

Abstract
The objective of this work was to select the most suitable procedure to immobilize lipase from Penicillium camembertii (Lipase G). Different techniques and supports were evaluated, including physical adsorption on hydrophobic supports octyl-agarose, poly(hydroxybutyrate) and Amberlite resin XAD-4; ionic adsorption on the anionic exchange resin MANAE-agarose and covalent attachment on glyoxyl-agarose, MANAE-agarose cross-linked with glutaraldehyde, MANAE-agarose-glutaraldehyde, and epoxy-silica-polyvinyl alcohol composite. Among the tested protocols, the highest hydrolytic activity (128.2 ± 8.10 IU·g−1 of support) was achieved when the lipase was immobilized on epoxy-SiO2-PVA using hexane as coupling medium. Lipase immobilized by ionic adsorption on MANAE-agarose also gave satisfactory result, attaining 55.6 ± 2.60 IU·g−1 of support. In this procedure, the maximum loading of immobilized enzyme was 9.3 mg·g−1 of gel, and the highest activity (68.8 ± 2.70 IU·g−1 of support) was obtained when 20 mg of protein·g−1 was offered. Immobilization carried out in aqueous medium by physical adsorption on hydrophobic supports and covalent attachment on MANAE-agarose-glutaraldehyde and glyoxyl-agarose was shown to be unfeasible for Lipase G. Thermal stability tests revealed that the immobilized derivative on epoxy-SiO2-PVA composite using hexane as coupling medium had a slight higher thermal stability than the free lipase.
1. Introduction
Lipases (triacylglycerol acyl hydrolases EC 3.1.1.3) are hydrolases that act on carboxylic ester bonds. The natural physiologic role of lipases is the hydrolysis of triglycerides into fatty acids and glycerol, but they can also catalyze esterifications and interesterifications in nonaqueous media [1–5]. A peculiarity mechanism action of lipases is the interfacial activation. Most lipases have a α-helical oligopeptide structure covering their active site (lid or flap) and making them inaccessible to substrates. In the absence of a hydrophobic interface, the active site is secluded from the reaction medium, and the enzyme is in the so-called “closed conformation.” However, in the presence of a hydrophobic interface (a drop of oil), the lipase changes its conformation and exposes the catalytic site to the hydrophobic phase, yielding the “open conformation” [6–9].
The limitations of the industrial use of lipases have been mainly due to their high cost, which may be overcome by immobilization techniques on solid supports. Immobilization facilitates the separation of products and provides more flexibility with enzyme/substrate contact by using various reactor configurations. Moreover, immobilization on solid supports may improve enzyme features, from stability to selectivity [10–14]. Lipases have been immobilized by using different protocols as physical adsorption on hydrophobic and ionic exchange resins, covalent attachment on highly activated supports and encapsulation in organic matrices [15–22]. Covalent attachment of lipases on highly activated supports (e.g., with epoxy or aldehyde groups) may promote an intense rigidification of their three-dimensional structure. The relative distances among all residues involved in the covalent immobilization have to be maintained during any conformational change induced by any distorting agent, for example, solvents and temperature [15–19]. Lipases can be also purified, immobilized, and stabilized via interfacial activation on hydrophobic supports. These enzymes are strongly adsorbed onto hydrophobic interfaces through the lid that covers their active site, recognizing these supports as their natural substrate (hydrophobic oil interfaces). This technique is easy, cheap, and allows facile recycling of the support at the end of the life of the enzyme [9, 20–22]. Enzymes can be also reversibly immobilized on ionic exchange supports. Stability of lipases immobilized on these supports can be significantly improved. After inactivation, the enzymes, including lipases, can be also fully desorbed from the support, and then the support can be reused for several cycles [13, 23].
This work deals with the selection of a suitable procedure to immobilize lipase from Penicillium camembertii (Lipase G). In order to achieve this goal, different protocols such as physical adsorption on octyl-agarose, poly(hydroxybutyrate) (PHB), and amberlite resin XAD-4; ionic adsorption on anionic exchange resin MANAE-agarose and covalent attachment on glyoxyl-agarose, MANAE-agarose cross-linked with glutaraldehyde, MANAE-agarose-glutaraldehyde, and epoxy-silica-polyvinyl alcohol composite (epoxy-SiO2-PVA) were tested.
Lipase G is a lipolytic enzyme preparation manufactured by Amano Enzyme Inc. by a selected strain belonging to Penicillium camembertii, which has unique substrate restricted to mono- and diglycerides reacting mainly with medium-chain molecules [24, 25]. This enzyme shows no activity towards triacylglycerols such as tripropionin, tributyrin, trioctanoin, and olive oil [26]. This lipase has a single polypeptide chain consisting of 276 amino acid residues with two short disulfide bridges (Cys36–Cys41 and Cys103–Cys106) and molecular weight of 30 kDa. The catalytic triad of Asp, His, and Ser, was conserved at positions 199, 259, and 145, respectively [25]. Penicillium camembertii lipase has a natural trend to form bimolecular aggregates by interaction between the hydrophobic surfaces surrounding the active centers via numerous hydrogen bonds and salt bridges [27]. This lipase has shown consistent high production of monoglyceride in the direct acylation of glycerol with fatty acids having different chain lengths [18]. Although several reports have been published using this lipase preparation [17, 18, 26], data relating its immobilization and the performance of the resulting immobilized derivative is still scarce in the literature.
2. Materials and Methods
2.1. Materials
Penicillium camembertii lipase (Lipase G) was purchased from Amano Enzyme (Nagoya, Japan), having protein concentration of 10 mg·g−1 of powder and specific activity of 85.9 ± 1.80 IU·mg−1 of protein. The lipase preparation was used without further purification. Amberlite resin XAD-4 was acquired from Fluka (Buchs, Swiss) and poly(hydroxybutyrate) (PHB) from PHB Industrial S.A. (São Paulo, Brazil). Agarose CL-6B (Sepharose CL-6B) and octyl-agarose CL-4B (40 μmoL of octyl groups·mL−1 of gel) were purchased from Pharmacia Biotech (Uppsala, Sweden). The characteristics of supports used in the present work are presented in Table 1. Glyoxyl-agarose (95 μmoL aldehyde·g−1 of support) was prepared as previously described [28]. MANAE-agarose was prepared as previously described [29]. MANAE-agarose-glutaraldehyde was prepared activating MANAE-agarose with glutaraldehyde dimeric form as previously described [30]. Tetraethylorthosilicate (TEOS) was purchased from Sigma-Aldrich Chemicals Co. (Milwaukee, Wis, USA). Epichlorohydrin, hydrochloric acid (minimum 36%), and polyethylene glycol (PEG, molecular weight 1500) were supplied by Reagen (Rio de Janeiro, RJ, Brazil). All other reagents used were of analytical grade.
Table 1.
Characteristics of the tested supports for immobilizing Lipase G by different protocols.
| Support | Basic structure | Particle diameter (μm) | Pore volume (mL·g−1) | Surface area (m2·g−1) |
|---|---|---|---|---|
| Amberlite XAD-4a | Macroreticular cross-linked aromatic polymer | 490–690 | 0.500 | ≥750 |
| Octyl-agarose CL-4Bb | Cross-linked agarose via stable ether linkages, 4% | 45–165 | — | — |
| PHBc | Polyester | 75–90 | — | — |
| SiO2-PVAd | Polysiloxane-polyvinyl alcohol hybrid matrix | 175 | 0.275 | 461 |
| Agarose CL-6Be | Cross-linked agarose via stable ether linkages, 6% | 45–165 | — | 25 |
2.2. Preparation of Epoxy-SiO2-PVA
Polysiloxane-polyvinyl alcohol hybrid composite was prepared by hydrolysis and polycondensation of tetraethylorthosilicate (TEOS) according to the methodology previously described [31]. The reagents TEOS (5 mL), ethanol (5 mL), and polyvinyl alcohol (PVA) solution 2% (w/v) (6 mL) were carefully mixed and stirred for 5 min at 60°C, followed by the addition of 0.1 mL of concentrated HCl in order to catalyze the reaction. After 40 min, the solution was transferred to microwells of tissue culture plates (disc shape) and kept at room temperature until complete gel solidification (formation of the interpenetrated network of SiO2-PVA). Then, the spheres were ground in a ball mill to attain particles with 80 MESH (Tyler standard). Activation of SiO2-PVA particles was carried out with epichlorohydrin 2.5% v/v at pH 7.0 for 1 h at room temperature, followed by exhaustive washings with distilled water [17].
2.3. Immobilization Protocols
2.3.1. Physical Adsorption
1.0 g of octyl-agarose CL-4B, amberlite resin XAD-4, or PHB were soaked into 5 mL of ethanol 96% (v/v) for 30 min, according to the methodology described by Adlercreutz [32]. The supports were filtered and incubated with 20 mL of 5 mmoL·L−1 buffer Tris-HCl pH 7.0 containing 0.25 mg protein·mL−1 for 24 h. The immobilized derivatives were recovered by vacuum filtration.
2.3.2. Ionic Adsorption
Powder lipase preparation was dissolved in 10 mL of 2.5 mmoL·L−1 buffer Tris-HCl pH 7.0 and mixed with 1.0 g of MANAE-agarose under mild stirring for 1 h at room temperature. MANAE-agarose was tested by offering different protein loadings (5.0 to 30·0 mg·g−1 of support) to determine the support saturation. The immobilized derivatives were recovered by vacuum filtration.
2.3.3. Covalent Attachment
In Aqueous Medium —
Powder lipase preparation was dissolved in 10 mL of 100 mmoL·L−1 of bicarbonate buffer pH 10.05 and mixed with 1.0 g of glyoxyl-agarose under mild stirring for 1 h at room temperature. Immobilization on MANAE-agarose-glutaraldehyde or SiO2-PVA composite previously activated with epichlorohydrin (epoxy-SiO2-PVA) was performed by incubating 1.0 g of support in 10 mL of buffer Tris-HCl (2.5 and 100 mmoL·L−1) pH 7.0 under mild stirring for 1 h at room temperature, offering protein loading of 5 mg·g−1 of support. Immobilization by ionic adsorption on MANAE-agarose followed by cross-linking with glutaraldehyde was performed by incubating 1.0 g of the immobilized lipase on MANAE-agarose in 9 mL of 1% (v/v) glutaraldehyde solution in buffer Tris-HCl 5 mmoL·L−1 pH 7.0 at room temperature for 40 min under mild stirring. The derivatives were then filtered and washed thoroughly with Milli-Q water.
In Organic Medium —
epoxy-SiO2-PVA composite (1.0 g) was soaked into hexane under stirring (100 rpm) for 1 h at room temperature. Then, excess of hexane was removed, and lipase was added at a ratio of 1 : 4 gram of enzyme per gram of support (2.5 mg of protein·g−1 of support). PEG-1500 was added together with the enzyme solution at a fixed amount (100 μL·g−1 of support). Lipase-support system was maintained in contact for 16 h at 4°C under static conditions. The immobilized derivative was filtered in nylon membrane and rinsed thoroughly with hexane [31].
2.4. Determination of Hydrolytic Activity
Hydrolytic activities of soluble and immobilized lipases were assayed using vinyl propionate as substrate, according to the methodology described by Chahinian et al. [33]. One international unit of activity was defined as the amount of enzyme that releases 1 μmoL of propionic acid per minute (1 IU) under the assay conditions.
2.5. Determination of Protein
Protein was determined according to the methodology described by Bradford [34] using bovine serum albumin (BSA) as standard.
2.6. Immobilization Parameters
Immobilized protein (IP) was calculated by determining the amount of protein disappeared from the supernatant and comparing to the initial protein concentration offered (mg·g−1 of support). Recovered activity percentage (RA) was calculated by determining the activity of the immobilized enzyme (apparent hydrolytic activity) and comparing with the number of enzyme units that disappeared from the supernatant (theoretically immobilized).
2.7. Thermal Studies of Free and Immobilized Lipase
The effect of temperature on the activity of free and immobilized lipase was determined at temperature range from 25 to 60°C. For thermal stability tests, both free and immobilized lipase preparations were incubated in buffer Tris-HCl pH 7.0 at 40 or 50°C for 2 h. Samples were removed and assayed for residual activity as previously described (hydrolysis of vinyl propionate), taking an unheated control to be 100% active.
3. Results and Discussion
3.1. Immobilization Parameters for Lipase G on Different Supports and Protocols
Different protocols to immobilize Lipase G on different supports were screened. All immobilization procedures were carried out in aqueous medium, except when epoxy-SiO2-PVA composite was used. For this matrix, the lipase was immobilized in both aqueous and organic media. Table 2 displays the immobilization parameters in terms of immobilized protein (IP), hydrolytic activity (HA), and recovered activity (RA) for each procedure evaluated.
Table 2.
Immobilization parameters for Lipase G on different supports and protocols.
| Immobilization protocols | Support | Activating agent | IP (mg·g−1) | HA (IU·g−1) | RA (%) |
|---|---|---|---|---|---|
| Ionic adsorption | MANAE-agarose* | none | 4.52 ± 0.18 | 55.6 ± 2.60 | 14.2 ± 0.40 |
| covalent attachment | Glyoxyl-agarose* | glycidol | 4.30 ± 0.20 | 0 | 0 |
| MANAE-agarose* | glutaraldehydea | 4.47 ± 0.12 | 0 | 0 | |
| MANAE-agarose* | glutaraldehydeb | 4.52 ± 0.18 | 31.2 ± 4.50 | 8.20 ± 0.80 | |
| MANAE-agarose* | glutaraldehydec | 4.05 ± 0.05 | 0 | 0 | |
| MANAE-agarose* | glutaraldehyded | 4.09 ± 0.08 | 0 | 0 | |
| Epoxy-SiO2-PVA* | epichlorohydrind | 3.00 ± 0.04 | 9.50 ± 0.30 | 4.00 ± 0.20 | |
| Epoxy-SiO2-PVA** | epichlorohydrin | 2.50 ± 0.02 | 128.2 ± 8.10 | 60.9 ± 5.30 |
IP: Immobilized protein; HA: hydrolytic activity; RA: recovered activity.
*Immobilization in aqueous medium.
**Immobilization in organic medium (hexane).
aCovalent attachment in 100 mmoL·L−1 buffer phosphate at pH 7.0.
bDerivative prepared by ionic adsorption cross-linked with glutaraldehyde solution.
cCovalent attachment in 100 mmoL·L−1 buffer Tris-HCl at pH 7.0.
dCovalent attachment in 2.5 mmoL·L−1 buffer Tris-HCl at pH 7.0.
The first methodology to be tested was the physical adsorption on highly hydrophobic supports with different characteristics such as octyl-agarose, PHB, and amberlite XAD-4 (Table 1) to take advantage of the complex mechanism of lipases as a tool that allows the immobilization via hydrophobic interaction at very low ionic strength [9, 20, 21]. Lipases recognize these surfaces similarly to those of their natural substrates (drops of oil), yielding immobilized derivative with open and hyperactivated structures [9, 21]. However, Lipase G was not immobilized by selective adsorption at very low ionic strength on three different hydrophobic supports tested, in agreement with data previously reported using hydrophobic matrix polypropylene [35]. Under these conditions, the aggregates of Lipase G formed by strong interaction of hydrophobic pockets surrounding the active site of lipase molecules in open conformation were not displaced to monomeric form (dissociation of the dimmers).
The most common and easy protocol to perform reversible immobilization of enzymes with an easy regeneration of the support is the ionic adsorption on anionic exchange resins such as MANAE-agarose [13, 23]. Figure 1(a) shows the immobilization course of the lipase G on MANAE-agarose by ionic adsorption. The procedure was quite fast, and after 60 min of contact between support and enzyme, the lipase immobilization was completed, resulting in protein immobilization up to 87% which corresponded to 4.52 ± 0.18 mg protein·g−1 and hydrolytic activity of 55.6 ± 2.60 IU·g−1 gel. Immobilization of lipase from different sources on MANAE-agarose by ionic adsorption has been previously reported to decrease the activity of the enzyme during immobilization procedure [20].
Figure 1.
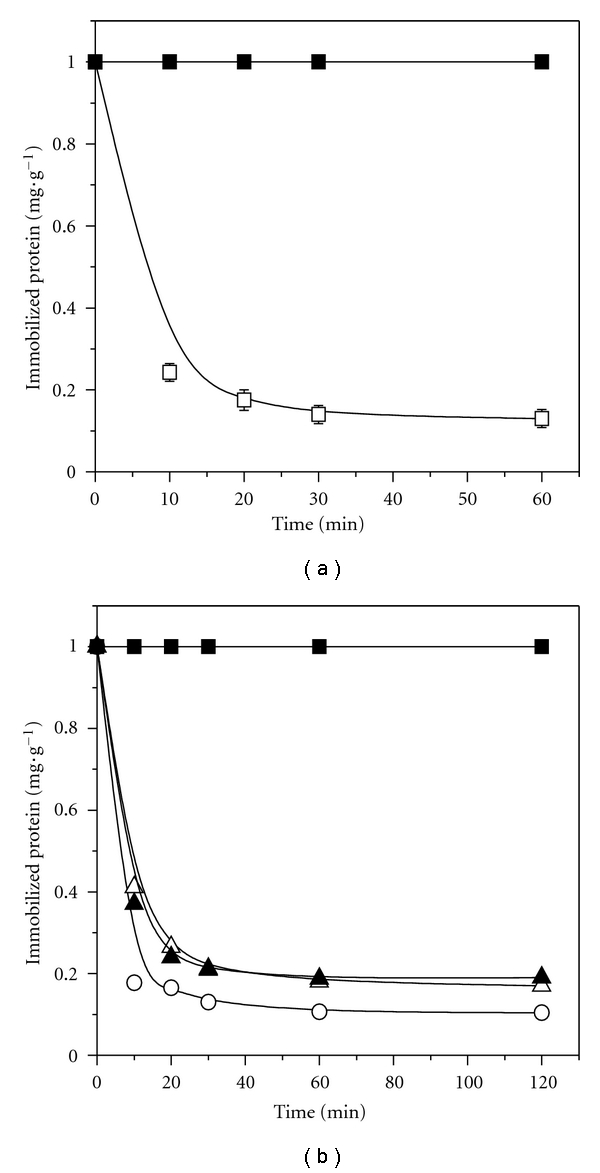
Immobilization course for Lipase G on MANAE-agarose by ionic adsorption (a) and covalent attachment (b) on MANAE-agarose-glutaraldehyde incubated in 100 mmoL·L−1 buffer phosphate pH 7.0 (open circle), 100 mmoL·L−1 buffer Tris-HCl pH 7.0 (open triangle), and 2.5 mmoL·L−1 buffer Tris-HCl pH 7.0 (full triangle) and supernatant (full square).
MANAE-agarose activated with dimmers of glutaraldehyde was used to immobilize Lipase G by covalent attachment at pH 7.0. An attempt was made to increase the activity recovered by evaluating the influence of the buffer (Tris-HCl and phosphate) and ionic strength (2.5 and 100 mmoL·L−1) on the immobilization course of Lipase G on MANAE-agarose-glutaraldehyde. Figure 1(b) shows that for both buffer solutions similar values for protein immobilized were found (4.10 and 4.47 mg·g−1 of support by incubating Lipase G in buffer Tris-HCl and phosphate, resp.). In this set of experiments, no significant effect of the ionic strength Tris-HCl buffer (2.5 and 100 mmoL·L−1) on the kinetic profile of the lipase immobilization was also verified. Although it has been shown that the use of amino compounds, such as Tris-HCl buffer, decreases the immobilization rate of the lipase preparation [36], for Lipase G this was not observed. In addition, screening for a suitable buffer to be used in the hydrolytic assay revealed that the highest hydrolytic activity of Lipase G was detected when incubated in Tris-HCl (85.9 ± 1.80 IU·mg−1 of protein), 6-fold higher than incubating in buffer phosphate (13.7 ± 1.80 IU·mg−1 of protein).
After 30 min of incubation, approximately 80% of the enzyme activity, corresponding to approximately 4.0 mg of protein·g−1 gel, was immobilized. Although high-immobilized protein was attained, using both protocols (lipase covalent attachment on glyoxyl-agarose and MANAE-agarose-glutaraldehyde), no hydrolytic activity was detected on the resulted immobilized derivatives. Such immobilizing protocols have probably led to the inactivation of the enzyme. Covalent interaction is followed by the formation of Schiff's bases (C=N double bond) between aldehyde groups and lysine residues placed on the enzyme surface. This type of interaction is likely to result in undesired mobility restriction of the enzyme or even displacement of the active site which may inactivate the lipase [20].
Multipoint covalent attachment of Lipase G on highly activated glyoxyl-agarose was performed which rendered a derivative with high-immobilized protein concentration (Table 2). However, free and immobilized Lipase G activities were completely inactivated when incubated at pH 10.05.
Covalent attachment of Lipase G ionically adsorbed on MANAE-agarose, followed by cross-linking with glutaraldehyde, was also performed. As can be seen, the hydrolytic activity of the immobilized derivative after cross-linking with glutaraldehyde at low concentration (1% v/v at pH 7.0) decreased from 55.6 ± 2.60 IU·g−1 to 31.2 ± 4.50 IU·g−1 of support which can be attributed to the distortion effect of the three-dimensional structure of the lipase after cross-linking step.
Lipase G was also immobilized by covalent attachment on epoxy-SiO2-PVA composite in aqueous medium (buffer Tris-HCl pH 7.0). Epoxy-SiO2-PVA is a composite with surface area of 461 m2·g−1, pore volume of 0.275 mL·g−1, and particles diameter of 175 μm (Table 1). This composite has been used extensively as a support for lipase from several sources [17, 18, 31, 35]. In this procedure, 60% of the protein initially offered was covalently attached, rendering maximum protein loading of 3.00 ± 0.04 mg·g−1 of support and hydrolytic activity of 9.50 ± 0.30 IU·g−1 of support (Table 2). Under these conditions, independent of the tested support, the immobilization of Lipase G in aqueous medium by covalent attachment appeared to be unsuitable for obtaining high-activity derivatives.
However, a different behavior was attained when the lipase G was immobilized on epoxy-SiO2-PVA composite by covalent attachment using hexane as coupling medium. Under this condition, high hydrolytic activity (128.2 ± 8.10 IU·g−1 of support) was attained which corresponded to 60.9 ± 5.30% of recovered activity. This may be explained by the dissociation of the dimmeric form of the lipase in the presence of a nonmiscible solvent interface. Similar results have been described by several researchers, indicating a new trend in the use of organic apolar solvent as coupling medium for lipase immobilization on different support types [17–19, 22, 31, 35]. Immobilization of lipases from several sources on activated supports using hexane as coupling medium allows high retention of the catalytic activity in biotransformation reactions. For example, lipase from Burkholderia cepacia (BCL) immobilized on epoxy-SiO2-PVA prepared by this methodology was found to have high and stable activity in the synthesis of monoglycerides by glycerolysis of babassu oil in packed-bed reactor with an estimated half-life of 50 days [37]. However, such satisfactory performance depends on the support nature and substrate polarity. In the ester hydrolysis, for instance, Candida rugosa immobilized in zirconium phosphate showed low stability after successive hydrolysis batches by either desorption or solubilization of the aggregates of enzymes in aqueous medium [19]. The immobilization of Lipase G on activated supports using hexane as coupling medium can be a very attractive for further use in nonaqueous medium reactions [18, 35].
Based on these results, the protocols that rendered higher hydrolytic activities were selected for further studies. Although the immobilization of Lipase G on MANAE-agarose by ionic adsorption rendered lower hydrolytic activity (55.6 ± 2.60 IU·g−1 of support) than the immobilized derivative on epoxy-SiO2-PVA (128.2 ± 8.10 IU·g−1 of support), this methodology is expected of have potential to thermal stabilization of the enzyme.
3.2. Maximum Protein Loading Immobilized on MANAE-Agarose by Ionic Adsorption
To determine the maximum protein loading that could be used in the immobilization of Lipase G on MANAE-agarose by ionic adsorption, the amount of protein offered to the support varied from 5 to 30 mg·g−1. Results are displayed in Table 3.
Table 3.
Immobilization parameters for different lipase loadings offered to MANAE-agarose.
| Protein loading (mg·g−1 of support) | IP (mg·g−1) | HA (IU·g−1) | RA (%) |
|---|---|---|---|
| 5 | 4.52 ± 0.18 | 55.6 ± 2.60 | 14.2 ± 0.40 |
| 10 | 6.80 ± 0.50 | 59.2 ± 2.80 | 10.4 ± 0.60 |
| 15 | 9.38 ± 0.21 | 66.7 ± 4.60 | 8.45 ± 0.50 |
| 20 | 9.31 ± 0.16 | 68.8 ± 2.70 | 8.80 ± 0.20 |
| 30 | 9.23 ± 0.10 | 63.8 ± 4.60 | 8.20 ± 0.50 |
IP: immobilized protein; HA: hydrolytic activity; RA: recovered activity.
For the maximum protein loading (30 mg·g−1), 9.23 ± 0.10 mg of protein.g−1 of support was immobilized. The highest activity (68.8 ± 2.70 IU·g−1 of support) was obtained when 20 mg of protein·g−1 was offered. The recovered activity decreased with the increase of the protein loading for all derivatives. The value of this parameter decreased from 14.2 ± 0.43% for the lowest lipase loading (5 mg·g−1 of support) to 8.22 ± 0.50% for the maximum offered loading (30 mg·g−1 of support). The intrinsic velocity, which would be measured in the absence of mass transfer effects, is proportional to the enzyme loading. However, when this loading increases, the rate-controlling step will shift from the intrinsic reaction to pore diffusion, and an intraparticle profile of substrate concentrations would cause a reduction of the effectiveness factor of the biocatalyst beads. In addition, the effective diffusion coefficient within the gel may decrease even more in the presence of a high amount of immobilized enzyme, due to a reduction of the porous effective diameter caused by spatial hindrances [15, 16].
3.3. Thermal Studies of Lipase G Immobilized on Epoxy-SiO2-PVA
The influence of temperature on the hydrolytic activity of free and immobilized Lipase G was assessed in the range from 25 to 60°C on the vinyl propionate hydrolysis (20 mmoL·L−1 buffer Tris-HCl pH 7.0). The optimum temperature for both free and immobilized lipases was found to be at 35°C, as shown in Figure 2. Although no change on the optimum temperature was verified, the immobilized lipase had a slight higher hydrolytic activity in the range from 35 to 45°C, indicating better stability of the three-dimensional structure of the immobilized derivative. Such increase may be explained by the change of the conformational integrity of the enzyme structure upon covalent binding to the support and can be considered as an important feature for a possible industrial application because it allows to reduce substrate viscosity, favoring high-yield process [18].
Figure 2.
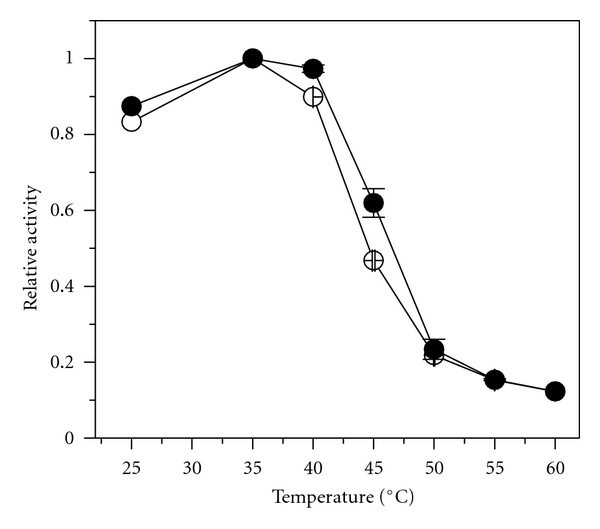
Influence of temperature on the hydrolytic activity of free (⚪) and immobilized lipase G on epoxy-SiO2-PVA composite (●).
The thermal stability of the immobilized lipase was evaluated by measuring the residual activity of lipase G exposed to 40 (Figure 3(a)) and 50°C (Figure 3(b)) in 100 mmoL·L−1 buffer Tris-HCl at pH 7.0 for 2 h. According to Figure 3(a), the free lipase at 40°C shows a half-life (t1/2) of 25.6 min, whereas the half-life of the immobilized lipase at this temperature was 43 min. These results are in agreement with the hypothesis that the conformational flexibility of the enzyme was reduced after immobilization. However, at 50°C, no difference was found between the curve profile for both free and immobilized lipases (Figure 3(b)). Therefore, the immobilization of the lipase on epoxy-SiO2-PVA led to a slight increase of enzyme rigidity at 40°C.
Figure 3.
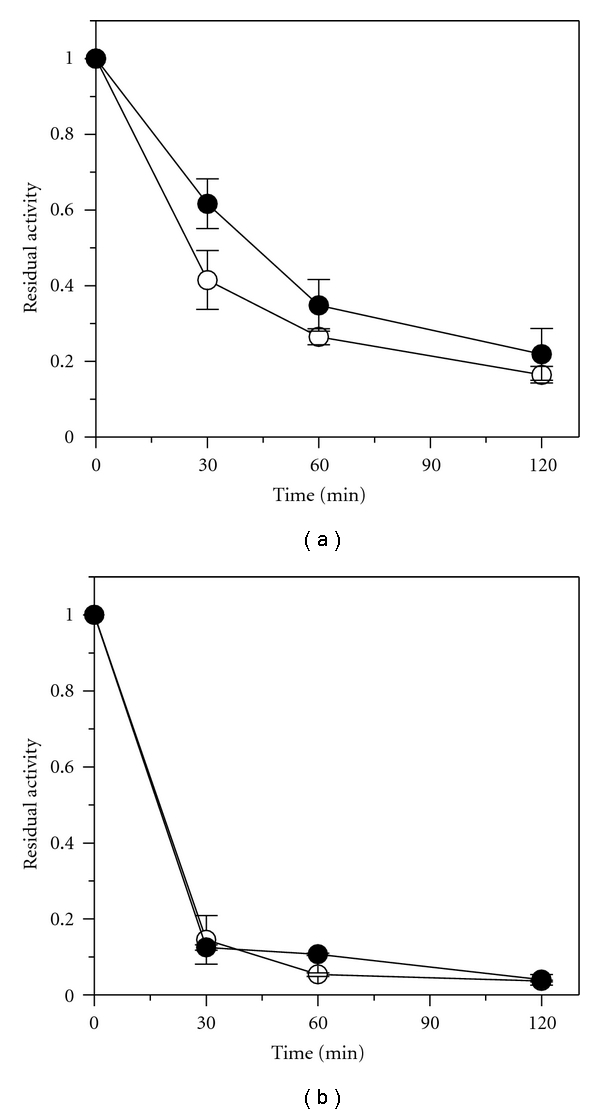
Residual activity for free (⚪) and immobilized Lipase G on epoxy-SiO2-PVA composite (●) incubated at 40 (a) and 50°C (b).
4. Conclusion
In this work, immobilizing procedures to stabilize Lipase G were screened. Different protocols such as physical adsorption, covalent attachment, and ionic adsorption were tested. In aqueous medium, only the immobilization on MANAE-agarose by ionic adsorption gave derivative with satisfactory hydrolytic activity. Immobilization on epoxy-SiO2-PVA by covalent attachment in organic medium was shown to be the most promising protocol for immobilizing Lipase G rendering a derivative with the highest hydrolytic activity. The replacement of buffer by hexane was able to create a microenvironment favorable to the immobilization of the lipase and that enhanced its thermal stability. The immobilization of Lipase G on epoxy-SiO2-PVA composite was shown to be a promising strategy for obtaining an active and stable biocatalyst.
Acknowledgment
The authors gratefully acknowledge CNPq, CAPES, FAPESP, and FAPEMIG (Brazil) for financial support.
References
- 1.Klibanov AM. Improving enzymes by using them in organic solvents. Nature. 2001;409(6817):241–246. doi: 10.1038/35051719. [DOI] [PubMed] [Google Scholar]
- 2.Sharma D, Sharma B, Shukla AK. Biotechnological approach of microbial lipase: a review. Biotechnology. 2011;10(1):23–40. [Google Scholar]
- 3.de Castro HF, Mendes AA, Dos Santos JC, de Aguiar CL. Modification of oils and fats by biotransformation. Quimica Nova. 2004;27(1):146–156. [Google Scholar]
- 4.Hasan F, Shah AA, Hameed A. Industrial applications of microbial lipases. Enzyme and Microbial Technology. 2006;39(2):235–251. [Google Scholar]
- 5.Sharma R, Chisti Y, Banerjee UC. Production, purification, characterization, and applications of lipases. Biotechnology Advances. 2001;19(8):627–662. doi: 10.1016/s0734-9750(01)00086-6. [DOI] [PubMed] [Google Scholar]
- 6.Sarda L, Desnuelle P. Action de la lipase pancreatique sur les esteres en emulsion. Biochimica et Biophysica Acta. 1958;30:513–521. doi: 10.1016/0006-3002(58)90097-0. [DOI] [PubMed] [Google Scholar]
- 7.Verger R. “Interfacial activation” of lipases: facts and artifacts. Trends in Biotechnology. 1997;15(1):32–38. [Google Scholar]
- 8.Reis P, Holmberg K, Watzke H, Leser ME, Miller R. Lipases at interfaces: a review. Advances in Colloid and Interface Science. 2009;147-148:237–250. doi: 10.1016/j.cis.2008.06.001. [DOI] [PubMed] [Google Scholar]
- 9.Bastida A, Sabuquillo P, Armisen P, Fernández-Lafuente R, Huguet J, Guisán JM. A single step purification, immobilization, and hyperactivation of lipases via interfacial adsorption on strongly hydrophobic supports. Biotechnology and Bioengineering. 1998;58(5):486–493. doi: 10.1002/(sici)1097-0290(19980605)58:5<486::aid-bit4>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 10.Hanefeld U, Gardossi L, Magner E. Understanding enzyme immobilisation. Chemical Society Reviews. 2009;38(2):453–468. doi: 10.1039/b711564b. [DOI] [PubMed] [Google Scholar]
- 11.Fjerbaek L, Christensen KV, Norddahl B. A review of the current state of biodiesel production using enzymatic transesterification. Biotechnology and Bioengineering. 2009;102(5):1298–1315. doi: 10.1002/bit.22256. [DOI] [PubMed] [Google Scholar]
- 12.Mateo C, Palomo JM, Fernandez-Lorente G, Guisan JM, Fernandez-Lafuente R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme and Microbial Technology. 2007;40(6):1451–1463. [Google Scholar]
- 13.Mateo C, Abian O, Fernández-Lafuente R, Guisán JM. Reversible enzyme immobilization via a very strong and nondistorting ionic adsorption on support-polyethylenimine composites. Biotechnology and Bioengineering. 2000;68(1):98–105. doi: 10.1002/(sici)1097-0290(20000405)68:1<98::aid-bit12>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 14.Mateo C, Palomo JM, Fuentes M, et al. Glyoxal agarose: a fully inert and hydrophilic support for immobilization and high stabilization of proteins. Enzyme and Microbial Technology. 2006;39:274–280. [Google Scholar]
- 15.Mendes AA, de Castro HF, Rodrigues DS, et al. Multipoint covalent immobilization of lipase on chitosan hybrid hydrogels: influence of the polyelectrolyte complex type and chemical modification on the catalytic properties of the biocatalysts. doi: 10.1007/s10295-010-0880-9. Journal of Industrial Microbiology & Biotechnology. In press. [DOI] [PubMed] [Google Scholar]
- 16.Mendes AA, Giordano RC, Giordano RDLC, de Castro HF. Immobilization and stabilization of microbial lipases by multipoint covalent attachment on aldehyde-resin affinity: application of the biocatalysts in biodiesel synthesis. Journal of Molecular Catalysis B. 2011;68(1):109–115. [Google Scholar]
- 17.Freitas L, Da Rós PCM, Santos JC, de Castro HF. An integrated approach to produce biodiesel and monoglycerides by enzymatic interestification of babassu oil (Orbinya sp) Process Biochemistry. 2009;44(10):1068–1074. [Google Scholar]
- 18.Freitas L, Paula AV, Santos JC, Zanin GM, Castro HF. Enzymatic synthesis of monoglycerides by esterification reaction using Penicillium camembertii lipase immobilized on epoxy SiO2-PVA composite. Journal of Molecular Catalysis B. 2010;65:87–90. [Google Scholar]
- 19.Mendes AA, Barbosa BCM, da Silva MLCP, de Castro HF. Morphological, biochemical and kinetic properties of lipase from Candida rugosa immobilized in zirconium phosphate. Biocatalysis and Biotransformation. 2007;25(5):393–400. [Google Scholar]
- 20.Cunha AG, Fernández-Lorente G, Bevilaqua JV, et al. Immobilization of Yarrowia lipolytica lipase—a comparison of stability of physical adsorption and covalent attachment techniques. Applied Biochemistry and Biotechnology. 2008;146(1–3):49–56. doi: 10.1007/s12010-007-8073-3. [DOI] [PubMed] [Google Scholar]
- 21.Palomo JM, Muñoz G, Fernández-Lorente G, Mateo C, Fernández-Lafuente R, Guisán JM. Interfacial adsorption of lipases on very hydrophobic support (octadecyl-Sepabeads): immobilization, hyperactivation and stabilization of the open form of lipases. Journal of Molecular Catalysis B. 2002;19-20:279–286. [Google Scholar]
- 22.de Oliveira PC, Alves GM, de Castro HF. Immobilisation studies and catalytic properties of microbial lipase onto styrene-divinylbenzene copolymer. Biochemical Engineering Journal. 2000;5(1):63–71. [Google Scholar]
- 23.Filho M, Pessela BC, Mateo C, Carrascosa AV, Fernández-Lafuente R, Guisán JM. Reversible immobilization of a hexameric α-galactosidase from Thermus sp. strain T2 on polymeric ionic exchangers. Process Biochemistry. 2008;43(10):1142–1146. [Google Scholar]
- 24.Yamaguchi S, Takeuchi K, Mase T, et al. The consequences of engineering an extra disulfide bond in the Penicillium camembertii mono- and diglyceride specific lipase. Protein Engineering. 1996;9(9):789–795. doi: 10.1093/protein/9.9.789. [DOI] [PubMed] [Google Scholar]
- 25.Isobe K, Nokihara K. Primary structure determination of mono- and diacylglycerol lipase from Penicillium camembertii. FEBS Letters. 1993;320(2):101–106. doi: 10.1016/0014-5793(93)80071-2. [DOI] [PubMed] [Google Scholar]
- 26.Nini L, Sarda L, Comeau LC, Boitard E, Dubès JP, Chahinian H. Lipase-catalysed hydrolysis of short-chain substrates in solution and in emulsion: a kinetic study. Biochimica et Biophysica Acta. 2001;1534(1):34–44. doi: 10.1016/s1388-1981(01)00172-x. [DOI] [PubMed] [Google Scholar]
- 27.Derewenda U, Swenson L, Green R, et al. Current progress in crystallographic studies of new lipases from filamentous fungi. Protein Engineering. 1994;7(4):551–557. doi: 10.1093/protein/7.4.551. [DOI] [PubMed] [Google Scholar]
- 28.Guisán JM. Aldehyde-agarose gels as activated supports for immobilization-stabilization of enzymes. Enzyme and Microbial Technology. 1988;10(6):375–382. [Google Scholar]
- 29.Fernández-Lafuente R, Rosell CM, Rodriguez V, et al. Preparation of activated supports containing low pK amino groups. A new tool for protein immobilization via the carboxyl coupling method. Enzyme and Microbial Technology. 1993;15(7):546–550. doi: 10.1016/0141-0229(93)90016-u. [DOI] [PubMed] [Google Scholar]
- 30.Betancor L, López-Gallego F, Hidalgo A, et al. Different mechanisms of protein immobilization on glutaraldehyde activated supports: effect of support activation and immobilization conditions. Enzyme and Microbial Technology. 2006;39:877–882. [Google Scholar]
- 31.Paula AV, Moreira ABR, Braga LP, de Castro HF, Bruno LM. Comparative performance of Candida rugosa lipase immobilized on polysiloxane polyvinyl alcohol hybrid support using different methodologies. Quimica Nova. 2008;31(1):35–40. [Google Scholar]
- 32.Adlercreutz P. Immobilization of enzymes for use in organic media. In: Guisan JM, editor. Immobilization of Enzymes and Cells. Totowa, NJ, USA: Humana Press; 2006. pp. 251–256. [Google Scholar]
- 33.Chahinian H, Nini L, Boitard E, Dubès JP, Sarda L, Comeau LC. Kinetic properties of Penicillium cyclopium lipases studied with vinyl esters. Lipids. 2000;35(8):919–925. doi: 10.1007/s11745-000-0601-3. [DOI] [PubMed] [Google Scholar]
- 34.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Analytical Biochemistry. 1976;72(1-2):248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 35.Salis A, Pinna M, Monduzzi M, Solinas V. Comparison among immobilised lipases on macroporous polypropylene toward biodiesel synthesis. Journal of Molecular Catalysis B. 2008;54(1-2):19–26. [Google Scholar]
- 36.Fernández-Lafuente R, Cowan DA, Wood ANP. Hyperstabilization of a thermophilic esterase by multipoint covalent attachment. Enzyme and Microbial Technology. 1995;17:366–372. [Google Scholar]
- 37.Freitas L, dos Santos JC, Zanin GM, de Castro HF. Packed-bed reactor running on babassu oil and glycerol to produce monoglycerides by enzymatic route using immobilized Burkholderia cepacia lipase. Applied Biochemistry and Biotechnology. 2010;161(1–8):372–381. doi: 10.1007/s12010-009-8858-7. [DOI] [PubMed] [Google Scholar]