

Abstract
Genetic manipulation is an important facet of synthetic biology but can be complicated by undesired nuclease degradation. Incorporating non-natural nucleic acids into a gene could convey resistance to nucleases and promote expression. The compatibility of non-natural nucleosides with polymerases is reviewed with a focus on results from the past two years. Details are provided about how the different systems could be useful in synthetic biology.
Synthetic biology aims to fabricate living systems and at the same time achieve a better understanding of life in general [1-3]. A number of recent reviews develop the finer points of synthetic biology [4-6], which include bioengineering, biotechnology, molecular evolution, and systems biology [7]. The deletion and insertion of genes is often an important component of synthetic biology, and the use of non-natural nucleic acids could facilitate genetic manipulation. Over the past few decades, a number of chemistry-based research groups have clearly demonstrated that DNA and RNA are not the only molecular systems capable of storing genetic information [8-10]. At the same time, studies on polymerase replication and transcription of oligonucleotide sequences reveal that chemical entities other than the typical nucleoside triphosphates can be tolerated to varying degrees [11]. Therefore, a non-natural genetic system could be considered as the carrier of genetic information in a synthetic cell. It is intriguing to consider the possibility that a cell could be designed to rely on a genome in which DNA and RNA are replaced by alternative chemical entities.
Most synthetic biology borrows and/or hijacks the existing components of cells while reprogramming the hosts genome, but this process can be complicated by degradation of the non-natural gene. Some of the recent problems in accomplishing gene transfer between species of bacteria and yeast were due to restriction endonuclease activity that needed to be suppressed before success was realized [12]. A possible advantage to using non-natural alternatives to DNA and RNA in synthetic biology would be added stability to endogenous proteins that are eager to degrade natural nucleic acids. Therefore, a gene partially composed of non-natural nucleic acids could have a higher chance of being expressed due to its resistance to degradation. The challenge of incorporating non-natural nucleosides has typically stemmed from their incompatibility with biological machinery that replicates nucleic acids, namely DNA and RNA polymerases. Recent advances, however, point to the possibility of overcoming such impediments.
For application in synthetic biology a non-natural nucleic acid must be recognized and properly used by polymerases while not being degraded by nucleases. The majority of nucleic acid mimics developed over the past few decades were made for applications in the field of medicine and drug discovery, and such molecules were designed to inhibit cellular machinery associated with progression of a disease [13]. For synthetic biology, however, only a handful of nucleic acid mimics are tolerated in gene replication and transcription in bacteria. This review specifically focuses on the examples of DNA and RNA mimics that satisfy one or more of the criteria below:
There is some type of base pairing between the non-natural nucleic acid and a complementary oligonucleotide sequence.
There is a mechanism to incorporate the non-natural nucleic acid across from a template.
The information copied from the synthetic gene can be converted into a RNA message.
Before describing the specific examples of the different analogs, there are a few caveats to consider. Most of the examples below satisfy criteria 1; in some cases Watson-Crick hydrogen bonding between canonical base pairs is maintained but in other examples there are non-natural base pairs that rely on alternative hydrogen bonding arrangements or on hydrophobic interactions. Criteria 2 would be important in engineering genes that need to be copied as cells divide. This criterion is achieved with varying degrees of efficiency, some very good, some mediocre, and others are too preliminary to thoroughly evaluate. There are only a few cases where a chemically different form of messenger RNA is translated by ribosomes (which satisfies criteria 3). Additional complications of selecting the proper cell membrane and delivering the non-natural nucleosides across a cell membrane are not evaluated in this review, but recent work of Szostak and Luisi addresses some of these concerns in the context of synthetic biology [14,15]. This review focuses primarily on incorporation of analogs into DNA and RNA; the stability of non-natural oligonucleotides to nuclease degradation can be found in recent reviews covering the areas of antisense and siRNA research [16-18]. Finally, this article should be regarded as a partial survey of the chemical systems that could satisfy some of the requirements important to synthetic biology.
Phosphate Analogs
Synthetic changes to the phosphodiester backbone of DNA and RNA are good initial options to explore for applications in synthetic biology [16]. Numerous alternatives to this chemical linkage have been investigated with the explicit intention of stabilizing the modified oligonucleotide to nuclease degradation [19]. In some cases, the effects of such changes on polymerase activity have also been studied. The most common phosphodiester analog is the phosphorothioate in which one phosphate oxygen is replaced with a sulfur atom (Figure 1A). These derivatives have been extensively examined for antisense activity [13]. A number of DNA polymerases accept a triphosphate version of this derivative in which the sulfur is present at the α-position of the triphosphate [11]. Phosphorothioate linkages are chiral, with a stereogenic center present at the phosphorous atom. Mechanistic studies have demonstrated that several polymerases selectively use one stereoisomer of a sulfur-modified nucleoside triphosphate (the Sp configuration) and that polymerization proceeds with inversion of configuration at phosphorous [20]. Recent examination of this mechanism by Benner indicates that the stereochemical requirements may not be so rigid for all polymerases, especially when a sulfur-modified nucleoside triphosphate contains a non-natural nucleobase (which in Benners work was the nucleobase P, Figure 2C) [21]. Ribosomes also accept mRNA messages with the phosphorothioate modification [22]. Recent reports have even revealed that certain bacteria naturally incorporate phosphorothioates into their genomes [23], and the genes encoding this process have been determined [24,25]. An alternative to phosphorothioates are the boranophosphates, a linkage that is similarly tolerated by polymerases and ribosomes [26]. The ability of polymerases to incorporate phosphonate nucleosides has been recently examined [27]. These derivatives contain an extra methylene group between the 5′ oxygen and the phosphorous atom and are highly resistant to nuclease degradation. A mutant DNA polymerase (Therminator) was able to incorporate 20 consecutive adenine-bearing phosphonate units across from a DNA duplex that had a thymine overhang of the same number. A subsequent study indicated that adenine- and cytosine-phosphonate derivatives are favored by polymerases for incorporation while the thymine and uracil versions are incorporated to a limited extent. In a competition study, the natural adenine nucleoside triphosphate is still preferred compared to the phosphonate version [28]. Continued studies with these analogues, along with efforts of polymerase engineering [29], could provide optimum conditions for extended polymerization of phosphonates.
Figure 1.
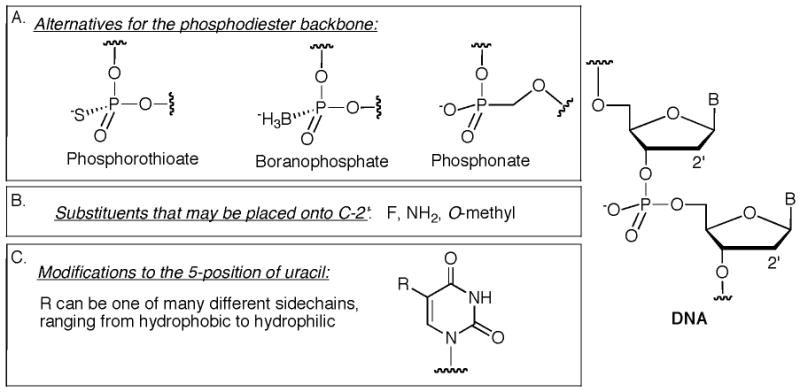
Figure 2.
Aptamers
The field of aptamers has also tackled the incorporation of synthetic nucleotides, focusing on both phosphodiester replacements as well as modified bases [30,31]. Aptamers are single-stranded oligonucleotides that are generated through an in vitro selection and evolution process called SELEX (which stands for Systematic Evolution of Ligands by Exponential enrichment). The evolution process for SELEX depends on employing several rounds of enzymatic amplification of oligonucleotide sequences that are selected to have good binding properties to a particular target. Since aptamers composed of DNA and RNA are themselves susceptible to nuclease degradation, the field has made significant efforts to find synthetic derivatives that are sufficiently stable for therapeutic applications. Therefore, techniques to perform SELEX with modified nucleosides has been developed, and also recently reviewed [30,31]. To briefly summarize, in vitro evolution using phosphorothioates has been successfully accomplished [32]. In addition, a number of mutant T7 RNA polymerases have been developed that are tolerant of substitutions at the 2′ position of the furanose ring (Figure 1B). Substitutions that may be attached to C2′ include: fluorine, an amine, and a methoxy group [33]. One of the most impressive accomplishments from aptamer development is the extensive number of modifications that are tolerated at the 5-position of uracil (Figure 1C) [34]. A wide variety of sidechains may be attached to this position, ranging from simple hydrophobic groups to extended sidechains terminated by polar groups. These modified nucleotides are tolerated by a variety of T7 RNA polymerases [35].
Nucleobase Analogs
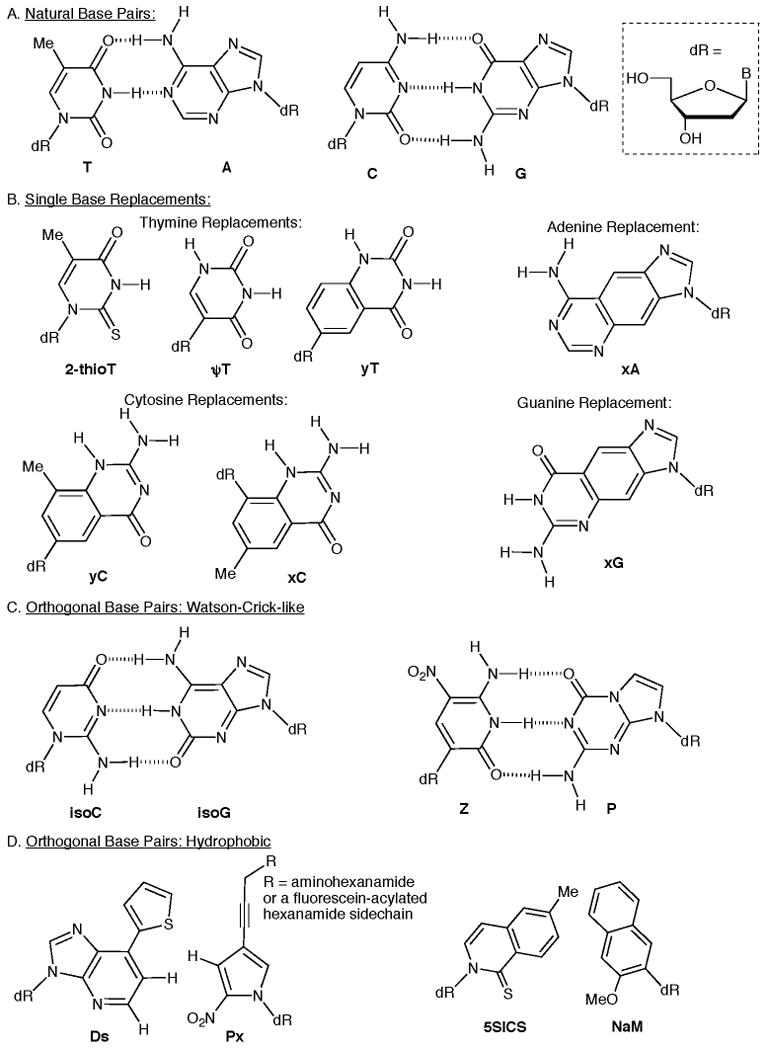
The development of non-natural base pairs for incorporation into DNA and RNA has been the focus of several research groups for a number of years [36]. There are two basic strategies in this area: the first aims to maintain a Watson-Crick-like network of hydrogen bonding between bases, while the second strategy uses hydrophobic groups that mimic the shape and polarity of the natural bases. In the first of these strategies, Benner is developing synthetic biology based on “artificially expanded genetic information systems (AEGIS) [37].” These systems contain six or more nucleotide base pairs, each one still conforming to the basic Watson-Crick geometry but differing in the arrangement of hydrogen-bond donors and acceptors (compare the natural T:A and C:G (Figure 2A) to the synthetic isoC:isoG and Z:P base pairs (Figure 2C)). Benner initially investigated the ability of an isoC:isoG base pair to be incorporated and replicated by DNA polymerases, but the simultaneous use of 2-thiothymine (2-thioT, Figure 2B) was essential to maintain high isoC:isoG selectivity [38]. Recently, Benner and his group have shown that one of these new base-pairs (Z:P) [39] can be readily incorporated by DNA polymerases with 97.5% retention of the non-natural base pair in a PCR amplification [40]. In addition, Benner has shown that an α-thiotriphosphate containing the nucleobase P can be used by polymerases to make a phosphorothioate-containing oligonucleotide [21]. Benner has also recently investigated the ability of pseudothymidine (ψT, Figure 2B) to be incorporated into DNA as a substitute for thymidine [41].
Kool has pioneered the development of expanded DNA bases to create xDNA and yDNA [36,42]. A recent paper by Kool has shown that two nucleotides (xA and xC, Figure 2B) of his extended nucleotide system can be replicated by polymerases in E. Coli to develop a normal message [43]. Similarly, two purines from yDNA (yT and yC) can be incorporated into E. Coli, but fidelity of replication is less efficient [44].
The hydrophobic approach to develop synthetic base pairs aims to mimic the size, shape, and polarity (in some cases) of a natural base pair, but without the hydrogen bonding component. Seminal work by Kool showed that the hydrophobic group difluorotoluene mimics thymine and is efficiently incorporated by polymerases on an oligonucleotide template [45]. Since this discovery, other hydrophobic base pairs have been developed. Hirao and coworkers recently reported a Ds:Px base pair (Figure 2D). This system allows multiple functional groups to be incorporated on the propargyl position of Px. Sequences with this pair can be amplified by 107 fold with 30 cycles of PCR and >99.9% fidelity per cycle [46]. Romesburg has developed his own system of hydrophobic base pairs and has recently shown that both replication and translation of the 5SICS:NaM pair (Figure 2D) proceeds efficiently [47]. Romesburgs bases also can be present in both the sense and antisense strand of DNA without loss in replication or translation efficiency [48].
Substitutions of the Furanose Ring
One of the most fundamental changes that could be made in a synthetic cell would be to alter the structure of the furanose ring that supports both the nucleotide base and the phosphodiester backbone. Three specific examples highlighted below are promising candidates for advancing into synthetic biology.
Threose Nucleic Acid (TNA)
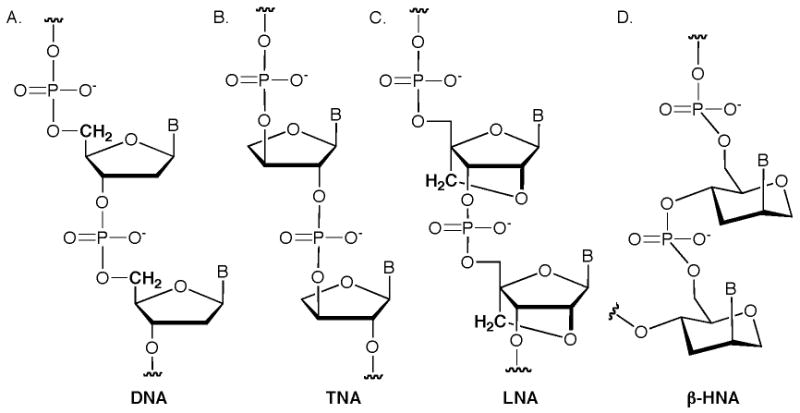
Among the numerous nucleic acid mimics studied by Eschenmoser, the version called TNA (an abbreviation for α-(L)-threofuranosyl-(3′-2′) nucleic acid, Figure 3B) could have the most immediate application to synthetic biology [49]. TNA differs from DNA in that the phosphates are linked to oxygens on the 3′ and 2′ positions of the furanose and there is no methylene group present between the sugar ring and an oxygen atom on the phosphate [50]. TNA forms hydrogen-bonded duplexes to complementary TNA sequences (called intrasystem cross-pairing) and similarly interacts with complementary sequences of DNA or RNA (intersystem cross-pairing). Structural details on such complexes reveal that a TNA-TNA duplex bears structural resemblance to A-form DNA or RNA with a slightly shorter distance between adjacent phosphorous atoms [51,52]. Perhaps the most intriguing aspect of TNA is the ability with which genetic information transfer can be catalyzed by polymerases. In particular, Szostak has demonstrated that Therminator DNA polymerase is adept at incorporation of TNA triphosphates using a DNA template [53]. Under the optimal conditions, it was estimated that TNAs of about 200 nucleotides could be generated with acceptable fidelity for in vitro selection experiments [54]. Another important facet of this study is the development of a TNA sequencing scheme to transfer the message from TNA back to DNA followed by DNA sequencing. A recent study of phosphonate analogs of TNA also indicate that Therminator polymerase has some potential to incorporate the adenine analogs [27].
Figure 3.
Locked Nucleic Acid (LNA)
A nucleic acid analog that has seen an enormous amount of development across numerous fields of nucleic acid-based applications is LNA (an abbreviation for Locked Nucleic Acid, Figure 3C) [55,56]. These molecules were developed by Wengel and first reported about 10 years ago. The LNA unit derives from RNA in that it has a 2′-oxygen, but the special feature of LNA is the extra methylene group that is covalently linked to C4′ and the oxygen of C2′ of the furanose ring. This linkage of LNA locks the sugar ring into an N-type or C3′-endo conformation that is favorable for binding natural nucleic acids, RNA in particular. LNAs engage in both intra- and intersystem cross-pairing, and LNA-LNA duplexes are extremely stable [57]. Structural studies have also shown that an LNA-RNA duplex has an A-form double helical structure while an LNA-DNA duplex resembles the structure seen in RNA-DNA duplexes [58,59]. Small numbers of LNA units have been used to replace portions of natural RNA units in aptamers or ribozymes to improve resistance to degradation by nucleases and even improve activity [60-62]. Investigations into the ability of polymerases to use LNA triphosphates began a couple of years ago and so far the results are preliminary but encouraging [63]. To date, incorporation of LNA units on a DNA or RNA template is limited to around 8 consecutive bases using KOD DNA polymerase or T7 RNA polymerase [64]. More work is needed to optimize such conditions, but it should be noted that there are a number of other LNA stereoisomers that show similar nucleic acid binding properties [65-70]. One of these other isomers could be more compatible with polymerases.
Hexitol Nucleic Acid (HNA)
The work of Herdewijn has extensively contributed to the general field of nucleic acid mimics, especially for medicinal applications and bioengineering [71-75]. Among the numerous nucleic acid analogues Herdewijn has developed, HNA (an abbreviation for hexitol nucleic acid) in which the nucleobase is present in the β position (β-HNA, Figure 3D) is a promising candidate for introduction into synthetic biology applications [76]. HNAs engage in both intra- and intersystem cross-pairing, and bind very tightly to RNA [77]. Similar to LNA, structures of HNA-HNA and HNA-RNA duplexes adopt an A-form helix [78]. There are a few polymerases that demonstrate good activity for incorporating HNA, namely the Vent(exo-) DNA polymerase, and the kinetics of incorporation are similar to the natural deoxynucleotide triphosphates [79]. However, the fidelity of HNA incorporation with Vent(exo-) can be maintained only if no more than two consecutive HNAs are inserted. Another study of HNA explored the ability of HIV Reverse Transcriptase (RT) to insert HNA, and a mutant form of HIV-RT (M184V) was adept at this process, specifically for the adenine derivative [80]. Taking this further, Herdewijn also demonstrated that a 33 nucleotide mRNA containing six consecutive HNAs (encompassing the AUG start codon plus the adjacent UUC codon) did not interfere with ribosome binding, tRNA binding, or translation of the message in an in vitro expression system [81]. More recently, a series of HNA:DNA duplexes (in which 1 to 6 consecutive HNAs were present in one strand of the duplex) were incorporated into a functional and selectable E. coli gene for thymidylate synthase [82]. Following ligation to a vector and transformation into E. coli, functional transformants were obtained, although the yield of such transformants decreased with increasing number of HNA residues (possibly due to the inefficient translation of the synthetic plasmid by bacterial polymerases). More recent results of Herdewijn demonstrate polymerase incorporation of other isomers related to HNA [83].
Acyclic Analogues
Acyclic nucleic acid analogs are conformationally more flexible compared to most of the other nucleic acid mimics. At the same time, the synthetic routes to make such analogs are typically much simpler compared to the methods to make the cyclic mimics of nucleic acids. Some of these flexible backbones still have the ability to cross-pair with nucleic acids as well as, in some cases, to be utilized by polymerases.
Flexible Nucleic Acid (FNA)
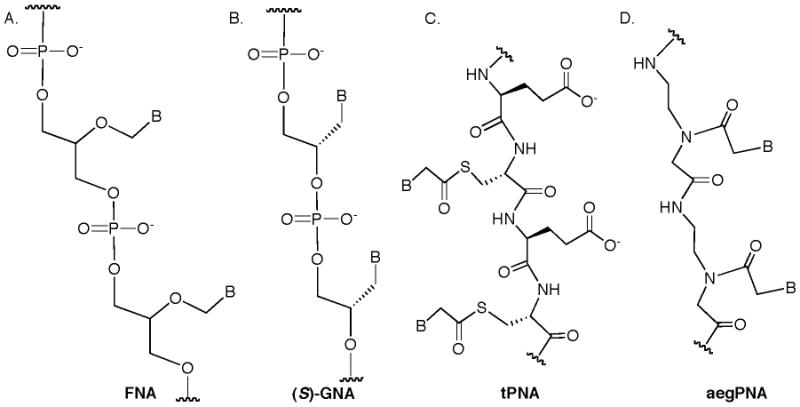
One of the earliest proposals for an evolutionary precursor of DNA and RNA is a nucleic acid based on formyl glycerol (FNA, Figure 4A) [84]. This system was initially described by Leslie Orgel (d. 2007); one of the first scientists to propose that an RNA-world preceded the molecular evolution of DNA and proteins [85,86]. While FNA shows no tendency to cross-pair with DNA, a recent investigation by Switzer has demonstrated that either FNA enantiomer (as well as a racemic mixture) can be used by DNA polymerases, with a preference for the R enantiomer [87]. With Therminator DNA polymerase, extensions of up to 7 nucleotides are possible with good fidelity.
Figure 4.
Glycerol Nucleic Acid (GNA)
Meggers has introduced an alternative acyclic nucleic acid mimic based on commercially available glycidol [88]. The resulting propylene glycol phosphosphate diester backbone (GNA, Figure 4B) is highly flexible, yet the oligomer with the S enantiomer cross-pairs well with RNA. GNA also cross-pairs with itself [89]. Recently, Szostak investigated the ability of GNA triphosphates to be used by DNA polymerases [90]. Therminator DNA polymerase successfully incorporates 2 GNA triphosphates on a DNA template, but efficiency decreases substantially with 5 GNAs and the pyrimidine GNA triphosphates are weak binders to the polymerase. An interesting solution to improve the activity of pyrimidine GNA triphosphates was to make thymine-substituted versions with a propyne group at the 5 position in an effort to improve base stacking with the terminal nucleotide of the primer. This strategy worked for cytosine but not for thymine. In a separate study by Szostak, polymerases were also able to use GNA as a template to make DNA [91]. Aminopropyl versions of GNA have also been reported recently, but compatibility with polymerases has not yet been investigated [92].
Peptide Nucleic Acid (PNA)
One of the most successful strategies to mimic nucleic acids has been to graft nucleobases onto a peptide backbone to make, “Peptide Nucleic Acids,” usually abbreviated PNA. A number of different PNAs have been explored over the past few decades, including aminoethylglycine PNA (aegPNA) [93] and very recently thioester PNA (tPNA) [94]. Ghadiri and Orgel have recently reported a very intriguing thioester PNA (tPNA, Figure 4C) in which the nucleobases self-assemble onto the peptide backbone in the presence of an oligonucleotide template [94]. The tPNA is constructed from a sequence of cysteine residues at every other position along a polypeptide backbone. Residues at the non-cysteine locations can be variable, but are typically amino acids with polar, ionic sidechains. The formation of the tPNA itself is the product of combining the cysteine-bearing polypeptide with individual nucleobases (derivatized with a thioester) and an oligonucleotide template. After incubation for a couple of hours, the tPNA enriched with the sequence of nucleobases complementary to the oligonucleotide template is obtained. This unique self-assembly process could potentially become the basis of a replication mechanism in a synthetic biology construct.
Nielsen and Buchardt developed what has become the most commonly used PNA [93]. The aminoethyl glycine version of PNA (sometimes called aegPNA, Figure 4D) displays both intra- and intersystem cross-pairing with remarkably high thermal stability and sequence specificity [95]. As a result, aegPNA-based applications have been developed in areas of biomedical research [96-105]. The simplicity of the aegPNA backbone has also contributed to proposals that such a molecule could have been a prebiotic precursor to nucleic acids [106,107]. While polymerase-based synthesis of aegPNA has not been reported, a number of template-based strategies have been developed. Nielsen and Orgel, for example, have reported that oligonucleotides can template the formation of aegPNA and vice versa, although chemically activated monomers have to be used [108]. A recent report has indicated the potential of a dipeptide to promote aegPNA formation, but not in a templated manner [109]. Other recent reports by Liu and coworkers demonstrate that short aegPNA segments can assemble onto a DNA template and then be chemically ligated together [110,111]. This technique was developed for the in vitro evolution of aegPNAs that bear proteinogenic sidechains of amino acids. At the moment, there is no PNA system that would be an immediate choice for applications in synthetic biology since there is no compatibility with existing biological machinery for replication. While the template-based systems represent important steps forward, the fidelity and efficiency of these systems is not quite at the level that would likely be needed in synthetic biology.
Conclusions
Nucleic acid analogs that are useful in synthetic biology are close to becoming widely applicable. Currently, the most promising analogues that could be applied include alterations to the phosphodiester backbone and to the nucleobases, as well as some of the alternatives to the ribofuranosyl sugar (namely TNA, LNA, HNA, and GNA). It is interesting to note that relatively few attempts have been made to simultaneously incorporate different strategies for mimicking nucleic acids into a single monomeric unit. For example, what are the properties of an oligomer made with a Z:P base pair attached to a threofuranosyl sugar where each monomer is linked together by phosphorothioate units? The numerous possible combinations of the existing components of synthetic nucleic acid mimics constitute a promising area for further development.
Acknowledgments
Support from the intramural research program at NIDDK, NIH is gratefully acknowledged. I would also like to sincerely thank Dr. Ethan Englund for critical reading and comments on this manuscript.
References
- 1.Luisi PL. Chemical aspects of synthetic biology. Chem Biodivers. 2007;4:603–621. doi: 10.1002/cbdv.200790053. [DOI] [PubMed] [Google Scholar]
- 2.Sismour AM, Benner SA. Synthetic biology. Expert Opin Biol Ther. 2005;5:1409–1414. doi: 10.1517/14712598.5.11.1409. [DOI] [PubMed] [Google Scholar]
- 3.Benner SA, Sismour AM. Synthetic biology. Nat Rev Genet. 2005;6:533–543. doi: 10.1038/nrg1637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Potthast T. Paradigm shifts versus fashion shifts?Systems and synthetic biology as new epistemic entities in understanding and making ‘life’. EMBO Rep. 2009;10 1:S42–45. doi: 10.1038/embor.2009.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Morange M. A new revolution? The place of systems biology and synthetic biology in the history of biology. EMBO Rep. 2009;10 1:S50–53. doi: 10.1038/embor.2009.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Deamer D. On the origin of systems. Systems biology, synthetic biology and the origin of life. EMBO Rep. 2009;10 1:S1–4. doi: 10.1038/embor.2009.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7•.Loakes D, Holliger P. Darwinian chemistry: towards the synthesis of a simple cell. Mol Biosyst. 2009;5:686–694. doi: 10.1039/b904024b. An excellent account of the challenges faced in the synthesis of a simple protocell. [DOI] [PubMed] [Google Scholar]
- 8.Mittapalli GK, Reddy KR, Xiong H, Munoz O, Han B, De Riccardis F, Krishnamurthy R, Eschenmoser A. Mapping the landscape of potentially primordial informational oligomers: oligodipeptides and oligodipeptoids tagged with triazines as recognition elements. Angew Chem Int Ed Engl. 2007;46:2470–2477. doi: 10.1002/anie.200603207. [DOI] [PubMed] [Google Scholar]
- 9.Mittapalli GK, Osornio YM, Guerrero MA, Reddy KR, Krishnamurthy R, Eschenmoser A. Mapping the landscape of potentially primordial informational oligomers: oligodipeptides tagged with 2,4-disubstituted 5-aminopyrimidines as recognition elements. Angew Chem Int Ed Engl. 2007;46:2478–2484. doi: 10.1002/anie.200603209. [DOI] [PubMed] [Google Scholar]
- 10.Eschenmoser A. Chemical etiology of nucleic acid structure. Science. 1999;284:2118–2124. doi: 10.1126/science.284.5423.2118. [DOI] [PubMed] [Google Scholar]
- 11.Jung KH, Marx A. Nucleotide analogues as probes for DNA polymerases. Cell Mol Life Sci. 2005;62:2080–2091. doi: 10.1007/s00018-005-5117-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12•.Lartigue C, Vashee S, Algire MA, Chuang RY, Benders GA, Ma L, Noskov VN, Denisova EA, Gibson DG, Assad-Garcia N, et al. Creating Bacterial Strains from Genomes That Have Been Cloned and Engineered in Yeast. Science. 2009 doi: 10.1126/science.1173759. A report on how to transfer genes between bacteria and yeast. [DOI] [PubMed] [Google Scholar]
- 13.Wilson C, Keefe AD. Building oligonucleotide therapeutics using non-natural chemistries. Curr Opin Chem Biol. 2006;10:607–614. doi: 10.1016/j.cbpa.2006.10.001. [DOI] [PubMed] [Google Scholar]
- 14.Mansy SS, Schrum JP, Krishnamurthy M, Tobe S, Treco DA, Szostak JW. Template-directed synthesis of a genetic polymer in a model protocell. Nature. 2008;454:122–125. doi: 10.1038/nature07018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pereira de Souza T, Stano P, Luisi PL. The minimal size of liposome-based model cells brings about a remarkably enhanced entrapment and protein synthesis. Chembiochem. 2009;10:1056–1063. doi: 10.1002/cbic.200800810. [DOI] [PubMed] [Google Scholar]
- 16.Faria M, Ulrich H. Sugar boost: when ribose modifications improve oligonucleotide performance. Curr Opin Mol Ther. 2008;10:168–175. [PubMed] [Google Scholar]
- 17.Behlke MA. Chemical modification of siRNAs for in vivo use. Oligonucleotides. 2008;18:305–319. doi: 10.1089/oli.2008.0164. [DOI] [PubMed] [Google Scholar]
- 18.Prakash TP, Bhat B. 2′-Modified oligonucleotides for antisense therapeutics. Curr Top Med Chem. 2007;7:641–649. doi: 10.2174/156802607780487713. [DOI] [PubMed] [Google Scholar]
- 19.Micklefield J. Backbone modification of nucleic acids: synthesis, structure and therapeutic applications. Curr Med Chem. 2001;8:1157–1179. doi: 10.2174/0929867013372391. [DOI] [PubMed] [Google Scholar]
- 20.Griffiths AD, Potter BV, Eperon IC. Stereospecificity of nucleases towards phosphorothioate-substituted RNA: stereochemistry of transcription by T7 RNA polymerase. Nucleic Acids Res. 1987;15:4145–4162. doi: 10.1093/nar/15.10.4145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yang Z, Sismour AM, Benner SA. Nucleoside alpha-thiotriphosphates, polymerases and the exonuclease III analysis of oligonucleotides containing phosphorothioate linkages. Nucleic Acids Res. 2007;35:3118–3127. doi: 10.1093/nar/gkm168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ueda T, Tohda H, Chikazumi N, Eckstein F, Watanabe K. Phosphorothioate-containing RNAs show mRNA activity in the prokaryotic translation systems in vitro. Nucleic Acids Res. 1991;19:547–552. doi: 10.1093/nar/19.3.547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23•.Wang L, Chen S, Xu T, Taghizadeh K, Wishnok JS, Zhou X, You D, Deng Z, Dedon PC. Phosphorothioation of DNA in bacteria by dnd genes. Nat Chem Biol. 2007;3:709–710. doi: 10.1038/nchembio.2007.39. A remarkable discovery that bacteria naturally make phosphorothioate linkages in their genes. [DOI] [PubMed] [Google Scholar]
- 24.Xu T, Liang J, Chen S, Wang L, He X, You D, Wang Z, Li A, Xu Z, Zhou X, et al. DNA phosphorothioation in Streptomyces lividans: mutational analysis of the dnd locus. BMC Microbiol. 2009;9:41. doi: 10.1186/1471-2180-9-41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ou HY, He X, Shao Y, Tai C, Rajakumar K, Deng Z. dndDB: a database focused on phosphorothioation of the DNA backbone. PLoS One. 2009;4:e5132. doi: 10.1371/journal.pone.0005132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Shaw BR, Dobrikov M, Wang X, Wan J, He K, Lin JL, Li P, Rait V, Sergueeva ZA, Sergueev D. Reading, writing, and modulating genetic information with boranophosphate mimics of nucleotides, DNA, and RNA. Ann N Y Acad Sci. 2003;1002:12–29. doi: 10.1196/annals.1281.004. [DOI] [PubMed] [Google Scholar]
- 27.Renders M, Emmerechts G, Rozenski J, Krecmerova M, Holy A, Herdewijn P. Enzymatic synthesis of phosphonomethyl oligonucleotides by therminator polymerase. Angew Chem Int Ed Engl. 2007;46:2501–2504. doi: 10.1002/anie.200603435. [DOI] [PubMed] [Google Scholar]
- 28.Renders M, Lievrouw R, Krecmerova M, Holy A, Herdewijn P. Enzymatic Polymerization of Phosphonate Nucleosides. Chembiochem. 2008;9:2883–2888. doi: 10.1002/cbic.200800494. [DOI] [PubMed] [Google Scholar]
- 29.Loakes D, Holliger P. Polymerase engineering: towards the encoded synthesis of unnatural biopolymers. Chem Commun (Camb) 2009:4619–4631. doi: 10.1039/b903307f. [DOI] [PubMed] [Google Scholar]
- 30.Mayer G. The chemical biology of aptamers. Angew Chem Int Ed Engl. 2009;48:2672–2689. doi: 10.1002/anie.200804643. [DOI] [PubMed] [Google Scholar]
- 31.Keefe AD, Cload ST. SELEX with modified nucleotides. Curr Opin Chem Biol. 2008;12:448–456. doi: 10.1016/j.cbpa.2008.06.028. [DOI] [PubMed] [Google Scholar]
- 32.Kang J, Lee MS, Copland JA, 3rd, Luxon BA, Gorenstein DG. Combinatorial selection of a single stranded DNA thioaptamer targeting TGF-beta1 protein. Bioorg Med Chem Lett. 2008;18:1835–1839. doi: 10.1016/j.bmcl.2008.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chelliserrykattil J, Ellington AD. Evolution of a T7 RNA polymerase variant that transcribes 2′-O-methyl RNA. Nat Biotechnol. 2004;22:1155–1160. doi: 10.1038/nbt1001. [DOI] [PubMed] [Google Scholar]
- 34.Hocek M, Fojta M. Cross-coupling reactions of nucleoside triphosphates followed by polymerase incorporation. Construction and applications of base-functionalized nucleic acids. Org Biomol Chem. 2008;6:2233–2241. doi: 10.1039/b803664k. [DOI] [PubMed] [Google Scholar]
- 35.Knudsen SM, Robertson MP, Ellington AD. In vitro selection using modified or unnatural nucleotides. CurrProtoc Nucleic Acid Chem. 2002;Chapter 9(Unit 9 6) doi: 10.1002/0471142700.nc0906s07. [DOI] [PubMed] [Google Scholar]
- 36•.Krueger AT, Kool ET. Redesigning the architecture of the base pair: toward biochemical and biological function of new genetic sets. Chem Biol. 2009;16:242–248. doi: 10.1016/j.chembiol.2008.12.004. A review on the analogs of nucleobases. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Benner SA. Understanding nucleic acids using synthetic chemistry. Acc Chem Res. 2004;37:784–797. doi: 10.1021/ar040004z. [DOI] [PubMed] [Google Scholar]
- 38.Sismour AM, Benner SA. The use of thymidine analogs to improve the replication of an extra DNA base pair: a synthetic biological system. Nucleic Acids Res. 2005;33:5640–5646. doi: 10.1093/nar/gki873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yang Z, Hutter D, Sheng P, Sismour AM, Benner SA. Artificially expanded genetic information system: a new base pair with an alternative hydrogen bonding pattern. Nucleic Acids Res. 2006;34:6095–6101. doi: 10.1093/nar/gkl633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yang Z, Sismour AM, Sheng P, Puskar NL, Benner SA. Enzymatic incorporation of a third nucleobase pair. Nucleic Acids Res. 2007;35:4238–4249. doi: 10.1093/nar/gkm395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Havemann SA, Hoshika S, Hutter D, Benner SA. Incorporation of multiple sequential pseudothymidines by DNA polymerases and their impact on DNA duplex structure. Nucleosides Nucleotides Nucleic Acids. 2008;27:261–278. doi: 10.1080/15257770701853679. [DOI] [PubMed] [Google Scholar]
- 42.Krueger AT, Lu H, Lee AH, Kool ET. Synthesis and properties of size-expanded DNAs: toward designed, functional genetic systems. Acc Chem Res. 2007;40:141–150. doi: 10.1021/ar068200o. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43•.Delaney JC, Gao J, Liu H, Shrivastav N, Essigmann JM, Kool ET. Efficient replication bypass of size-expanded DNA base pairs in bacterial cells. Angew Chem Int Ed Engl. 2009;48:4524–4527. doi: 10.1002/anie.200805683. A report on how bacteria tolerate xDNA bases. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chelliserrykattil J, Lu H, Lee AH, Kool ET. Polymerase amplification, cloning, and gene expression of benzo-homologous “yDNA” base pairs. Chembiochem. 2008;9:2976–2980. doi: 10.1002/cbic.200800339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kool ET, Sintim HO. The difluorotoluene debate--a decade later. Chem Commun (Camb) 2006:3665–3675. doi: 10.1039/b605414e. [DOI] [PubMed] [Google Scholar]
- 46.Kimoto M, Kawai R, Mitsui T, Yokoyama S, Hirao I. An unnatural base pair system for efficient PCR amplification and functionalization of DNA molecules. Nucleic Acids Res. 2009;37:e14. doi: 10.1093/nar/gkn956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Seo YJ, Hwang GT, Ordoukhanian P, Romesberg FE. Optimization of an Unnatural Base Pair toward Natural-Like Replication. J Am Chem Soc. 2009;131:3246–3252. doi: 10.1021/ja807853m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48•.Seo YJ, Matsuda S, Romesberg FE. Transcription of an expanded genetic alphabet. J Am Chem Soc. 2009;131:5046–5047. doi: 10.1021/ja9006996. A report on the use of 5SICS:NaM nucleobase pair in replication and transcription. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Schoning K, Scholz P, Guntha S, Wu X, Krishnamurthy R, Eschenmoser A. Chemical etiology of nucleic acid structure: the alpha-threofuranosyl-(3′-->2′) oligonucleotide system. Science. 2000;290:1347–1351. doi: 10.1126/science.290.5495.1347. [DOI] [PubMed] [Google Scholar]
- 50.Eschenmoser A. The TNA-family of nucleic acid systems: properties and prospects. Orig Life Evol Biosph. 2004;34:277–306. doi: 10.1023/b:orig.0000016450.59665.f4. [DOI] [PubMed] [Google Scholar]
- 51.Ebert MO, Mang C, Krishnamurthy R, Eschenmoser A, Jaun B. The structure of a TNA-TNA complex in solution: NMR study of the octamer duplex derived from alpha-(L)-threofuranosyl-(3′-2′)-CGAATTCG. J Am Chem Soc. 2008;130:15105–15115. doi: 10.1021/ja8041959. [DOI] [PubMed] [Google Scholar]
- 52.Pallan PS, Wilds CJ, Wawrzak Z, Krishnamurthy R, Eschenmoser A, Egli M. Why does TNA cross-pair more strongly with RNA than with DNA? an answer from X-ray analysis. Angew Chem Int Ed Engl. 2003;42:5893–5895. doi: 10.1002/anie.200352553. [DOI] [PubMed] [Google Scholar]
- 53.Horhota A, Zou K, Ichida JK, Yu B, McLaughlin LW, Szostak JW, Chaput JC. Kinetic analysis of an efficient DNA-dependent TNA polymerase. J Am Chem Soc. 2005;127:7427–7434. doi: 10.1021/ja0428255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ichida JK, Horhota A, Zou K, McLaughlin LW, Szostak JW. High fidelity TNA synthesis by Therminator polymerase. Nucleic Acids Res. 2005;33:5219–5225. doi: 10.1093/nar/gki840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Petersen M, Wengel J. LNA: a versatile tool for therapeutics and genomics. Trends in Biotechnology. 2003;21:74–81. doi: 10.1016/S0167-7799(02)00038-0. [DOI] [PubMed] [Google Scholar]
- 56.Vester B, Wengel J. LNA (Locked nucleic acid): High-affinity targeting of complementary RNA and DNA. Biochemistry. 2004;43:13233–13241. doi: 10.1021/bi0485732. [DOI] [PubMed] [Google Scholar]
- 57.Koshkin AA, Nielsen P, Meldgaard M, Rajwanshi VK, Singh SK, Wengel J. LNA (locked nucleic acid): An RNA mimic forming exceedingly stable LNA : LNA duplexes. Journal of the American Chemical Society. 1998;120:13252–13253. [Google Scholar]
- 58.Nielsen KE, Rasmussen J, Kumar R, Wengel J, Jacobsen JP, Petersen M. NMR studies of fully modified locked nucleic acid (LNA) hybrids: Solution structure of an LNA: RNA hybrid and characterization of an LNA: DNA hybrid. Bioconjugate Chemistry. 2004;15:449–457. doi: 10.1021/bc034145h. [DOI] [PubMed] [Google Scholar]
- 59.Petersen M, Bondensgaard K, Wengel J, Jacobsen JP. Locked nucleic acid (LNA) recognition of RNA: NMR solution structures of LNA: RNA hybrids. Journal of the American Chemical Society. 2002;124:5974–5982. doi: 10.1021/ja012288d. [DOI] [PubMed] [Google Scholar]
- 60.Lebars I, Richard T, Di Primo C, Toulme JJ. LNA derivatives of a kissing aptamer targeted to the trans-activating responsive RNA element of HIV-1. Blood Cells Mol Dis. 2007;38:204–209. doi: 10.1016/j.bcmd.2006.11.008. [DOI] [PubMed] [Google Scholar]
- 61.Lebars I, Richard T, Di Primo C, Toulme JJ. NMR structure of a kissing complex formed between the TAR RNA element of HIV-1 and a LNA-modified aptamer. Nucleic Acids Res. 2007;35:6103–6114. doi: 10.1093/nar/gkm655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Vester B, Lundberg LB, Sorensen MD, Babu BR, Douthwaite S, Wengel J. LNAzymes: Incorporation of LNA-type monomers into DNAzymes markedly increases RNA cleavage. Journal of the American Chemical Society. 2002;124:13682–13683. doi: 10.1021/ja0276220. [DOI] [PubMed] [Google Scholar]
- 63.Veedu RN, Wengel J. Locked nucleic acid nucleoside triphosphates and polymerases: on the way towards evolution of LNA aptamers. Molecular Biosystems. 2009;5:787–792. doi: 10.1039/b905513b. [DOI] [PubMed] [Google Scholar]
- 64.Veedu RN, Vester B, Wengel J. Polymerase chain reaction and transcription using locked nucleic acid nucleotide triphosphates. Journal of the American Chemical Society. 2008;130:8124–+. doi: 10.1021/ja801389n. [DOI] [PubMed] [Google Scholar]
- 65.Babu BR, Raunak, Poopeiko NE, Juhl M, Bond AD, Parmar VS, Wengel J. XNA (xylo nucleic acid): A summary and new derivatives. European Journal of Organic Chemistry. 2005:2297–2321. [Google Scholar]
- 66.Christensen NK, Bryld T, Sorensen MD, Arar K, Wengel J, Nielsen P. Parallel nucleic acid recognition by the LNA (locked nucleic acid) stereoisomers beta-L-LNA and alpha-D-LNA; studies in the mirror image world. Chemical Communications. 2004:282–283. doi: 10.1039/b312321a. [DOI] [PubMed] [Google Scholar]
- 67.Lauritsen A, Dahl BM, Dahl O, Vester B, Wengel J. Methylphosphonate LNA: A locked nucleic acid with a methylphosphonate linkage. Bioorganic & Medicinal Chemistry Letters. 2003;13:253–256. doi: 10.1016/s0960-894x(02)00882-x. [DOI] [PubMed] [Google Scholar]
- 68.Sorensen MD, Kvaerno L, Bryld T, Hakansson AE, Verbeure B, Gaubert G, Herdewijn P, Wengel J. alpha-L-ribo-configured locked nucleic acid (alpha-L-LNA): Synthesis and properties. Journal of the American Chemical Society. 2002;124:2164–2176. doi: 10.1021/ja0168763. [DOI] [PubMed] [Google Scholar]
- 69.Rajwanshi VK, Hakansson AE, Sorensen MD, Pitsch S, Singh SK, Kumar R, Nielsen P, Wengel J. The eight stereoisomers of LNA (Locked nucleic acid): A remarkable family of strong RNA binding molecules. Angewandte Chemie-International Edition. 2000;39:1656–+. doi: 10.1002/(sici)1521-3773(20000502)39:9<1656::aid-anie1656>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- 70.Rajwanshi VK, Hakansson AE, Dahl BM, Wengel J. LNA stereoisomers: xylo- LNA (beta-D-xylo configured locked nucleic acid) and alpha-L-LNA (alpha-L-ribo configured locked nucleic acid) Chemical Communications. 1999:1395–1396. [Google Scholar]
- 71••.Herdewijn P, Marliere P. Toward Safe Genetically Modified Organisms through the Chemical Diversification of Nucleic Acids. Chemistry & Biodiversity. 2009;6:791–808. doi: 10.1002/cbdv.200900083. An overview of many nucleic acid analogs and their potential applications. [DOI] [PubMed] [Google Scholar]
- 72.Abramov M, Schepers G, Van Aerschot A, Van Hummelen P, Herdewijn P. HNA and ANA high-affinity arrays for detections of DNA and RNA single-base mismatches. Biosens Bioelectron. 2008;23:1728–1732. doi: 10.1016/j.bios.2008.01.033. [DOI] [PubMed] [Google Scholar]
- 73.Nauwelaerts K, Lescrinier E, Sclep G, Herdewijn P. Cyclohexenyl nucleic acids: conformationally flexible oligonucleotides. Nucleic Acids Research. 2005;33:2452–2463. doi: 10.1093/nar/gki538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang J, Verbeure B, Luyten I, Froeyen M, Hendrix C, Rosemeyer H, Seela F, Van Aerschot A, Herdewijn P. Cyclohexene nucleic acids (CeNA) form stable duplexes with RNA and induce RNase H activity. Nucleosides Nucleotides Nucleic Acids. 2001;20:785–788. doi: 10.1081/NCN-100002430. [DOI] [PubMed] [Google Scholar]
- 75.Verbeure B, Lescrinier E, Wang J, Herdewijn P. RNase H mediated cleavage of RNA by cyclohexene nucleic acid (CeNA) Nucleic Acids Research. 2001;29:4941–4947. doi: 10.1093/nar/29.24.4941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Hendrix C, Rosemeyer H, Verheggen I, Seela F, VanAerschot A, Herdewijn P. 1′,5′-anhydrohexitol oligonucleotides: Synthesis, base pairing and recognition by regular oligodeoxyribonucleotides and oligoribonucleotides. Chemistry-a European Journal. 1997;3:110–120. [Google Scholar]
- 77.Allart B, Khan K, Rosemeyer H, Schepers G, Hendrix C, Rothenbacher K, Seela F, Van Aerschot A, Herdewijn P. D-Altritol nucleic acids (ANA): Hybridisation properties, stability, and initial structural analysis. Chemistry-a European Journal. 1999;5:2424–2431. [Google Scholar]
- 78.Lescrinier E, Esnouf R, Schraml J, Busson R, Heus HA, Hilbers CW, Herdewijn P. Solution structure of a HNA-RNA hybrid. Chemistry & Biology. 2000;7:719–731. doi: 10.1016/s1074-5521(00)00017-x. [DOI] [PubMed] [Google Scholar]
- 79.Vastmans K, Pochet S, Peys A, Kerremans L, Van Aerschot A, Hendrix C, Marliere P, Herdewijn P. Enzymatic incorporation in DNA of 1,5-anhydrohexitol nucleotides. Biochemistry. 2000;39:12757–12765. doi: 10.1021/bi001297g. [DOI] [PubMed] [Google Scholar]
- 80.Vastmans K, Froeyen M, Kerremans L, Pochet S, Herdewijn P. Reverse transcriptase incorporation of 1,5-anhydrohexitol nucleotides. Nucleic Acids Res. 2001;29:3154–3163. doi: 10.1093/nar/29.15.3154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lavrik IN, Avdeeva ON, Dontsova OA, Froeyen M, Herdewijn PA. Translational properties of mHNA, a messenger RNA containing anhydrohexitol nucleotides. Biochemistry. 2001;40:11777–11784. doi: 10.1021/bi010836u. [DOI] [PubMed] [Google Scholar]
- 82.Pochet S, Kaminski PA, Van Aerschot A, Herdewijn P, Marliere P. Replication of hexitol oligonucleotides as a prelude to the propagation of a third type of nucleic acid in vivo. C R Biol. 2003;326:1175–1184. doi: 10.1016/j.crvi.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 83.Renders M, Abramov M, Froeyen M, Herdewijn P. Polymerase-Catalysed Incorporation of Glucose Nucleotides into a DNA Duplex. Chemistry-a European Journal. 2009;15:5463–5470. doi: 10.1002/chem.200801951. [DOI] [PubMed] [Google Scholar]
- 84.Joyce GF, Schwartz AW, Miller SL, Orgel LE. The case for an ancestral genetic system involving simple analogues of the nucleotides. Proc Natl Acad Sci U S A. 1987;84:4398–4402. doi: 10.1073/pnas.84.13.4398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Orgel LE. Prebiotic chemistry and the origin of the RNA world. Crit Rev Biochem Mol Biol. 2004;39:99–123. doi: 10.1080/10409230490460765. [DOI] [PubMed] [Google Scholar]
- 86.Orgel LE. Some consequences of the RNA world hypothesis. Orig Life Evol Biosph. 2003;33:211–218. doi: 10.1023/a:1024616317965. [DOI] [PubMed] [Google Scholar]
- 87.Heuberger BD, Switzer C. A pre-RNA candidate revisited: both enantiomers of flexible nucleoside triphosphates are DNA polymerase substrates. J Am Chem Soc. 2008;130:412–413. doi: 10.1021/ja0770680. [DOI] [PubMed] [Google Scholar]
- 88.Zhang L, Peritz A, Meggers E. A simple glycol nucleic acid. J Am Chem Soc. 2005;127:4174–4175. doi: 10.1021/ja042564z. [DOI] [PubMed] [Google Scholar]
- 89.Schlegel MK, Peritz AE, Kittigowittana K, Zhang L, Meggers E. Duplex formation of the simplified nucleic acid GNA. Chembiochem. 2007;8:927–932. doi: 10.1002/cbic.200600435. [DOI] [PubMed] [Google Scholar]
- 90.Chen JJ, Tsai CH, Cai X, Horhota AT, McLaughlin LW, Szostak JW. Enzymatic primer-extension with glycerol-nucleoside triphosphates on DNA templates. PLoS One. 2009;4:e4949. doi: 10.1371/journal.pone.0004949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Tsai CH, Chen J, Szostak JW. Enzymatic synthesis of DNA on glycerol nucleic acid templates without stable duplex formation between product and template. Proc Natl Acad Sci U S A. 2007;104:14598–14603. doi: 10.1073/pnas.0704211104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Zhou D, Froeyen M, Rozenski J, Van Aerschot A, Herdewijn P. Chemical etiology of nucleic acids: aminopropyl nucleic acids (APNAs) Chem Biodivers. 2007;4:740–761. doi: 10.1002/cbdv.200790061. [DOI] [PubMed] [Google Scholar]
- 93.Nielsen PE, Egholm M, Berg RH, Buchardt O. Sequence-selective recognition of DNA by strand displacement with a thymine-substituted polyamide. Science. 1991;254:1497–1500. doi: 10.1126/science.1962210. [DOI] [PubMed] [Google Scholar]
- 94••.Ura Y, Beierle JM, Leman LJ, Orgel LE, Ghadiri MR. Self-assembling sequence-adaptive peptide nucleic acids. Science. 2009;325:73–77. doi: 10.1126/science.1174577. A unique approach to template recognition and assembly. [DOI] [PubMed] [Google Scholar]
- 95.Egholm M, Buchardt O, Christensen L, Behrens C, Freier SM, Driver DA, Berg RH, Kim SK, Norden B, Nielsen PE. PNA hybridizes to complementary oligonucleotides obeying the Watson-Crick hydrogen-bonding rules. Nature. 1993;365:566–568. doi: 10.1038/365566a0. [DOI] [PubMed] [Google Scholar]
- 96.Abes S, Ivanova GD, Abes R, Arzumanov AA, Williams D, Owen D, Lebleu B, Gait MJ. Peptide-based delivery of steric-block PNA oligonucleotides. Methods Mol Biol. 2009;480:85–99. doi: 10.1007/978-1-59745-429-2_6. [DOI] [PubMed] [Google Scholar]
- 97.Galbiati S, Foglieni B, Travi M, Curcio C, Restagno G, Sbaiz L, Smid M, Pasi F, Ferrari A, Ferrari M, et al. Peptide-nucleic acid-mediated enriched polymerase chain reaction as a key point for noninvasive prenatal diagnosis of beta-thalassemia. Haematologica-the Hematology Journal. 2008;93:610–614. doi: 10.3324/haematol.11895. [DOI] [PubMed] [Google Scholar]
- 98.Chaubey B, Tripathi S, Pandey VN. Single acute-dose and repeat-doses toxicity of anti-HIV-1 PNA TAR-penetratin conjugate after intraperitoneal administration to mice. Oligonucleotides. 2008;18:9–20. doi: 10.1089/oli.2007.0088. [DOI] [PubMed] [Google Scholar]
- 99.Zhang N, Appella DH. Colorimetric detection of anthrax DNA with a peptide nucleic acid sandwich-hybridization assay. Journal of the American Chemical Society. 2007;129:8424–8425. doi: 10.1021/ja072744j. [DOI] [PubMed] [Google Scholar]
- 100.Tovar-Salazar A, Dhawan J, Lovejoy A, Liu QA, Gifford AN. Preparation of radioiodinated peptide nucleic acids with high specific activity. Anal Biochem. 2007;360:92–98. doi: 10.1016/j.ab.2006.10.008. [DOI] [PubMed] [Google Scholar]
- 101.Tian X, Aruva MR, Zhang K, Shanthly N, Cardi CA, Thakur ML, Wickstrom E. PET imaging of CCND1 mRNA in human MCF7 estrogen receptor positive breast cancer xenografts with oncogene-specific [64Cu]chelator-peptide nucleic acid-IGF1 analog radiohybridization probes. J Nucl Med. 2007;48:1699–1707. doi: 10.2967/jnumed.107.042499. [DOI] [PubMed] [Google Scholar]
- 102.Liu ZC, Shin DS, Shokouhimehr M, Lee KN, Yoo BW, Kim YK, Lee YS. Light- directed synthesis of peptide nucleic acids (PNAs) chips. Biosens Bioelectron. 2007;22:2891–2897. doi: 10.1016/j.bios.2006.12.005. [DOI] [PubMed] [Google Scholar]
- 103.Forrest GN. PNA FISH: Present and future impact on patient management. Expert Review of Molecular Diagnostics. 2007;7:231–236. doi: 10.1586/14737159.7.3.231. [DOI] [PubMed] [Google Scholar]
- 104.Fabris L, Dante M, Braun G, Lee SJ, Reich NO, Moskovits M, Nguyen TQ, Bazan GC. A heterogeneous PNA-based SERS method for DNA detection. J Am Chem Soc. 2007;129:6086–6087. doi: 10.1021/ja0705184. [DOI] [PubMed] [Google Scholar]
- 105.Englund EA, Appella DH. gamma-substituted peptide nucleic acids constructed from L-lysine are a versatile scaffold for multifunctional display. Angew Chem Int Ed Engl. 2007;46:1414–1418. doi: 10.1002/anie.200603483. [DOI] [PubMed] [Google Scholar]
- 106.Nielsen PE. Question 1: Peptide nucleic acids and the origin and homochirality of life. Orig Life Evol Biosph. 2007;37:323–328. doi: 10.1007/s11084-007-9105-2. [DOI] [PubMed] [Google Scholar]
- 107.Nielsen PE. Peptide nucleic acids and the origin of life. Chem Biodivers. 2007;4:1996–2002. doi: 10.1002/cbdv.200790166. [DOI] [PubMed] [Google Scholar]
- 108.Bohler C, Nielsen PE, Orgel LE. Template switching between PNA and RNA oligonucleotides. Nature. 1995;376:578–581. doi: 10.1038/376578a0. [DOI] [PubMed] [Google Scholar]
- 109.Gorlero M, Wieczorek R, Adamala K, Giorgi A, Schinina ME, Stano P, Luisi PL. Ser-His catalyses the formation of peptides and PNAs. FEBS Lett. 2009;583:153–156. doi: 10.1016/j.febslet.2008.11.052. [DOI] [PubMed] [Google Scholar]
- 110••.Kleiner RE, Brudno Y, Birnbaum ME, Liu DR. DNA-templated polymerization of side-chain-functionalized peptide nucleic acid aldehydes. J Am Chem Soc. 2008;130:4646–4659. doi: 10.1021/ja0753997. A great application of PNA in templated polymeration. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Rosenbaum DM, Liu DR. Efficient and sequence-specific DNA-templated polymerization of peptide nucleic acid aldehydes. J Am Chem Soc. 2003;125:13924–13925. doi: 10.1021/ja038058b. [DOI] [PubMed] [Google Scholar]