

Abstract
When encountering individuals with a potential inclination to harm them, people face a dilemma: Staring at them provides useful information about their intentions but may also be perceived by them as intrusive and challenging—thereby increasing the likelihood of the very threat the people fear. One solution to this dilemma would be an enhanced ability to efficiently encode such individuals—to be able to remember them without spending any additional direct attention on them. In two experiments, the authors primed self-protective concerns in perceivers and assessed visual attention and recognition memory for a variety of faces. Consistent with hypotheses, self-protective participants (relative to control participants) exhibited enhanced encoding efficiency (i.e., greater memory not predicated on any enhancement of visual attention) for Black and Arab male faces—groups stereotyped as being potentially dangerous—but not for female or White male faces. Results suggest that encoding efficiency depends on the functional relevance of the social information people encounter.
Keywords: encoding, memory, visual attention, threat, evolutionary psychology
If you spent an hour people watching in a large city, which of the many passersby would you later be able to identify? Common sense suggests that the faces you looked at longer would be better remembered. Sometimes, however, the very people most relevant to your immediate goals are the last ones you would want to stare at. For example, if you are new to the city and have just been reminded of the high crime rate by a story on the local news, you might become more vigilant toward strange males but nevertheless not want to assume the potential interpersonal costs of staring at them for too long. How might the cognitive system maintain the informational benefits of attending to others without paying the interactional costs of doing so? The present study suggests one solution to this dilemma—that people more efficiently encode individuals believed to threaten fundamental social goals such as self-protection.
Our hypotheses stem from two emerging and increasingly related literatures. The first is the work on automatically elicited changes in cognition and behavior that result from primed social goals (see Bargh & Williams, 2007). This literature illustrates, for instance, that exposing participants to stimuli that prime the goals of cooperating or mating engages motivational systems that shift cognitive processing and behavior toward the pursuit of these goals (Bargh, Gollwitzer, Lee-Chai, Barndollar, & Troetschel, 2001; Huang & Bargh, 2008). The second line of research builds on the premise that there are fundamental, recurring problems that humans have long faced and that evolved motivational systems manage these challenges by modulating how perceivers cognitively process information potentially relevant to these problems (Kenrick, 1994; Neuberg, Kenrick, & Schaller, in press). Physical aggression by others is one such problem, and one would thus expect social perceivers with activated self-protection goals to become especially vigilant toward potential sources of threat in the environment.
Which social targets are most likely to be seen as relevant to self-protection concerns? Historically, men are more likely to be aggressors and perpetrators of violent crime (Daly & Wilson, 1988), and out-group men are especially likely to be seen as posing a physical threat (e.g., Navarrete, McDonald, Molina, & Sidanius, in press; Sidanius & Pratto, 1999; Van Vugt, De Cremer, & Janssen, 2007). Although in-group men are also often seen as threatening, they may also be viewed as potential allies, especially when males of readily identifiable out-groups are also present. Hence, when individuals are concerned with self-protection—for example, when primed by media depictions of violent crime—one would expect them to become particularly vigilant toward out-group men. Of course, increasing overt visual attention (staring) can be costly: Direct gaze may be perceived as a challenge (Ellsworth, 1975), and indeed people often avert their gaze from ostensibly dangerous individuals (Rohner, 2004). To the extent perceivers could encode the features of faces presumed to be especially threatening into memory with special efficiency, they could obtain desired informational benefits without incurring the costs of overt staring. This suggests a threat-encoding superiority hypothesis—that perceivers with an active self-protection goal will encode potentially threatening individuals (in this case, out-group males) with special efficiency.
To test this hypothesis, we first primed participants to be in a self-protective goal state. Participants then viewed arrays of faces varying in their sex and in-group versus out-group status while a hidden remote eye tracker monitored overt visual attention toward each of the faces. Following this, participants were given a surprise recognition memory test.
The design employed here is an example of what cognitive psychologists have labeled an “ideal strategy manipulation” (Stone & Van Orden, 1993). By displaying each stimulus in exactly the same manner in both the control and self-protection conditions, any differences in encoding efficiency can be attributed only to the manipulation. Thus, even though strong processing differences may exist between face types (e.g., stronger memory for female faces relative to male faces because of the greater variability in hair styles and makeup in photographs of women or stronger memory by White perceivers for White faces than Black faces, owing to extremely robust out-group homogeneity processes; Anthony, Copper, & Mullen, 1992; Ostrom & Sedikides, 1992), any changes in encoding efficiency within face type that result from the priming manipulations strongly implicate strategic (although not necessarily conscious) modulations of information processing. Our interest in threat-facilitated encoding efficiency—the increase in memory “bang” for the visual attention “buck”—requires that we measure the effects of the motivation manipulation on both recognition memory and the amount of time participants spend looking at each of the faces. We thus analyze memory and attention separately, then combine these measures in an multilevel regression model (the first stage of a varying covariate ANCOVA) as a way of testing our efficiency hypothesis.
Experiment 1
Out-group men, in general, are likely to elicit vigilance. This is clearly the case for Black men in North America, who are stereotypically associated with the potential to do harm (e.g., Quillian & Pager, 2001). Thus, we might expect that when self-protection goals are active, non-Black perceivers’ memory for Black male faces would be enhanced relative to the poor memory such perceivers typically have for Black faces (i.e., the cross-race recognition deficit; Anthony et al., 1992; Ostrom & Sedikides, 1992). Indeed this motivated memory might even equal or surpass that for White male faces, which ordinarily generate better memory.
Method
Participants
Participants were 89 non-Black women (39) and men (50) recruited from the psychology participant pool at Arizona State University.1
Procedures
To minimize the possibility that participants would consciously try to control eye movements, they were told the study investigated visual perception using a portable electroencephalograph (actually an Applied Science Laboratories Series 5000 eye tracker).
After calibrating the eye tracking software, participants watched one of two brief film clips designed to activate motivational states but presented to the participants as the first of many stimuli to which they would be exposed as we measured changes in brain activity. Participants in the self-protection condition viewed scenes from The Silence of the Lambs depicting a female protagonist being stalked by a (White) serial killer. Participants in the control condition viewed scenes from Koyaanisqatsi depicting images of high-speed urban activity. Pretesting in prior research indicated these film clips produced the intended motivational states and elicited equivalent amounts of general arousal.2
After watching their assigned film, participants viewed a slideshow consisting of four different circular arrays (for 10 s each). Each array was composed of eight neutrally expressive faces reflecting the factorial combination of sex, race (Black, White), and physical attractiveness (highly attractive vs. average looking).3 The eye tracking software tracked participant eye movements during the slideshow and recorded the amount of time each participant spent looking at each face.4
Measures
Participants next completed an unexpected face recognition task consisting of 64 faces presented serially, in random order. This set included the 32 faces from the stimulus set and an additional 32 matched foils not seen before. The two sets were counterbalanced as stimuli for some participants versus memory foils for others. For each face, participants were asked to indicate whether or not they had previously seen the face.
Results
Overt visual attention
We first calculated the total time spent looking at each face for each participant and log transformed the measure to reduce the distribution’s skew. Some participants failed to look at some of the faces or had no observed dwell times because eye tracker capture was lost for a portion of their session, so we eliminated individuals with missing dwell times for 6 or more of the 32 faces. Anticipating the varying covariate ANCOVA that would be our final analysis, we conducted all analyses as multilevel models, in this case with the dwell times as the dependent variable at the first level and participants as the second level. This resulting split-plot design is formally equivalent to a repeated measures ANOVA with participant motivational condition (control vs. self-protection), gender of the face, and race (Black or White) of the face, as well as the factorial interactions among these terms, as fixed factors and participants (and the within-subjects effects) as a random factor.5 Note that the restricted maximum likelihood estimation procedure employed here automatically adjusts the df for each effect to more adequately estimate the p values, and we present df to one decimal to keep the reader aware of this adjustment.
Participants looked more at White faces than Black faces, F(1, 76.5) = 20.11, p < .001, and women more than men, F(1, 128.4) = 10.32, p = .002. These main effects were qualified by a significant face race × gender interaction, F(1, 96.3) = 7.2, p = .009, which was driven by the much lower gaze durations for Black male faces relative to all others. There was no main effect of the self-protection prime, but it did interact with face gender, F(1, 128.4) = 6.65, p = .011. Relative to control, self-protection increased the amount of overt visual attention to female faces and decreased it to male faces. No other effects were significant, all Fs < 1. Figure 1 displays the average gaze durations as a function of face type and motivation condition.
Figure 1.

Mean gaze duration (in seconds) as a function of face type and motivation condition (control vs. self-protection), Experiment 1
Recognition memory
Recognition accuracy was corrected for guessing by subtracting the participant’s false alarm rate (for a given type of face) from the memory decision for that face (1 = hit and 0 = miss), then linearly transforming (0.5x + 0.5) so that means would reflect proportions correctly recognized. The resulting memory scores were highly correlated (r = .93) with A-Prime (a nonparametric measure of recognition sensitivity) but had better distributional characteristics (specifically, A-prime scores clustered more at the extremes, inflating standard errors). These scores were used as the dependent variable in multilevel regression analyses that included participant as a random factor and participant motivational condition (control vs. self-protection), gender of the face, race of the face, and the factorial interactions among these fixed factors as predictors.
Participants remembered White faces better than Black faces, F(1, 63.5) = 5.53, p = .022, and they remembered women better than men, F(1, 79.2) = 15.17, p < .001; both of these main effects reflect the differences in overt visual attention paid to these face types. Also in accordance with the analysis of gaze duration, there was a marginally significant interaction of the race and gender of the face, F(1, 114.8) = 2.69, p = .104, driven by lower memory for Black male faces relative to all other face types. Although there was no main effect of the self-protection prime and it did not interact with face gender (both Fs < 1), there was a significant interaction with face race, F(1, 63.5) = 4.12, p = .047; relative to control, self-protection increased recognition accuracy for Black faces whereas it decreased it for White faces. The three-way interaction was not significant (F < 1). Figure 2 shows the average recognition accuracy as a function of face type and motivation condition.
Figure 2.
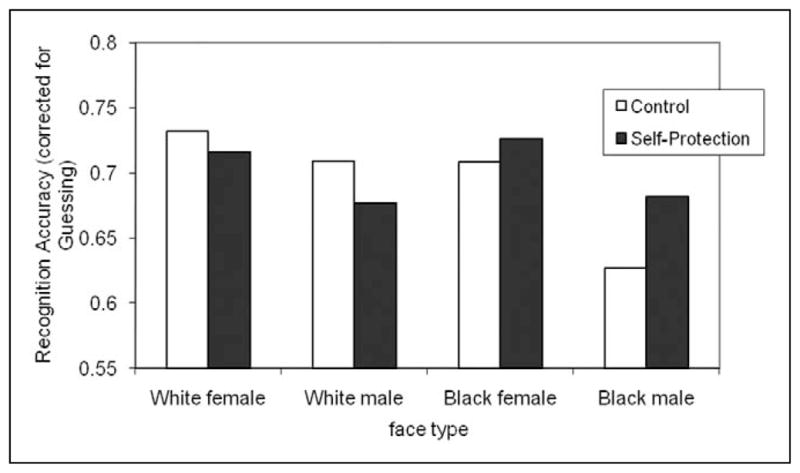
Mean recognition accuracy (corrected for guessing) as a function of face type and motivation condition (control vs. self-protection), Experiment 1
These memory results, considered in isolation, suggest that priming thoughts of self-protection leads to a general enhancement of memory for out-group faces. However, when we consider the attention results, in which self-protection led to an increase in attention to Black females but a decrease in attention to Black males, we see evidence of a disjunction of attention and memory for the Black males. This disjunction suggests that self-protective concerns evoke an encoding efficiency boost for Black males—enhanced memory despite diminished visual attention—relatively unique to them. The multilevel modeling framework provides a means of testing this encoding efficiency boost by employing gaze duration per face as a varying covariate in a model assessing recognition memory.
Memory controlling for gaze duration: Testing the hypothesis that threat enhances encoding efficiency
To ensure that these benefits truly reflected an enhanced efficiency of encoding and not merely an increase in looking time, gaze duration per face within each participant was included as a covariate. Gaze duration was grand-mean centered to allow inferences to be made across participants (essential because they were nested within motivation condition; see Kreft, DeLueew, & Aiken, 1995), and gaze duration slopes (within participants and face types) were specified as a random factor to allow for individual differences in the efficiency of encoding overtly attended items into memory. Across the following analyses, we included all higher order interactions of the fixed factors and the covariates (even when nonsignificant) to protect against model misspecification and to generate the best estimates of the means and slopes in the reported comparisons.6
To test whether self-protection motivation led participants to more efficiently encode out-group males, we assessed the simple effect of recognition accuracy for these faces covarying out differences in gaze duration. This analysis revealed that self-protection significantly boosted memory for Black males (t328.2 = 1.98, p = .047). Importantly, this threat-facilitated increase in encoding efficiency was highly selective: Activating a self-protective motivation did not enhance memory for Black or White females (both |t|s < 1), despite the fact that these faces were looked at longer in the self-protection condition, revealing that any memory advantage for these faces, albeit nonsignificant, was mediated by longer gaze durations. Moreover, there was no encoding efficiency boost for White males (t < 1). These motivation effects (i.e., changes in encoding efficiency for self-protection motivation relative to control) are depicted in Figure 3.
Figure 3.
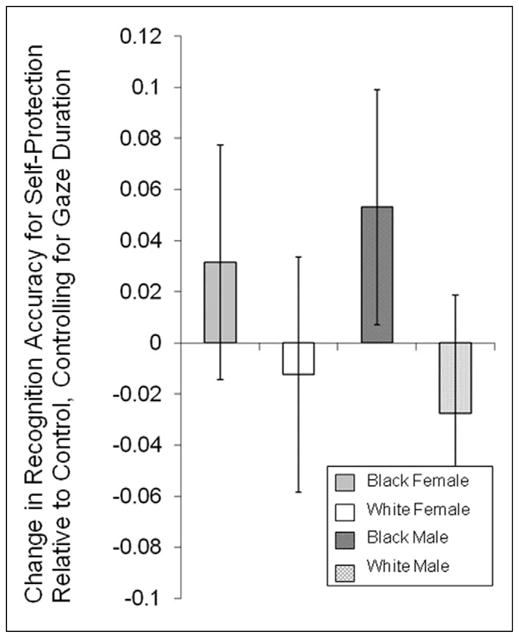
This figure depicts the priming effects in Experiment 1—the mean in the self-protection condition minus the mean in the control condition—as estimated by the multilevel regression model including gaze duration as a varying covariate
Note: The bars indicate 95% confidence intervals for the priming benefits (or costs) evaluated at the grand mean for gaze duration.
Experiment 2
Results of the first experiment suggest that priming self-protection concerns increases the efficiency of encoding for faces that have the greatest heuristic association with physical threat (out-group males). To explore the generality of these findings, we conducted a second experiment that differed in three primary ways from Experiment 1: (a) Rather than focusing on race to cue in-group versus out-group status, we presented participants with stimulus targets wearing Arabic versus Western clothing, (b) we manipulated self-protection concern not with the edited clip from The Silence of the Lambs but with a guided visualization exercise requiring participants to imagine themselves as a soldier under enemy fire in a foreign country, and (c) we employed an additional manipulation designed to explore the extent to which any encoding benefit might automatically accrue even when attention was directed away from the threatening faces; to do this, we instructed participants to study certain faces (indicated by a red border) while ignoring the others.
Method
Participants
Participants were 73 women (39) and men (34) recruited from the psychology participant pool at Arizona State University.
Procedures
Participants were told the study would examine how well they could focus their attention on certain faces but not others. Unlike in Experiment 1, participants here were told that we were monitoring where they looked to verify that they complied with the instructions.
To activate motivational states, participants listened to one of two 5-min audio clips, which they were told were designed to facilitate their performance in the primary task. In the self-protection condition, the audio narrative guided participants to visualize being a soldier on patrol in an unspecified foreign country. As the narration proceeded, participants imagined coming under attack by hostile forces, being cut off from the rest of their platoon, and being pinned down by enemy fire. Participants in the control condition instead were guided to visualize visiting a vibrant foreign market. Pretesting indicated that the visualization task (relative to control) significantly enhanced the participants’ concerns about self-protection.
After performing the guided visualization exercise, participants viewed a slide show consisting of four different circular arrays (for 8 s each). Each array was composed of eight neutrally expressive faces reflecting the factorial combination of sex, group (Arab or European, as implied by the presence or absence, respectively, of Arabic headgear), and whether the face was surrounded by a red border (directing the participant to study the face) or not (directing the participant to ignore the face).7 The eye tracking software tracked participant eye movements during the slideshow and recorded the amount of time each participant spent looking at each face.
Results
We initially followed the same analytical strategies as in Study 1, and the results broadly replicated the earlier study. To focus on the out-group male effect, however, we present here a simpler treatment in which Arab males are contrasted against the other face types. This serves to emphasize the most interesting effects from the full set of analyses. We include the means broken down by all four face types in Table 1 and the encoding boosts for each in Figure 4.
Table 1.
For Study 2, Mean Looking Time and Recognition Memory Accuracy as a Function of Whether the Face Was Attended or Ignored, the Gender and Race of the Face, and the Priming Condition
| Looking time (in seconds)
|
Memory accuracy
|
||||||
|---|---|---|---|---|---|---|---|
| Attention? | Face type | Control | Self-protection | Difference | Control | Self-protection | Difference |
| Ignored | European female | 0.28 | 0.23 | −0.06 | 0.53 | 0.51 | −0.021 |
| Ignored | Arabic female | 0.33 | 0.24 | −0.09 | 0.56 | 0.54 | −0.018 |
| Ignored | European male | 0.28 | 0.25 | −0.03 | 0.5 | 0.52 | 0.015 |
| Ignored | Arabic male | 0.24 | 0.24 | 0 | 0.45 | 0.51 | 0.059 |
| Attended | European female | 1.46 | 1.48 | 0.02 | 0.74 | 0.73 | −0.011 |
| Attended | Arabic female | 1.28 | 1.4 | 0.13 | 0.65 | 0.65 | 0.006 |
| Attended | European male | 1.37 | 1.62 | 0.24 | 0.77 | 0.76 | −0.011 |
| Attended | Arabic male | 1.32 | 1.38 | 0.06 | 0.65 | 0.68 | 0.025 |
Note: The priming benefit or cost is in the column labeled “Difference.”
Figure 4.
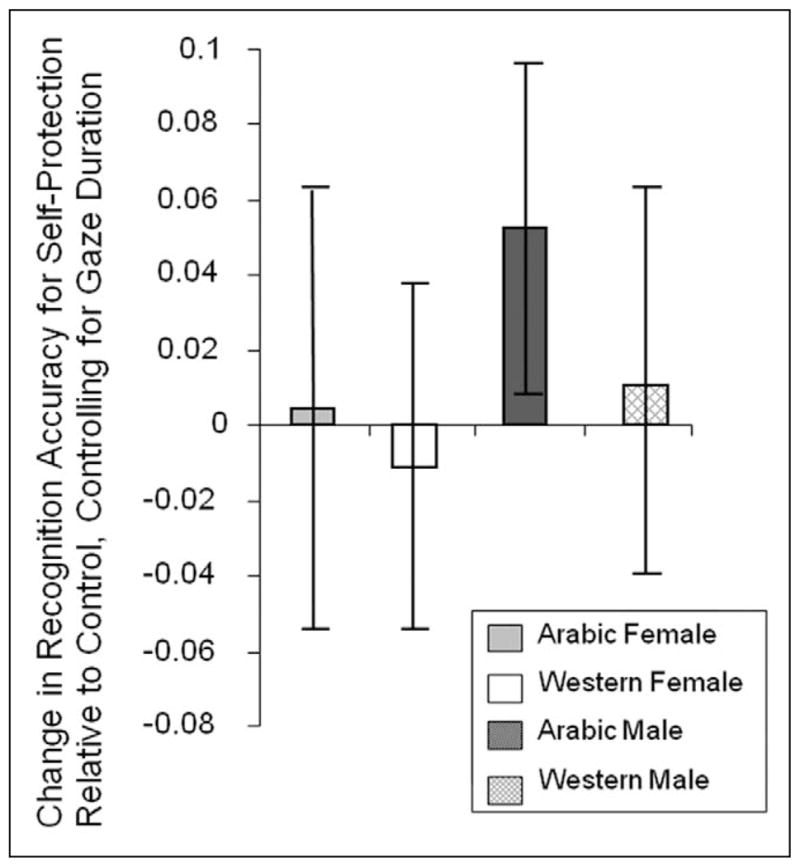
This figure depicts the priming effects in Experiment 2—the mean in the self-protection condition minus the mean in the control condition—as estimated by the multilevel regression model including gaze duration as a varying covariate
Note: The bars indicate 95% confidence intervals for the priming benefits (or costs) evaluated at the grand mean for gaze duration.
Overt visual attention
Demonstrating the effectiveness of the directed attention manipulation, participants spent much more time looking at the faces they were instructed to attend to (M = 1.35 s) relative to those they were told to ignore (M = 0.28 s), F(1, 1145.5) = 758.0, p < .0001.8 There was also an effect of the contrast of Arab male versus other face types, F(1, 196.87) = 6.65, p = .011, with Arab males garnering lower attention than the other faces.
Recognition memory (corrected for guessing)
Not surprisingly, lower overall visual attention to the faces that were actively ignored led to near chance memory for these faces (.507) relative to others (.690), F(1, 180.3) = 157.9, p < .0001. Memory for Arab males was worse than memory for the other groups, but this effect was moderated by an interaction of this contrast and the self-protection manipulation, F(1, 180.3) = 4.90, p = .028; As predicted, there was a significant increase in memory for Arab males in the self-protection condition relative to the control condition but no significant change in aggregate memory for the other face types.
Memory controlling for gaze duration
As indicated above, the self-protection manipulation caused no significant increase in gaze duration for the Arab male faces that could account for its effect on memory, which replicates the key attention–memory disjunction observed in Experiment 1. Formal analyses including gaze duration as a covariate in the model predicting memory confirmed this, producing an interaction of the Arab male contrast and the self-protection manipulation, F(1, 204.1) = 4.02, p = .046, which, as we predicted, corresponded to a significant increase in memory for Arab males in the self-protection condition relative to the control condition, effectively erasing any memory deficit these faces showed in the control condition. No such memory enhancements occurred for any of the other face types (all Fs < 1; see Figure 4).
Discussion
Does the efficiency with which one can encode a stranger’s face depend on whether that person might pose a threat? The present results suggest that concerns about interpersonal violence boost the encoding efficiency of faces heuristically associated with intergroup violence. Specifically, priming a self-protective motivation with a threatening movie clip or guided visualization exercise boosted the efficiency with which out-group male faces were remembered: Although people under a threat motivation did not especially attend to threatening faces, these faces were the recipients of greater memory bang for the perceivers’ visual attention buck.
Several features of our findings are worth briefly expanding on. First, threat-enhanced encoding efficiency was specific to out-group male faces and was not applied to individuals typically viewed as relatively nonthreatening (i.e., in-group male faces or female faces from either the in-group or the out-group).9 Focused specificity of this sort is a hallmark of a functional system—of a system operating to solve a particular problem—and has been observed in other research from our labs (e.g., Maner at al., 2005; Schaller, Park, & Mueller, 2003; for a broader discussion of functional specificity, see Kenrick, Neuberg, Griskevicius, Becker, & Schaller, in press; Neuberg et al., in press).10
Second, in Experiment 1, the threat-facilitated encoding efficiency for Black males occurred even though the aggressor in the film clip was a White male. This suggests that threat-based encoding benefits resonate not merely with the specific details of the priming incident but also with underlying, longstanding self-protective heuristics. If the encoding boosts reflected only the situational details of the priming incident, then White male targets would have received the encoding efficiency benefit because the aggressor in the film clip was White. Our results suggest that activating self-protective concerns enhances the encoding of faces associated with the greatest assumed potential and inclination to do physical harm, even if such faces are unrelated to the incidents that aroused the self-protective concerns. These results are consistent with those obtained by Navarrete and colleagues (2009), who showed that extinction of conditioned fear responses was slowest for out-group (Black) male faces but relatively rapid for female faces and in-group male faces.
Third, these self-protection boosts happened against the backdrop of the poor memory typically expected for out-group male faces. For example, the boost for Black male faces effectively eliminated the cross-race recognition deficit typically observed for other-race faces (Anthony et al., 1992; Ostrom & Sedikides, 1992). Specifically, in the control condition Black male faces elicited the worst memory of any face type, but once self-protection was aroused memory for the Black males increased whereas memory for White males decreased, such that memory was equivalent across male faces. This trade-off is consistent with a functional, strategic reallocation of limited cognitive resources from the less threatening faces to the more threatening faces.11
Finally, it bears reiterating that the encoding benefit for Black males was found in the absence of additional visual attention to the potentially threatening targets. Thus, any encoding benefits were not mediated by changes in overt visual attention; activating the fundamental goal of self-protection apparently caused central and covert processes to generate more memory bang for the equivalent visual attentional buck. This phenomenon therefore suggests an adaptive modulation of encoding, sensitive to the need to gain informational benefits without running the interactional risk of staring too long at a potentially dangerous face.
Conclusion
These results suggest that memory processes can be modulated by motivational or emotional systems such that they more effectively serve fundamental goals such as self-protection. Goals and needs have long been known to modulate the initial stages of perception. Indeed, in the early days of cognitive psychology, Bruner (1957) showed that poor children perceived coins to be physically bigger than did affluent children, ushering in a movement known as the “New Look” in perception. We believe that all cognitive processes are on the threshold of receiving a similar New Look (see Kenrick et al., in press). Of course, this version of the New Look might more properly be labeled the “Old Look,” as it postulates that regularities in the ancestral past account for the present resonance between self-protective goals and out-group men. Such a functionalist or ecological position also takes as axiomatic that every stage of cognitive processing serves these fundamental social goals, from early perception and attentional processes all the way through to the schemas and associative networks that capture our social world. Memory researchers, in particular, are beginning to investigate how survival concerns differentially engage memory systems (e.g., the recent findings that words are better recalled to the extent they are more relevant to survival; Nairne, Thompson, & Pandeirada, 2007). Our findings suggest that encoding efficiency can depend on perceivers’ current motivational states and the functional relevance of the social information being processed. The presence of pronounced encoding efficiencies may reveal key insights into how the mind deals with the important opportunities and threats of social life.
Acknowledgments
We thank Brian Hofer for assistance in data collection.
Biographies
D. Vaughn Becker is an assistant professor at Arizona State University.
Uriah S. Anderson is a doctoral candidate in psychology at Arizona State University.
Steven L. Neuberg is a professor at Arizona State University.
Jon K. Maner is an associate professor at Florida State University.
Jenessa R. Shapiro is an assistant professor at the University of California, Los Angeles.
Joshua M. Ackerman is an assistant professor in the Sloan School of Management at M.I.T.
Mark Schaller is a professor at the University of British Columbia.
Douglas T. Kenrick is a professor at Arizona State University.
Footnotes
Includes 16 Hispanic, 10 Asian, and 2 Native Americans. Not included in the analysis were 4 Black participants and 5 of mixed race.
In several early (unpublished) studies using these primes in other experimental paradigms (e.g., functional projection; see Maner et al., 2005), we included manipulation checks that reliably indicated that The Silence of the Lambs film clip made participants feel both threat and its concomitant emotion, fear, whereas Koyaanisqatsi led only to general arousal.
This factor was included to keep stimulus features consistent across a series of studies using other methods but is collapsed across in the present analyses (including it as a factor did not change the results we report here).
Note that the design of these arrays and the eye tracker resolution allowed a good measure of how long the participants looked at each face but did not allow us to discriminate the locations fixated within a particular face with sufficient accuracy.
More traditional repeated measures analyses aggregating the data at the participant level produced equivalent results, as did treating the faces (rather than the participants) as random factors. We also included participant sex, but as this produced no significant interactions with the variables of interest, we collapsed across this factor in the analyses reported here.
It is important to note that the resulting statistical models are a special case of the first stage of an analysis of covariance, and they provide a test of whether treatment effects are mediated by the covariate (indicated by attenuation of treatment effects when the covariate is included in the model; see Maxwell & Delaney, 2004, p. 339). In the following analyses, significant motivation effects represent enhancements of memory that cannot be attributed to changes in looking time and are unmediated by gaze duration in the sense that the enhancements become larger when the covariate is included in the model.
These faces also showed slight variations in facial affect, but this factor was evenly counterbalanced across the other factors and did not interact with any of the effects reported here and so is collapsed across.
Note that because of the layout of the arrays, participants had to saccade past the faces that were to be ignored, and so average looking times per unattended face were not expected to be zero.
This is not to say that in-group males are never viewed as threatening physical safety; clearly, they are. In an intergroup context, however—recall that participants viewed Black males along with White males or Arab males along with non-Arab males—a coalitional psychology is especially likely to come to the fore and bias threat-relevant processing toward out-group males.
The specific pattern of these findings also makes alternative interpretations of our findings less compelling. For instance, one might argue that our film or guided visualization manipulations created general arousal, or simple negative affect, and that these effects might account for our findings. We mentioned above that the control film elicits an equivalent amount of general arousal, rendering that alternative explanation less plausible. Even so, there are no compelling arguments of which we are aware for why general arousal, in and of itself, would direct attention and memory in the very specific way observed here—such that enhanced encoding efficiency would be observed only for out-group male targets. Simple negative affect would have a similar difficulty: If it merely directed processing toward groups seen as generally negative, the sex of out-group targets should make little difference; male and female Arabs, for instance, should be processed similarly to one another as negatively viewed out-groups. But they are not. Moreover, simple negative affect would have great difficulty explaining why out-group men would be attended to less than other groups. In all, just as the specificity of these findings strongly suggests that they represent a functional response to a particular threat-induced dilemma, they also serve to rule out simple alternative explanations.
Note the importance of differentiating the benefits that resulted from the self-protective priming manipulation from overall memory effects. The priming benefits stemmed from an experimental manipulation and index a purely psychological phenomenon, whereas the relatively better memory for female as compared to male faces observed in Experiment 1 (and similar effects in Experiment 2) is confounded with stimulus-based differences that may not illuminate anything about the psychology of the perceiver.
Reprints and permission: sagepub.com/journalsPermissions.nav
Declaration of Conflicting Interests
The authors declared no potential conflicts of interests with respect to the authorship and/or publication of this article.
Financial Disclosure/Funding
The authors disclosed receipt of the following financial support for the research and/or authorship of this article: This research was supported by National Institute of Mental Health Grant MH64734 to Douglas Kenrick, Steven Neuberg, and Mark Schaller, U.S. Army Research Institute for the Behavioral and Social Sciences Grant W74V8H-05-K-0003 to Douglas T. Kenrick and Steven Neuberg, and National Science Foundation Grant BCS-0642873 to Vaughn Becker, Douglas Kenrick, and Steven Neuberg.
References
- Anthony T, Copper C, Mullen B. Cross-racial facial identification: A social cognitive integration. Personality and Social Psychology Bulletin. 1992;18:296–301. [Google Scholar]
- Bargh JA, Gollwitzer PM, Lee-Chai A, Barndollar K, Troetschel R. The automated will: Nonconscious activation and pursuit of behavioral goals. Journal of Personality and Social Psychology. 2001;81:1014–1027. [PMC free article] [PubMed] [Google Scholar]
- Bargh JA, Williams EL. The automaticity of social life. Current Directions in Psychological Science. 2006;15:1–4. doi: 10.1111/j.0963-7214.2006.00395.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruner JS. On perceptual readiness. Psychological Review. 1957;64:123–152. doi: 10.1037/h0043805. [DOI] [PubMed] [Google Scholar]
- Daly M, Wilson M. Homicide. New York: Aldine de Gruyter; 1988. [Google Scholar]
- Ellsworth PC. Direct gaze as a social stimulus: The example of aggression. In: Pliner P, Krames L, Alloway T, editors. Non-verbal communication of aggression. New York: Plenum; 1975. pp. 53–75. [Google Scholar]
- Huang JY, Bargh JA. Peak of desire: Activating the mating goal changes life stage preferences across living kinds. Psychological Science. 2008;19:573–578. doi: 10.1111/j.1467-9280.2008.02125.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenrick DT. Evolutionary social psychology: From sexual selection to social cognition. In: Zanna MP, editor. Advances in Experimental Social Psychology. Vol. 26. San Diego, CA: Academic Press; 1994. pp. 75–121. [Google Scholar]
- Kenrick DK, Neuberg SL, Griskevicius V, Becker DV, Schaller M. Goal-driven cognition and functional behavior: The fundamental motives framework. Current Directions in Psychological Science. doi: 10.1177/0963721409359281. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreft IGG, de Leeuw J, Aiken LS. The effect of different forms of centering in hierarchical linear models. Multivariate Behavioral Research. 1995;30:1–21. doi: 10.1207/s15327906mbr3001_1. [DOI] [PubMed] [Google Scholar]
- Maner JK, Kenrick DT, Becker DV, Robertson TE, Hofer B, Neuberg SL, et al. Functional projection: How fundamental social motives can bias interpersonal perception. Journal of Personality and Social Psychology. 2005;88:63–78. doi: 10.1037/0022-3514.88.1.63. [DOI] [PubMed] [Google Scholar]
- Maxwell SE, Delaney HD. Designing experiments and analyzing data: A model comparison perspective. 2. Mahwah, NJ: Lawrence Erlbaum; 2004. [Google Scholar]
- Nairne JS, Thompson SR, Pandeirada JNS. Adaptive memory: Survival processing enhances retention. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2007;33:263–273. doi: 10.1037/0278-7393.33.2.263. [DOI] [PubMed] [Google Scholar]
- Navarrete CD, McDonald M, Molina L, Sidanius J. Prejudice at the nexus of race and gender: An out-group male target hypothesis. Journal of Personality and Social Psychology. doi: 10.1037/a0017931. in press. [DOI] [PubMed] [Google Scholar]
- Navarrete CD, Olsson A, Ho A, Mendes W, Thomsen L, Sidanius J. Fear extinction to an outgroup face: The role of target gender. Psychological Science. 2009;20:155–158. doi: 10.1111/j.1467-9280.2009.02273.x. [DOI] [PubMed] [Google Scholar]
- Neuberg SL, Kenrick DT, Schaller M. Evolutionary social psychology. In: Fiske ST, Gilbert D, Lindzey G, editors. Handbook of social psychology. New York: John Wiley; in press. [Google Scholar]
- Ostrom TM, Sedikides C. Out-group homogeneity effects in natural and minimal groups. Psychological Bulletin. 1992;112:536–552. [Google Scholar]
- Quillian L, Pager D. Black neighbors, higher crime? The role of racial stereotypes in evaluations of neighborhood crime. American Journal of Sociology. 2001;107:717–767. [Google Scholar]
- Rohner J. Memory-based attentional biases: Anxiety is linked to threat avoidance. Cognition and Emotion. 2004;18:1027–1054. [Google Scholar]
- Schaller M, Park JH, Mueller A. Fear of the dark: Interactive effects of beliefs about danger and ambient darkness on ethnic stereotypes. Personality and Social Psychology Bulletin. 2003;29:637–649. doi: 10.1177/0146167203029005008. [DOI] [PubMed] [Google Scholar]
- Sidanius J, Pratto F. Social dominance: An intergroup theory of social hierarchy and oppression. New York: Cambridge University Press; 1999. [Google Scholar]
- Stone GO, Van Orden GC. Strategic control of processing in word recognition. Journal of Experimental Psychology: Human perception and Performance. 1993;19:744–774. doi: 10.1037//0096-1523.19.4.744. [DOI] [PubMed] [Google Scholar]
- Van Vugt M, De Cremer D, Janssen DP. Gender differences in cooperation and competition: The male-warrior hypothesis. Psychological Science. 2007;18:19–23. doi: 10.1111/j.1467-9280.2007.01842.x. [DOI] [PubMed] [Google Scholar]