

Abstract
Purpose
We have previously identified specific epithelial proteins with altered expression in human diabetic central corneas. Decreased hepatocyte growth factor receptor (c-met) and increased proteinases were functionally implicated in the changes of these proteins in diabetes. The present study examined whether limbal stem cell marker patterns were altered in diabetic corneas and whether c-met gene overexpression could normalize these patterns.
Methods
Cryostat sections of 28 ex vivo and 26 organ-cultured autopsy human normal and diabetic corneas were examined by immunohistochemistry using antibodies to putative limbal stem cell markers including ATP-binding cassette sub-family G member 2 (ABCG2), N-cadherin, ΔNp63α, tenascin-C, laminin γ3 chain, keratins (K) K15, K17, K19, β1 integrin, vimentin, frizzled 7, and fibronectin. Organ-cultured diabetic corneas were studied upon transduction with adenovirus harboring c-met gene.
Results
Immunostaining for ABCG2, N-cadherin, ΔNp63α, K15, K17, K19, and β1 integrin, was significantly decreased in the stem cell-harboring diabetic limbal basal epithelium either by intensity or the number of positive cells. Basement membrane components, laminin γ3 chain, and fibronectin (but not tenascin-C) also showed a significant reduction in the ex vivo diabetic limbus. c-Met gene transduction, which normalizes diabetic marker expression and epithelial wound healing, was accompanied by increased limbal epithelial staining for K17, K19, ΔNp63α, and a diabetic marker α3β1 integrin, compared to vector-transduced corneas.
Conclusions
The data suggest that limbal stem cell compartment is altered in long-term diabetes. Gene therapy, such as with c-met overexpression, could be able to restore normal function to diabetic corneal epithelial stem cells.
Introduction
In pathological conditions, such as diabetes mellitus, the cornea is significantly affected and this can cause visual impairment. The most recognized diabetic complications in the cornea include neurotrophic corneal ulcers, filamentous keratitis, loss of corneal sensation, and a characteristic epithelial keratodystrophy, which is referred to as diabetic keratopathy [1-9]. Diabetic cornea exhibits basement membrane abnormalities, reduced numbers of hemidesmosomes, altered growth factor content and signaling, epithelial cellular enlargement, edema, and delayed wound healing resulting in persistent epithelial defects [2-4,8-11]. Treatment for diabetic keratopathy remains symptomatic [2].
Corneal epithelial renewal and healing of epithelial wounds largely depend on corneal stem cells that, at least in humans, reside in the basal epithelial layer of the corneoscleral junction, limbus [12-21]. These cells represent less than 10% of the total limbal basal epithelial cell population [22,23]. Deficiencies of or damage to these limbal epithelial stem cells (LESC) have serious implications for corneal function such as in-growth of conjunctival cells and neovascularization of the corneal stroma, which eventually lead to corneal opacity and vision loss [20,24-26]. These cells have a high capacity for self-renewal, which is retained throughout life. Corneal maintenance depends on LESC as a source of epithelial proliferation and rapid renewal through generation of transient amplifying (TA) cells, which in turn differentiate into epithelial cells during their centripetal movement [21,27-29].
Because of its role in epithelial renewal and wound healing, deficiency of the limbal niche and its residing LESC may be responsible for abnormalities in diabetic corneal epithelium. In the present paper we examined various putative stem cell markers in ex vivo diabetic and normal epithelial limbal compartment, as well as in organ-cultured diabetic corneas upon overexpression of c-met proto-oncogene shown to normalize wound healing time and epithelial marker expression [30]. Immunostaining patterns of several putative stem cell markers were altered in the diabetic limbus, and some of these patterns could be normalized by c-met overexpression. The data suggest that limbal compartment may play an important role in diabetic corneal alterations that can be corrected by gene therapy.
Methods
Tissues
Age-matched normal, diabetic (with insulin-dependent [IDDM] or non-insulin-dependent [NIDDM] diabetes), and diabetic retinopathy (DR) autopsy human corneas were obtained from the National Disease Research Interchange (NDRI, Philadelphia, PA), within 24 (for ex vivo) to 48 h after death. NDRI has a human tissue collection protocol approved by a managerial committee and subject to National Institutes of Health oversight. In this study (Table 1), 15 normal (from 13 donors, mean age 57.8±21.8 years) and 13 diabetic (from 9 donors; mean age 71.2±6.3 years; 7 with IDDM, 2 with NIDDM, 4 with DR) ex vivo corneas, as well as 13 pairs of organ-cultured diabetic corneas (from 13 donors; mean age 68.5±14.4 years; 6 with IDDM, 7 with NIDDM, 4 with DR) were used. Mean ages in all groups as well as mean disease durations for known cases in ex vivo and organ culture diabetic groups did not differ significantly. The corneas were embedded in Optimal Cutting Temperature (OCT) compound (Sakura Finetek USA, Inc., Torrance, CA) and stored at –80 °C for immunohistochemistry, or were processed for organ culture.
Table 1. Donor characteristics.
| Case number | Diabetes type | Age, sex | Diabetes duration, years | Cause of death |
|---|---|---|---|---|
|
Ex vivo normal | ||||
| 95–12 |
- |
17, F |
- |
gunshot wound |
| 95–15 |
- |
59, M |
- |
cardiac arrest |
| 95–44 |
- |
68, M |
- |
massive hemorrhage |
| 99–19 |
- |
79, F |
- |
respiratory arrest |
| 99–61 |
- |
80, F |
- |
myocardial infarction |
| 03–3 |
- |
71, M |
- |
myocardial infarction |
| 03–09 |
- |
72, M |
- |
lung cancer |
| 05–16 |
- |
56, M |
- |
ruptured aortic dissection |
| 05–25 |
- |
60, F |
- |
COPD |
| 05–26 |
- |
60, F |
- |
COPD |
| 05–45 |
- |
10, M |
- |
exsanguination |
| 05–46 |
- |
10, M |
- |
exsanguination |
| 05–56 |
- |
45, M |
cardiovascular accident |
|
| 05–60 |
- |
65, F |
- |
pneumonia |
| 10–03 |
- |
69, M |
- |
respiratory failure |
|
Ex vivo diabetic | ||||
| 95–17 |
IDDM |
79, M |
5 |
cardiac arrest |
| 95–18 |
IDDM |
79, M |
5 |
cardiac arrest |
| 96–06 |
IDDM |
69, M |
22 |
acute cardiac event |
| 96–30 |
NIDDM |
77, F |
>5 |
cardiorespiratory arrest |
| 96–46 |
IDDM, PDR |
68, F |
>48 |
cardiorespiratory arrest |
| 96–47 |
IDDM, PDR |
68, F |
>48 |
cardiorespiratory arrest |
| 96–95 |
IDDM |
63, M |
3 |
cardiovascular accident |
| 96–96 |
IDDM |
63, M |
3 |
cardiovascular accident |
| 99–08 |
IDDM, DR |
67, F |
30 |
myocardial infarction |
| 99–79 |
IDDM, PDR |
77, F |
unknown |
cardiac arrest |
| 99–80 |
IDDM, PDR |
77, F |
unknown |
cardiac arrest |
| 01–47 |
IDDM, DR |
64, F |
unknown |
ventricular arrhythmia |
| 06–26 |
NIDDM |
77, M |
15 |
intracerebral hemorrhage |
|
Organ-cultured diabetic | ||||
| 07–27 |
IDDM |
81, M |
>10 |
acute renal failure |
| 07–32 |
NIDDM |
84, M |
20 |
stroke |
| 07–34 |
IDDM, DR |
37, M |
22 |
intracranial hemorrhage |
| 08–35 |
IDDM, DR |
88, M |
38 |
cardiac arrest |
| 08–36 |
NIDDM |
82, M |
15 |
stroke |
| 08–38 |
IDDM |
73 F |
>10 |
diabetic ketoacidosis |
| 08–40 |
NIDDM |
59, M |
20 |
cardiac arrest |
| 08–44 |
IDDM, DR |
71, M |
15 |
cardiopulmonary arrest |
| 08–49 |
NIDDM |
59, F |
28 |
intracranial hemorrhage |
| 08–54 |
NIDDM |
57, M |
10 |
myocardial infarction |
| 08–57 |
IDDM |
78, F |
15 |
respiratory failure |
| 09–12 |
NIDDM, DR |
61, F |
unknown |
myocardial infarction |
| 09–16 | NIDDM | 61, F | >10 | congestive heart failure |
M, male; F, female; PDR, proliferative diabetic retinopathy; COPD, chronic obstructive pulmonary disease. In the ex vivo groups, each case number refers to one cornea; in organ-cultured diabetic group, each case number refers to a pair of fellow corneas.
Corneal organ culture and viral transduction
As described previously [30,31], after filling the corneal concavity with warm agar-collagen mixture, corneas were cultured in serum-free medium with insulin-transferrin-selenite, antibiotics and antimycotic (Invitrogen, Carlsbad, CA), at a liquid-air interface with epithelium facing upwards. Organ-cultured diabetic corneas were transduced for 48 h with 1.0–2.0×108 plaque-forming units of recombinant adenoviruses, rAV-cmet (harboring full-length c-met open reading frame) and the fellow corneas with rAV-vector (no gene inserted) as a control. Seventy-five μg/ml of sterile sildenafil citrate (Viagra®; Pfizer Corp., New York, NY) was added to the culture medium along with the viruses to increase rAV transduction efficiency [31]. Some transduced corneas were processed after 7–10 days in culture, some after wound healing experiments [30]. They were embedded in OCT and 5 μm cryostat sections cut for immunostaining on a Leica CM1850 cryostat (McBain Instruments, Chatsworth, CA).
Immunohistochemistry
The list of primary antibodies to putative stem cell markers is presented in Table 2. Different fixations such as 100% acetone at –20 °C for 10 min, 100% methanol at –20 °C for 10 min, 1% formalin (0.37% formaldehyde) in saline at room temperature for 5 min were used for different antibodies. For each marker the same exposure time was used when photographing stained sections of fellow corneas using a MicroFire digital camera (Optronics, Goleta, CA) attached to an Olympus BX40 microscope (Olympus USA, Melville, NY) and operated using PictureFrame software. Negative controls without a primary antibody were included in each experiment.
Table 2. Antibodies used in the study.
| Antigen | Antibody | Source | Dilution | Immunostaining |
|---|---|---|---|---|
| ABCG2 |
Mouse mAb MAB4155 |
Millipore |
1:50 |
- |
| ABCG2 |
Mouse mAb sc-58222 |
Santa Cruz Biotechnology |
1:5 |
+ |
| ABCG2 |
Rabbit pAb sc-25821 |
Santa Cruz Biotechnology |
1:20 |
+ |
| C/EBPδ |
Rabbit pAb sc-636 |
Santa Cruz Biotechnology |
1:20 |
- |
| Fibronectin |
Mouse mAb 568 |
[72] |
1:60 |
+ |
| Integrin β1 |
Mouse mAb MAB1959 |
Millipore |
1:50 |
+ |
| Integrin β1 |
Mouse mAb MAB2000 |
Millipore |
1:50 |
+ |
| Keratin 15 |
Mouse mAb sc-47697 |
Santa Cruz Biotechnology |
1:10 |
+ |
| Keratin 17 |
Mouse mAb E3 |
[73] |
straight |
+ |
| Keratin 17 |
Rabbit mAb #4543 |
Cell Signaling |
1:50 |
+ |
| Keratin 19 |
Mouse mAb MAB1607 |
Millipore |
1:10 |
+ |
| Keratin 19 |
Mouse mAb MA1–35554 |
Thermo Scientific |
1:10 |
+ |
| Keratin 19 |
Rabbit pAb PA1–38014 |
Thermo Scientific |
1:20 |
+ |
| Keratin 19 |
Mouse mAb MAB1608 |
Millipore |
1:10 |
+ |
| Laminin β1 |
Rat mAb LT3 |
[74] |
straight |
+ |
| Laminin β2 |
Mouse mAb C4 |
Developmental Hybridoma Bank |
straight |
+ |
| Laminin γ1 |
Rat mAb A5 |
[75] |
straight |
+ |
| Laminin γ3 |
Rabbit pAb R96 |
[76] |
1:500 |
+ |
| Laminin γ3 |
Rabbit pAb sc-25719 |
Santa Cruz Biotechnology |
1:20 |
- |
| Laminin γ3 |
Goat pAb sc-16601 |
Santa Cruz Biotechnology |
1:10 |
+ |
| N-Cadherin |
Mouse mAb 3B9 |
Invitrogen |
1:20 |
+ |
| N-Cadherin |
Rabbit pAb 12221 |
Abcam |
1:50 |
- |
| N-Cadherin |
Rabbit pAb sc-7939 |
Santa Cruz Biotechnology |
1:20 |
+ |
| Nidogen-1 |
Mouse mAb MAB2570 |
R&D Systems |
1:50 |
+ |
| Nidogen-2 |
Rabbit pAb 1080 |
[77] |
1:200 |
+ |
| Nidogen-2 |
Goat pAb sc-26132 |
Santa Cruz Biotechnology |
1:25 |
- |
| Nidogen-2 |
Goat pAb sc-26133 |
Santa Cruz Biotechnology |
1:25 |
- |
| ΔNp63 |
Goat pAb sc-8609 |
Santa Cruz Biotechnology |
1:20 |
+ |
| ΔNp63 |
Rabbit pAb 619001 |
Biolegend |
1:50 |
- |
| P63α |
Rabbit pAb |
[23] |
1:100 |
+ |
| SOD2 |
Rabbit pAb sc-30080 |
Santa Cruz Biotechnology |
1:10 |
+ |
| Vimentin |
Goat pAb sc-7558 |
Santa Cruz Biotechnology |
1:20 |
+ |
| Frizzled 7 |
Rat mAb 1981 |
R&D Systems |
1:20 |
+ |
| Tenascin-C | Mouse mAb BC2 | [78] | straight | + |
mAb, monoclonal antibody; pAb, polyclonal antibody.
Statistics
Immunostaining results were analyzed by unpaired (for ex vivo corneas; n=15 for normal, n=13 for diabetic) or paired (for organ cultured corneas; n=13 pairs) Student’s t-test (InStat, GraphPad Software, San Diego, CA). Staining intensity was scored arbitrarily as: 0 (negative), 0.5 (weak), 1 (distinct), 2 (moderate), 3 (strong), 4 (very strong). Most cases were stained at least twice with good reproducibility between experiments, and a mean intensity score from independent experiments was used for each case. The mean scores between groups (e.g., normal versus diabetic) were then compared. A p-value <0.05 was considered significant. Data are expressed as mean±standard error (SEM).
Results
Distribution of putative stem cell markers in normal and diabetic ex vivo corneas
The immunostaining patterns of several limbal and/or putative stem cell markers were altered in the ex vivo diabetic limbus compared to the normal one. Figure 1, left column, shows the normal staining patterns of cytoskeletal structural proteins of epithelial cells, keratins 15, 17, and 19 (K15, K17 and K19). These three keratins were expressed in limbal compartment but not in central ex vivo corneal epithelium, in good agreement with previous data [17,32-34]. K15 and K19 were prominently expressed in the basal limbal cells and in lesser amounts in the suprabasal and superficial layers of the limbal epithelium (Figure 1). K17 protein was usually found in clusters of limbal basal cells (Figure 1), as was vimentin (not shown here). As shown in Figure 1, right column, in the diabetic limbus staining for all three keratins decreased by both intensity and number of positive cells; the latter was most pronounced for K17. Decreased staining for these keratins in the diabetic limbus reached significance. The most commonly used putative LESC markers [20,23,35-37], ATP-binding cassette transporter Bcrp1/ABCG2 and transcription factor ΔNp63α isoform, were also significantly decreased in the diabetic limbus (Figure 2 and Figure 3). The same was true for another putative LESC marker [38], N-cadherin (Figure 3). β1 Integrin [39] that is found in basal cells as well as the suprabasal and superficial layers of the corneal epithelium was expressed much less in the diabetic limbus (Figure 4), although its localization did not agree with the presence in LESC niche only. In the diabetic limbus, reduced and discontinuous immunostaining was observed for select ECM markers, such as laminin γ3 chain (Figure 4), expressed mostly in the limbal basement membrane [40,41]. Fibronectin staining was also significantly decreased in the diabetic limbal basement membrane (Figure 4). No significant changes were observed in the diabetic limbal cells for total tenascin-C (Figure 3), a ubiquitous laminin γ1 chain, and putative LESC markers superoxide dismutase 2 (SOD2), vimentin, and a Wnt receptor, frizzled 7 (data not shown).
Figure 1.
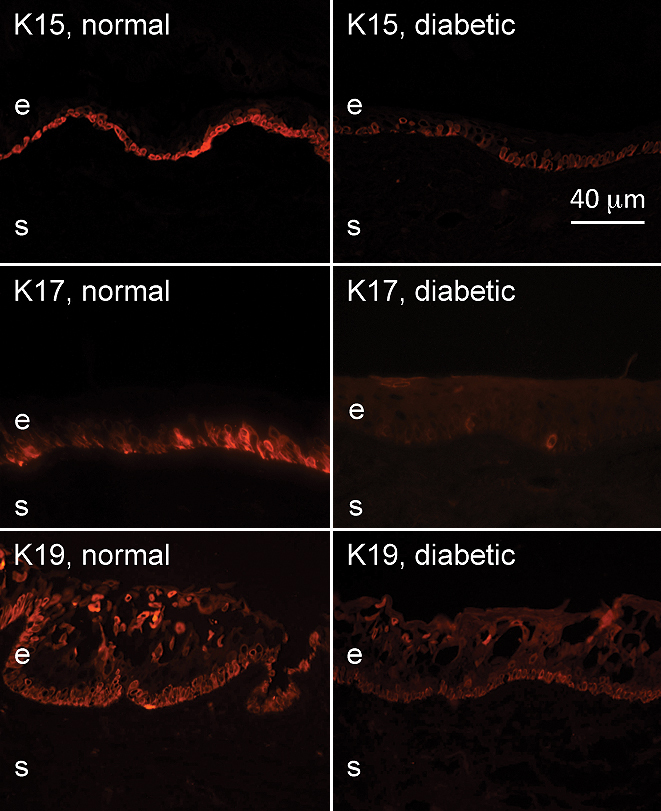
Keratin expression patterns in normal and diabetic ex vivo limbus. The staining intensity of K15, K17, and K19 was significantly decreased in the diabetic limbus. Note a reduction of K17-positive cells in the diabetic limbus as well. Here and in all other figures, each normal and diabetic pair was photographed at the same exposure times in the same staining experiments. e, epithelium, s, stroma. Bar=40 μm.
Figure 2.
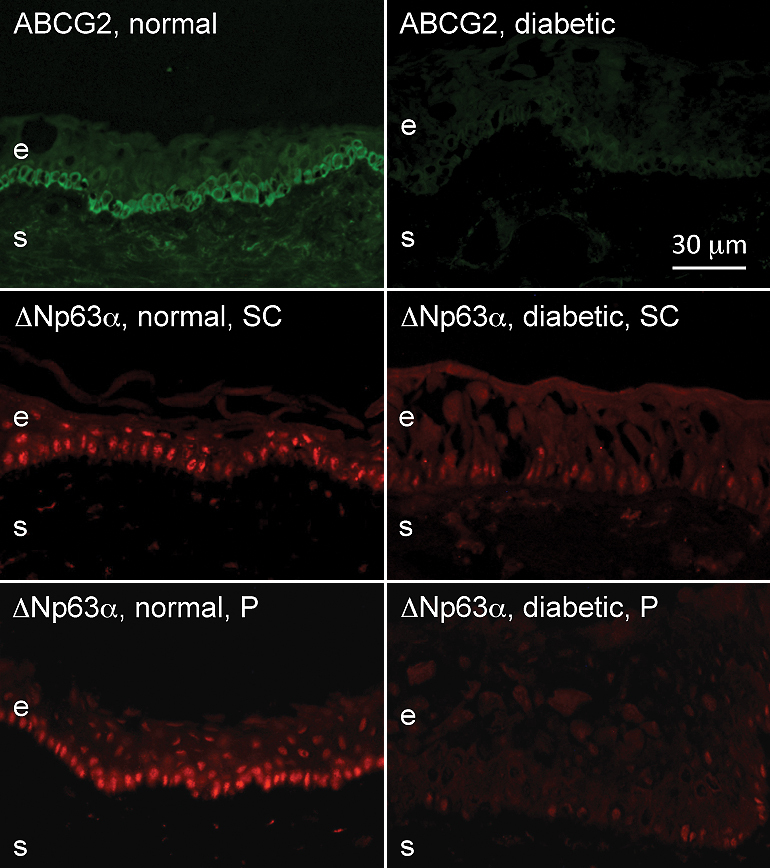
Putative LESC marker expression patterns in normal and diabetic ex vivo limbus. Note a dramatic decrease in staining intensity and the number of positive basal epithelial cells for ABCG2 and ΔNp63α in the diabetic limbus. ΔNp63α was revealed with two different antibodies (Santa Cruz, SC) and Pellegrini (P) with the same result. e, epithelium, s, stroma. Bar=30 μm.
Figure 3.

Statistical analysis of changes in the staining for various markers in diabetic versus normal ex vivo limbus. Significant staining decrease was observed for K15, K17, K19, ΔNp63α, N-cadherin, ABCG2, fibronectin, β1 integrin, and laminin γ3 chain. Data are mean±SEM. Normal, n=15; diabetic, n=13. *p<0.05; **p<0.01. Details are in the Methods section.
Figure 4.
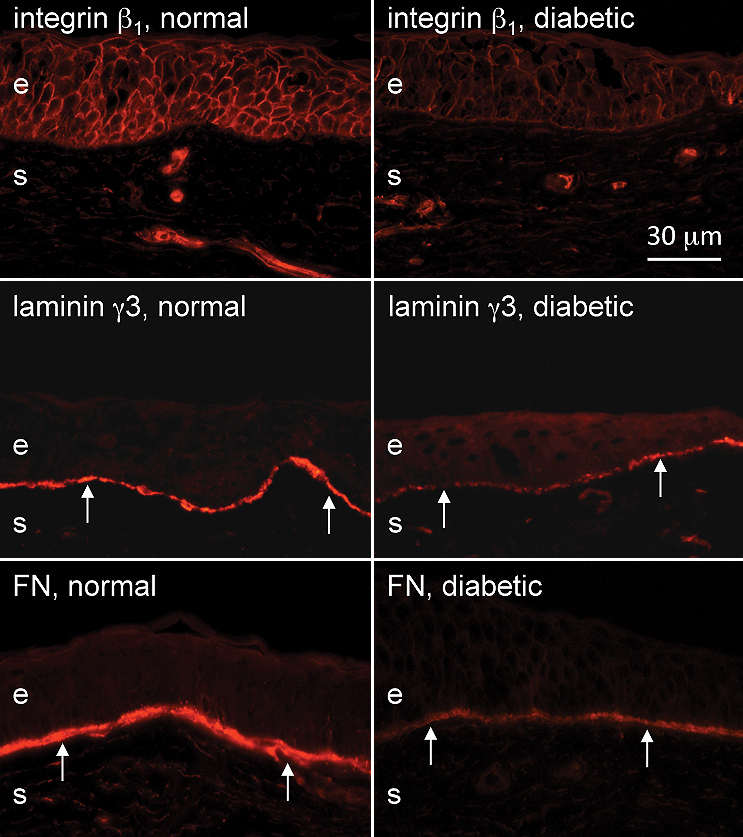
Integrin and basement membrane protein expression patterns in normal and diabetic ex vivo limbus. Integrin β1 staining is markedly reduced in the diabetic limbus, which occurs in all epithelial layers. A limbal-specific laminin γ3 chain staining is weak and discontinuous in the diabetic limbal epithelial basement membrane (arrows). This is also true for fibronectin. e, epithelium, s, stroma. Bar=30 μm.
Normalization of putative stem cell marker patterns upon c-met overexpression
We have recently shown that the expression of certain markers altered in the ex vivo diabetic corneas including integrin α3β1, some laminin chains, nidogen-1, nidogen-2, and phosphorylated p38 MAP kinase (p-p38) returned to almost normal patterns in the central region of diabetic organ-cultured corneas after c-met overexpression using rAV-driven c-met transduction [30]. As shown in Figure 5, c-met transduction brought about an increase in staining for some of these markers, such as integrin α3β1 and p-p38, so that they became closer to normal (see [30]) in the diabetic limbus. We next examined if altered expressions of putative LESC markers could also be normalized by rAV-cmet transduction of diabetic organ-cultured corneas. Indeed, c-met overexpression was accompanied by increased limbal staining for K15 (did not reach significance), K17, and K19, as well as ΔNp63α isoform compared to vector-transduced corneas (Figure 6; compare with Figure 1 and Figure 2), so that the staining became similar to normal corneas. Some of these increases were significant (Figure 7), although certain markers did not show an appreciable change in staining intensity. It should be mentioned that in organ-cultured diabetic corneas some K17 and K19 immunostaining could also be found in the suprabasal layers of the limbal epithelium as well as in the central cornea, whereas K15 was still expressed exclusively in the limbus.
Figure 5.
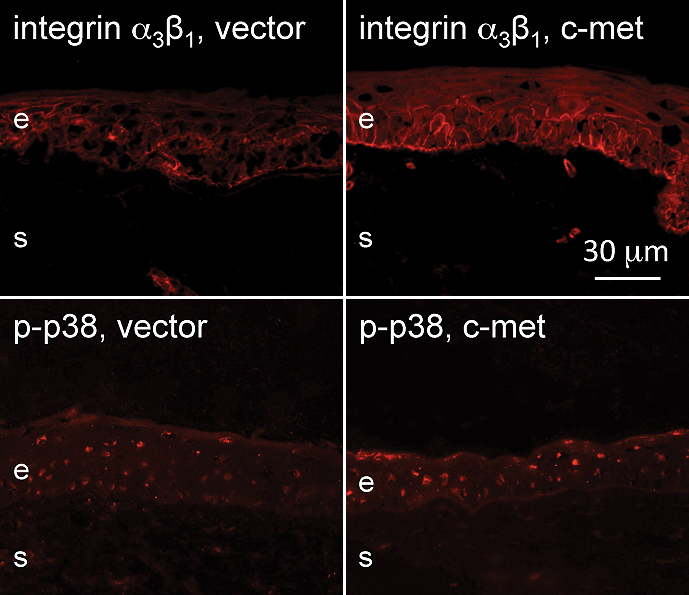
Increased diabetic marker expression in the diabetic limbus in organ culture upon c-met overexpression. Both integrin α3β1 and p-p38 staining in the limbal epithelium is increased upon c-met gene transduction and becomes similar to normal. e, epithelium, s, stroma. Bar=30 μm.
Figure 6.
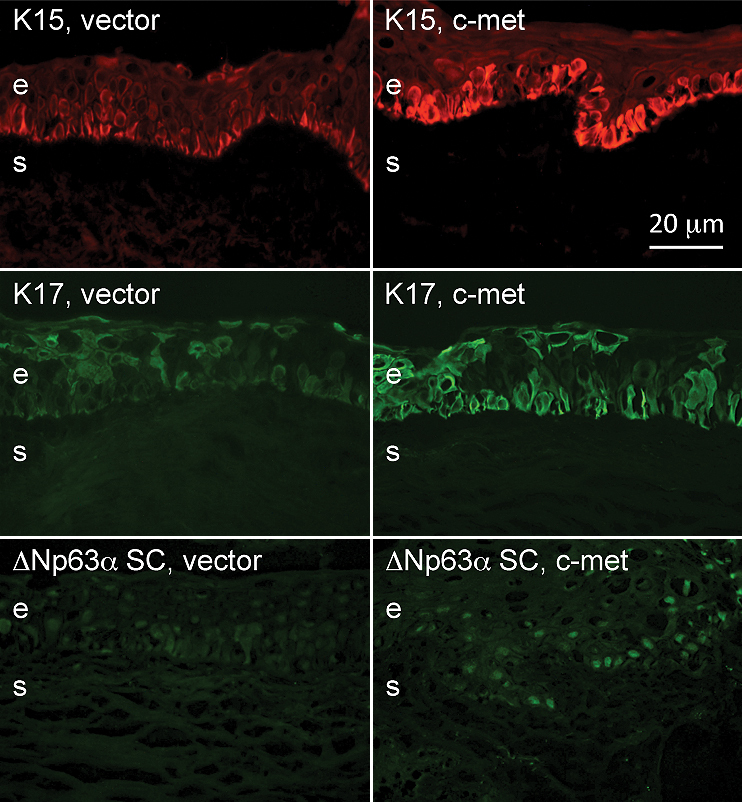
Increased putative LESC marker expression in the diabetic limbus in organ culture upon c-met overexpression. c-Met gene transduction leads to elevated expression of K15, K17, and ΔNp63α in the limbus of organ-cultured diabetic corneas. The staining intensity and regularity appear more normal (compare with Figure 1 and Figure 2). Note that in organ cultures keratins (especially K17) can also be seen in suprabasal epithelial layers. e, epithelium, s, stroma. Bar=20 μm.
Figure 7.

Statistical analysis of changes in the staining for various markers in the diabetic limbus in organ culture upon c-met overexpression. Significant staining increase after c-met gene transduction was observed for K17, K19, and ΔNp63α. Changes in the expression levels of K15, N-cadherin and β1 integrin did not reach significance. Data are mean±SEM. Thirteen pairs of c-met or vector treated organ cultured diabetic corneas were used. *p<0.05. Details are in the Methods section.
Discussion
As a systemic disease, diabetes has significant impact on all tissues. In the eye, the major vision-threatening effect is on retina (DR), whereas the other ocular parts are generally thought to be less affected [42,43]. However, more than a half of diabetics suffer from corneal problems related to neuropathy and epitheliopathy [44]. Various epithelial abnormalities are present in diabetic corneas that appear to be related to cell adhesion and basement membrane alterations, decreased innervation, and poor wound healing [44-46]. Using adenoviral gene therapy with overexpression of c-met that is downregulated in diabetic corneas [47], we were able to bring basement membrane protein patterns and wound healing times in organ-cultured diabetic corneas close to normal [30]. Other possible ways to normalize these corneas could be a restoration of normal signaling of the epidermal growth factor receptor (EGFR) axis that is downregulated by high glucose and diabetes [48,49], silencing of specific proteinases [31] or a blockade of opioid growth factor - opioid growth factor receptor system with naltrexone [50].
Mechanisms responsible for the epithelial changes in diabetic corneas are still not well understood. One possibility is that epithelial alterations could be triggered or exacerbated by abnormal innervation, which may be the case with ulcers [51]. However, several lines of evidence support direct detrimental action of hyperglycemia in diabetes on corneal epithelium. In diabetic rabbits, corneal wound healing is not delayed [52], although corneal neuropathy develops [53]. In human corneal organ culture where corneas are denervated, delayed diabetic epithelial wound healing persists [30]. In normal organ-cultured porcine corneas, high glucose causes delayed epithelial wound healing [48]. Because LESC and their immediate progeny (TA cells) play a key role in the epithelial maintenance and renewal, these data support the hypothesis that LESC and or/TA cells may be altered in the course of diabetes. Using a large panel of antibodies to putative LESC/limbal basal epithelial markers we tested this hypothesis using ex vivo human corneas, as well as organ-cultured diabetic corneas upon viral-induced c-met overexpression.
In the ex vivo corneas, many tested markers including keratins 15, 17 and 19, as well as Bcrp1/ABCG2, ΔNp63α isoform, N-cadherin, laminin γ3 chain, and β1 integrin were significantly downregulated in diabetic compared to normal limbus. In some cases, such as with integrin β1 or laminin γ3 chain, the immunostaining intensity was diminished. In other cases, such as with K17, ABCG2, or N-cadherin, the number of positive cells was markedly reduced in the diabetic limbus. This is the first demonstration of changes in LESC marker expression in a common disease that does not involve LESC deficiency. It may be suggested that the observed differences in marker expression between normal and diabetic corneal limbus relate to functional abnormalities of stem cell niche in diabetes. At present one can only speculate on what kind of dysfunction such reduced marker expression would be related. A plausible candidate would be impaired cell migration translating into slower and incomplete wound healing in diabetic corneas. The data support the idea that stem cell niche alterations may underlie poor wound healing and other epithelial abnormalities typical for diabetic corneas. It would also be important to understand whether diabetes changes marker expression in LESC, TA cells or both. The generally even distribution of most studied putative LESC markers in the limbal basal cells (comprised by LESC and TA cells) would favor the hypothesis that reduced expression of these markers in diabetic corneas is applicable to both LESC and TA cells.
It was interesting to examine whether gene therapy that can bring diabetic corneas closer to normal in terms of specific protein expression and wound healing rates would also change the expression of putative LESC markers in diabetic corneas toward normal patterns. To this end, we used organ-cultured diabetic corneas following gene therapy with c-met, which had significantly improved epithelial wound healing and expression of basement membrane markers and signaling intermediates [30].
Compared to vector treatment, c-met treated corneas displayed enhanced staining for several putative LESC markers, which became similar to normal ex vivo limbus. These data attest to the feasibility of using specific gene therapy to normalize the functions of LESC in diabetic corneas, which may be useful for LESC transplantation in diabetics. However, not all the markers altered in diabetic ex vivo corneas showed increased staining upon c-met gene transduction (e.g., β1 integrin) suggesting that one-gene therapy was not enough for corneal normalization.
A partial effect of c-met upregulation on LESC marker expression could be related to the influence of this gene therapy only on certain cell signaling pathways. As we showed before [30], c-met overexpression in diabetic corneas causes normalization of epithelial wound healing by restoring signaling through p38. However, overexpression of proteinases cathepsin F and matrix metalloproteinase-10 (MMP-10) in diabetic corneas or incubation of normal corneas in high glucose appear to reduce migration-promoting EGFR signaling through Akt phosphorylation [31,48]. Our preliminary data showed that a combined gene therapy with c-met overexpression and shRNA silencing of cathepsin F and MMP-10 brought diabetic corneas significantly closer to normal in terms of epithelial protein expression, p38 and Akt phosphorylation, and wound healing time than c-met upregulation alone. Therefore, by a concerted acting on several key signaling pathways, this combination could possibly exert a greater positive effect on putative LESC marker expression in the diabetic limbus.
Based on limbal location, little or no expression in the central cornea, preferential expression in basal limbal epithelial cells, various putative LESC markers have been proposed, such as K8, K15, K17, K19, Bcrp1/ABCG2, ΔNp63α, N-cadherin, laminin γ3 chain, β1 integrin, TCF4, frizzled 7, SOD2, epiregulin, Notch-1, α-enolase, vimentin, C/EBPδ, SPON1, and nectin-3 [23,32,33,35,36,41,54-65]. However, despite numerous attempts, no single and reliable LESC marker has been identified so far. This is in part due to the fact that unambiguous identification of LESC has been difficult. These cells are generally considered as largely non-proliferating, or slow cycling. Based on this criterion many authors agree that if corneal cultures or animal corneas in vivo are labeled with tritiated thymidine or bromodeoxyuridine and then chased for a while (at least several weeks), the few corneal cells that retain the label should be considered LESC. This promising strategy has been used to examine which markers are expressed by these cells. They were found to stain for K14, K15, CDH3 (P-cadherin), Wnt-4 [61], as well as to contain high levels of integrins β1 and β4 [60]. However, these markers are expressed not only in putative LESC but also in other limbal cells, as well as in central corneal cells (e.g., both integrins) and thus cannot be considered specific for LESC. Additional experiments with label-retaining cells using a large panel of antibodies are definitely needed to establish which existing markers are more specific for LESC. Currently, it is generally agreed that a combination of several markers should be used to characterize the presence of LESC in tissues and cultures.
The problem of LESC markers has gained wide attention because of recent success in transplantation of cultured limbal epithelium to patients with LESC deficiency [26,66-71]. Unfortunately, not all such cultures, especially when only small amounts of biopsied tissue were available for autologous transplantation, have been characterized as to the expression of putative LESC markers. Some authors, however, to standardize the cultures for successful transplantation, did examine one to several markers, e.g., p63 and K19 [69,70], confirming the presence of LESC-like cells in the transplanted cultures. In line with low content of LESC in corneal tissue, successful transplantations could be achieved when the fraction of p63-positive cells exceeded 3% [70].
In summary, we provide here the first account of significant alterations of limbal stem cell compartment in human diabetic corneas with respect to several commonly used putative LESC markers. These abnormalities may lead to diabetic LESC dysfunction and to clinically observed epithelial problems in diabetics including poor wound healing. Partial normalization of these pathological changes by c-met overexpression may offer a possibility of improving LESC function and general corneal health in diabetes by specific gene therapy. Another promising approach could be autologous transplantation of limbal epithelial cells to diabetic patients with advanced disease after prior normalization of their marker expression levels by gene therapy during culture.
Acknowledgments
The authors thank Prof. Luciano Zardi (Sirius biotech S.r.l., Genoa, Italy) for a kind gift of BC-2 antibody to human tenascin-C. Presented in part at the Annual Meeting of the Association for Research in Vision and Ophthalmology (ARVO), Fort Lauderdale, FL, May, 2010. This work was supported by NIH R01 EY13431, M01 RR00425 and Winnick Family Foundation, Eye Defects Research Foundation, Skirball Program in Molecular Ophthalmology.
References
- 1.Herse PR. A review of manifestations of diabetes mellitus in the anterior eye and cornea. Am J Optom Physiol Opt. 1988;65:224–30. doi: 10.1097/00006324-198803000-00013. [DOI] [PubMed] [Google Scholar]
- 2.Cavallerano J. Ocular manifestations of diabetes mellitus. Optom Clin. 1992;2:93–116. [PubMed] [Google Scholar]
- 3.Saini JS, Khandalavla B. Corneal epithelial fragility in diabetes mellitus. Can J Ophthalmol. 1995;30:142–6. [PubMed] [Google Scholar]
- 4.Sánchez-Thorin JC. The cornea in diabetes mellitus. Int Ophthalmol Clin. 1998;38:19–36. [PubMed] [Google Scholar]
- 5.Malik RA, Kallinikos P, Abbott CA, van Schie CH, Morgan P, Efron N, Boulton AJ. Corneal confocal microscopy: a non-invasive surrogate of nerve fibre damage and repair in diabetic patients. Diabetologia. 2003;46:683–8. doi: 10.1007/s00125-003-1086-8. [DOI] [PubMed] [Google Scholar]
- 6.Negi A, Vernon SA. An overview of the eye in diabetes. J R Soc Med. 2003;96:266–72. doi: 10.1258/jrsm.96.6.266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Saito J, Enoki M, Hara M, Morishige N, Chikama T, Nishida T. Correlation of corneal sensation, but not of basal or reflex tear secretion, with the stage of diabetic retinopathy. Cornea. 2003;22:15–8. doi: 10.1097/00003226-200301000-00004. [DOI] [PubMed] [Google Scholar]
- 8.Gekka M, Miyata K, Nagai Y, Nemoto S, Sameshima T, Tanabe T, Maruoka S, Nakahara M, Kato S, Amano S. Corneal epithelial barrier function in diabetic patients. Cornea. 2004;23:35–7. doi: 10.1097/00003226-200401000-00006. [DOI] [PubMed] [Google Scholar]
- 9.Wylegała E, Moćko L, Woyna-Orlewicz A, Teper S, Orzhechowska-Wylegała B. Diabetic complications within ocular surface. Pol Merkur Lekarski. 2006;21:495–7. [PubMed] [Google Scholar]
- 10.Ben Osman N, Jeddi A, Sebai L, Zghal I, Kaoueche M, Gaigi S, Ayed S. The cornea of diabetics. J Fr Ophtalmol. 1995;18:120–3. [PubMed] [Google Scholar]
- 11.Inoue K, Kato S, Ohara C, Numaga J, Amano S, Oshika T. Ocular and systemic factors relevant to diabetic keratoepitheliopathy. Cornea. 2001;20:798–801. doi: 10.1097/00003226-200111000-00004. [DOI] [PubMed] [Google Scholar]
- 12.Schermer A, Galvin S, Sun TT. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J Cell Biol. 1986;103:49–62. doi: 10.1083/jcb.103.1.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cotsarelis G, Cheng SZ, Dong G, Sun TT, Lavker RM. Existence of slow-cycling limbal epithelial basal cells that can be preferentially stimulated to proliferate: implications on epithelial stem cells. Cell. 1989;57:201–9. doi: 10.1016/0092-8674(89)90958-6. [DOI] [PubMed] [Google Scholar]
- 14.Tseng SC. Concept and application of limbal stem cells. Eye (Lond) 1989;3:141–57. doi: 10.1038/eye.1989.22. [DOI] [PubMed] [Google Scholar]
- 15.Lauweryns B, van den Oord JJ, Missotten L. The transitional zone between limbus and peripheral cornea. An immunohistochemical study. Invest Ophthalmol Vis Sci. 1993;34:1991–9. [PubMed] [Google Scholar]
- 16.Majo F, Rochat A, Nicolas M, Jaoudé GA, Barrandon Y. Oligopotent stem cells are distributed throughout the mammalian ocular surface. Nature. 2008;456:250–4. doi: 10.1038/nature07406. [DOI] [PubMed] [Google Scholar]
- 17.Secker GA, Daniels JT. Corneal epithelial stem cells: deficiency and regulation. Stem Cell Rev. 2008;4:159–68. doi: 10.1007/s12015-008-9029-x. [DOI] [PubMed] [Google Scholar]
- 18.Sun TT, Tseng SC, Lavker RM. Location of corneal epithelial stem cells. Nature. 2010;463:E10–1. doi: 10.1038/nature08805. [DOI] [PubMed] [Google Scholar]
- 19.Castro-Muñozledo F, Gómez-Flores E. Challenges to the study of asymmetric cell division in corneal and limbal epithelia. Exp Eye Res. 2011;92:4–9. doi: 10.1016/j.exer.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 20.Pellegrini G, Rama P, De Luca M. Vision from the right stem. Trends Mol Med. 2010;17:1–7. doi: 10.1016/j.molmed.2010.10.003. [DOI] [PubMed] [Google Scholar]
- 21.Di Girolamo N. Stem cells of the human cornea. Br Med Bull. 2011 doi: 10.1093/bmb/ldr026. [DOI] [PubMed] [Google Scholar]
- 22.Dua HS, Shanmuganathan VA, Powell-Richards AO, Tighe PJ, Joseph A. Limbal epithelial crypts: a novel anatomical structure and a putative limbal stem cell niche. Br J Ophthalmol. 2005;89:529–32. doi: 10.1136/bjo.2004.049742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Di Iorio E, Barbaro V, Ruzza A, Ponzin D, Pellegrini G, De Luca M. Isoforms of ΔNp63 and the migration of ocular limbal cells in human corneal regeneration. Proc Natl Acad Sci USA. 2005;102:9523–8. doi: 10.1073/pnas.0503437102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dua HS, Saini JS, Azuara-Blanco A, Gupta P. Limbal stem cell deficiency: concept, aetiology, clinical presentation, diagnosis and management. Indian J Ophthalmol. 2000;48:83–92. [PubMed] [Google Scholar]
- 25.Hatch KM, Dana R. The structure and function of the limbal stem cell and the disease states associated with limbal stem cell deficiency. Int Ophthalmol Clin. 2009;49:43–52. doi: 10.1097/IIO.0b013e3181924e54. [DOI] [PubMed] [Google Scholar]
- 26.Nakamura T, Kinoshita S. New hopes and strategies for the treatment of severe ocular surface disease. Curr Opin Ophthalmol. 2011;22:274–8. doi: 10.1097/ICU.0b013e3283477d4d. [DOI] [PubMed] [Google Scholar]
- 27.Thoft RA, Friend J. The X, Y, Z hypothesis of corneal epithelial maintenance. Invest Ophthalmol Vis Sci. 1983;24:1442–3. [PubMed] [Google Scholar]
- 28.Wiley L, SundarRaj N, Sun TT, Thoft RA. Regional heterogeneity in human corneal and limbal epithelia: an immunohistochemical evaluation. Invest Ophthalmol Vis Sci. 1991;32:594–602. [PubMed] [Google Scholar]
- 29.Lehrer MS, Sun TT, Lavker RM. Strategies of epithelial repair: modulation of stem cell and transit amplifying cell proliferation. J Cell Sci. 1998;111:2867–75. doi: 10.1242/jcs.111.19.2867. [DOI] [PubMed] [Google Scholar]
- 30.Saghizadeh M, Kramerov AA, Yu FS, Castro MG, Ljubimov AV. Normalization of wound healing and diabetic markers in organ cultured human diabetic corneas by adenoviral delivery of c-met gene. Invest Ophthalmol Vis Sci. 2010;51:1970–80. doi: 10.1167/iovs.09-4569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Saghizadeh M, Kramerov AA, Yaghoobzadeh Y, Hu J, Ljubimova JY, Black KL, Castro MG, Ljubimov AV. Adenovirus-driven overexpression of proteinases in organ-cultured normal human corneas leads to diabetic-like changes. Brain Res Bull. 2010;81:262–72. doi: 10.1016/j.brainresbull.2009.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yoshida S, Shimmura S, Kawakita T, Miyashita H, Den S, Shimazaki J, Tsubota K. Cytokeratin 15 can be used to identify the limbal phenotype in normal and diseased ocular surfaces. Invest Ophthalmol Vis Sci. 2006;47:4780–6. doi: 10.1167/iovs.06-0574. [DOI] [PubMed] [Google Scholar]
- 33.Lyngholm M, Vorum H, Nielsen K, Østergaard M, Honoré B, Ehlers N. Differences in the protein expression in limbal versus central human corneal epithelium - a search for stem cell markers. Exp Eye Res. 2008;87:96–105. doi: 10.1016/j.exer.2008.05.001. [DOI] [PubMed] [Google Scholar]
- 34.Merjava S, Neuwirth A, Tanzerova M, Jirsova K. The spectrum of cytokeratins expressed in the adult human cornea, limbus and perilimbal conjunctiva. Histol Histopathol. 2011;26:323–31. doi: 10.14670/HH-26.323. [DOI] [PubMed] [Google Scholar]
- 35.Kawasaki S, Tanioka H, Yamasaki K, Connon CJ, Kinoshita S. Expression and tissue distribution of p63 isoforms in human ocular surface epithelia. Exp Eye Res. 2006;82:293–9. doi: 10.1016/j.exer.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 36.Kubota M, Shimmura S, Miyashita H, Kawashima M, Kawakita T, Tsubota K. The anti-oxidative role of ABCG2 in corneal epithelial cells. Invest Ophthalmol Vis Sci. 2010;51:5617–22. doi: 10.1167/iovs.10-5463. [DOI] [PubMed] [Google Scholar]
- 37.Echevarria TJ, Di Girolamo N. Tissue-regenerating, vision-restoring corneal epithelial stem cells. Stem Cell Rev. 2011;7:256–68. doi: 10.1007/s12015-010-9199-1. [DOI] [PubMed] [Google Scholar]
- 38.Hayashi R, Yamato M, Sugiyama H, Sumide T, Yang J, Okano T, Tano Y, Nishida K. N-cadherin is expressed by putative stem/progenitor cells and melanocytes in the human limbal epithelial stem cell niche. Stem Cells. 2007;25:289–96. doi: 10.1634/stemcells.2006-0167. [DOI] [PubMed] [Google Scholar]
- 39.Chen Z, de Paiva CS, Luo L, Kretzer FL, Pflugfelder SC, Li DQ. Characterization of putative stem cell phenotype in human limbal epithelia. Stem Cells. 2004;22:355–66. doi: 10.1634/stemcells.22-3-355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kabosova A, Azar DT, Bannikov GA, Campbell KP, Durbeej M, Ghohestani RF, Jones JC, Kenney MC, Koch M, Ninomiya Y, Patton BL, Paulsson M, Sado Y, Sage EH, Sasaki T, Sorokin LM, Steiner-Champliaud MF, Sun TT, Sundarraj N, Timpl R, Virtanen I, Ljubimov AV. Compositional differences between infant and adult human corneal basement membranes. Invest Ophthalmol Vis Sci. 2007;48:4989–99. doi: 10.1167/iovs.07-0654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schlötzer-Schrehardt U, Dietrich T, Saito K, Sorokin L, Sasaki T, Paulsson M, Kruse FE. Characterization of extracellular matrix components in the limbal epithelial stem cell compartment. Exp Eye Res. 2007;85:845–60. doi: 10.1016/j.exer.2007.08.020. [DOI] [PubMed] [Google Scholar]
- 42.Khan ZA, Farhangkhoee H, Chakrabarti S. Towards newer molecular targets for chronic diabetic complications. Curr Vasc Pharmacol. 2006;4:45–57. doi: 10.2174/157016106775203081. [DOI] [PubMed] [Google Scholar]
- 43.Jackson GR, Barber AJ. Visual dysfunction associated with diabetic retinopathy. Curr Diab Rep. 2010;10:380–4. doi: 10.1007/s11892-010-0132-4. [DOI] [PubMed] [Google Scholar]
- 44.Abdelkader H, Patel DV, McGhee CNj, Alany RG. New therapeutic approaches in the treatment of diabetic keratopathy: a review. Clin Experiment Ophthalmol. 2011;39:259–70. doi: 10.1111/j.1442-9071.2010.02435.x. [DOI] [PubMed] [Google Scholar]
- 45.Messmer EM, Schmid-Tannwald C, Zapp D, Kampik A. In vivo confocal microscopy of corneal small fiber damage in diabetes mellitus. Graefes Arch Clin Exp Ophthalmol. 2010;248:1307–12. doi: 10.1007/s00417-010-1396-8. [DOI] [PubMed] [Google Scholar]
- 46.Ljubimov AV, Huang ZS, Huang GH, Burgeson RE, Gullberg D, Miner JH, Ninomiya Y, Sado Y, Kenney MC. Human corneal epithelial basement membrane and integrin alterations in diabetes and diabetic retinopathy. J Histochem Cytochem. 1998;46:1033–41. doi: 10.1177/002215549804600907. [DOI] [PubMed] [Google Scholar]
- 47.Saghizadeh M, Kramerov AA, Tajbakhsh J, Aoki AM, Wang C, Chai NN, Ljubimova JY, Sasaki T, Sosne G, Carlson MR, Nelson SF, Ljubimov AV. Proteinase and growth factor alterations revealed by gene microarray analysis of human diabetic corneas. Invest Ophthalmol Vis Sci. 2005;46:3604–15. doi: 10.1167/iovs.04-1507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xu KP, Li Y, Ljubimov AV, Yu FS. High glucose suppresses epidermal growth factor receptor/phosphatidylinositol 3-kinase/Akt signaling pathway and attenuates corneal epithelial wound healing. Diabetes. 2009;58:1077–85. doi: 10.2337/db08-0997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Xu K, Yu FS. Impaired epithelial wound healing and EGFR signaling pathways in the corneas of diabetic rats. Invest Ophthalmol Vis Sci. 2011;52:3301–8. doi: 10.1167/iovs.10-5670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.McLaughlin PJ, Sassani JW, Klocek MS, Zagon IS. Diabetic keratopathy and treatment by modulation of the opioid growth factor (OGF)-OGF receptor (OGFr) axis with naltrexone: a review. Brain Res Bull. 2010;81:236–47. doi: 10.1016/j.brainresbull.2009.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lockwood A, Hope-Ross M, Chell P. Neurotrophic keratopathy and diabetes mellitus. Eye (Lond) 2006;20:837–9. doi: 10.1038/sj.eye.6702053. [DOI] [PubMed] [Google Scholar]
- 52.Zagon IS, Sassani JW, Carroll MA, McLaughlin PJ. Topical application of naltrexone facilitates reepithelialization of the cornea in diabetic rabbits. Brain Res Bull. 2010;81:248–55. doi: 10.1016/j.brainresbull.2009.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li J, Mai C, Hu Y. A study on histochemistry and electron microscopy of corneal nerves in experimental diabetic rabbits. Zhonghua Yan Ke Za Zhi. 1996;32:258–9. [PubMed] [Google Scholar]
- 54.Budak MT, Alpdogan OS, Zhou M, Lavker RM, Akinci MA, Wolosin JM. Ocular surface epithelia contain ABCG2-dependent side population cells exhibiting features associated with stem cells. J Cell Sci. 2005;118:1715–24. doi: 10.1242/jcs.02279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Barbaro V, Testa A, Di Iorio E, Mavilio F, Pellegrini G, De Luca M. C/EBPδ regulates cell cycle and self-renewal of human limbal stem cells. J Cell Biol. 2007;177:1037–49. doi: 10.1083/jcb.200703003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kusanagi R, Umemoto T, Yamato M, Matsuzaki Y, Nishida K, Kobayashi Y, Fukai F, Okano T. Nectin-3 expression is elevated in limbal epithelial side population cells with strongly expressed stem cell markers. Biochem Biophys Res Commun. 2009;389:274–8. doi: 10.1016/j.bbrc.2009.08.130. [DOI] [PubMed] [Google Scholar]
- 57.Mei H, Nakatsu MN, Ng M, Truong TT, Lee G, Deng SX. Differential expression of frizzled 7 in human corneal epithelial stem cells. ARVO Annual Meeting; 2011 May 1-May 5; Fort Lauderdale (FL). [Google Scholar]
- 58.Zhou M, Li XM, Lavker RM. Transcriptional profiling of enriched populations of stem cells versus transient amplifying cells. A comparison of limbal and corneal epithelial basal cells. J Biol Chem. 2006;281:19600–9. doi: 10.1074/jbc.M600777200. [DOI] [PubMed] [Google Scholar]
- 59.Bian F, Liu W, Yoon KC, Lu R, Zhou N, Ma P, Pflugfelder SC, Li DQ. Molecular signatures and biological pathway profiles of human corneal epithelial progenitor cells. Int J Biochem Cell Biol. 2010;42:1142–53. doi: 10.1016/j.biocel.2010.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pajoohesh-Ganji A, Pal-Ghosh S, Simmens SJ, Stepp MA. Integrins in slow-cycling corneal epithelial cells at the limbus in the mouse. Stem Cells. 2006;24:1075–86. doi: 10.1634/stemcells.2005-0382. [DOI] [PubMed] [Google Scholar]
- 61.Figueira EC, Di Girolamo N, Coroneo MT, Wakefield D. The phenotype of limbal epithelial stem cells. Invest Ophthalmol Vis Sci. 2007;48:144–56. doi: 10.1167/iovs.06-0346. [DOI] [PubMed] [Google Scholar]
- 62.Thomas PB, Liu YH, Zhuang FF, Selvam S, Song SW, Smith RE, Trousdale MD, Yiu SC. Identification of Notch-1 expression in the limbal basal epithelium. Mol Vis. 2007;13:337–44. [PMC free article] [PubMed] [Google Scholar]
- 63.Schlötzer-Schrehardt U, Kruse FE. Identification and characterization of limbal stem cells. Exp Eye Res. 2005;81:247–64. doi: 10.1016/j.exer.2005.02.016. [DOI] [PubMed] [Google Scholar]
- 64.Takács L, Tóth E, Losonczy G, Szanto A, Bähr-Ivacevic T, Benes V, Berta A, Vereb G. Differentially expressed genes associated with human limbal epithelial phenotypes: new molecules that potentially facilitate selection of stem cell-enriched populations. Invest Ophthalmol Vis Sci. 2011;52:1252–60. doi: 10.1167/iovs.10-5242. [DOI] [PubMed] [Google Scholar]
- 65.Merjava S, Brejchova K, Vernon A, Daniels JT, Jirsova K. Cytokeratin 8 is expressed in human corneoconjunctival epithelium, particularly in limbal epithelial cells. Invest Ophthalmol Vis Sci. 2011;52:787–94. doi: 10.1167/iovs.10-5489. [DOI] [PubMed] [Google Scholar]
- 66.Ahmad S, Kolli S, Lako M, Figueiredo F, Daniels JT. Stem cell therapies for ocular surface disease. Drug Discov Today. 2010;15:306–13. doi: 10.1016/j.drudis.2010.02.001. [DOI] [PubMed] [Google Scholar]
- 67.Kolli S, Lako M, Figueiredo F, Ahmad S. Corneal epithelial stem cells and their therapeutic application. In: Bharavand H, editor. Trends in Stem Cell Biology and Technology. Dordrecht (The Netherlands): Humana Press, a Part of Springer Science + Business Media, LLC; 2009. p. 319–65. [Google Scholar]
- 68.Burman S, Sangwan V. Cultivated limbal stem cell transplantation for ocular surface reconstruction. Clin Ophthalmol. 2008;2:489–502. [PMC free article] [PubMed] [Google Scholar]
- 69.Shortt AJ, Secker GA, Rajan MS, Meligonis G, Dart JK, Tuft SJ, Daniels JT. Ex vivo expansion and transplantation of limbal epithelial stem cells. Ophthalmology. 2008;115:1989–97. doi: 10.1016/j.ophtha.2008.04.039. [DOI] [PubMed] [Google Scholar]
- 70.Rama P, Matuska S, Paganoni G, Spinelli A, De Luca M, Pellegrini G. Limbal stem-cell therapy and long-term corneal regeneration. N Engl J Med. 2010;363:147–55. doi: 10.1056/NEJMoa0905955. [DOI] [PubMed] [Google Scholar]
- 71.Baylis O, Figueiredo F, Henein C, Lako M, Ahmad S. 13 years of cultured limbal epithelial cell therapy: a review of the outcomes. J Cell Biochem. 2011;112:993–1002. doi: 10.1002/jcb.23028. [DOI] [PubMed] [Google Scholar]
- 72.Ljubimov AV, Burgeson RE, Butkowski RJ, Michael AF, Sun TT, Kenney MC. Human corneal basement membrane heterogeneity: Topographical differences in the expression of type IV collagen and laminin isoforms. Lab Invest. 1995;72:461–73. [PubMed] [Google Scholar]
- 73.Guelstein VI, Tchypysheva TA, Ermilova VD, Litvinova LV, Troyanovsky SM, Bannikov GA. Monoclonal antibody mapping of keratins 8 and 17 and of vimentin in normal human mammary gland, benign tumors, dysplasias and breast cancer. Int J Cancer. 1988;42:147–53. doi: 10.1002/ijc.2910420202. [DOI] [PubMed] [Google Scholar]
- 74.Ljubimov AV, Afanasjeva AV, Litvinova LV, Senin VM. Basement membrane components produced by a mouse ascites teratocarcinoma TB24. Analysis with monoclonal and polyclonal antibodies. Exp Cell Res. 1986;165:530–40. doi: 10.1016/0014-4827(86)90605-1. [DOI] [PubMed] [Google Scholar]
- 75.Ljubimov AV, Bartek J, Couchman JR, Kapuller LL, Veselov VV, Kovarik J, Perevoshchikov AG, Krutovskikh VA. Distribution of individual components of basement membrane in human colon polyps and adenocarcinomas as revealed by monoclonal antibodies. Int J Cancer. 1992;50:562–6. doi: 10.1002/ijc.2910500412. [DOI] [PubMed] [Google Scholar]
- 76.Aisenbrey S, Zhang M, Bacher D, Yee J, Brunken WJ, Hunter DD. Retinal pigment epithelial cells synthesize laminins, including laminin 5, and adhere to them through α3- and α6-containing integrins. Invest Ophthalmol Vis Sci. 2006;47:5537–44. doi: 10.1167/iovs.05-1590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kohfeldt E, Sasaki T, Göhring W, Timpl R. Nidogen-2: a new basement membrane protein with diverse binding properties. J Mol Biol. 1998;282:99–109. doi: 10.1006/jmbi.1998.2004. [DOI] [PubMed] [Google Scholar]
- 78.Balza E, Siri A, Ponassi M, Caocci F, Linnala A, Virtanen I, Zardi L. Production and characterization of monoclonal antibodies specific for different epitopes of human tenascin. FEBS Lett. 1993;332:39–43. doi: 10.1016/0014-5793(93)80479-e. [DOI] [PubMed] [Google Scholar]