

Abstract
7,8-Dihydroxyflavone is a recently identified small molecular tropomyosin-receptor-kinase B (TrkB) agonist. Our preliminary structural activity relationship (SAR) study showed that the 7,8-dihydroxy groups are essential for the agonistic effect. To improve the lead compound's agonistic activity, we have conducted an extensive SAR study and synthesized numerous derivatives. We have successfully identified 4'-dimethylamino-7,8-dihydroxyflavone that displays higher TrkB agonistic activity than the lead. This novel compound also exhibits a more robust and longer TrkB activation effect in animals. Consequently, this new compound reveals more potent anti-apoptotic activity. Interestingly, chronic oral administration of 4'-dimethylamino-7,8-dihydroxyflavone and its lead strongly promotes neurogenesis in dentate gyrus and demonstrates marked antidepressant effects. Hence, our data support that the synthetic 4'-dimethylamino-7,8-dihydroxyflavone and its lead both are orally bioavailable TrkB agonists and possess potent antidepressant effects.
Keywords: TrkB agonist, BDNF, synthetic derivatives, antidepressant, neurogenesis
Introduction
Neurotrophins are growth factors that regulate the development and maintenance of the peripheral and the central nervous system1. Brain-derived neurotrophic factor (BDNF) is a member of the neurotrophin family, which includes nerve growth factor (NGF), NT-3, NT-4 and NT-52. BDNF, like the other neurotrophins, exerts its biological functions on neurons through TrkB3. BDNF binding to TrkB triggers its dimerization through conformational changes and autophosphorylation of tyrosine residues in its intracellular domain, resulting in activation of the three major signaling pathways involving mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K) and phospholipase C-γ1. To pharmacologically mimic BDNF, we have developed a cell-based assay and identified a small molecule 7,8-dihydroxyflavone that binds the extracellular domain of TrkB, and triggers its dimerization and activation. Intraperitoneal administration of this compound elicits robust TrkB activation in mouse brain. Moreover, like BDNF, this compound displays potent neurotrophic effect in stroke and Parkinson disease animal models in a TrkB-dependent manner. Thus, 7,8-dihydroxyflavone is a novel TrkB agonist4.
Although BDNF/TrkB signaling is traditionally thought of in terms of synaptic plasticity and neuroprotection5, 6, recent evidence indicates that these molecules are also critical for neurogenesis and antidepressant drug efficacy. Antidepressant treatment induces BDNF mRNA expression, as well as autophosphorylation and activation of TrkB in the brain7, 8. The behavioral effects of antidepressants in the forced swim test are attenuated in mice with only one active BDNF allele (BDNF+/− mice), completely lacking BDNF specifically in forebrain or expressing a dominant negative form of TrkB (trkB.T1)8, 9. However, in this work, TrkB activation and the behavioral effects of these drugs are observed with acute antidepressant treatment, leaving the questions concerning chronic antidepressant efficacy unanswered. A breakthrough occurred recently linking antidepressants, TrkB, and neurogenesis with the publication of a landmark paper by Parada's group10. They showed that ablation of TrkB specifically in hippocampal neural progenitor cells prevents chronic antidepressant-induced neurogenesis and renders the mice behaviorally non-responsive to chronic antidepressant treatment. Combined, these results suggest a model whereby chronic antidepressant treatment induces BDNF expression and long-term activation of TrkB, leading to increased neurogenesis and an antidepressant effect11–13. As the result of a rash of recent data, a “neurotrophic” hypothesis of antidepressant activity has been suggested. Administration of chronic, but not acute, monoamine antidepressant drugs enhances adult neurogenesis in the subgranular zone of the dentate gyrus of rodents and non-human primates, while blocking neurogenesis by irradiation attenuates the behavioral antidepressant-like effects of these drugs in some rodent strains14–17. These results suggest that enhanced adult hippocampal neurogenesis is required for some aspects of antidepressant drug efficacy, and may help to explain why chronic treatment is required for therapeutic benefit.
In this study, we report that a hydrogen bond acceptor on the 4'-position of the flavone B ring is critical for 7,8-dihydroxyflavone's TrkB agonistic effect. The synthetic 4'-dimethylamino-7,8-dihydroxyflavone possesses a more potent agonistic effect on TrkB than the lead. Moreover, these two compounds are orally bio-available. They both strongly provoke neurogenesis and display robust antidepressant effects in a TrkB-dependent manner.
Results
Structure-activity relationship study
7,8-dihydroxyflavone (7,8-DHF) is a small molecular TrkB agonist. Our preliminary SAR supports that the 7,8-catechol moiety is essential for the agonistic effect by 7,8-DHF4. To explore the SAR in depth, we examined the TrkB stimulatory activity by numerous flavonoid derivatives. The numeric positions and each ring's nomenclature are designed (Figure 1A). The compounds were dissolved in dimethyl sulfoxide (DMSO), then diluted into 500 μM with 1× PBS (final vehicle contains 10% DMSO/phosphate buffered saline (PBS)) (Figure 1B). The primary rat cortical neurons (13 day in vitro (DIV)) were treated with 500 nM compounds for 20 min. The cell lysates were analyzed by immunoblotting. The positive control BDNF (100 ng/ml) strongly activated TrkB, as TrkB was robustly phosphorylated. As expected, 7,8,2'-trihydroxyflavone, 5,7,8-trihydroxyflavone, 3,7,8,2'-tetrahydroxyflavone, 7,8-DHF and 3,7-dihydroxyflavone all stimulated TrkB activation as compared to control (Figure 1C, top panel lane 3 and 7–10). Interestingly, 7,3'-dihydroxyflavone and 7,8,3'-trihydroxyflavone displayed an even more robust stimulatory effect on TrkB phosphorylation than the lead 7,8-DHF (Figure 1C, top panel lane 4–5), indicating that the 3'-hydroxy group in B ring can effectively escalate 7,8-DHF's agonistic activity. By contrast, 7,8,4'-trihydroxyflavone, 7-hydroxy-4'methoxyflavone and 8-hydroxy-7-methoxyflavone barely activated TrkB as compared to vehicle control (Figure 1C, top panel lane 6, 11 and 12). Strikingly, the highly hydroxylated flavone derivative, 3,5,7,8,3',4'-hexahydroxylatedflavone completely blocked TrkB phosphorylation (Figure 1C, top panel, last lane), suggesting the hyper-hydroxylated compound even antagonizes the TrkB activity. Immunoblotting analysis demonstrated Akt phosphorylation tightly coupled to TrkB activation (Figure 1C, bottom panel). Since no quantitative approach for mouse or rat TrkB phosphorylation is currently available, we monitored Akt activity by enzyme-linked immunosorbent assay (ELISA) in the primary neurons instead. In alignment with the immunoblotting results, Akt phosphorylation by these compounds exhibited the similar activation patterns. 7,8,3'-trihydroxyflavone displayed a more than 2-fold higher activity than vehicle control, and the 4'-hydroxylated derivatives failed to activate Akt (Figure 1D). Therefore, the hydroxy groups on the B ring can regulate 7,8-DHF's stimulatory activity on TrkB receptor. The 2'-hydroxy and, especially, the 3'-hydroxy groups elevate the agonistic effect, whereas the 4'-hydroxy group diminishes its stimulatory effect.
Figure 1. Structure-activity relationship study.

(A) 7,8-dihydroxyflavone chemical structure with every position in each ring labeled. (B) Chemical Structures of flavonoids from Indofine, Inc. (C) Immunoblotting analysis with neuronal lysates. Primary rat cortical neurons from E17 embryos (13 DIV) were treated with 500 nM various chemicals for 15 min. The neuronal cell lysates were collected and resolved on 10% SDS-PAGE. Immunoblotting was conducted with various antibodies. p-TrkB Y817 antibody was employed at 1:20,000–40,000 dilution. (D). Phospho-Akt 473 ELISA. The cell lysates (20 μg/sample) from the neurons treated with various indicated drugs were analyzed by p-Akt ELISA. The quantitative p-Akt activity in ELISA correlated with TrkB activity. Results were expressed as mean ± SEM (*P<0.05, compared with the vehicle control group, Student t-test, n=3).
To further explore the structure-activity relationships, we examined the role of different hydroxy groups in each ring (Supplemental Figure 1A &B). Akt ELISA analysis showed that an 8-hydroxy group on the A ring was essential for the stimulatory effect, as 5,7-DHF, 5,6-DHF and 5,6,7-THF each lost the effect as compared to 7,8-DHF. These compounds even decreased Akt activity as compared to the DMSO control. The dihydroxy groups on the B ring displayed relatively higher activity when compared to compounds with dihydroxy groups on the A ring. However, they were not significantly higher than the DMSO control. None of the single hydroxy flavone derivatives exhibited a notable stimulatory effect. Moreover, none of the tested trihydroxy flavones or dimethoxy flavones demonstrated any substantial effect on Akt activation. Similarly, neither coumarin derivatives nor 4-Cl-7-hydroxy-8-methoxy-isoflavone had any effect as compared to the DMSO control. It is noteworthy that 6,7-DHF and 6,7,3'-THF exhibited significant activity, suggesting that 6,7-dihydroxy groups retain partial activity (Supplemental Figure 1C). Together, the SAR study suggests that the catechol group (7,8-dihydroxy on the A ring) might be indispensable for the agonistic activity. A 4'-hydroxy group on the B ring reduces the activity, whereas a 3'-hydroxy group increases the activity.
Chemical synthesis of 7,8-dihydroxyflavone derivatives
The above SAR suggests that the 7,8-dihydroxy groups on the A ring should be kept intact. Hence we chose to leave the A ring intact and focused on modifying the B and C rings. To optimize the lead compound and conduct a more thorough SAR study, we synthesized a few more derivatives of 7,8-DHF. The synthetic pathways are outlined in Figure 2. In Figure 2A, 2-hydroxy-3,4-dimethoxyacetophenone (1) was first treated with pyridine and various benzoyl chlorides under refluxing conditions, followed by an acid-induced dehydrative cyclization to generate the desired unprotected flavones. To synthesize the NH-replaced flavones in the middle C ring, a mixture of ethyl 3-(4-fluorophenyl)-3-oxopropanoate and 2,3-dimethoxybenzenamine were refluxed in the presence of AcOH(cat.) and CaSO4 in EtOH (100 mL) at 75oC under N2, followed by cyclization and deprotection to generate the desired products in good yields (Figure 2B). The yield for each step in the synthesis was indicated.
Figure 2. Organic synthesis of various 7,8-dihydroxyflavone derivatives.

(A) & (B) Schematic diagram of synthetic pathways for various 7,8-DHF derivatives.
4'-Dimethylamino-7,8-dihydroxyflavone displays more potent stimulatory effect on TrkB receptor than the lead
To compare the TrkB activation by these synthetic compounds, we prepared primary cortical cultures and treated them with 500 nM of various compounds for 15 min and collected the cell lysates. Phospho-Akt ELISA analysis revealed that 4'-dimethylamino-7,8-dihydroxyflavone (compound #6, 4'-DMA-7,8-DHF) and 7,8-dihydroxy-2(pyrimidin-5-yl)-4H-chromen-4-one (compound #8) strongly activated Akt, followed by 4'-fluoro-7,8-DHF (compound #7). However, 2-(4-fluoro-phenyl)-7,8-dihydroxyquinolin-4(1H)-one (compound #13) and 7,8-dihydroxy-2-phenylquinolin-4(1H)-one (compound #14) failed to provoke Akt activation, indicating that the oxygen in the middle C ring is essential for 7,8-DHF's agonistic effect to TrkB (Figure 3A & B). These data suggest that replacing the H-bond accepting oxygen with H-bond donating NH abolishes its stimulatory effect. To quantitatively compare the activity between 4'-DMA-7,8-DHF and 7,8-DHF, we conducted a titration assay. Both 7,8-DHF and 4'-DMA-7,8-DHF triggered Akt activation at concentrations as low as 10 nM, and Akt activity gradually increased as drug concentration escalated. However, 4'-DMA-7,8-DHF displayed more robust activity than 7,8-DHF at 10 and 50 nM. At concentration of 100 nM or higher concentration, p-Akt ELISA by 4'-DMA-7,8-DHF was slightly decreased and displayed comparable activity as that of 7,8-DHF. This effect might be caused by other unknown pathways activated by 4'-DMA-7.8-DHF that subsides Akt activation at higher concentration (Figure 3B, right panels). The immunoblotting of TrkB phosphorylation correlated with the observed Akt activation pattern (Supplemental Figure 2). To further study 4'-DMA-7,8-DHF's kinetics on TrkB activation, we treated the primary neurons for various time points. 4'-DMA-7,8-DHF swiftly activated TrkB as soon as 5 min, and the signal slightly reduced at 10 min, and decayed back to baseline at 30 ~ 180 min (Figure 3C, top left panel). Once again, Akt activation pattern appears temporally coupled to TrkB activation (Figure 3C, 3rd left panel). However, MAPK phosphorylation peaked at 10 min (Figure 3C, 4th left panel). To compare the stimulatory effect on TrkB receptor in mouse brain, we orally administrated 1 mg/kg of these compounds into C57BL/6J mice and monitored TrkB activation at different time points. Clearly, both compounds elicited TrkB activation (p-TrkB 706) at 1 h and the activity of TrkB gradually escalated with time. 7,8-DHF-triggered TrkB activation climaxed at about 1~2 h, whereas 4'-DMA-7,8-DHF peaked at 4 h and partially decayed at 8~16 h. The downstream effectors of Akt and MAPK were potently activated by both compounds. As expected, both p-Akt and p-MAPK activated by 4'-DMA-7,8-DHF were stronger than 7,8-DHF, and the signals lasted longer for 4'-DMA-7,8-DHF than 7,8-DHF (Figure 3C, middle panels). Quantitative analysis with p-Akt 473 ELISA also supported that 4'-DMA-7,8-DHF was more potent than 7,8-DHF in triggering Akt activation in mouse brain (Figure 3C, right panel). Hence, 4'-DMA-7,8-DHF possesses a higher agonistic effect on TrkB than the parental compound 7,8-DHF and its stimulatory effect sustained longer in animals as well.
Figure 3. 4'-dimethylamino-7,8-dihydroxyflavone displays more potent TrkB stimulatory effect than parental 7,8-dihydroxyflavone.

(A) Chemical structures of various 7,8-DHF derivatives. (B) Phospho-Akt ELISA assay by the synthetic compounds in cortical neurons. Primary cortical cultures from E17 rat embryos were treated with 500 nM of various 7,8-DHF derivatives. The cell lysates were analyzed by the ELISA (left panel) (*: P<0.05, ***: P<0.001 vs vehicle, Student's t-test). Different doses of 4'-DMA-7,8-DHF and 7,8-DHF were incubated with primary cortical neurons for 15 min. The cell lysates (20 μg) were analyzed with p-Akt ELISA (right panel) (*: P<0.05, **: P<0.01, ***: P<0.001 vs control, one-way ANOVA; b: P<0.01, c: P<0.001 vs 7,8-DHF at same concentration, Student's t-test). The data were from 2 sets of replicated experiments (mean ± SEM). (C) Time course assay with 4'-DMA-7,8-DHF. Rat primary neurons were treated with 500 nM 4'-DMA-7,8-DHF fro various time points. The neuronal lysates were analyzed with various antibodies. 4'-DMA-7,8-DHF rapidly activated TrkB and its downstream signaling cascades (left panels). 4'-DMA-7,8-DHF revealed longer period of TrkB activation in mouse brain. 1 mg/kg of 4'-DMA-7,8-DHF and 7,8-DHF were orally administrated into C57 BL/6J mice and TrkB phosphorylation and its downstream signaling cascades including Akt and MAPK in mouse brain were analyzed by immunoblotting at various time points. TrkB activation by 4'-DMA-7,8-DHF peaked at 4 h, whereas the maximal TrkB activation by 7,8-DHF in mouse brain occurred at 1–2 h (middle panels). P-Akt 4734 ELISA in drug treated mouse brain was analyzed (right panel) (***: P<0.001 vs control, one-way ANOVA; a: P<0.05, c: P<0.001 vs 7,8-DHF at same concentration, Student's t-test). The data were from 2 sets of replicated experiments (mean ± SEM). (D) 7,8-dihydroxy groups are essential for the flavone's agonistic effect. Different methoxy replaced derivatives were tested on primary neurons by immunoblotting assays.
To further examine the role of each hydroxy group in 4'-DMA-7,8-DHF's agonistic activity, we prepared the 7-methoxy and 7,8-dimethoxy derivatives. Immunoblotting analysis demonstrated that replacement of the 7-hydroxy with a 7-methoxy group slightly reduced its agonistic effect. Switching both hydroxy groups to dimethoxy groups substantially decreased the agonistic effect (Figure 3D). Thus, the 7,8-dihydroxy groups are essential for the flavonoid's agonistic activity.
4'-Dimethylamino-7,8-dihydroxyflavone possesses more robust anti-apoptotic activity than the lead compound
To quantitatively compare the anti-apoptotic activity of these two TrkB agonists, we prepared cortical neurons, and pretreated the cells with various indicated concentrations of 4'-DMA-7,8-DHF and 7,8-DHF for 30 min, followed by 50 μM glutamate for 16 h. The cell lysates were quantitatively analyzed with an active caspase-3 ELISA. Glutamate-provoked caspase-3 activation was substantially blocked by both compounds at 50 nM or higher concentrations. However, at 10 nM, 4'-DMA-7,8-DHF displayed a more robust inhibitory effect than 7,8-DHF (Figure 4A & B). These results fit with the TrkB receptor activation status by 4'-DMA-7,8-DHF and 7,8-DHF (Figure 3B). To investigate whether these compounds exert any neuroprotective effects in animals, we conducted a time course assay. A titration assay revealed that 7,8-DHF induced TrkB activation in a dose-dependent manner upon oral administration (Supplemental Figure 3). Our previous study suggested that 7,8-DHF displayed a robust neuroprotective effect at a dose of 5 mg /kg4. Hence, we administrated both compounds (5 mg/kg) orally by gavage. At either 0 h, 2 h or 6 h, we intraperitoneally administrated kainic acid (KA)(20 mg/kg). After 2 h, we sacrificed the mice and prepared the hippocampal regions. Immunoblotting with mouse brain lysates demonstrated that KA-induced caspase-3 activation was slightly reduced with time lapse, which inversely correlated with TrkB activation by 4'-DMA-7,8-DHF (Figure 4C, left panels). Nonetheless, KA-induced caspase-3 activation was reduced by 7,8-DHF at 4 h, and active caspase-3 was slightly increased at 8 h. This kinetic spectrum tightly coupled to the TrkB activation status by 7,8-DHF (Figure 4C, right panel).
Figure 4. 4'-dimethylamino-7,8-dihydroxyflavone prevents neurons from apoptosis in a TrkB-dependent manner.

(A & B) Active caspase-3 ELISA assay. Cortical neurons were prepared from E16 rat embryonic. The neurons were pretreated with different doses of compounds as indicated for 30 min, followed by 50 μM glutamate for 16 h. The cell lysates were analyzed by active caspse-3 ELISA. (C) 4'-DMA-7,8-DHF and 7,8-DHF prevent KA-elicited neuronal cell death. C57BL/6J mice were orally administrated with 5 mg/kg of 4'-DMA-7,8-DHF and 7,8-DHF, at different time points, the mice were i.p. injected with 20 mg/kg KA for 2 h. The brain lysates were analyzed by immunoblotting with anti-p-TrkB, anti-active caspase-3 antibodies, respectively. (D) TrkB activation is indispensable for the neuroprotective effect of 4'-DMA-7,8-DHF. 4'-DMA-7,8-DHF and 7,8-DHF suppressed KA-induced caspase-3 activation in TrkB F616A mutant knockin mice, which can not be blocked by 1NMPP1 (top panel). TrkB F616A was strongly activated by 4'-DMA-7,8-DHF and 7,8-DHF, which was blocked by 1NMPP1 (middle panel).
To explore whether the neuroprotective actions of these small molecules is dependent on TrkB activation in vivo, we employed TrkB F616A knock-in mice, where it has been shown that TrkB F616A can be selectively blocked by 1-(1,1-dimethylethyl)-3-(1-naphthalenylmethyl)-1H-pyrazolo[3,4-d]pyrimidin-4-amine (1NMPP1), a TrkB F616A inhibitor, resulting in effective TrkB-null phenotypes18. Since 1NMPP1 selectively inhibits TrkB F616A activation by 7,8-DHF, we reasoned that blockade of TrkB F616A signaling by 1NMPP1 in mice would make the neurons vulnerable to KA-provoked neuronal cell death. As expected, KA caused significant caspase-3 activation, which was reduced by 4'-DMA-7,8-DHF and 7,8-DHF pretreatment. Interestingly, 1NMPP1 pretreatment abolished 4'-DMA-7,8-DHF and 7,8-DHF's protective effect in F616A mice (Figure 4D, top panel). Accordingly, TrkB phosphorylation by 4'-DMA-7,8-DHF and 7,8-DHF was notably blocked by 1NMPP1 pretreatment (Figure 4D, 2nd panel). Hence, these data demonstrate that 4'-DMA-7,8-DHF and 7,8-DHF selectively activate the TrkB receptor and enhance neuronal survival in mice.
4'-Dimethylamino-7,8-dihydroxyflavone and 7,8-dihydroxyflavone promote neurogenesis
Administration of chronic, but not acute, monoamine antidepressant drugs enhances adult neurogenesis in dentate gyrus of rodents and non-human primates, while blocking neurogenesis by irradiation attenuates the behavioral antidepressant-like effects of these drugs in some rodent strains14–17. This suggests that enhanced adult hippocampal neurogenesis is required for the efficacy of antidepressant drugs. Ablation of TrkB specifically in hippocampal neural progenitor cells prevents chronic antidepressant-induced neurogenesis and renders the mice behaviorally non-responsive to chronic antidepressant treatment10. To test whether increasing TrkB activation by these small agonists would elevate the neurogenesis, we administrated either vehicle, 7,8-DHF and 4'-DMA-7,8-DHF (5 mg/kg), orally by gavage to adult male C57BL/6J mice for 21 days. At the end of treatment (day 21), the animals were injected with BrdU (50 mg/kg, intraperitoneal (i.p.)) to label the dividing cells and were sacrificed 2 h later. BrdU immunohistochemistry was then used to assess progenitor proliferation (Figure 5A). We observed that long-term (21 days) TrkB agonists treatment significantly increased neurogenesis as compared to the vehicle control. Immunohistochemistry staining demonstrated that TrkB was markedly activated by these two chemicals in dentate gyrus (Figure 5B) after 3 weeks treatment. Therefore, chronic treatment with TrkB agonists promotes neurogenesis in the hippocampus of mice.
Figure 5. 4'-dimethylamino-7,8-dihydroxyflavone and 7,8-dihydroxyflavone promote neurogenesis.

A, Neurogenesis assay. Male C57BL/6J mice were orally administrated with 5 mg/kg 4'-DMA-7,8-DHF and 7,8-DHF and vehicle solvent for 21 days, and followed by 50 mg/kg BrdU i.p. injection. In 2 h, the mice were perfused and brain sections were immunostained with anti-BrdU and DAPI. The positive cells in dentate gyrus were highlighted by arrow (left panels). Quantitative analysis of the BrdU positive cells in dentate gyrus (right panel). B, 7,8-DHF and its derivative upregulate TrkB activation in dentate gyrus. Paraffin section were deparaffinized in xylene and rehydrated gradient ethanol solution. Samples were boiled in 10 mM sodium citrate buffer for 20 min for antigen retrieval purpose. Brain sections were incubated with anti-TrkB (BD biosciences, San Jose, CA) 1:50, and anti-p-TrkB Y816 was used at 1:300 dilution. Secondary antibody were applied using anti-rabbit-Alexa 594 (red), anti-mouse-FITC (green). DAPI (blue) was used for nuclear staining.
4'-dimethylamino-7,8-dihydroxyflavone and 7,8-dihydroxyflavone demonstrate antidepressant effect in a TrkB-dependent manner
Accumulating evidence supports that BDNF plays an essential role in mediating antidepressants' therapeutic effects8, 9, 19, 20. Infusion of exogenous BDNF into hippocampus or brain stem has anti-depressant-like behavioral effect21, 22. A forced swim test is broadly used for screening of potential antidepressant drugs and is widely used to measure antidepressant activity23, 24. To explore whether 4'-DMA-7,8-DHF and 7,8-DHF have any antidepressant effect like BDNF, we conducted a forced swim test after chronic treatment of the mice for 21 days via oral administration. When mice were treated with 7,8-DHF (5 mg/kg), the swimming immobility was significantly decreased. Interestingly, 4'-DMA-7,8-DHF (5 mg/kg) also evidently reduced the immobility (Figure 6A), suggesting that 7,8-DHF and its derivative imitate BDNF and exert potent anti-depressant effects. Immunoblotting analysis revealed that both compounds evidently provoked TrkB but not TrkA activation in mouse brain (Figure 6B).
Figure 6. 4'-dimethylamino-7,8-dihydroxyflavone and 7,8-dihydroxyflavone demonstrate antidepressant effect in a TrkB-dependent manner.

(A) Forced swim test with 4'-DMA-7,8-DHF and 7,8-DHF compounds. Male C57BL/6J mice (8 mice/group) were orally administrated by gavage with 5 mg/kg 4'-DMA-7,8-DHF and 7,8-DHF and vehicle solvent saline for 21 days, and subjected to a forced swim test (6 min, immobility recorded in the last 4 min). Data are presented as mean ± SEM. Analysis of variance (ANOVA) revealed significant difference between vehicle and either 7,8-DHF or 4'-DMA-7,8-DHF (n=6, ***P<0.0001 vs vehicle). (B) TrkB but not TrkA is activated by 4'-DMA-7,8-DHF and 7,8-DHF in mouse brain. The brain lysates from above chronically treated mice were analyzed by immunoblotting with anti-p-TrkA 794 and p-TrkB 817. (C) Forced swim test with TrkB F616A knockin mice. Male TrkB knockin mice were given the regular drinking water or 1NMPP1 (25 μM) containing drinking water one day before we started to inject the drugs and sustained throughout the whole experiment. The indicated control (saline) and drugs were administrated for 5 days. Data are presented as mean ± SEM; Analysis of variance (ANOVA) revealed significant effect between vehicle and either 7,8-DHF (n=7 mice, **P<0.001) or 4'-DMA-7,8-DHF (**P<0.001) in TrkB KI mice. None of the drugs produced a significant change in 1NMPP1 treated TrkB KI mice (n=6 to 7 mice) as compared to control.
To assess whether the behavior responses by 7,8-DHF and its derivative are mediated by the TrkB receptor, we utilized TrkB F616A knockin mice. The transgenic mice were subjected to saline or 1NMPP1 pretreatment, respectively. No significant difference was observed in the immobility time between saline and 1NMPP1 treated control groups. In saline group, both 7,8-DHF and 4'-DMA-7,8-DHF substantially reduced the immobility time; in contrast, neither 4'-DMA-7,8-DHF nor 7,8-DHF had any significant effect on the immobility time after 1NMPP1 treatment (Figure 6C), suggesting that inhibition of TrkB signaling cascade blocks the antidepressant effect by the TrkB agonists. Thus, these data demonstrate that 4'-DMA-7,8-DHF and its parental lead mimic BDNF and act as potent antidepressant drugs in mice through activating TrkB receptor.
Discussion
Flavonoids are a large group of polyphenolic compounds containing a basic flavan nucleus with two aromatic rings (the A and the B rings) interconnected by a three-carbon-atom heterocyclic ring (the C ring). Our previous structure-activity relationship study and the current work both support that 7,8-dihydroxy groups are critical for the lead compound's agonistic effect. Chrysin and 5,7-dihydroxyflavone have no effect at all, supporting that the ortho-dihydroxy groups have to be integral4. Hence, when we synthesized new derivatives, we kept the ortho-hydroxy groups on the A ring intact. In this report, we show that the oxygen atom in the C ring is also essential for the stimulatory activity. Replacement of this O atom with NH group abolishes the agonistic activity (Figure 3). However, the B ring might tolerate some modification. We have demonstrated that B-ring-modified compounds bearing either a 4'-dimethylamino, 2'- or 3'-hydroxy-group (and, specifically, not 4'-hydroxy) display robust TrkB stimulatory effects. Interestingly, 4'-DMA-7,8-DHF exhibits a higher agonistic activity and prolonged effect in mouse brain than the lead. Fitting with its robust agonistic effect to TrkB receptor, this derivative reveals potent neuroprotective activities. Notably, these agonists exert a prominent neurogenesis effect and significant antidepressant activity, in alignment with the physiological actions of BDNF/TrkB signalings.
Flavonoids represent one of the largest and the most diverse class of plant secondary metabolites. These compounds are naturally present in vegetables, fruits, and beverages and thus are important components of the daily diet. Flavonoids have been known for a long time to exert diverse biological effects (bioflavonoids) and in particular to act as antioxidants and preventive agents against cancer25. Accumulating evidence suggests that flavonoids have the potential to improve human memory and neuro-cognitive performance via their ability to protect vulnerable neurons, enhance existing neuronal function and stimulate neuronal regeneration26. Flavonoids exert effects on LTP, one of the major mechanisms underlying learning and memory, and consequently memory and cognitive performance, through their interactions with the signaling pathways including PI3K /Akt27, MAPK28, 29 protein kinase C30 etc. The pleiotropic activity profile often results in a complicated structure-activity relationship particularly in cell-based and in vivo systems where metabolism adds further complexity in data interpretation. Our data support that 4'-DMA-7,8-DHF possesses more robust neuroprotective action than the lead compound. Oral administration of these two compounds into mice elicits robust TrkB activation in mouse brain, suggesting that these compounds are orally bio-available. In addition, 4'-DMA-7,8-DHF displays a longer TrkB stimulatory activity in mouse brain (Figure 3). Presumably, this modified derivative or its metabolites have a longer half-life in animals. However, it has to be pointed out that we did not determine drug levels either systemically or centrally and it is unclear if the differential effects are simply a reflection of differential in vivo exposures of the analogs. To explore whether these compounds have any intolerable toxicity, we have analyzed all of the major organs from the mice, which were treated with both compounds at 5 mg/kg for 3 weeks. No appreciable adverse pathological change was detectable in the drug-treated mice (Supplemental Figure 4). In addition, a complete blood count (CBC) analysis shows that there is no significant difference between drug-treated and the control saline-treated mice (Supplemental Table 1). Hence, these data support that both compounds are not toxic to the mice at 5 mg/kg dose over the 3-week chronic treatment. The titration experiment also supports that neither compound is toxic for HEK293 cell proliferation at a dose up to 50 μM. Nonetheless, at 100 μM 7,8-DHF displays demonstrable anti-proliferative effect, whereas 4'-DMA-7,8-DHF has no effect (Supplemental Figure 5A & B). Neither of the compounds provokes cell death in HEK293 cells (Supplemental Figure 5C & D). Neuronal cell death assay reveals that both compounds are non-toxic for primary cortical neurons up to 5 μM (Supplemental Figure 5E & F).
Emerging evidence supports that BDNF and TrkB receptor are implicated in both the development of mood disorders and the action of antidepressants31–33. BDNF-mediated TrkB signaling is both sufficient and necessary for antidepressant-like behaviors21, 34. The antidepressant effect by imipramine or fluoxetine are abolished in transgenic mice with reduced BDNF or TrkB signaling9, 35. All these findings indicate that TrkB signaling is crucial in the antidepressant effect. Here, we provide further evidence supporting that TrkB signaling is critical for this process. We show that both 4'-DMA-7,8-DHF and 7,8-DHF, which mimic BDNF and potently activate the TrkB receptor, exhibit evident antidepressant effects in both wild-type mice and TrkB F616A knockin mice. However, the antidepressant activity by 7,8-DHF and 4'-DMA-7,8-DHF are blunted in TrkB F616A knockin mice, when the TrkB receptor is inhibited by 1NMPP1 (Figure 6). These data demonstrate that these compounds render the antidepressant effect through activating the TrkB receptor signaling cascade. In wild-type mice, both compounds reveal comparable antidepressant effects at 5 mg/kg. At a dose of 1 mg/kg, 4,-DMA-7,8-DHF displays a more potent effect than 7,8-DHF (Figure 3C). Conceivably, we might be able to distinguish these two compounds' antidepressant effects at this low dose. Interestingly, 4'-DMA-7,8-DHF exhibits a much more potent antidepressant effect than the parental compound 7,8-DHF in TrkB F616A knockin mice, and the anti-depressant effect is completely inhibited by 1NMPP1, supporting that this antidepressant effect is exerted through TrkB receptor. Collectively, these findings demonstrate that 4'-DMA-7,8-DHF possesses the stronger agonistic effect on TrkB than the lead.
Accumulating evidence supports that hippocampal neurogenesis is required for the efficacy of antidepressant drugs36–38. Ablation of TrkB specifically in hippocampal neural progenitor cells prevents chronic antidepressant-induced neurogenesis and renders the mice behaviorally non-responsive to chronic antidepressant treatment10. Our neurogenesis assay on the animal with chronic drug treatment supports that both 4'-DMA-7,8-DHF and the parental lead evidently provoke neurogenesis in the denta gyrus of drug-treated mice (Figure 5). This finding fits with the previous report that exogenous BDNF promotes proliferation of hippocampal NPCs39. The robust antidepressant effects of these two TrkB agonists support an important role for BDNF in mediating the biological response to chronic AD treatments40. Together, our data support that 4'-DMA-7,8-DHF is a more potent synthetic TrkB agonist than 7,8-DHF. The SAR study points a clear direction for the future optimization of a 7,8-DHF TrkB agonist.
Experimental Section
Cells, reagents and mice
NGF and BDNF were from Roche. Anti-p-TrkB 817 was from Epitomics. Anti-TrkB antibody was from Biovision. Anti-TrkA was from Cell Signaling. TrkBF616A mice have been described previously18. TrkBF616A mice and wild-type C57BL/6 mice were bred in a pathogen-free environment in accordance with Emory Medical School guidelines. All chemicals not included above were purchased from Sigma. 7,8-dihydroxyflavone was purchased from TCI. The flavanoids were from Indofine (Hillsborough, NJ 08833, USA). NMR spectrum (Bruker AV300K, 300MHz), MS spectrum (Shimadzu LCMS), HPLC (PE, dual pumper, SPD detector, ODS -C18 reverse phase, 254 nm, CH3CN-H2O-0.1%TFA). Phospho-TrkB Y816 antibody was raised against [H]-CKLQNLAKASPV-pY-LDILG-[OH] (a.a. 806 – 822)(EM437 and EM438) as rabbit polyclonal antibody in Covance. The antiserum was purified by affinity columns. This phospho-TrkB was utilized for immunostaining the brain sections. Anti-TrkB (Cell Signaling, which recognizes both full-length and truncated TrkB) and anti-TrkB (Biovision, which only recognizes full-length TrkB) were used for immunoblotting. Mouse Monoclonal anti-TrkB (BD Bioscience) was used for immunostaining. P-Akt 473 Sandwich ELISA was from Cell Signaling. BDNF was from Roche. Anti-phospho-TrkA 785, anti-TrkA, Phospho-Akt-473, anti-Akt and Anti-phospho-Erk1/2 antibodies were from Cell Signaling. Anti-p-TrkB Y817 antibody were from Epitomics.
Synthesis of 7,8-dihydroxyflavone derivatives
1H and 13C NMR spectra were recorded with a Varian 300 spectrometer. Chemical shifts are reported as δ values (parts per million, ppm). Infrared spectra were recorded on a Bruker Equinox 55 FTIR spectrophotometer. Peaks are reported in cm−1. Melting points were obtained with a Barnstead Electrothermal MelTemp apparatus. ESI MS spectra were recorded on an Applied Biosystems 4000 QTrap spectrometer. Gallacetophenone, Gallacetophenone 3',4'-dimethyl ether (1) and all acid chlorides were purchased from Sigma Aldrich (acid chlorides were also prepared directly from the corresponding carboxylic acids). All other reagents and solvents were purchased from commercial sources and purified by standard procedures. All reactions were performed under a dry N2 atmosphere in flame-dried glassware. The purity of the compounds (> 95%) is confirmed by HPLC.
General procedures for the synthesis of 7,8-DHF derivatives according to Figure 2
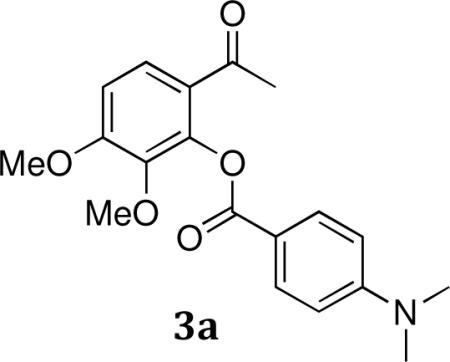
General synthesis of aryl benzoates (3a–c): 6-Acetyl-2,3-dimethoxyphenyl 4-(dimethylamino)benzoate (3a)
To a solution of gallacetophenone 3',4'-dimethyl ether (5.83 g, 29.72 mmol) in dry pyridine (25 mL) was added (4-dimethylamino)benzoyl chloride (8.19 g, 44.58 mmol) in three portions over 15 min. The mixture was stirred at room temperature for 2 h. The reaction was acidified with 2M HCl and extracted with ethyl acetate. The combined organics were washed with brine, dried with MgSO4, and concentrated under reduced pressure. The crude product is purified by flash chromatography (10% EtOAc/hexanes) to afford 8.45 g (83%) of 3 as a white crystalline solid.
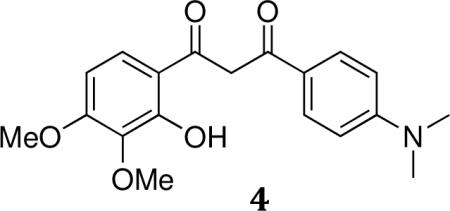
General synthesis of 1,3-diketones (4a–c): 1-(4-(Dimethylamino)phenyl)-3-(2-hydroxy-3,4-dimethoxyphenyl)propane-1,3-dione (4a)
A solution containing 6-acetyl-2,3-dimethoxyphenyl 4-(dimethylamino)benzoate 3 (8.45 g, 24.6 mmol), anhydrous powdered potassium hydroxide (2.08 g, 36.9 mmol), and pyridine (50 mL) was heated at 50°C for 2 hour. Reaction was cooled to room temperature, acidified with 2M HCl, extracted with ethyl acetate, washed with brine, dried with MgSO4, and evaporated under reduced pressure to yield 7.59 g (90%) of crude propanedione 4. The crude reaction mixture was carried forward without further purification.
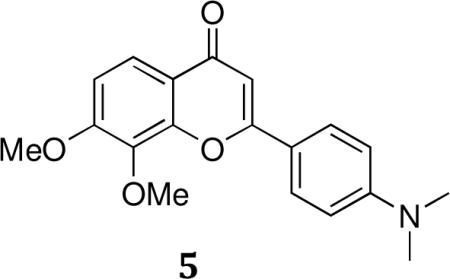
General synthesis of dimethoxy chromen-4-ones (5a–c): 2-(4-(Dimethylamino)phenyl)-7,8-dimethoxy-4H-chromen-4-one (5a)
A solution of 1-(4-(dimethylamino)phenyl)-3-(2- hydroxy-3,4-dimethoxyphenyl)propane-1,3-dione (4) (11.5 g, 33.4 mmol) in glacial acetic acid (100 mL) and concentrated sulfuric acid (1 mL) was refluxed for 1 hour. The reaction mixture was then poured into ice and extracted with ethyl acetate. The combined organics were washed with brine, dried with MgSO4 and concentrated under reduced pressure to yield a dark solid. Flash column chromatography (30% EtOAc/hexane) yielded 6.41 g (60%) as a yellow-brown solid.
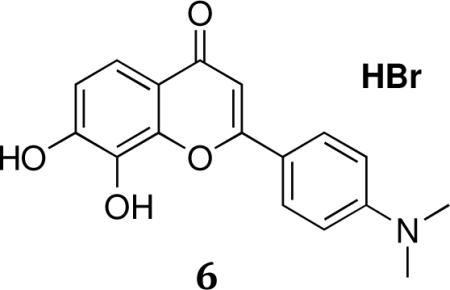
2-(4-(Dimethylamino)phenyl)-7,8-dihydroxy-4H-chromen-4-one.HBr (4'-DMA-7,8-DHF.HBr, 6)
A solution of 2-(4-(dimethylamino)phenyl)-7,8-dimethoxy-4H-chromen-4-one (v) (0.462 g, 1.42 mmol) in aqueous hydrobromic acid (48%, 10 mL) is refluxed overnight. After cooling, the reaction mixture is diluted with water, neutralized with saturated NaHCO3, and extracted with 1-butanol. The organic phase is washed with water, dried with MgSO4, and evaporated under reduced pressure. Recrytallization from 50% methanol/dichloromethane provided 0.221 g (52%) of 6.
1H NMR (300 MHz, DMSO-d6) 7.96(d,J=8.80,2H), 7.35 (d, J = 8.56, 1H),6.91 (d, J = 8.64, 1H), 6.85 (d, J = 8.76,2H), 6.64(s,1H),3.02 (s, 6H);MS-ESI: cal. 297; found: 298(M+H)+. Anal. for C17H15NO4. 1.06HBr; calcd: C, 53.30; H, 4.23; N, 3.66; Br, 22.11;Found: C, 53.08; H, 4.23; N, 3.42; Br, 21.81; m.p.:267.8–270.4 °C; HPLC: 100%.
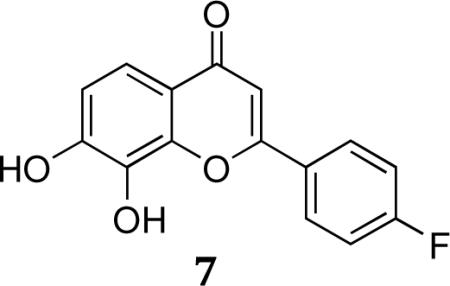
2-(4-Fluorophenyl)-7,8-dihydroxy-4H-chromen-4-one.HBr (4'-F-7,8-DHF.HBr, 7)
1H NMR (300 MHz, DMSO-d6) • 8.20–8.25 (dd, J = 8.80, 2H), 7.35 (d,J = 8.56, 1H),6.91 (d, J = 8.64,1H), 6.85 (d, J=8.76,2H), 6.64(s,1H),3.02 (s, 6H);MS-ESI: cal. 272.0478; found: 272.0485.
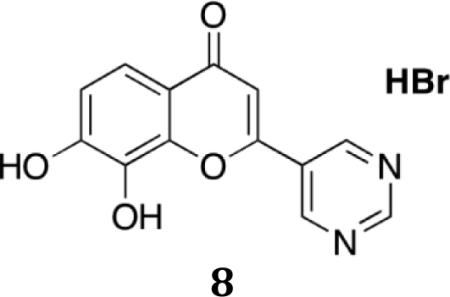
7,8-Dihydroxy-2-(pyrimidin-5-yl)-4H-chromen-4-one.HBr (8)
1H NMR (300 MHz, CDCl3) • 10.27 (brs, 1H), 9.65(brs, 1H), 9.52 (s, 2H), 9.32 (s, 1H), 7.39 (d, J =8.7, 1H), 7.10 (s, 1H), 6.94(d,J=8.7,1H); cal. 256; found: 257(M+H)+. Anal. for C13H8N2O4. 0.45H2O. 0.3HBr; calcd: C, 54.10; H, 3.21; N, 9.71; Br, 8.31; Found: C: 54.30; H, 3.39; N, 9.41; Br, 8.37. m.p.>300°C. HPLC: 89.3%.
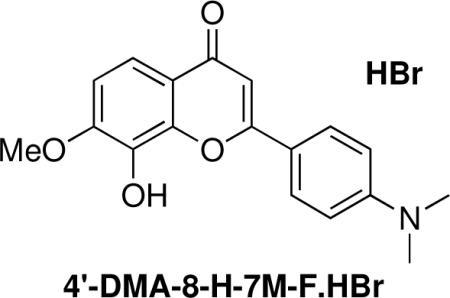
2-(4-(Dimethylamino)phenyl)-7-hydroxy-8-methoxy-4H-chromen-4-one. HBr (4'-DMA-8-H-7M-F.HBr)
1H (CDCl3) δ 7.95 (d, J = 9 Hz, 1H), 7.75 (d, J = 9 Hz, 2H), 7.08 (d, J = 9 Hz, 1H), 6.78 (d, J = 9 Hz, 2H), 6.65 (s, 1H), 3.99 (s, 3H), 3.08 (s, 6H); 13C (CDCl3) δ 177.1, 163.2, 153.0, 128.1, 121.0, 111.9, 109.8, 103.6, 61.7, 57.2.
Representative procedures for the synthesis of 7,8-dihydroxy quinolin-4(1H)-one derivatives according to Figure 2B
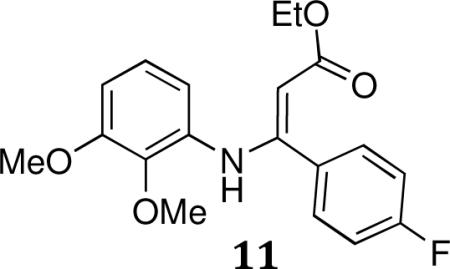
(E)-ethyl 3-(2,3-dimethoxyphenylamino)-3-(4-fluorophenyl)acrylate
A mixture of ethyl 3-(4-fluorophenyl)-3-oxopropanoate (5.00 g, 23.8 mmol), 2,3-dimethoxybenzenamine (3.64g, 23.8 mmol), AcOH(cat.) and CaSO4(6.5g, 47.6 mmol) in EtOH (100mL) was refluxed at 75°C under N2 for 144h. The reaction mixture was concentrated to afford crude product which was purified by flash chromatography to afford (E)-ethyl 3-(2,3-dimethoxyphenylamino)-3-(4-fluorophenyl)acrylate (11).
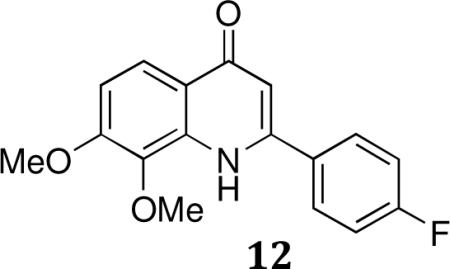
2-(4-fluorophenyl)-7,8-dimethoxyquinolin-4(1H)-one (12)
A solution of (E)-ethyl 3-(2,3-dimethoxyphenylamino)-3-(4-fluorophenyl)acrylate(11) in Ph2O was heated to 250°C for 10min. concentrated. The residue was purified by flash chromatography (CH2Cl2 –MeOH, 40:1) to afford 2-(4-fluorophenyl)-7,8-dimethoxyquinolin-4(1H)-one(12).
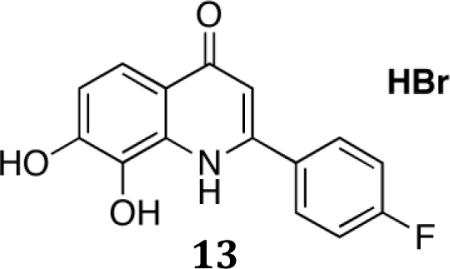
2-(4-Fluorophenyl)-7,8-dihydroxyquinolin-4(1H)-one.HBr (13)
To a solution of 2-(4-fluorophenyl)-7,8-dimethoxyquinolin-4(1H)-one (500mg) in CH2Cl2 was added slowly BBr3 at 0°C under N2, then the reaction was stirred at rt for 3h. The reaction was cooled down to 0°C and quenched by adding slowly MeOH. Concentrated and the residue was purified by recrystallization from methyl tert-Butyl ether to afford 2-(4-fluorophenyl)-7,8-dihydroxyquinolin-4(1H)-one(13) as a red solid (200mg). 1H NMR (300 MHz, DMSO-d6) • 10.83(br s,1H),10.09(br s,1H),7.92(q, J1=5.1,J2=8.1 2H), 7.70 (d,J = 8.7, 1H),7.46 (t, J=8.7,2H), 7.30 (d, J=9,1H), 6.87(s,1H),MS-ESI: cal. 271; found: 272(M+H)+. Anal. For C15H10FNO3. 0.87HBr.2.4H2O; calcd: C, 46.81; H, 4.10; N, 3.64; Br, 18.06; Found: C, 46.75; H, 3.80; N, 3.26; Br, 17.46; m.p.>300. HPLC: 98.9%;
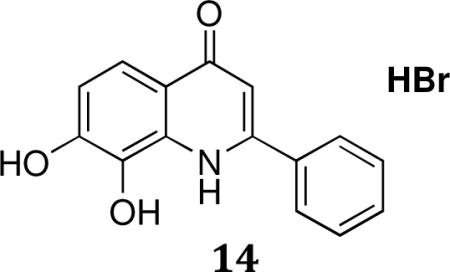
7,8-dihydroxy-2-phenylquinolin-4(1H)-one.HBr (14)
1H NMR (300 MHz, DMSO-d6) • 10.71(br s,1H),7.81(d,J=7.2,2H), 7.65 (d,J = 9.3, 1H),7.57–7.66 (m, 3H), 7.23 (d, J=9.3,1H), 6.81(s,1H),MS-ESI(negative): cal. 253; found: 252(M-1). Anal. For C15H11NO3. 0.8HBr.0.96H2O; calcd: C, 53.74; H, 4.12; N, 4.18; Br, 18.90; Found: C, 53.45; H, 3.96; N, 4.43; Br, 19.11; m.p.:269.8–274.2°C. HPLC: 100%;
Phospho-AktS473 ELISA
The ELISA was carried out using 96-well Nunc-Immuno MaxSorp plates (VWR Cat.# 62409-024). PathScan Phospho-Akt1 (Ser473) Sandwich ELISA kit was purchased from Cell Signaling (Cat.# 7160). The cell lysates from primary neurons or mouse brain that were treated with various compounds were employed in the assay. 100 μl sample (20–40 g) in sample diluent buffer (supplied in the kit) was added to each well and incubated overnight at 4 °C. After 4× wash with 200 μl wash buffer (supplied in the kit), 100 μl/well detection antibody was added and incubated for 1 h at 37 °C. After 4× wash, 100 μl of HRP-linked secondary antibody (supplied in the kit) was added and incubated for 30 min at 37 °C. After a final wash, 100 μl of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate was added to each well and incubated for 10 minutes at 37 °C. The reaction was stopped by adding 100 μl/well stop solution. The values of each well were recorded using the microplate reader at 450 nm and 650 nm. The optical density was determined by the subtraction of the reading at 650 nm from the readings at 450 nm.
Kainic Acid/TrkB agonists drug Administration
Male C57BL/6 mice aged of 60 days were administrated orally with a single dose of 4'-DMA-7,8-DHF or 7,8-DHF (1 mg/kg each). KA (20 mg/kg) (Sigma, MO) was i.p. injected. Animals were continually monitored for 2 h for the onset of seizure activity. At 0, 4 and 8 h following TrkB agonist treatment, the animals were sacrificed and the hippocampal section lysates were analyzed by immunoblotting with p-TrkB, active caspase-3 and total TrkB antibodies.
TrkB agonists suppress KA-induced neuronal cell death in TrkB F616A mice
TrkB F616A knockin mice (2–3 months old) were fed with 1NMPP1 (25 μM) in drinking water one day before pharmacological reagent treatment. Next day, the mice were administrated orally by gavage with 7,8-DHF or 4'-DMA-7,8-DHF (5 mg/kg) 4 h before kainic acid (20 mg/kg). The control mice were injected with saline, 1NMPP1, kainic acid alone or administrated 7,8-DHF or 4'-DMA-7,8-DHF 4 h before kainic acid. In 4 days, the mice were sacrificed and brains were homogenated and ultracentrafuged. The supernatant (40 μg) was employed for SDS-PAGE and immunoblotting analysis with indicated antibodies, respectively.
Immunohistochemistry staining
Brain tissues were fixed in 4% paraformaldehyde overnight followed by paraffin embedding. Sections of 6 μm were cut. For immunohistochemical staining, brain sections were deparaffinized in xylene and rehydrated in graded alcohols. Endogenous peroxidase activity was blocked by 3% hydrogen peroxide for 5 minutes and all slides were boiled in 10 mM sodium citrate buffer (pH 6.0) for 10 minutes. Phosphorylated Trk B816 and Trk B were detected using specific antibodies. Paraffin section were deparaffinized in xylene and rehydrated gradient ethanol solution. Samples were boiled in 10mM sodium citrate buffer for 20 min for antigen retrieval purpose. Brain sections were incubated with anti-TrkB (BD biosciences, San Jose, CA) 1:50, p-TrkB 1:300 dilution. Secondary antibody was applied using anti-rabbit-Alexa 594 (red), anti-mouse- Fluorescein isothiocyanate (FITC) (green). DAPI (blue) was used for nuclear staining.
Force swim test
Adult male mice (2–3 months old) were randomly submitted to a forced swim test without a pre-swim. Saline, 4'-DMA-7,8-DHF and 7,8-DHF (5 mg/kg) were orally administrated by gavage for 21 days. The mice were allowed to adapt to the test room for 2 days. The mice were placed in a clear glass cylinder with a diameter of 16 cm, half-filled with clear water at 24 °C (water depth of 14 cm did not allow the mice to reach the bottom of the cylinder; water was changed after each mouse) for a total of 6 min, and immobility was recorded during the last 4 min by an investigator blind to the genotype and treatment.
Neurogenesis analysis in TrkB agonists treated hippocampus
Adult male mice (2–3 months old) were orally administrated with Saline, 4'-DMA-7,8-DHF and 7,8-DHF (5 mg/kg) for 21 days. Then BrdU (50 mg/kg) was i.p. injected. In 2 h, the mice were perfused with 4% paraformaldehyde. Immunohistochemical staining was performed on formalin-fixed paraffin-embedded sections. Sections from brain were cut, deparaffinized in xylene and rehydrated in graded alcohols. The slides were boiled in 10 mM citric acid (pH 6.0) for 10 min followed by an incubation in 2 N HCl for 10 min in room temperature. The slides were then permeabilized and blocked with 1 % BSA in 0.2% PBS tween-20 (PBST). The incorporated BrdU were stained using anti-BrdU-FITC (Abcam, USA) at 4 °C for 16 hr. After three washing in PBS, the cells were then stained with DAPI for another 10 min at room temperature. The slides were finally mounted with AquaMount (Lerner Laboratories, USA) containing 0.01% 1,4-diazobicyclo(2,2,2)octane and examined under a fluorescence microscope.
Supplementary Material
Acknowledgement
This work is supported by grants from National Institute of Health RO1 NS045627 to K. Ye. The Authors are thankful to Dr. David Ginty at Johns Hopkins University for TrkB F616A knockin mice.
Abbreviations
- 1NMPP1
1-(1,1-dimethylethyl)-3-(1-naphthalenylmethyl)-1H-pyrazolo[3,4-d]pyrimidin-4-amine
- 4'-DMA-7,8-DHF
4'-dimethylamino-7,8-dihydroxyflavone
- 7,8-DHF
7,8-dihydroxyflavone
- BDNF
brain-derived neurotrophic factor
- CBC
complete blood count
- DIV
day in vitro
- DMSO
dimethyl sulfoxide
- ELISA
enzyme-linked immunosorbent assay
- FITC
Fluorescein isothiocyanate
- i.p.
intraperitoneal
- KA
kainic acid
- LDH
lactate dehydrogenase
- MAPK
mitogen-activated protein kinase
- MTT
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- NGF
nerve growth factor
- PBS
phosphate buffered saline
- PBST
phosphate buffered saline Tween-20
- ppm
parts per million
- TMB
3,3′,5,5′-tetramethylbenzidine
- TrkB
tropomyosin-receptor-kinase B
Footnotes
Supporting Information: Supporting information (Supplemental table 1 and Fig S1–S5) is available free of charge via the Internet at http://pubs.acs.org.
References
- 1.Lewin GR, Barde YA. Physiology of the neurotrophins. Annu Rev Neurosci. 1996;19:289–317. doi: 10.1146/annurev.ne.19.030196.001445. [DOI] [PubMed] [Google Scholar]
- 2.Thoenen H. The changing scene of neurotrophic factors. Trends Neurosci. 1991;14:165–170. doi: 10.1016/0166-2236(91)90097-e. [DOI] [PubMed] [Google Scholar]
- 3.Kaplan DR, Miller FD. Neurotrophin signal transduction in the nervous system. Curr Opin Neurobiol. 2000;10:381–391. doi: 10.1016/s0959-4388(00)00092-1. [DOI] [PubMed] [Google Scholar]
- 4.Jang SW, Liu X, Yepes M, Shepherd KR, Miller GW, Liu Y, Wilson WD, Xiao G, Blanchi B, Sun YE, Ye K. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc Natl Acad Sci U S A. 2010;107:2687–2692. doi: 10.1073/pnas.0913572107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nagappan G, Lu B. Activity-dependent modulation of the BDNF receptor TrkB: mechanisms and implications. Trends Neurosci. 2005;28:464–471. doi: 10.1016/j.tins.2005.07.003. [DOI] [PubMed] [Google Scholar]
- 6.Hennigan A, O'Callaghan RM, Kelly AM. Neurotrophins and their receptors: roles in plasticity, neurodegeneration and neuroprotection. Biochem Soc Trans. 2007;35:424–427. doi: 10.1042/BST0350424. [DOI] [PubMed] [Google Scholar]
- 7.Nibuya M, Morinobu S, Duman RS. Regulation of BDNF and trkB mRNA in rat brain by chronic electroconvulsive seizure and antidepressant drug treatments. J Neurosci. 1995;15:7539–7547. doi: 10.1523/JNEUROSCI.15-11-07539.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Saarelainen T, Hendolin P, Lucas G, Koponen E, Sairanen M, MacDonald E, Agerman K, Haapasalo A, Nawa H, Aloyz R, Ernfors P, Castren E. Activation of the TrkB neurotrophin receptor is induced by antidepressant drugs and is required for antidepressant-induced behavioral effects. J Neurosci. 2003;23:349–357. doi: 10.1523/JNEUROSCI.23-01-00349.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Monteggia LM, Barrot M, Powell CM, Berton O, Galanis V, Gemelli T, Meuth S, Nagy A, Greene RW, Nestler EJ. Essential role of brain-derived neurotrophic factor in adult hippocampal function. Proc Natl Acad Sci U S A. 2004;101:10827–10832. doi: 10.1073/pnas.0402141101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Li Y, Luikart BW, Birnbaum S, Chen J, Kwon CH, Kernie SG, Bassel-Duby R, Parada LF. TrkB regulates hippocampal neurogenesis and governs sensitivity to antidepressive treatment. Neuron. 2008;59:399–412. doi: 10.1016/j.neuron.2008.06.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sahay A, Hen R. Adult hippocampal neurogenesis in depression. Nat Neurosci. 2007;10:1110–1115. doi: 10.1038/nn1969. [DOI] [PubMed] [Google Scholar]
- 12.Banasr M, Duman RS. Regulation of neurogenesis and gliogenesis by stress and antidepressant treatment. CNS Neurol Disord Drug Targets. 2007;6:311–320. doi: 10.2174/187152707783220929. [DOI] [PubMed] [Google Scholar]
- 13.Schmidt HD, Duman RS. The role of neurotrophic factors in adult hippocampal neurogenesis, antidepressant treatments and animal models of depressive-like behavior. Behav Pharmacol. 2007;18:391–418. doi: 10.1097/FBP.0b013e3282ee2aa8. [DOI] [PubMed] [Google Scholar]
- 14.Malberg JE, Eisch AJ, Nestler EJ, Duman RS. Chronic antidepressant treatment increases neurogenesis in adult rat hippocampus. J Neurosci. 2000;20:9104–9110. doi: 10.1523/JNEUROSCI.20-24-09104.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Santarelli L, Gobbi G, Debs PC, Sibille ET, Blier P, Hen R, Heath MJ. Genetic and pharmacological disruption of neurokinin 1 receptor function decreases anxiety-related behaviors and increases serotonergic function. Proc Natl Acad Sci U S A. 2001;98:1912–1917. doi: 10.1073/pnas.041596398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Surget A, Saxe M, Leman S, Ibarguen-Vargas Y, Chalon S, Griebel G, Hen R, Belzung C. Drug-dependent requirement of hippocampal neurogenesis in a model of depression and of antidepressant reversal. Biol Psychiatry. 2008;64:293–301. doi: 10.1016/j.biopsych.2008.02.022. [DOI] [PubMed] [Google Scholar]
- 17.Perera TD, Coplan JD, Lisanby SH, Lipira CM, Arif M, Carpio C, Spitzer G, Santarelli L, Scharf B, Hen R, Rosoklija G, Sackeim HA, Dwork AJ. Antidepressant-induced neurogenesis in the hippocampus of adult nonhuman primates. J Neurosci. 2007;27:4894–4901. doi: 10.1523/JNEUROSCI.0237-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chen X, Ye H, Kuruvilla R, Ramanan N, Scangos KW, Zhang C, Johnson NM, England PM, Shokat KM, Ginty DD. A chemical-genetic approach to studying neurotrophin signaling. Neuron. 2005;46:13–21. doi: 10.1016/j.neuron.2005.03.009. [DOI] [PubMed] [Google Scholar]
- 19.Castren E. Neurotrophic effects of antidepressant drugs. Curr Opin Pharmacol. 2004;4:58–64. doi: 10.1016/j.coph.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 20.Groves JO. Is it time to reassess the BDNF hypothesis of depression? Mol Psychiatry. 2007;12:1079–1088. doi: 10.1038/sj.mp.4002075. [DOI] [PubMed] [Google Scholar]
- 21.Siuciak JA, Lewis DR, Wiegand SJ, Lindsay RM. Antidepressant-like effect of brain-derived neurotrophic factor (BDNF) Pharmacol Biochem Behav. 1997;56:131–137. doi: 10.1016/S0091-3057(96)00169-4. [DOI] [PubMed] [Google Scholar]
- 22.Shirayama Y, Chen AC, Nakagawa S, Russell DS, Duman RS. Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of depression. J Neurosci. 2002;22:3251–3261. doi: 10.1523/JNEUROSCI.22-08-03251.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Porsolt RD, Le Pichon M, Jalfre M. Depression: a new animal model sensitive to antidepressant treatments. Nature. 1977;266:730–732. doi: 10.1038/266730a0. [DOI] [PubMed] [Google Scholar]
- 24.Cryan JF, Markou A, Lucki I. Assessing antidepressant activity in rodents: recent developments and future needs. Trends Pharmacol Sci. 2002;23:238–245. doi: 10.1016/s0165-6147(02)02017-5. [DOI] [PubMed] [Google Scholar]
- 25.Harborne JB, Williams CA. Advances in flavonoid research since 1992. Phytochemistry. 2000;55:481–504. doi: 10.1016/s0031-9422(00)00235-1. [DOI] [PubMed] [Google Scholar]
- 26.Spencer JP. Food for thought: the role of dietary flavonoids in enhancing human memory, learning and neuro-cognitive performance. Proc Nutr Soc. 2008;67:238–252. doi: 10.1017/S0029665108007088. [DOI] [PubMed] [Google Scholar]
- 27.Vauzour D, Vafeiadou K, Rice-Evans C, Williams RJ, Spencer JP. Activation of pro-survival Akt and ERK1/2 signalling pathways underlie the anti-apoptotic effects of flavanones in cortical neurons. J Neurochem. 2007;103:1355–1367. doi: 10.1111/j.1471-4159.2007.04841.x. [DOI] [PubMed] [Google Scholar]
- 28.Schroeter H, Bahia P, Spencer JP, Sheppard O, Rattray M, Cadenas E, Rice-Evans C, Williams RJ. (−)Epicatechin stimulates ERK-dependent cyclic AMP response element activity and up-regulates GluR2 in cortical neurons. J Neurochem. 2007;101:1596–1606. doi: 10.1111/j.1471-4159.2006.04434.x. [DOI] [PubMed] [Google Scholar]
- 29.Maher P, Akaishi T, Abe K. Flavonoid fisetin promotes ERK-dependent long-term potentiation and enhances memory. Proc Natl Acad Sci U S A. 2006;103:16568–16573. doi: 10.1073/pnas.0607822103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Levites Y, Amit T, Youdim MB, Mandel S. Involvement of protein kinase C activation and cell survival/ cell cycle genes in green tea polyphenol (−)-epigallocatechin 3-gallate neuroprotective action. J Biol Chem. 2002;277:30574–30580. doi: 10.1074/jbc.M202832200. [DOI] [PubMed] [Google Scholar]
- 31.Altar CA. Neurotrophins and depression. Trends Pharmacol Sci. 1999;20:59–61. doi: 10.1016/s0165-6147(99)01309-7. [DOI] [PubMed] [Google Scholar]
- 32.Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM. Neurobiology of depression. Neuron. 2002;34:13–25. doi: 10.1016/s0896-6273(02)00653-0. [DOI] [PubMed] [Google Scholar]
- 33.Castren E, Voikar V, Rantamaki T. Role of neurotrophic factors in depression. Curr Opin Pharmacol. 2007;7:18–21. doi: 10.1016/j.coph.2006.08.009. [DOI] [PubMed] [Google Scholar]
- 34.Koponen E, Rantamaki T, Voikar V, Saarelainen T, MacDonald E, Castren E. Enhanced BDNF signaling is associated with an antidepressant-like behavioral response and changes in brain monoamines. Cell Mol Neurobiol. 2005;25:973–980. doi: 10.1007/s10571-005-8468-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rantamaki T, Hendolin P, Kankaanpaa A, Mijatovic J, Piepponen P, Domenici E, Chao MV, Mannisto PT, Castren E. Pharmacologically diverse antidepressants rapidly activate brain-derived neurotrophic factor receptor TrkB and induce phospholipase-Cgamma signaling pathways in mouse brain. Neuropsychopharmacology. 2007;32:2152–2162. doi: 10.1038/sj.npp.1301345. [DOI] [PubMed] [Google Scholar]
- 36.Santarelli L, Saxe M, Gross C, Surget A, Battaglia F, Dulawa S, Weisstaub N, Lee J, Duman R, Arancio O, Belzung C, Hen R. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science. 2003;301:805–809. doi: 10.1126/science.1083328. [DOI] [PubMed] [Google Scholar]
- 37.Encinas JM, Vaahtokari A, Enikolopov G. Fluoxetine targets early progenitor cells in the adult brain. Proc Natl Acad Sci U S A. 2006;103:8233–8238. doi: 10.1073/pnas.0601992103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wang JW, David DJ, Monckton JE, Battaglia F, Hen R. Chronic fluoxetine stimulates maturation and synaptic plasticity of adult-born hippocampal granule cells. J Neurosci. 2008;28:1374–1384. doi: 10.1523/JNEUROSCI.3632-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Scharfman H, Goodman J, Macleod A, Phani S, Antonelli C, Croll S. Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp Neurol. 2005;192:348–356. doi: 10.1016/j.expneurol.2004.11.016. [DOI] [PubMed] [Google Scholar]
- 40.Wang JW, Dranovsky A, Hen R. The when and where of BDNF and the antidepressant response. Biol Psychiatry. 2008;63:640–641. doi: 10.1016/j.biopsych.2008.01.008. [DOI] [PubMed] [Google Scholar]
- 41.Menichincheri M, Ballinari D, Bargiotti A, Bonomini L, Ceccarelli W, D'Alessio R, Fretta A, Moll J, Polucci P, Soncini C, Tibolla M, Trosset JY, Vanotti E. Catecholic flavonoids acting as telomerase inhibitors. J Med Chem. 2004;47:6466–6475. doi: 10.1021/jm040810b. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.