

Abstract
C57BL/6 mice are widely used in biomedical research for the background of genetically engineered mice (GEM) and wild-type controls with the belief that the genetic background of GEM and control mice differ significantly by only one or more altered gene. This principle, however, does have limitations due in part to the existence of multiple substrains of C57BL/6 mice that should not be used interchangeably as they can differ both genetically and phenotypically. We show here that these mispairings do occur frequently and can lead to inaccurate and conflicting findings.
Keywords: C57BL/6, Substrains, Knockout Mice, JNK2, Acetaminophen, Concanavalin-A Liver Injury and Nicotinamide Nucleotide Transhydrogenase
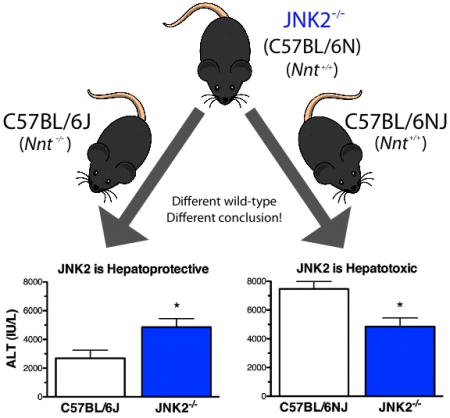
In a recent study, we found genetically engineered mice (GEM) lacking c-Jun N-terminal kinase 2 (JNK2-/-, also known as Mapk9-/-) to be more susceptible than their WT controls to acetaminophen-induced liver injury (AILI). This result suggested that JNK2 played a protective role in AILI. Another research group doing a similar experiment at the same dose of acetaminophen reported JNK2-/- mice to be less susceptible to AILI (1), a finding consistent with JNK2 having a pathologic role in AILI, as did other studies where JNK inhibitors protected WT mice from AILI (1-3). The major difference in the experimental design of the two JNK2-/- studies was that The Jackson Laboratory (JAX) was the source of our JNK2-/- mice and C57BL/6J WT controls as recommended by JAX (Table 1) (http://jaxmice.jax.org/strain/004321.html), while the other research group used a different JNK2-/- mice line that were apparently backcrossed to the same C57BL/6 mice used as controls for their studies (1). These differences led us to question whether the JNK2-/- mice used in our study were actually on a C57BL/6J background. If this were not the case and the JNK2-/- mice were actually on a different C57BL/6 substrain background, then this mismatch could likely explain the conflicting findings as it has been reported but not widely known that C57BL/6 substrains can differ genetically (4-8) and phenotypically (4, 6, 7, 9-13), especially C57BL/6J mice compared to other C57BL/6 substrains. Although other researchers have studied the mechanism of AILI in JNK2 and WT mice, direct comparison to our work is difficult because of dose differences of APAP and other confounding factors as discussed previously (14).
Table 1.
Genetically engineered C57BL/6 mice found with Nnt+/+ or Nnt+/- genotype after PCR analysis using DNA stock samples from The Jackson Laboratory
| JAX Stock # | Strain | Date of Birth | Nnt Genotypea | JAX Suggested Control | Times Backcrossedb |
|---|---|---|---|---|---|
| 004321 | Mapk9-/- | 08/19/02 | +/+ | C57BL/6J | 0 |
| 002251 | Il10-/- | 12/21/02 | +/+ | C57BL/6J | 0 |
| 005693 | Cxcr6-/- | 06/04/04 | +/+ | C57BL/6J | 0 |
| 003611 | Cd80-/- | 10/22/01 | +/+ | C57BL/6J | +1 |
| 002287 | Ifng-/- | 04/24/96 | +/- | C57BL/6J | +1 |
| 002663 | Cd4-/- | 04/15/97 | +/- | C57BL/6J | +1 |
| 003245 | Il1r1-/- | 08/18/02 | +/+ | C57BL/6J | +2 |
| 002508 | Plat-/- | 06/11/96 | +/+ | C57BL/6J | +2 |
| 002620 | Tnfrsf1b-/- | 05/23/00 | +/+ | C57BL/6J | +5 |
| 003171 | Fcgr3-/- | 06/16/04 | +/+ | C57BL/6J | +2 |
| 003641 | C3-/- | 12/06/02 | +/+ | C57BL/6J | +1 |
| 001021 | Fasl-/- | 09/04/96 | +/+ | C57BL/6J | 0 |
| 002295 | Il7r-/- | 02/16/97 | +/+ | C57BL/6J | +6 |
| 002509 | Plau-/- | 04/21/02 | +/+ | From Colony or C57BL/6J | +1 |
| 002818 | Tnfrsf1a-/- | 12/13/02 | +/+ | C57BL/6J | 0 |
| 003991 | Itgam-/- | 12/02/02 | +/+ | C57BL/6J | +1 |
| 004183 | Bak1-/- | 04/15/05 | +/+ | C57BL/6J (approximate) | +1 |
| 004650 | Tlr2-/- | 11/28/04 | +/+ | C57BL/6J | +6 |
| 004859 | Icos-/- | 4/11/05 | +/+ | C57BL/6J | 0 |
| 005576 | P2rx7-/- | 08/25/07 | +/+ | C57BL/6J (approximate) | 0 |
| 006416 | Map2k3-/- | 02/17/07 | +/+ | From Colony or C57BL/6J | 0 |
| 006659 | Cxcr5-/- | 11/03/07 | +/+ | C57BL/6J | 0 |
| 002252 | Il2-/- | 07/16/96 | +/- | From Colony or C57BL/6J | 0 |
| 002405 | Ncam1-/- | 10/01/01 | +/- | From Colony or C57BL/6J | 0 |
| 002619 | Tgfb3+/- | 08/20/97 | +/- | From Colony or C57BL/6J | 0 |
| 005582 | Cx3cr1-/- | 11/13/05 | +/- | C57BL/6J | 0 |
| 006440 | Pten+/- | 08/07/07 | +/- | C57BL/6J | +3 |
Genotyping was done using DNA stock samples collected at various times after the birth of the genetically engineered mice.
Number of times the genetically engineered mice were backcrossed again, as of April 2011, to C57BL/6 WT mice since the original DNA stock sample was collected.
When a DNA stock sample from a JNK2-/- mouse was sent to us from JAX, PCR analysis revealed that the mouse was homozygous for the intact WT allele of nicotinamide nucleotide transhydrogenase (Nnt+/+) and not the homozygous mutant Nnt-/- allele, which is unique to the C57BL/6J substrain of C57BL/6 mice (5) (Table 1 and Fig. S1). This finding was confirmed when PCR analysis was repeated with DNA from tails of several JNK2-/- mice more recently (results not shown), establishing beyond doubt that the JNK2-/- colony at JAX was definitely not on a C57BL/6J (Nnt-/-) background. We next assessed the effect of mispairing wild-type controls for the JNK2-/- mice by comparing them to C57BL/6NJ (Nnt+/+) and C57BL/6J (Nnt-/-) mice. The designations 6J and 6N in the nomenclature of C57BL/6 substrains refers to them being derived from C57BL/6 colonies which were isolated from the Jackson Laboratory (6J) and sent to the National Institutes of Health (6N) in 1951(5). As the first step, we compared the susceptibility of C57BL/6J (Nnt-/-) and C57BL/6NJ (Nnt+/+) substrains to AILI. C57BL/6J mice were less susceptible to AILI than the C57BL/6NJ substrain, as determined biochemically by the measurement of serum ALT activities (Fig. 1A), a biomarker of liver injury (15), and histochemically by reduced amounts of hepatic perivenous necrosis (Fig. 1B). Lastly and most importantly, we confirmed the results of our original study that JNK2-/- mice from JAX were more susceptible than C57BL/6J WT mice to AILI (14) and also showed that this finding could be reversed when JNK2-/- mice were paired with C57BL/6NJ (Nnt+/+) WT mice (Fig. 1). Overall, these results support the findings by other researchers as discussed earlier that JNK2 has a pathologic role in AILI.
Figure 1.
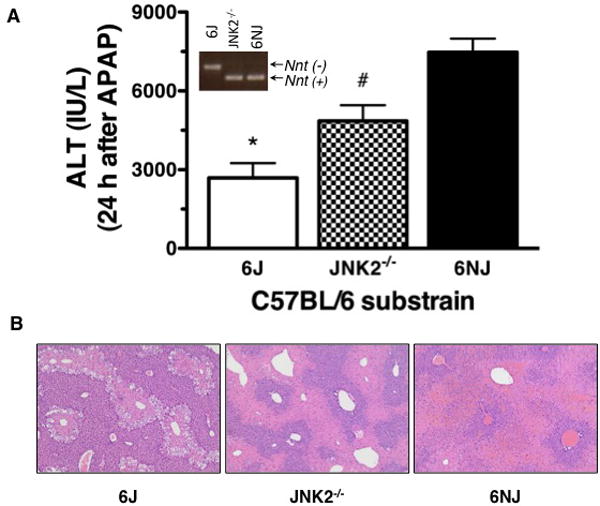
Role of c-Jun N-terminal kinase 2 in liver injury caused by acetaminophen was dependent on the substrain of C57BL/6 wild-type mice paired with c-Jun N-terminal kinase 2 deficient mice. Mice were treated with acetaminophen (APAP, 300 mg/kg intraperitoneally). Liver injury was assessed 24 hours after treatment by measurement of serum alanine aminotransferase (ALT) activity and by histopathologic examination of H&E stained liver sections; magnification 200×. (A) Serum ALT activities of mice treated with APAP represent the means ± SEM where *P < 0.05 when C57BL/6J (6J, n=5) mice were compared to C57BL/6 c-Jun N-terminal kinase 2 deficient mice (JNK2-/-, n=5) and C57BL/6NJ (6NJ, n=4) mice, while #P < 0.05 when JNK2-/- mice were compared to 6NJ mice. (B) Representative photomicrographs of liver sections of mice treated with APAP showed that the severity of hepatic necrosis followed the order of 6NJ > JNK2-/- > 6J mice. Two experiments were performed with similar results. The inset shows genotyping results where the 6J mice in this study were confirmed to be homozygous Nnt-/- whereas the JNK2-/- and 6NJ mice were homozygous Nnt+/+.
Our findings raised the possibility that mispairings of JNK2-/- and C57BL/6J WT controls mice from JAX in other studies dealing with JNK2 signaling may have also led to inaccurate interpretations of data (see Supporting Information for such misparings). A case in point is the conflicting role of JNK2 in a T-cell model of liver injury induced by concanavalin A that can also be explained by these mismatches (Fig. S2).
The studies with JNK2-/- mice prompted us to explore whether similar C57BL/6 background problems might also have occurred in investigations with other GEM. We picked an additional 79 GEM from JAX for our studies based upon the recommendation by JAX that C57BL/6J WT mice could be used as controls and the wide-spread use of these strains in studies dealing with innate and adaptive immune systems in physiology and pathology. When DNA samples from each of the GEM were genotyped, 26 of them were found to be either homozygous Nnt+/+ (19 samples) or heterozygous Nnt+/- (7 samples) (Table 1 and Fig. S1), while all others were homozygous Nnt-/- (Table S1). Since 12 of the 26 in addition to the JNK2-/- mice had not been backcrossed again to any C57BL/6 substrain as of April 2011 according to information from JAX websites for each of the GEM strains (Table 1), it is likely that these colonies remain Nnt+/+ except possibly for the heterozygous Nnt+/- strains. Similarly, among the 19 GEM strains that were found Nnt+/+, the 6 that were backcrossed only one extra time to C57BL/6 substrain (Table 1) would be on Nnt mixed background. Together, this suggests that many strains are still incorrectly mispaired with C57BL/6J and therefore could lead to confounding results.
Although recent studies have not uncovered genetic differences among substrains of C57BL/6N (Nnt+/+) mice (5, 8) it is possible that more complete genetic analyses in the future may reveal genetic diversity within these substrains that results in phenotypic differences among C57BL/6N substrains. If this were case, then mismatches in C57BL/6N GEM and WT controls may also be a source of inaccuracies in the literature. Another potential complicating factor is that genomic copy number variations may also lead to phenotypic differences within and between substrains of mice (16, 17). As it is known that genetic and phenotypic differences can also exist within strains of other mice that have diverged at various sites around the world, including the widely used Balb/c mouse (16, 18), it is possible that background mismatches of GEM with WT controls have occurred with these mice as well.
Our findings underscore the critical need for researchers to be extremely careful when designing experiments with GEM and WT controls on a C57BL/6 background, obtained from vendors and other researchers. Investigators can help avoid mispairing C57BL/6J (Nnt-/-) and C57BL/6N (Nnt+/+) substrains by Nnt genotyping mice prior to beginning experiments. This problem could also be alleviated if journals required authors to include in their manuscripts information concerning the source and C57BL/6 substrain background of GEM including number of times backcrossed as well as and relevant information about the WT controls used in their studies. Similarly, vendors of GEM should report similar information and as well as details of any further backcrossing of their GEM onto C57BL/6 WT mice on their websites. However, it always best to choose control WT mice that are either age-matched WT littermates of GEM or age-matched WT mice on to which the GEM were backcrossed (19). This approach would have been the optimal choice to show that JNK2-/- mice are less susceptible than WT controls to AILI.
We have also found in two distinctive animal models of liver pathology, AILI and concanavalin A-induced liver injury, that C57BL/6N mice are more susceptible than C57BL/6J mice to liver injury (Figs.1 and S2, respectively). These results were surprising as mitochondrial oxidative stress plays a pathologic role in both models of liver injury (20, 21), and because the Nnt mutation in C57BL/6J mice encodes for an inactive form of mitochondrial Nnt that normally catalyzes the interconversion of NADH into NADPH (22), which has an important antioxidant role in regenerating reduced glutathione and thioredoxin during mitochondrial oxidative stress (22). Moreover, recent studies show that additional genetic differences exist between C57BL/6J (Nnt-/-) and C57BL/6 (Nnt+/+) substrains(5, 8). It is anticipated that mechanistic studies in the future that can unravel the susceptible differences of C57BL/6 substrains mice to AILI and concanavalin A-induced liver injury could lead to the identification of potential risk factors for not drug-induced liver injury, but also for liver injury caused by other pathologies.
Supplementary Material
Acknowledgments
Funding Sources This work was supported by the Intramural Research Program of the National Institutes of Health and the National Heart, Lung, and Blood Institute.
Abbreviations
- GEM
genetically engineered mice
- WT
wild-type
- JAX
The Jackson Laboratory
- Nnt
nicotinamide nucleotide transhydrogenase
- JNK2-/-
c-Jun N-terminal kinase 2 deficient
- AILI
acetaminophen-induced liver injury
- APAP
acetaminophen
- ALT
alanine aminotransferase
Footnotes
Supporting Information. Experimental procedures; Table of genotyping results showing GEM from JAX with Nnt-/- genotype corresponding to mice with a C57BL/6J background; Figure of genotyping results showing GEM from JAX with Nnt+/+ and Nnt+/- genotype that do not correspond to mice with a C57BL/6J background; Figure and results showing that the role of JNK2 in concanavalin A-induced liver injury can be misleading when C57BL/6 substrain backgrounds of JNK2 mice and WT controls are mispaired; Additional mispairings of C57BL/6 Substrains of GEM and WT Controls in the literature.
References
- 1.Nakagawa H, Maeda S, Hikiba Y, Ohmae T, Shibata W, Yanai A, Sakamoto K, Ogura K, Noguchi T, Karin M, Ichijo H, Omata M. Deletion of apoptosis signal-regulating kinase 1 attenuates acetaminophen-induced liver injury by inhibiting c-Jun N-terminal kinase activation. Gastroenterology. 2008;135:1311–1321. doi: 10.1053/j.gastro.2008.07.006. [DOI] [PubMed] [Google Scholar]
- 2.Gunawan BK, Liu ZX, Han D, Hanawa N, Gaarde WA, Kaplowitz N. c-Jun N-terminal kinase plays a major role in murine acetaminophen hepatotoxicity. Gastroenterology. 2006;131:165–178. doi: 10.1053/j.gastro.2006.03.045. [DOI] [PubMed] [Google Scholar]
- 3.Henderson NC, Pollock KJ, Frew J, Mackinnon AC, Flavell RA, Davis RJ, Sethi T, Simpson KJ. Critical role of c-jun (NH2) terminal kinase in paracetamol- induced acute liver failure. Gut. 2007;56:982–990. doi: 10.1136/gut.2006.104372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mulligan MK, Ponomarev I, Boehm SL, Owen JA, Levin PS, Berman AE, Blednov YA, Crabbe JC, Williams RW, Miles MF, Bergeson SE. Alcohol trait and transcriptional genomic analysis of C57BL/6 substrains. Genes Brain Behav. 2008;7:677–689. doi: 10.1111/j.1601-183X.2008.00405.x. [DOI] [PubMed] [Google Scholar]
- 5.Mekada K, Abe K, Murakami A, Nakamura S, Nakata H, Moriwaki K, Obata Y, Yoshiki A. Genetic differences among C57BL/6 substrains. Exp Anim. 2009;58:141–149. doi: 10.1538/expanim.58.141. [DOI] [PubMed] [Google Scholar]
- 6.Bothe GW, Bolivar VJ, Vedder MJ, Geistfeld JG. Genetic and behavioral differences among five inbred mouse strains commonly used in the production of transgenic and knockout mice. Genes Brain Behav. 2004;3:149–157. doi: 10.1111/j.1601-183x.2004.00064.x. [DOI] [PubMed] [Google Scholar]
- 7.Bryant CD, Zhang NN, Sokoloff G, Fanselow MS, Ennes HS, Palmer AA, McRoberts JA. Behavioral differences among C57BL/6 substrains: implications for transgenic and knockout studies. J Neurogenet. 2008;22:315–331. doi: 10.1080/01677060802357388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Zurita E, Chagoyen M, Cantero M, Alonso R, Gonzalez-Neira A, Lopez-Jimenez A, Lopez-Moreno JA, Landel CP, Benitez J, Pazos F, Montoliu L. Genetic polymorphisms among C57BL/6 mouse inbred strains. Transgenic Res. 2010 doi: 10.1007/s11248-010-9403-8. in press. [DOI] [PubMed] [Google Scholar]
- 9.Kajioka EH, Andres ML, Nelson GA, Gridley DS. Immunologic variables in male and female C57BL/6 mice from two sources. Comp Med. 2000;50:288–291. [PubMed] [Google Scholar]
- 10.Andersson KE, Immerstrand T, Sward K, Bergenstahl B, Lindholm MW, Oste R, Hellstrand P. Effects of oats on plasma cholesterol and lipoproteins in C57BL/6 mice are substrain specific. Br J Nutr. 2010;103:513–521. doi: 10.1017/S000711450999211X. [DOI] [PubMed] [Google Scholar]
- 11.Diwan BA, Blackman KE. Differential susceptibility of 3 sublines of C57BL/6 mice to the induction of colorectal tumors by 1,2-dimethylhydrazine. Cancer Lett. 1980;9:111–115. doi: 10.1016/0304-3835(80)90114-7. [DOI] [PubMed] [Google Scholar]
- 12.Nicholson A, Reifsnyder PC, Malcolm RD, Lucas CA, MacGregor GR, Zhang W, Leiter EH. Diet-induced obesity in two C57BL/6 substrains with intact or mutant nicotinamide nucleotide transhydrogenase (Nnt) gene. Obesity (Silver Spring) 2010;18:1902–1905. doi: 10.1038/oby.2009.477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ramachandra V, Phuc S, Franco AC, Gonzales RA. Ethanol preference is inversely correlated with ethanol-induced dopamine release in 2 substrains of C57BL/6 mice. Alcohol Clin Exp Res. 2007;31:1669–1676. doi: 10.1111/j.1530-0277.2007.00463.x. [DOI] [PubMed] [Google Scholar]
- 14.Bourdi M, Korrapati MC, Chakraborty M, Yee SB, Pohl LR. Protective role of c-Jun N-terminal kinase 2 in acetaminophen-induced liver injury. Biochem Biophys Res Commun. 2008;374:6–10. doi: 10.1016/j.bbrc.2008.06.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yee SB, Bourdi M, Masson MJ, Pohl LR. Hepatoprotective role of endogenous interleukin-13 in a murine model of acetaminophen-induced liver disease. Chem Res Toxicol. 2007;20:734–744. doi: 10.1021/tx600349f. [DOI] [PubMed] [Google Scholar]
- 16.Velez L, Sokoloff G, Miczek KA, Palmer AA, Dulawa SC. Differences in Aggressive Behavior and DNA Copy Number Variants Between BALB/cJ and BALB/cByJ Substrains. Behav Genet. 2010;40:201–210. doi: 10.1007/s10519-009-9325-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Watkins-Chow DE, Pavan WJ. Genomic copy number and expression variation within the C57BL/6J inbred mouse strain. Genome Res. 2008;18:60–66. doi: 10.1101/gr.6927808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Teuscher C, Blankenhorn EP, Hickey WF. Differential Susceptibility to Actively Induced Experimental Allergic Encephalomyelitis and Experimental Allergic Orchitis among Balb/C Substrains. Cellular Immunology. 1987;110:294–304. doi: 10.1016/0008-8749(87)90124-9. [DOI] [PubMed] [Google Scholar]
- 19.Sigmund CD. Viewpoint: are studies in genetically altered mice out of control? Arterioscler Thromb Vasc Biol. 2000;20:1425–1429. doi: 10.1161/01.atv.20.6.1425. [DOI] [PubMed] [Google Scholar]
- 20.Hanawa N, Shinohara M, Saberi B, Gaarde WA, Han D, Kaplowitz N. Role of JNK translocation to mitochondria leading to inhibition of mitochondria bioenergetics in acetaminophen-induced liver injury. Journal Of Biological Chemistry. 2008;283:13565–13577. doi: 10.1074/jbc.M708916200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ni HM, Chen X, Ding WX, Schuchmann M, Yin XM. Differential roles of JNK in ConA/GalN and ConA-induced liver injury in mice. Am J Pathol. 2008;173:962–972. doi: 10.2353/ajpath.2008.080358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Huang TT, Naeemuddin M, Elchuri S, Yamaguchi M, Kozy HM, Carlson EJ, Epstein CJ. Genetic modifiers of the phenotype of mice deficient in mitochondrial superoxide dismutase. Hum Mol Genet. 2006;15:1187–1194. doi: 10.1093/hmg/ddl034. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.