2003b, Rolls et al., 2008, Winston et al., 2007) and a number of studies have examined the functional and structural development of these regions (Crone et al., 2006, Geier and Luna, 2009, Gogtay et al., 2004, Huttenlocher and Dabholkar, 1997, Steinberg et al., 2009). In the next sections we discuss VPFC functions applicable in generating social flexibility and development of these associations.
4. Functional overview of the VPFC
Within the VPFC, OFC lesions have long been known to produce perseverative patterns of behavior (e.g., repeatedly performing the same act without reinforcement) (Dias et al., 1996), which have led some to suggest that this region facilitates response reversal and response flexibility (Rolls et al., 2008). Although similar processes have been reported for sub-cortical structures such as the amygdala and striatum, recent conceptualizations have argued that the OFC specifically plays an important role in the more sophisticated process of computing an expected value of a stimulus, a process called valuation (Murray and Wise, 2010, Schoenbaum et al., 2009). While the OFC is not the only region in the brain where expected valuation is generated, the OFC is thought to be particularly important when this value computation must also integrate complex sequences, response sets, or temporal dynamics (Murray and Wise, 2010, Schoenbaum et al., 2009). In line with this proposition, Rangel et al. (2008) put forth a compelling framework for different valuation systems within Pavlovian, instrumental, or goal-directed learning contexts (Rangel et al., 2008). All three of these contexts likely play an important role in social interaction, and the OFC likely plays a key role in computing that emotional valuation under all three circumstances. Thus, the OFC may interact with regions such as the amygdala or striatum, where more elemental stimulus–response contingencies are formed, and where behavior patterns are executed, in order to guide behavior appropriately with respect to value expectations within a complex environment.
The more elemental stimulus-value associations generated in subcortical structures are thought to be integrated and updated with other information within the OFC. Thus, while a number of studies have shown that the OFC responds to emotional stimuli of both positive and negative valence (Elliott et al., 2000, Murray et al., 2007, O’Doherty et al., 2003a, Winston et al., 2007), recent studies have shown clearly that one of the central functions of the OFC is valuation or assigning a subjective value to a stimulus based on the integration of a number of different factors (Murray and Wise, 2010, Padoa-Schioppa and Assad, 2006, Schoenbaum et al., 2009). Schoenbaum et al. (2009) have further suggested that the expected value computation in the OFC may serve as a teaching signal through back projections onto areas such as striatum and amygdala, and possibly also regions relevant for social perception such as the superior temporal sulcus or fusiform gyrus to enable updating the salience of entire networks based upon expected stimulus value (Schoenbaum et al., 2009). The contribution of the OFC to valuation comes from correlation approaches such as neuroimaging and single unit recordings (Kringelbach and Rolls, 2004, O’Doherty et al., 2007). However, the association between OFC and valuation also derives from a number of lesion studies in rodents, nonhuman primates, and humans (Bechara et al., 1994, Rudebeck and Murray, 2008, Simmons et al., 2010), which indicate that the medial region of the OFC is not only involved, but necessary for valuation to occur in some circumstances. Nevertheless, the rodent work must be treated somewhat cautiously in its application to primates, as there is considerable controversy over the degree of homology between rodent and primate OFC.
Whereas the OFC interacts with cortical and limbic structures to generate and update the value of a stimulus, the vlPFC appears to be more involved in flexible behavioral engagement. The vlPFC is thought to be a region where highly processed visual information from the ventral visual stream integrates with motivational significance from the midline OFC to generate an action pattern which is then sent to premotor and motor cortex (Sakagami and Pan, 2007). Thus, while a key function of the OFC is integration and expectation, a key function of the vlPFC is integration and output. Motor output can include behaviors from whole body approach or avoidance to directing attention toward or away from a particular stimulus (Lee and Siegle, 2009, Leitman et al., 2010, Okamoto et al., 2009) to inhibiting or overriding behavioral tendencies toward stimuli, particularly when those tendencies have an emotional characteristic (Dillon and Pizzagalli, 2007, Garavan et al., 2006).
The vlPFC is also thought to play an important role in acquiring and retrieving rules for behavioral responses under different contingencies (Bunge, 2004, Donohue et al., 2008). This rule learning function may be dependent on interconnections with dorsal regions of prefrontal cortex in conjunction with value computations of OFC (Sakagami and Pan, 2007). Integrating behavioral output under different contingencies may also occur in vlPFC because integrative processing occurs across a longer delay period than many other regions (Sakagami and Pan, 2007, Simmons et al., 2010). Such a prolonged period for the maturation of integration of different contingencies may be necessary for complex learning functions like set switching or contextual modulation. As with the valuation literature, our understanding of the functional attributes of vlPFC in humans has largely been informed by neuroimaging and other correlational approaches. However, several recent lesion studies in nonhuman primates and humans have begun to dissociate the functions of ventrolateral and ventromedial cortex along similar lines (Gozzi et al., 2009, Rygula et al., 2010, Simmons et al., 2010).
5. How development progresses in VPFC
5.1. Synaptogenesis and pruning
Developmental neuroanatomical studies have shown that the maturation of the PFC overall is a protracted process, not reaching full maturity until the third decade of life (Gogtay et al., 2004, O’Donnell et al., 2005). Detailed morphometric studies have shown that synaptogenesis in the PFC continues postnatally at least until toddlerhood in humans (Huttenlocher and Dabholkar, 1997). This period of new synapse formation is protracted in the PFC relative to other brain regions. The initial process of synapse generation is then followed by a prolonged period of synapse elimination, which continues beyond puberty and into late adolescence (Huttenlocher and Dabholkar, 1997). The process of synapse elimination has been called pruning and is thought to represent a process of developmental learning whereby neuronal connections that are utilized are strengthened and those that are not needed are eliminated (Casey et al., 2000).
In the cortex, like synaptogenesis, the pruning process is completed first in primary sensory regions and last in frontal and parietal regions (Huttenlocher and Dabholkar, 1997). While relatively few studies have used human tissue to examine synapse formation across development, the results of many magnetic resonance imaging (MRI) studies performed in recent years reach the same general conclusion that there is an increase in gray matter through early to middle childhood followed by a decrease through late adolescence or early adulthood, and in areas such as the PFC, this maturation process is not complete until middle to late adolescence (Bramen et al., 2010, Casey et al., 2000, Giedd and Rapoport, 2010, Raznahan et al., 2010, Toga et al., 2006).
5.2. Myelination
A second key neurodevelopmental process involves myelination. Myelination is the process by which myelin, the fatty insulation around axons, is added to neurons. Myelination speeds communication between cells and, like synaptogenesis, does not reach maturity until late adolescence or early adulthood (Giedd and Rapoport, 2010, Toga et al., 2006). In contrast to the inverted U-shape of synapse creation and elimination across development, myelination is generally thought to be a progressive and linear process, and not subject to an initial overproduction and pruning-like process. Also, in contrast to synaptogenesis, myelination does not have the same degree of regional variation in timing that synaptogenesis does (Giedd and Rapoport, 2010), although there is some evidence that frontal regions reach mature levels of myelination slightly later than other regions (Giedd and Rapoport, 2010, Huttenlocher and Dabholkar, 1997, Toga et al., 2006). The myelination process may be particularly relevant for functions of the PFC because of the importance of network formation and the integration of multiple sources of information (Stevens et al., 2009). Together, these studies suggest that OFC and vlPFC undergo marked changes at a structural level from early postnatal life through late adolescence and possibly even into early adulthood.
5.3. Gender differences in development
Recent studies have found important sex differences in the maturation of the OFC and vlPFC that are directly related to puberty. These studies indicate that the gonadal surge at puberty may represent an important developmental period for the VPFC more generally (Bramen et al., 2010, Raznahan et al., 2010). Not only do the puberty findings underscore the prolonged developmental nature of the VPFC, but they also suggest that rapid punctual changes may also occur as a function of the pubertal surge of gonadal steroids and may relate to gender differences in behavior (Raznahan et al., 2010). Because puberty occurs much earlier in girls than boys, this may lead to gender differences in VPFC maturation as it relates to social flexibility. Indeed Raznahan et al. (2010) found that cortical thinning occurred in the OFC and vlPFC in both genders, but this reached maturity much sooner in girls than boys. The authors suggest that this may be one reason for the adolescent male predominance of psychopathologies that indicate impaired inhibitory control (i.e., aggression and risk-taking). In the present context, this may also indicate that young adolescent females are more likely to be facile at learning new social rules (i.e., flirting and peer alliance formation); value updating (i.e., generating on-line or context specific social expectations); and other social inhibitory controls (i.e., inhibiting sexual engagement) than age-matched adolescent males.
5.4. Asynchronous regional maturation?
Most of the studies of structural development to date have reported on a relatively gross scale comparing cortical to sub-cortical structures or frontal to parietal lobes, for example (Giedd and Rapoport, 2010, Toga et al., 2006). These results temper the ability to comment on patterns of structural and functional development within more localized sub-regions, such as the OFC and vlPFC. However, several developmental trends have been consistently observed in cerebral development that may indicate different rates of development within these two regions. In general, development progresses outwardly from midline to lateral extents as well as from caudal to rostral (Stiles, 2008). Development also progresses along a phylogenetic course with evolutionarily older structures emerging before newer ones (Konner, 2010, Gogtay et al., 2004). These patterns suggest that midline OFC regions may reach maturity earlier than the more lateral vlPFC regions and, in particular, the phylogenetically older agranular and dysgranular regions of caudal OFC may reach maturity earlier than the more rostral and fully granular regions of OFC (Price, 2007). Verification of this proposition awaits more detailed study, however.
6. Development of valuation
While a number of studies have found that the OFC responds to salient social stimuli of both positive and negative valence (Kalin et al., 2007, Nitschke et al., 2004, O’Doherty et al., 2003b, Winston et al., 2007), further evidence of OFC involvement in valuation comes from neuroimaging studies that report this region to be particularly responsive in situations involving altruistic decisions or charitable giving (Moll et al., 2006), competitive or co-operative social decisions (Hampton et al., 2008, McCabe et al., 2001, Phan et al., 2010, Sanfey et al., 2003), and inequality aversion (Tricomi et al., 2010). Other work focused on primate lesions has also indicated that OFC lesions lead to impairments in social and emotional behavior, including OFC damage that occurs early in life (Eslinger et al., 1997). For example, a detailed analysis of the social behavior of OFC-lesioned monkeys found that, following the lesion, monkeys did not alter their social behavior in a context or social partner specific manner. These results suggest that a compromised ability to adjust the value of a social stimulus can produce a clear functional impairment in social flexibility. Interestingly, these monkeys also displayed more aggression overall in addition to their general inflexible pattern of behavior (Machado and Bachevalier, 2006). A similar increase in aggressive behavior has also been observed in rats following OFC lesions (Rudebeck and Murray, 2008). It is possible that the increased aggression may be secondary to an inability to flexibly modulate behavior due to difficulty in generating an online update of social value using cues such as facial expression or vocal feedback (Machado and Bachevalier, 2006). This would result in animals finding themselves in unpredictable social situations where aggression may be the only option in their behavioral repertoire, if the real-time social information updating process is impaired.
Relatively little is known about the neural development of social valuation, particularly in humans. The process of assigning value to a social stimulus is likely present from a very early age, and indeed maternal stimuli have been found to induce OFC activity in infants (Minagawa-Kawai et al., 2009). In a study measuring frontal electroencephalogram activity, preschool-aged children with relatively greater right frontal activity at rest showed more withdrawal behaviors after receiving an unappealing toy (Forbes et al., 2006). Although this study did not directly assay OFC functional activity in this context, it does provide empirical support of the VPFC's role more broadly defined in the relationship between assigning value to stimuli and social responding early in life and in relation to individual neural differences. These social valuation processes are likely to become increasingly complex with age as more and more social and contextual information is integrated into the valuation computation. Thus, as the OFC becomes increasingly interconnected with cortical, thalamic, limbic, and striatal structures across development, the amount and type of information that informs the calculation of stimulus value also increases dramatically and valuation becomes more intricate and nuanced in how it influences social behavior as a result.
One example of stimulus valuation can be found in both behavioral and neuroimaging studies that have used the temporal discounting task. Temporal discounting refers to the difference in the value assigned to a stimulus that is immediately available versus one that is available at some future point. An immediately available reward is always more valuable than one that is delayed, and the degree of that difference is measured in temporal discounting tasks. Neuroimaging work has shown that the OFC and striatum interact with frontal dorsolateral and parietal circuits, which are neural circuits involved in temporal assessment while performing this task (McClure et al., 2007, McClure et al., 2004). In a large behavioral study that examined the age-related changes in temporal discounting processing, Steinberg and colleagues found that mature behavioral response patterns (e.g., stronger orientation toward future versus immediate outcomes) did not emerge on this task until middle to late adolescence (Steinberg et al., 2009). Findings from this study suggested that, although children are able to understand and perform the temporal discounting task, they do not integrate all of the variables involved at a mature level until much later in development. Similar maturational differences are likely to occur with valuation performance in other contexts, such as assigning value to a social stimulus.
In terms of social maturation, the valuation process is also likely to become increasingly complex as development progresses. In a recent study, activity was found to co-occur in ventral striatum and OFC when positive social feedback was obtained from an individual that participants were expecting to like them (Gunther Moor et al., 2010). This suggests that positive social feedback may have been integrated into the valuation process via the OFC. The result of this integration appears to have been an overall increase in value attributed to expected positive feedback via the social neural circuitry drawing from both cortical and subcortical networks. This pattern displayed a linear increase from middle childhood through adolescence suggesting a linear pattern of the development of social valuation. Interestingly, social rejection feedback has not been associated with significant neural response within this circuit, a pattern that we have also documented in a similar task (Guyer et al., unpublished observations). This may indicate that, in the context of computerized social evaluation tasks, positive social feedback may be more relevant than negative social feedback for informing stimulus valuation, particularly with increased age and maturation.
In early life, functional input of the relatively immature OFC may be reflected in a less discriminate social response pattern. However, as development progresses, the value of different individuals becomes more varied and even context or experience dependent (i.e., some individuals are highly valued in some contexts, whereas other individuals are highly valued in other contexts) (Barbu, 2003, Furman and Buhrmester, 1992, Howes, 1990, Steinberg, 2005). Later in development, the functional input of the OFC likely supports the integration of more abstract information, such as goals and values in social contexts, into the assessment of social stimulus value. Another developmentally sensitive aspect of social value assessment is that the innate value of different classes of social stimuli likely changes across development. In other words, the inherent value of a mother, a peer, or a member of the opposite sex is different at different points in development, probably at least in part because of programmed changes within the limbic system that are additionally guided by pubertal changes signaling the onset of sexual maturity (Nelson et al., 2005). In Table 1, we provide some illustrative examples of what valuation may look like at different developmental periods. Because the valuation process within the OFC integrates this “bottom-up” limbic information with cortical and other “top-down” information, these innately fluctuating limbic responses are likely to have a large impact on the OFC-driven valuation as well.
Table 1.
Cognitive and behavioral examples of valuation, inhibitory control, and rule use in different social contexts across development.
| Early childhood | Middle childhood | Adolescence | |
|---|---|---|---|
| Valuation | |||
| Family | Mom is mad because she yelled; dad is happy because he smiled | Dad will be more likely to help me after dinner than before | She is in a good mood. Now I will ask her for the car |
| Peer (same-sex) | He is nice because he laughs | I want to play with her because she has an awesome toy | He will take me to lots of parties because he is on the football team |
| Peer (opposite-sex) | She is mean because she's bossy | He will be fun if other girls are around | He may not be handsome, but he is funny |
| Inhibitory control | |||
| Family | Don't hurt your sister | Play quietly | Don't scream at your parents |
| Peer (same-sex) | Don't play with other kids’ toys | Don't be chicken | Don't let them know you like stuffed animals |
| Peer (opposite-sex) | Don't push | Don't cry in front of them | Don't kiss her |
| Rule use | |||
| Family | We can't play in here | Dad will buy me ice cream, but mom won't | Tease your brother and his friends, but not when mom is around |
| Peer (same-sex) | Share your toys | Talk about sports; play word games | The clothes you wear define your group |
| Peer (opposite-sex) | In our game, I’ll be the girls, you be the boys | Boys don't do girl things, and girls don't do boy things | To flirt sit close, make eye contact, and laugh |
7. Development of response inhibition
The vlPFC clearly plays an important role in inhibiting prepotent cognitive, motor, and emotional responses (Dillon and Pizzagalli, 2007, Garavan et al., 2006). From a behavioral standpoint, this may be one of the earliest attributes of the ventral prefrontal cortex to emerge as the ability to inhibit a prepotent or motivated response tendency has been demonstrated in children as young as four years of age (Davidson et al., 2006, Luna et al., 2001, Mischel et al., 1989, Mischel et al., 2010). However, behavioral similarity does not necessarily indicate functional isomorphism (Schlaggar et al., 2002), and inhibition may be associated with a different neuronal signature in young children than in adults. Furthermore, a number of studies have shown that the ability to inhibit responses, as well as vlPFC activity elicited while individuals are engaged in an inhibitory task, continues to mature from late childhood through late adolescence even when task difficulty is controlled (Geier and Luna, 2009, Luna et al., 2001, Rubia et al., 2006, Rubia et al., 2007). In addition, the ability to inhibit has been shown to become increasingly willful and resistant to distraction through at least middle adolescence (Davidson et al., 2006).
In the context of social flexibility, children must engage this function of vlPFC to inhibit tendencies in a number of social situations. For example, many children experience some degree of fear when encountering new individuals, and children with a temperament of extreme behavioral inhibition demonstrate this reaction to an even greater extent (Fox et al., 2005, Kagan et al., 1984), a temperament which often evolves into clinical levels of social anxiety. Therefore, in order to engage in social interaction with new people, these behavioral inhibitory tendencies must be overcome, presumably at least in part through inhibitory control exerted by the vlPFC. Likewise, children who have aggressive tendencies when conflict emerges in social interactions must learn to inhibit these behavioral response tendencies in order for social interaction to progress adaptively. Another important aspect of response inhibition is the willful ability to direct attention toward or away from a salient aspect of the environment. In previous studies of this phenomenon, we and others have found that adults are better able to engage the vlPFC in tasks that required attention switching while viewing emotionally provocative faces. Under these task demands, adolescents had greater activity in emotional circuits, particularly while viewing these stimuli without any explicit attentional direction (Guyer et al., 2008b, Monk et al., 2003, Passarotti et al., 2009). We believe that these findings suggest an increasing ability to engage the vlPFC to override basic emotional responding to salient social stimuli. The ability to willfully control attention is likely to play a key role in social situations where an ultimate goal conflicts with an otherwise overriding tendency. For example, if an individual has to interact with someone higher on the dominance hierarchy or with someone with whom past interactions might bias behavior or attention away from the present interaction (e.g., in order to obtain access to a desired party), the individual must engage the vlPFC to inhibit the tendency to flee or to focus on the last fight. Table 1 provides examples of response inhibition changes across ages within different social contexts.
Eisenberg et al. (2009) have argued for a distinction between effortful inhibition of orienting and engagement, and a more reflexive inhibition of hyper-responsive motoric and affective responding (e.g., driven by regions such as the amygdala). While these two behavioral control systems likely overlap greatly both across individuals and in the neuronal patterns engaged, the effortful subcomponent may involve more lateral aspects and the reflexive more medial aspects of VPFC. Eisenberg has argued that this distinction may help distinguish between temperamental predisposition toward inhibition (e.g., high in effortful control) and active internalizing disorders such as anxiety (e.g., low in reflexive control), although both conditions are likely to be associated with hyperactivity in sub-cortical structures (e.g., the amygdala). Effortful control is thought to be highly heritable and to emerge early in development, although it may continue to mature until much later in life (Eisenberg et al., 2009).
8. Development of flexible rule use
Bunge and colleagues have shown that in complex cognitive tasks, the vlPFC is engaged when individuals acquire new rules for how behavior is governed and also when the rules are held in working memory during task execution. The vlPFC is not involved in the long term storage of these rules, however (Souza et al., 2009). The capacity of vlPFC to perform this function increases from middle childhood through early adulthood (Crone et al., 2006). We have found a similar pattern in a risk-taking task in which stronger activation in vlPFC was observed in adults than in children or adolescents when making risky decisions (Eshel et al., 2007). Given the purported role of vlPFC in rule use, we interpret this finding to indicate that decision-making in adults may involve greater computation of available options than it does in children. Rubia et al., 2006, Rubia et al., 2007 have also shown a gradual linear increase across both childhood and adolescence in set switching tasks that likely involves the temporary storage of different rules in vlPFC. Similar results have been obtained in behavioral tasks where flexible shifting is among the latest executive functions to mature (Davidson et al., 2006).
The development of social rule use is also likely to develop slowly and reach maturity in adolescence or early adulthood. While the protracted development of this function is in part the result of physical maturation of the vlPFC and its interconnections, it may also be protracted because the rules needed for social interaction differ across periods of development, must be acquired with experience, and vary by social context. For example, in early life, different rules may be needed for interacting with a sibling than with a parent. Further, within a small social group such as one's family, the number of rule sets demanded is likely minimal and become more routine as children age, thus, requiring less engagement of vlPFC (Bunge, 2004). See Table 1 for examples of social applications of rule use across development.
As children become more independent and begin interacting more with peers in contexts away from the established family system, social rule use becomes a more variable and novel process. For example, there are explicit rules for playing different games, as well as many socially imposed implicit rules regarding interactions with members of same versus opposite sex. This appears particularly evident in later childhood and early adolescence when there are further rules for how to interact with members in different levels of the dominance hierarchy (Bell et al., 2009, Cikara et al., 2010, Marsh et al., 2009). The rules change once again when children reach sexual maturity and must learn how to interact with same- and opposite-sex peers in the context of dating and mating. In adolescence, there are often many different cliques and peer subgroups to which one belongs. Each peer group or clique likely has their own explicit or implicit applicable rules of socialization that must be learned and brought into working memory. Indeed, a recent study found vlPFC activation when adults thought about in-group versus out-group members (Cikara et al., 2010). Therefore, the developmental process of vlPFC may depend not only on physical maturation, but also on socio-cultural experience. It may be that these different socio-cultural experiences emerge at different points in development precisely when they do because the refinement of the vlPFC's role reaches a maturational peak within a similar chronological age range in most individuals. In other words, brain development may be driving the expression of socio-cultural development to some extent.
9. Sensitive periods for the development of social flexibility
One of the consequences of prolonged brain and social development is that it provides substantial opportunity for the environment to fundamentally alter the structure of the nervous system as it is being built. In many ways, development can be thought of as learning in slow motion (Fox et al., 2010, Knudsen, 2004). Furthermore, in the development of many neural systems, there appear to be sensitive periods, where circuit formation is particularly affected by experience or environmental input (Forbes and Dahl, 2010, Fox et al., 2010, Knudsen, 2004, Steinberg, 2005, Tottenham and Sheridan, 2009). There are several studies that suggest sensitive periods in VPFC organization of social behavior may occur as well. Anderson and colleagues (Anderson et al., 1999) reported a finding from two individuals who received OFC lesions prior to 15 months of age. These two individuals displayed patterns of aggressive and uncooperative social behavior and marked difficulties in moral reasoning that had not been observed in individuals who received OFC lesions later in life. These findings suggest that OFC activity early in life may play an important role in learning and internalizing rules of social behavior. In the present conceptualization, to the extent that lesions are confined to the medial and orbital regions in these studies, the primary deficit in OFC is likely in the ability to generate accurate expectations of value from social stimuli (or consequences of social transgressions), which then leads to difficulties in generating meaningful rules in vlPFC because of degraded stimulus expectation information.
Emerging neuroimaging-based evidence indicates that adolescence may also be a period in which neurodevelopment is sensitive to social experiences, or at least a period during which previously acquired organization may be expressed (Brody et al., 2009, Guyer et al., 2009, Maheu et al., 2010, Whittle et al., 2009, Yap et al., 2008). Andersen et al. reported volumetric size reductions in specific brain regions are linked to vulnerability to psychosocial adversity within specific periods across childhood and adolescence. While limbic regions are most affected by adversity early in life, the PFC in general appears to be particularly sensitive to adverse experiences that occur during adolescence (Andersen et al., 2008). Adverse events may also have a cumulative effect specifically on VPFC development. A recent study of neurobiological development in children who were reared in an abusive environment reported volume reductions in VPFC that also related to decrements in social functioning (Hanson et al., 2010). This suggests that an adverse rearing environment may not foster optimal VPFC maturation and one consequence of this may be a decrement in social flexibility and social functioning. Interestingly, in another recent study in which behavioral interaction between mothers and sons was observed and related to structural MRI analysis, the OFC in particular was found to be larger in adolescents whose mothers displayed an emotionally negative interactive style (Whittle et al., 2009), and greater VPFC volume has been found among adolescents who, at age five, demonstrated extreme social inhibition during play tasks with novel peers, whereby they spent more time next to their mother and more time staring at the peer (Hill et al., 2010). While it is not clear why the directions of these effects differ in these studies, all four indicate that VPFC regions (vlPFC and OFC) differ in adolescents who had either adverse social environments or expressed extreme social behavior early in life and suggest that VPFC development may be particularly sensitive to social experience during development due to its role in internalizing flexible control of social behavior.
Observations of changes in rodent social behavior are also consistent with a developmentally sensitive role for VPFC in social behavior. Interestingly, these behavioral deficits do not appear at the time of lesion, but rather emerge much later in development. For example, neonatal lesions of OFC in rats result in an aberrant pattern of social behavior in rats when they reach puberty. Normal male adult rats express different patterns of social behavior toward other male rats depending on their position in the dominance hierarchy. However, rats who received OFC lesions early in life fail to show this moderation in their social behavior after puberty and instead behave similarly to all other males, regardless of their dominance position (Kolb et al., 2004). In addition, a recent study found that early life social experience can directly affect the complexity of neurons within the OFC. For the rat, the typical juvenile period is characterized by a great deal of rough and tumble play. Rats reared in a variety of social play experiences as juveniles had OFC neurons with longer dendrites and more arbors than did rats reared in conditions lacking in social variety (Bell et al., 2010). These findings suggest that the OFC plays an important role in discriminating among different individuals and adjusting behavior accordingly. Importantly, these data suggest that early life social experiences impact social behavior in mature animals. Among humans, a recent study found that style of rough and tumble play with fathers moderated aggressive behavior in young aggressive children. Play styles in which fathers exerted their dominance status resulted in significant overall reductions in aggressive behavior in children relative to play styles in which the father did not display dominance (Flanders et al., 2009). This suggests that, in humans as in rats, play in childhood may be an important means of acquiring inhibitory skills and learning different patterns of behavioral engagement via VPFC mediated processes. It important again, however, to highlight that there is considerable controversy surrounding the degree of homology between rodent and primate OFC regions.
The findings described above regarding the influence of social experience on neural complexity in rats suggest that juvenile play may be an important means of OFC organization during development, and that experiences occurring during this juvenile period may not be expressed behaviorally until much later (Bell et al., 2010). Whether or not this is truly a sensitive period awaits further experimentation, with specific focus on evaluating the effects of play at different periods in development. In general, while sensitive periods of organization may be a fundamental property of neural development (Fox et al., 2010, Knudsen, 2004), this may be particularly difficult to detect for VPFC-related processes for two reasons. First, the prolonged maturation period of the PFC (20+ years in humans) makes it difficult to detect sensitive periods of organization; in the PFC, this may be more like a sensitive epoch than a sensitive period. Secondly, the very nature of the PFC is to integrate with many other regions that are themselves undergoing prolonged maturation. Accordingly, there may be many sub-components of sensitivity. For example, organizing dominance-related behavioral patterns may be sensitive during the juvenile rough and play period, but principles of courting-related behavioral patterns may not be sensitive until after puberty. If there are sensitive periods for the development of social behaviors, then it may mean that rules of engagement, inhibitory tendencies, or methods of valuation calculations that are acquired at different points in development are not absolute. Further, rules acquired during earlier periods of social development may be more easily learned and more resistant to extinction than rules learned later, just as language is more easily acquired early in life than later in life.
10. VPFC in the genesis of mood and anxiety disorders in adolescence
While many of the basic roles of the VPFC in social flexibility have been established by the time of adolescence, they continue to be refined and altered by both changes in cortical organization and changes in subcortical (e.g., bottom up) processes related to puberty (Forbes and Dahl, 2010, Gardner and Steinberg, 2005, Nelson et al., 2005). The literature reviewed above informs our understanding of how adolescents learn to competently and flexibly navigate social interactions with peers. However, perturbations in the process of developing social flexibility may relate to the emergence of some forms of psychopathology in adolescence, such as social anxiety (La Greca and Harrison, 2005, Nelson et al., 2005, Pine et al., 2009).
Social anxiety is characterized by an impairing fear of negative evaluation from others, including situations that appear socially positive or at least neutral to individuals without social anxiety (Guyer et al., 2008a, La Greca and Stone, 1993, Rapee and Heimberg, 1997). Socially anxious individuals show difficulty in assigning appropriate valuations to social stimuli (e.g., fears of negative evaluation from friends), may have difficulty inhibiting emotional tendencies, and may not competently apply appropriate social rules to changing environmental demands or contingencies.
Findings from work on the neural underpinnings of cognitive flexibility and regulation suggest that the VPFC is engaged during events of potential social evaluation. In these situations, the VPFC may facilitate reframing of the situation and adjusting affect and goal-related behavior accordingly through a combined process of inhibiting tendencies and selecting appropriate responses (Badre and Wagner, 2006, Ochsner et al., 2004, Phan et al., 2005, Souza et al., 2009). For example, in a study of anticipated social evaluation, adolescents with clinical social anxiety showed positive co-activation of the amygdala and the left vlPFC while anticipating evaluation specifically from low- versus high-desirability peers (Guyer et al., 2008a). This suggests that, among socially anxious adolescents, vlPFC activity coupled with amygdala activity may reflect attentional processes directed to the need for inhibiting or regulating one's response to another individual who poses a subjectively experienced potential threat.
Of further relevance is research from two separate studies of adolescents with generalized anxiety disorder that found increased right vlPFC activation in anxious relative to control subjects when viewing angry faces (Maslowsky et al., 2010, Monk et al., 2006). In an initial study, the severity of anxiety among patients was inversely related to vlPFC activation and positively associated with amygdala activity (Monk et al., 2006). This pattern of results suggests that the detection of a threat engages both fear-related circuitry instantiated in the amygdala and inhibitory or other regulatory circuitry within vlPFC. One possibility is that the VLPC moderates amygdala response either directly by inhibiting (possibly via OFC) or indirectly by engaging other response patterns that reduce fear-related activity in the amygdala. Furthermore, the degree to which vlPFC may subsequently modulate behavior appears to be reflected in the symptom profile. In another example, adolescent anxiety patients showed increased vlPFC activation to threatening faces after receiving either cognitive-behavioral or psychopharmacological treatment (Maslowsky et al., 2010). Again, vlPFC response in anxious adolescents may facilitate signaling to other brain regions that correlate with anxiety, such as the amygdala or OFC, during social threat processing in order to maintain representation of task-related goals in the presence of salient social-emotional feedback. These patterns suggest a compensatory mechanism through engagement of the vlPFC in adolescent anxiety. While these studies have found a consistent pattern of vlPFC activity in combination with amygdala hyperactivation in anxious adolescents, the exact role of the vlPFC is not clear, particularly in relation to the amygdala, and awaits further study.
11. Summary and conclusion
Our goal in this review was to address questions about the involvement of specific PFC regions in supporting flexible control of behavior within the VPFC. This is can be viewed as a general function with application to a number of different behavioral domains. However, our purpose here was to focus on social development and delineate how this pattern of flexibility via neural maturation might affect social behavior across development.
The findings reviewed above indicate that the VPFC plays an important role in flexible social behavior, and like other VPFC-related processes, undergoes a prolonged period of maturation. We believe that three specific aspects of social flexibility are governed by the VPFC – valuation, inhibition, and generation and usage of behavioral rules. At a functional level, all three of these processes have been shown to display developmental changes from early childhood through adolescence and even into early adulthood. We believe the present framework will help to clarify the functional capabilities of individuals’ social behavior as they mature. Furthermore, although sensitive periods of VPFC development have not been clearly demonstrated, several suggestive studies have been published. Future studies of the VPFC should pay particular attention to the specific processes that are being identified by individual studies and to the developmental phase in which individuals are in. Individual differences in the sensitivity of VPFC functions and developmental trajectories of VPFC-limbic interactions may relate to varied patterns of social behavior present across development.
We believe that it is important to focus on social behavior in relation to the VPFC in part because of the prolonged maturation period of this region. In particular, adolescence is a highly salient time frame in which the VPFC undergoes protracted maturation and functional refinement. Adolescence also involves a period of tremendous social change. It is possible that this extended period of VPFC development is designed as such in order to accommodate the nuanced and complex types of social exchanges that adolescents encounter as they become sexually mature and prepare for adult roles.
Much of what we have proposed here is based on inferential reasoning derived from combining studies of cognitive neuroscience, cognitive development, and social behavior. Critical next steps in this area will involve studies that test hypotheses proposed in this review for scientific verification and revisions to the ideas proposed here. The results of such studies will help to integrate mechanism into theories of social development and to guide and constrain future theories of the neurobiology of social development.
Acknowledgements
The authors report no financial relationships with commercial interests. This research was supported by the Intramural Research Program of the National Institute of Mental Health and NIH Career Development Grants to A.E.G. (K99 MH080076 and R00 MH080076).
References
- Amodio D.M., Frith C.D. Meeting of minds: the medial frontal cortex and social cognition. Nat. Rev. Neurosci. 2006;7(4):268–277. doi: 10.1038/nrn1884. [DOI] [PubMed] [Google Scholar]
- Andersen S.L., Tomada A., Vincow E.S., Valente E., Polcari A., Teicher M.H. Preliminary evidence for sensitive periods in the effect of childhood sexual abuse on regional brain development. J. Neuropsychiatry Clin. Neurosci. 2008;20(3):292–301. doi: 10.1176/appi.neuropsych.20.3.292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson S.W., Bechara A., Damasio H., Tranel D., Damasio A.R. Impairment of social and moral behavior related to early damage in human prefrontal cortex. Nat. Neurosci. 1999;2(11):1032–1037. doi: 10.1038/14833. [DOI] [PubMed] [Google Scholar]
- Badre D., Wagner A.D. Computational and neurobiological mechanisms underlying cognitive flexibility. Proc. Natl. Acad. Sci. U.S.A. 2006;103(18):7186–7191. doi: 10.1073/pnas.0509550103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbas H. Specialized elements of orbitofrontal cortex in primates. Ann. N. Y. Acad. Sci. 2007;1121:10–32. doi: 10.1196/annals.1401.015. [DOI] [PubMed] [Google Scholar]
- Barbu S. Stability and flexibility in preschoolers’ social networks: a dynamic analysis of socially directed behavior allocation. J. Comp. Psychol. 2003;117(4):429–439. doi: 10.1037/0735-7036.117.4.429. [DOI] [PubMed] [Google Scholar]
- Bechara A., Damasio A.R., Damasio H., Anderson S.W. Insensitivity to future consequences following damage to human prefrontal cortex. Cognition. 1994;50(1–3):7–15. doi: 10.1016/0010-0277(94)90018-3. [DOI] [PubMed] [Google Scholar]
- Bell H.C., McCaffrey D.R., Forgie M.L., Kolb B., Pellis S.M. The role of the medial prefrontal cortex in the play fighting of rats. Behav. Neurosci. 2009;123(6):1158–1168. doi: 10.1037/a0017617. [DOI] [PubMed] [Google Scholar]
- Bell H.C., Pellis S.M., Kolb B. Juvenile peer play experience and the development of the orbitofrontal and medial prefrontal cortices. Behav. Brain Res. 2010;207(1):7–13. doi: 10.1016/j.bbr.2009.09.029. [DOI] [PubMed] [Google Scholar]
- Blair R.J. Dysfunctions of medial and lateral orbitofrontal cortex in psychopathy. Ann. N. Y. Acad. Sci. 2007;1121:461–479. doi: 10.1196/annals.1401.017. [DOI] [PubMed] [Google Scholar]
- Blakemore S.J. The social brain in adolescence. Nat. Rev. Neurosci. 2008;9(4):267–277. doi: 10.1038/nrn2353. [DOI] [PubMed] [Google Scholar]
- Bramen J.E., Hranilovich J.A., Dahl R.E., Forbes E.E., Chen J., Toga A.W. Puberty influences medial temporal lobe and cortical gray matter maturation differently in boys than girls matched for sexual maturity. Cereb. Cortex. 2010 doi: 10.1093/cercor/bhq137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brody G.H., Beach S.R., Philibert R.A., Chen Y.F., Murry V.M. Prevention effects moderate the association of 5HTTLPR and youth risk behavior initiation: gene x environment hypotheses tested via a randomized prevention design. Child Dev. 2009;80(3):645–661. doi: 10.1111/j.1467-8624.2009.01288.x. [DOI] [PubMed] [Google Scholar]
- Brothers L., Ring B., Kling A. Response of neurons in the macaque amygdala to complex social stimuli. Behav. Brain Res. 1990;41(3):199–213. doi: 10.1016/0166-4328(90)90108-q. [DOI] [PubMed] [Google Scholar]
- Bunge S.A. How we use rules to select actions: a review of evidence from cognitive neuroscience. Cogn. Affect. Behav. Neurosci. 2004;4(4):564–579. doi: 10.3758/cabn.4.4.564. [DOI] [PubMed] [Google Scholar]
- Carmichael S.T., Price J.L. Architectonic subdivision of the orbital and medial prefrontal cortex in the macaque monkey. J. Comp. Neurol. 1994;346(3):366–402. doi: 10.1002/cne.903460305. [DOI] [PubMed] [Google Scholar]
- Casey B.J., Giedd J.N., Thomas K.M. Structural and functional brain development and its relation to cognitive development. Biol. Psychol. 2000;54(1–3):241–257. doi: 10.1016/s0301-0511(00)00058-2. [DOI] [PubMed] [Google Scholar]
- Casey B.J., Jones R.M., Hare T.A. The adolescent brain. Ann. N. Y. Acad. Sci. 2008;1124:111–126. doi: 10.1196/annals.1440.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cikara M., Farnsworth R.A., Harris L.T., Fiske S.T. On the wrong side of the trolley track: neural correlates of relative social valuation. Soc. Cogn. Affect. Neurosci. 2010 doi: 10.1093/scan/nsq011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crone E.A., Donohue S.E., Honomichl R., Wendelken C., Bunge S.A. Brain regions mediating flexible rule use during development. J. Neurosci. 2006;26(43):11239–11247. doi: 10.1523/JNEUROSCI.2165-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson R.J., Putnam K.M., Larson C.L. Dysfunction in the neural circuitry of emotion regulation: a possible prelude to violence. Science. 2000;289:591–594. doi: 10.1126/science.289.5479.591. [DOI] [PubMed] [Google Scholar]
- Davidson M.C., Amso D., Anderson L.C., Diamond A. Development of cognitive control and executive functions from 4 to 13 years: evidence from manipulations of memory, inhibition, and task switching. Neuropsychologia. 2006;44(11):2037–2078. doi: 10.1016/j.neuropsychologia.2006.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dias R., Robbins T.W., Roberts A.C. Dissociation in prefrontal cortex of affective and attentional shifts. Nature. 1996;380(6569):69–72. doi: 10.1038/380069a0. [DOI] [PubMed] [Google Scholar]
- Dillon D.G., Pizzagalli D.A. Inhibition of action, thought, and emotion: a selective neurobiological review. Appl. Prev. Psychol. 2007;12(3):99–114. doi: 10.1016/j.appsy.2007.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolan R.J. Keynote address: revaluing the orbital prefrontal cortex. Ann. N. Y. Acad. Sci. 2007;1121:1–9. doi: 10.1196/annals.1401.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donohue S.E., Wendelken C., Bunge S.A. Neural correlates of preparation for action selection as a function of specific task demands. J. Cogn. Neurosci. 2008;20(4):694–706. doi: 10.1162/jocn.2008.20042. [DOI] [PubMed] [Google Scholar]
- Dunbar R.I., Shultz S. Understanding primate brain evolution. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 2007;362(1480):649–658. doi: 10.1098/rstb.2006.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenberg N., Valiente C., Spinrad T.L., Liew J., Zhou Q., Losoya S.H. Longitudinal relations of children's effortful control, impulsivity, and negative emotionality to their externalizing, internalizing, and co-occurring behavior problems. Dev. Psychol. 2009;45(4):988–1008. doi: 10.1037/a0016213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott R., Dolan R.J., Frith C.D. Dissociable functions in the medial and lateral orbitofrontal cortex: evidence from human neuroimaging studies. Cereb. Cortex. 2000;10(3):308–317. doi: 10.1093/cercor/10.3.308. [DOI] [PubMed] [Google Scholar]
- Eshel N., Nelson E.E., Blair R.J., Pine D.S., Ernst M. Neural substrates of choice selection in adults and adolescents: development of the ventrolateral prefrontal and anterior cingulate cortices. Neuropsychologia. 2007;45(6):1270–1279. doi: 10.1016/j.neuropsychologia.2006.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eslinger P.J., Biddle K.R., Grattan L.M. Development of the Prefrontal Cortex: Evolution, Neurobiology, and Behavior. Paul H. Brookes Publishing; Baltimore, MD: 1997. Cognitive and social development in children with prefrontal cortex lesions. [Google Scholar]
- Flanders J.L., Leo V., Paquette D., Pihl R.O., Seguin J.R. Rough-and-tumble play and the regulation of aggression: an observational study of father-child play dyads. Aggressive Behav. 2009;35(4):285–295. doi: 10.1002/ab.20309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forbes E.E., Dahl R.E. Pubertal development and behavior: hormonal activation of social and motivational tendencies. Brain Cogn. 2010;72(1):66–72. doi: 10.1016/j.bandc.2009.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forbes E.E., Fox N.A., Cohn J.F., Galles S.F., Kovacs M. Children's affect regulation during a disappointment: psychophysiological responses and relation to parent history of depression. Biol. Psychol. 2006;71(3):264–277. doi: 10.1016/j.biopsycho.2005.05.004. [DOI] [PubMed] [Google Scholar]
- Fox N.A., Henderson H.A., Marshall P.J., Nichols K.E., Ghera M.M. Behavioral inhibition: linking biology and behavior within a developmental framework. Annu. Rev. Psychol. 2005;56:235–262. doi: 10.1146/annurev.psych.55.090902.141532. [DOI] [PubMed] [Google Scholar]
- Fox S.E., Levitt P., Nelson C.A., 3rd. How the timing and quality of early experiences influence the development of brain architecture. Child Dev. 2010;81(1):28–40. doi: 10.1111/j.1467-8624.2009.01380.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furman W., Buhrmester D. Age and sex differences in perceptions of networks of personal relationships. Child Dev. 1992;63(1):103–115. doi: 10.1111/j.1467-8624.1992.tb03599.x. [DOI] [PubMed] [Google Scholar]
- Gallagher H.L., Frith C.D. Functional imaging of ‘theory of mind’. Trends Cogn. Sci. 2003;7(2):77–83. doi: 10.1016/s1364-6613(02)00025-6. [DOI] [PubMed] [Google Scholar]
- Garavan H., Hester R., Murphy K., Fassbender C., Kelly C. Individual differences in the functional neuroanatomy of inhibitory control. Brain Res. 2006;1105(1):130–142. doi: 10.1016/j.brainres.2006.03.029. [DOI] [PubMed] [Google Scholar]
- Gardner M., Steinberg L. Peer influence on risk taking, risk preference, and risky decision making in adolescence and adulthood: an experimental study. Dev. Psychol. 2005;41(4):625–635. doi: 10.1037/0012-1649.41.4.625. [DOI] [PubMed] [Google Scholar]
- Geier C., Luna B. The maturation of incentive processing and cognitive control. Pharmacol. Biochem. Behav. 2009;93(3):212–221. doi: 10.1016/j.pbb.2009.01.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giedd J.N., Rapoport J.L. Structural MRI of pediatric brain development: what have we learned and where are we going? Neuron. 2010;67(5):728–734. doi: 10.1016/j.neuron.2010.08.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gogtay N., Giedd J.N., Lusk L., Hayashi K.M., Greenstein D., Vaituzis A.C. Dynamic mapping of human cortical development during childhood through early adulthood. Proc. Natl. Acad. Sci. U.S.A. 2004;101(21):8174–8179. doi: 10.1073/pnas.0402680101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gozzi M., Raymont V., Solomon J., Koenigs M., Grafman J. Dissociable effects of prefrontal and anterior temporal cortical lesions on stereotypical gender attitudes. Neuropsychologia. 2009;47(10):2125–2132. doi: 10.1016/j.neuropsychologia.2009.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gunther Moor B., van Leijenhorst L., Rombouts S.A., Crone E.A., Van der Molen M.W. Do you like me? Neural correlates of social evaluation and developmental trajectories. Soc. Neurosci. 2010:1–22. doi: 10.1080/17470910903526155. [DOI] [PubMed] [Google Scholar]
- Guyer A.E., Lau J.Y., McClure-Tone E.B., Parrish J., Shiffrin N.D., Reynolds R.C. Amygdala and ventrolateral prefrontal cortex function during anticipated peer evaluation in pediatric social anxiety. Arch. Gen. Psychiatry. 2008;65(11):1303–1312. doi: 10.1001/archpsyc.65.11.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer A.E., McClure-Tone E.B., Shiffrin N.D., Pine D.S., Nelson E.E. Probing the neural correlates of anticipated peer evaluation in adolescence. Child Dev. 2009;80(4):1000–1015. doi: 10.1111/j.1467-8624.2009.01313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer A.E., Monk C.S., McClure-Tone E.B., Nelson E.E., Roberson-Nay R., Adler A.D. A developmental examination of amygdala response to facial expressions. J. Cogn. Neurosci. 2008;20(9):1565–1582. doi: 10.1162/jocn.2008.20114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer, A.E., Choate, V.R., Pine, D.S., Nelson, E.E., Neural circuitry underlying affective response to peer feedback in adolescence, unpublished observations. [DOI] [PMC free article] [PubMed]
- Hampton A.N., Bossaerts P., O’Doherty J.P. Neural correlates of mentalizing-related computations during strategic interactions in humans. Proc. Natl. Acad. Sci. U.S.A. 2008;105(18):6741–6746. doi: 10.1073/pnas.0711099105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson J.L., Chung M.K., Avants B.B., Shirtcliff E.A., Gee J.C., Davidson R.J. Early stress is associated with alterations in the orbitofrontal cortex: a tensor-based morphometry investigation of brain structure and behavioral risk. J. Neurosci. 2010;30(22):7466–7472. doi: 10.1523/JNEUROSCI.0859-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill S.Y., Tessner K., Wang S., Carter H., McDermott M. Temperament at 5 years of age predicts amygdala and orbitofrontal volume in the right hemisphere in adolescence. Psychiatry Res. 2010;182(1):14–21. doi: 10.1016/j.pscychresns.2009.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howes C. Social status and friendship from kindergarten to third grade. J. Appl. Dev. Psychol. 1990;11:321–330. [Google Scholar]
- Huttenlocher P.R., Dabholkar A.S. Regional differences in synaptogenesis in human cerebral cortex. J. Comp. Neurol. 1997;387(2):167–178. doi: 10.1002/(sici)1096-9861(19971020)387:2<167::aid-cne1>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- Izquierdo A., Suda R.K., Murray E.A. Comparison of the effects of bilateral orbital prefrontal cortex lesions and amygdala lesions on emotional responses in rhesus monkeys. J. Neurosci. 2005;25(37):8534–8542. doi: 10.1523/JNEUROSCI.1232-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kagan J., Reznick J., Clarke C., Snidman N., Garcia-Coll C. Behavioral inhibition to the unfamiliar. Child Dev. 1984;55(6):2212–2225. [Google Scholar]
- Kalin N.H., Shelton S.E., Davidson R.J. Role of the primate orbitofrontal cortex in mediating anxious temperament. Biol. Psychiatry. 2007;62(10):1134–1139. doi: 10.1016/j.biopsych.2007.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim H., Adolphs R., O’Doherty J.P., Shimojo S. Temporal isolation of neural processes underlying face preference decisions. Proc. Natl. Acad. Sci. U.S.A. 2007;104(46):18253–18258. doi: 10.1073/pnas.0703101104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knudsen E.I. Sensitive periods in the development of the brain and behavior. J. Cogn. Neurosci. 2004;16(8):1412–1425. doi: 10.1162/0898929042304796. [DOI] [PubMed] [Google Scholar]
- Kolb B., Pellis S., Robinson T.E. Plasticity and functions of the orbital frontal cortex. Brain Cogn. 2004;55(1):104–115. doi: 10.1016/S0278-2626(03)00278-1. [DOI] [PubMed] [Google Scholar]
- Konner M. Harvard University Press; Cambridge, MA: 2010. The Evolution of Childhood: Relationships, Emotion, Mind. [Google Scholar]
- Kringelbach M.L., Rolls E.T. The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neuropsychology. Prog. Neurobiol. 2004;72(5):341–372. doi: 10.1016/j.pneurobio.2004.03.006. [DOI] [PubMed] [Google Scholar]
- La Greca A.M., Harrison H.M. Adolescent peer relations, friendships, and romantic relationships: do they predict social anxiety and depression? J. Clin. Child Adolesc. Pschol. 2005;34(1):49–61. doi: 10.1207/s15374424jccp3401_5. [DOI] [PubMed] [Google Scholar]
- La Greca A.M., Stone W.L. Social Anxiety Scale for Children – Revised: factor structure and concurrent validity. J. Clin. Child Psychol. 1993;22(1):17–27. [Google Scholar]
- Lee K.H., Siegle G.J. Common and distinct brain networks underlying explicit emotional evaluation: a meta-analytic study. Soc. Cogn. Affect. Neurosci. 2009 doi: 10.1093/scan/nsp001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitman D.I., Wolf D.H., Loughead J., Valdez J.N., Kohler C.G., Brensinger C. Ventrolateral prefrontal cortex and the effects of task demand context on facial affect appraisal in schizophrenia. Soc. Cogn. Affect. Neurosci. 2010 doi: 10.1093/scan/nsq018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luna B., Thulborn K.R., Munoz D.P., Merriam E.P., Garver K.E., Minshew N.J. Maturation of widely distributed brain function subserves cognitive development. Neuroimage. 2001;13(5):786–793. doi: 10.1006/nimg.2000.0743. [DOI] [PubMed] [Google Scholar]
- Machado C.J., Bachevalier J. The impact of selective amygdala, orbital frontal cortex, or hippocampal formation lesions on established social relationships in rhesus monkeys (Macaca mulatta) Behav. Neurosci. 2006;120(4):761–786. doi: 10.1037/0735-7044.120.4.761. [DOI] [PubMed] [Google Scholar]
- Maheu F.S., Dozier M., Guyer A.E., Mandell D., Peloso E., Poeth K., Jenness J., Lau J.Y., Ackerman J.P., Pine D.S., Ernst M. A preliminary study of the mdeial temporal lobe in youths with a history of caregiver deprivation and emotional neglect. Cogn. Affect. Behav. Neurosci. 2010;10(1):34–39. doi: 10.3758/CABN.10.1.34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsh A.A., Blair K.S., Jones M.M., Soliman N., Blair R.J. Dominance and submission: the ventrolateral prefrontal cortex and responses to status cues. J. Cogn. Neurosci. 2009;21(4):713–724. doi: 10.1162/jocn.2009.21052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maslowsky J., Mogg K., Bradley B.P., McClure-Tone E., Ernst M., Pine D.S. A preliminary investigation of neural correlates of treatment in adolescents with generalized anxiety disorder. J. Child Adolesc. Psychopharmacol. 2010;20(2):105–111. doi: 10.1089/cap.2009.0049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCabe K., Houser D., Ryan L., Smith V., Trouard T. A functional imaging study of cooperation in two-person reciprocal exchange. Proc. Natl. Acad. Sci. U.S.A. 2001;98(20):11832–11835. doi: 10.1073/pnas.211415698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClure S.M., Laibson D.I., Loewenstein G., Cohen J.D. Separate neural systems value immediate and delayed monetary rewards. Science. 2004;306(5695):503–507. doi: 10.1126/science.1100907. [DOI] [PubMed] [Google Scholar]
- McClure S.M., Ericson K.M., Laibson D.I., Loewenstein G., Cohen J.D. Time discounting for primary rewards. J. Neurosci. 2007;27(21):5796–5804. doi: 10.1523/JNEUROSCI.4246-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minagawa-Kawai Y., Matsuoka S., Dan I., Naoi N., Nakamura K., Kojima S. Prefrontal activation associated with social attachment: facial-emotion recognition in mothers and infants. Cereb. Cortex. 2009;19(2):284–292. doi: 10.1093/cercor/bhn081. [DOI] [PubMed] [Google Scholar]
- Mischel W., Shoda Y., Rodriguez M.I. Delay of gratification in children. Science. 1989;244(4907):933–938. doi: 10.1126/science.2658056. [DOI] [PubMed] [Google Scholar]
- Mischel W., Ayduk O., Berman M.G., Casey B.J., Gotlib I.H., Jonides J. ‘Willpower’ over the life span: decomposing self-regulation. Soc. Cogn. Affect. Neurosci. 2010 doi: 10.1093/scan/nsq081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moll J., Krueger F., Zahn R., Pardini M., de Oliveira-Souza R., Grafman J. Human fronto-mesolimbic networks guide decisions about charitable donation. Proc. Natl. Acad. Sci. U.S.A. 2006;103(42):15623–15628. doi: 10.1073/pnas.0604475103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monk C.S., McClure E.B., Nelson E.E., Zarahn E., Bilder R.M., Leibenluft E. Adolescent immaturity in attention-related brain engagement to emotional facial expressions. Neuroimage. 2003;20(1):420–428. doi: 10.1016/s1053-8119(03)00355-0. [DOI] [PubMed] [Google Scholar]
- Monk C.S., Nelson E.E., McClure E.B., Mogg K., Bradley B.P., Leibenluft E. Ventrolateral prefrontal cortex activation and attentional bias in response to angry faces in adolescents with generalized anxiety disorder. Am. J. Psychiatry. 2006;163(6):1091–1097. doi: 10.1176/ajp.2006.163.6.1091. [DOI] [PubMed] [Google Scholar]
- Monk C.S., Telzer E.H., Mogg K., Bradley B.P., Mai X., Louro H.M. Amygdala and ventrolateral prefrontal cortex activation to masked angry faces in children and adolescents with generalized anxiety disorder. Arch. Gen. Psychiatry. 2008;65(5):568–576. doi: 10.1001/archpsyc.65.5.568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moriguchi Y., Ohnishi T., Mori T., Matsuda H., Komaki G. Changes of brain activity in the neural substrates for theory of mind during childhood and adolescence. Psychiatry Clin. Neurosci. 2007;61(4):355–363. doi: 10.1111/j.1440-1819.2007.01687.x. [DOI] [PubMed] [Google Scholar]
- Murray E.A., Wise S.P. Interactions between orbital prefrontal cortex and amygdala: advanced cognition, learned responses and instinctive behaviors. Curr. Opin. Neurobiol. 2010;20(2):212–220. doi: 10.1016/j.conb.2010.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray E.A., O’Doherty J.P., Schoenbaum G. What we know and do not know about the functions of the orbitofrontal cortex after 20 years of cross-species studies. J. Neurosci. 2007;27(31):8166–8169. doi: 10.1523/JNEUROSCI.1556-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nelson E.E., Leibenluft E., McClure E.B., Pine D.S. The social re-orientation of adolescence: a neuroscience perspective on the process and its relation to psychopathology. Psychol. Med. 2005;35:163–174. doi: 10.1017/s0033291704003915. [DOI] [PubMed] [Google Scholar]
- Nitschke J.B., Nelson E.E., Rusch B.D., Fox A.S., Oakes T.R., Davidson R.J. Orbitofrontal cortex tracks positive mood in mothers viewing pictures of their newborn infants. Neuroimage. 2004;21(2):583–592. doi: 10.1016/j.neuroimage.2003.10.005. [DOI] [PubMed] [Google Scholar]
- Ochsner K.N., Ray R.D., Cooper J.C., Robertson E.R., Chopra S., Gabrieli J.D. For better or for worse: neural systems supporting the cognitive down- and up-regulation of negative emotion. Neuroimage. 2004;23(2):483–499. doi: 10.1016/j.neuroimage.2004.06.030. [DOI] [PubMed] [Google Scholar]
- O’Doherty J., Critchley H., Deichmann R., Dolan R.J. Dissociating valence of outcome from behavioral control in human orbital and ventral prefrontal cortices. J. Neurosci. 2003;23(21):7931–7939. doi: 10.1523/JNEUROSCI.23-21-07931.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Doherty J., Winston J., Critchley H., Perrett D., Burt D.M., Dolan R.J. Beauty in a smile: the role of medial orbitofrontal cortex in facial attractiveness. Neuropsychologia. 2003;41(2):147–155. doi: 10.1016/s0028-3932(02)00145-8. [DOI] [PubMed] [Google Scholar]
- O’Doherty J.P., Hampton A., Kim H. Model-based fMRI and its application to reward learning and decision making. Ann. N. Y. Acad. Sci. 2007;1104:35–53. doi: 10.1196/annals.1390.022. [DOI] [PubMed] [Google Scholar]
- O’Donnell S., Noseworthy M.D., Levine B., Dennis M. Cortical thickness of the frontopolar area in typically developing children and adolescents. Neuroimage. 2005;24(4):948–954. doi: 10.1016/j.neuroimage.2004.10.014. [DOI] [PubMed] [Google Scholar]
- Okamoto M., Dan H., Clowney L., Yamaguchi Y., Dan I. Activation in ventro-lateral prefrontal cortex during the act of tasting: an fNIRS study. Neurosci. Lett. 2009;451(2):129–133. doi: 10.1016/j.neulet.2008.12.016. [DOI] [PubMed] [Google Scholar]
- Padoa-Schioppa C., Assad J.A. Neurons in the orbitofrontal cortex encode economic value. Nature. 2006;441(7090):223–226. doi: 10.1038/nature04676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Passarotti A.M., Sweeney J.A., Pavuluri M.N. Neural correlates of incidental and directed facial emotion processing in adolescents and adults. Soc. Cogn. Affect. Neurosci. 2009;4(4):387–398. doi: 10.1093/scan/nsp029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelphrey K.A., Morris J.P., McCarthy G. Grasping the intentions of others: the perceived intentionality of an action influences activity in the superior temporal sulcus during social perception. J. Cogn. Neurosci. 2004;16(10):1706–1716. doi: 10.1162/0898929042947900. [DOI] [PubMed] [Google Scholar]
- Petrides M. Lateral prefrontal cortex: architectonic and functional organization. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 2005;360(1456):781–795. doi: 10.1098/rstb.2005.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeifer J.H., Lieberman M.D., Dapretto M. “I know you are but what am I?!”: neural bases of self- and social knowledge retrieval in children and adults. J. Cogn. Neurosci. 2007;19(8):1323–1337. doi: 10.1162/jocn.2007.19.8.1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan K.L., Fitzgerald D.A., Nathan P.J., Moore G.J., Uhde T.W., Tancer M.E. Neural substrates for voluntary suppression of negative affect: a functional magnetic resonance imaging study. Biol. Psychiatry. 2005;57(3):210–219. doi: 10.1016/j.biopsych.2004.10.030. [DOI] [PubMed] [Google Scholar]
- Phan K.L., Sripada C.S., Angstadt M., McCabe K. Reputation for reciprocity engages the brain reward center. Proc. Natl. Acad. Sci. U.S.A. 2010;107(29):13099–13104. doi: 10.1073/pnas.1008137107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pine D.S., Helfinstein S.M., Bar-Haim Y., Nelson E., Fox N.A. Challenges in developing novel treatments for childhood disorders: lessons from research on anxiety. Neuropsychopharmacology. 2009;34(1):213–228. doi: 10.1038/npp.2008.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price J.L. Definition of the orbital cortex in relation to specific connections with limbic and visceral structures and other cortical regions. Ann. N. Y. Acad. Sci. 2007;1121:54–71. doi: 10.1196/annals.1401.008. [DOI] [PubMed] [Google Scholar]
- Rangel A., Camerer C., Montague P.R. A framework for studying the neurobiology of value-based decision making. Nat. Rev. Neurosci. 2008;9(7):545–556. doi: 10.1038/nrn2357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rapee R.M., Heimberg R.G. A cognitive-behavioral model of anxiety in social phobia. Behav. Res. Ther. 1997;35(8):741–756. doi: 10.1016/s0005-7967(97)00022-3. [DOI] [PubMed] [Google Scholar]
- Raznahan A., Lee Y., Stidd R., Long R., Greenstein D., Clasen L. Longitudinally mapping the influence of sex and androgen signaling on the dynamics of human cortical maturation in adolescence. Proc. Natl. Acad. Sci. U.S.A. 2010;107(39):16988–16993. doi: 10.1073/pnas.1006025107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rolls E.T., McCabe C., Redoute J. Expected value, reward outcome, and temporal difference error representations in a probabilistic decision task. Cereb. Cortex. 2008;18(3):652–663. doi: 10.1093/cercor/bhm097. [DOI] [PubMed] [Google Scholar]
- Rubia K., Smith A.B., Woolley J., Nosarti C., Heyman I., Taylor E. Progressive increase of frontostriatal brain activation from childhood to adulthood during event-related tasks of cognitive control. Hum. Brain Mapp. 2006;27(12):973–993. doi: 10.1002/hbm.20237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rubia K., Smith A.B., Taylor E., Brammer M. Linear age-correlated functional development of right inferior fronto-striato-cerebellar networks during response inhibition and anterior cingulate during error-related processes. Hum. Brain Mapp. 2007;28(11):1163–1177. doi: 10.1002/hbm.20347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudebeck P.H., Murray E.A. Amygdala and orbitofrontal cortex lesions differentially influence choices during object reversal learning. J. Neurosci. 2008;28(33):8338–8343. doi: 10.1523/JNEUROSCI.2272-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rygula R., Walker S.C., Clarke H.F., Robbins T.W., Roberts A.C. Differential contributions of the primate ventrolateral prefrontal and orbitofrontal cortex to serial reversal learning. J. Neurosci. 2010;30(43):14552–14559. doi: 10.1523/JNEUROSCI.2631-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakagami M., Pan X. Functional role of the ventrolateral prefrontal cortex in decision making. Curr. Opin. Neurobiol. 2007;17(2):228–233. doi: 10.1016/j.conb.2007.02.008. [DOI] [PubMed] [Google Scholar]
- Sanfey A.G., Rilling J.K., Aronson J.A., Nystrom L.E., Cohen J.D. The neural basis of economic decision-making in the Ultimatum Game. Science. 2003;300(5626):1755–1758. doi: 10.1126/science.1082976. [DOI] [PubMed] [Google Scholar]
- Schlaggar B.L., Brown T.T., Lugar H.M., Visscher K.M., Miezin F.M., Petersen S.E. Functional neuroanatomical differences between adults and school-age children in the processing of single words. Science. 2002;296(5572):1476–1479. doi: 10.1126/science.1069464. [DOI] [PubMed] [Google Scholar]
- Schoenbaum G., Roesch M.R., Stalnaker T.A., Takahashi Y.K. A new perspective on the role of the orbitofrontal cortex in adaptive behaviour. Nat. Rev. Neurosci. 2009;10(12):885–892. doi: 10.1038/nrn2753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmons J.M., Minamimoto T., Murray E.A., Richmond B.J. Selective ablations reveal that orbital and lateral prefrontal cortex play different roles in estimating predicted reward value. J. Neurosci. 2010;30(47):15878–15887. doi: 10.1523/JNEUROSCI.1802-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Souza M.J., Donohue S.E., Bunge S.A. Controlled retrieval and selection of action-relevant knowledge mediated by partially overlapping regions in left ventrolateral prefrontal cortex. Neuroimage. 2009;46(1):299–307. doi: 10.1016/j.neuroimage.2009.01.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinberg L. Cognitive and affective development in adolescence. Trends Cogn. Sci. 2005;9(2):69–74. doi: 10.1016/j.tics.2004.12.005. [DOI] [PubMed] [Google Scholar]
- Steinberg L., Graham S., O’Brien L., Woolard J., Cauffman E., Banich M. Age differences in future orientation and delay discounting. Child Dev. 2009;80(1):28–44. doi: 10.1111/j.1467-8624.2008.01244.x. [DOI] [PubMed] [Google Scholar]
- Stevens M.C., Skudlarski P., Pearlson G.D., Calhoun V.D. Age-related cognitive gains are mediated by the effects of white matter development on brain network integration. Neuroimage. 2009;48(4):738–746. doi: 10.1016/j.neuroimage.2009.06.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiles J. Harvard University Press; Cambridge, MA: 2008. The Fundamentals of Brain Devleopment: Integrating Nature and Nurture. [Google Scholar]
- Toga A.W., Thompson P.M., Sowell E.R. Mapping brain maturation. Trends Neurosci. 2006;29(3):148–159. doi: 10.1016/j.tins.2006.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tottenham N., Sheridan M.A. A review of adversity, the amygdala and the hippocampus: a consideration of developmental timing. Front. Hum. Neurosci. 2009;3:68. doi: 10.3389/neuro.09.068.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tricomi E., Rangel A., Camerer C.F., O’Doherty J.P. Neural evidence for inequality-averse social preferences. Nature. 2010;463(7284):1089–1091. doi: 10.1038/nature08785. [DOI] [PubMed] [Google Scholar]
- Wang A.T., Lee S.S., Sigman M., Dapretto M. Developmental changes in the neural basis of interpreting communicative intent. Soc. Cogn. Affect. Neurosci. 2006;1(2):107–121. doi: 10.1093/scan/nsl018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittle S., Yap M.B., Yucel M., Sheeber L., Simmons J.G., Pantelis C. Maternal responses to adolescent positive affect are associated with adolescents’ reward neuroanatomy. Soc. Cogn. Affect. Neurosci. 2009;4(3):247–256. doi: 10.1093/scan/nsp012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winston J.S., O’Doherty J., Kilner J.M., Perrett D.I., Dolan R.J. Brain systems for assessing facial attractiveness. Neuropsychologia. 2007;45(1):195–206. doi: 10.1016/j.neuropsychologia.2006.05.009. [DOI] [PubMed] [Google Scholar]
- Yap M.B., Whittle S., Yucel M., Sheeber L., Pantelis C., Simmons J.G., Allen N.B. Interactions of parenting experience and brain structure in the prediction of depression in adolescents. Arch. Gen. Psychiatry. 2008;65(12):1377–1385. doi: 10.1001/archpsyc.65.12.1377. [DOI] [PubMed] [Google Scholar]