

Abstract
Fundamental motives have direct implications for evolutionary fitness and orchestrate attention, memory, and social inference in functionally specific ways. Motivational states linked to self-protection and mating offer illustrative examples. When self-protective motives are aroused, people show enhanced attention to, and memory for, angry male strangers; they also perceive out-group members as especially dangerous. In contrast, when mating motives are aroused, men show enhanced attention to and memory for attractive members of the opposite sex; mating motives also lead men (but not women) to perceive sexual arousal in attractive members of the opposite sex. There are further functionally specific consequences for social behavior. For example, self-protective motives increase conformity among both men and women, whereas mating motives lead men (but not women) to engage in anticonformist behavior. Other motivational systems trigger different adaptive patterns of cognitive and behavioral responses. This body of research illustrates the highly specific consequences of fitness-relevant motivational states for cognition and behavior, and highlights the value of studying human motivation and cognition within an evolutionary framework.
Keywords: motivation, social cognition, evolutionary psychology, threat, mating
You walk into a crowded room. Who do you notice? Who do you later remember? Do you try to fit in, or do you instead try to stand out from others? The answers to these questions likely depend critically on your current motivational state. Emerging evidence shows that attention to, memory for, and behavioral responses toward other people differ—sometimes dramatically—depending on whether individual decision makers are concerned with personal safety, are interested in romance, or are motivated to attain some other adaptively significant goal. A growing body of research suggests that biologically significant motivational states inspire us to be functionally attuned to particular events in our social environments. Those motivational states can be considered “fundamental” in an evolutionary sense (Kenrick, Griskevicius, Neuberg, & Schaller, in press). Here, we review the fundamental-motives framework, examining some of the functionally specific effects of such motivational states on cognition and behavior. This approach provides a distinct alternative to traditional domain-general models of motivation, which have often adopted overly simple dimensional approaches that bypass much of the important action in the functional links between motivation and cognition.
The Fundamental Motives Framework
Motivational systems have been shaped by natural selection to produce behaviors that increase reproductive fitness. For any social animal, including Homo sapiens, reproduction involves much more than just sex. To reproduce successfully—to produce viable offspring and raise them to reproductive age—human beings need to achieve many subsidiary goals, including affiliation, self-protection, status, mate acquisition, mate retention, and child rearing. Some of these goals may appear superficially similar (e.g., finding mates, making friends, and caring for children are all associated with rewards and concomitant positive feelings). But they are qualitatively distinct: Successful attainment of each goal requires different, and sometimes opposing, cognitive and behavioral responses. An emerging literature at the intersection of evolutionary biology and cognitive science suggests that these goals—representing conceptually distinct domains of adaptive behavior—are managed by distinct motivational systems (Kenrick, Li, & Butner, 2003). Given the important implications these goals have had for reproductive fitness and human evolution, the underlying motives can be considered “fundamental.”
This domain-specific approach to human motivation is buttressed by a wealth of research on human and nonhuman animals. For example, birds use distinct, domain-specific neuropsychological systems for learning and remembering information about poisonous foods, about the song of their species, and about the spatial location of their food caches. Similarly, humans use distinct, domain-specific systems and neural architectures for learning and remembering words, faces, and nausea-inducing foods (Sherry & Schacter, 1987).
This fundamental-motives framework leads to more specific predictions than traditional approaches to human motivation— such as those that characterize motives simply according to approach versus avoidance behavior, or according to an association with positive versus negative affect (Griskevicius et al., 2009). These dichotomous classification schemes fail to capture the psychologically distinct nature of specific motivational states and cannot predict the highly specific ways in which fitness-relevant motivational states orchestrate social cognition and behavior. For example, both insults and scorpions arouse “negative” feelings, but their affective and motivational consequences are qualitatively distinct. In addition, either an insult or a scorpion could trigger approach or avoidance, depending on an individual’s preexisting motivational state or on the immediate context. Insults may elicit avoidance if you are afraid, approach if you’re angry. A scorpion may elicit avoidance if it’s sitting alone on a rock, but approach if it’s crawling across your sleeping daughter’s leg. And, of course, even when insults and scorpions elicit approach behavior, they do so for motivational reasons that are psychologically different from those that incline a person to approach a parent, a pal, or a potential mate (Kenrick & Shiota, 2008).
In contrast to these traditional dichotomies, the fundamental-motives framework draws on the logic of evolutionary biology to identify a set of functionally discrete motivational states, each of which has specific implications for reproductive fitness (Kenrick et al., in press). Each motivational state, when aroused, is expected to exert a specific set of consequences for human attention, perception, cognition, and behavior in relation to specific kinds of fitness-relevant stimuli in the social environment. We illustrate this approach by focusing on two paradigmatic fundamental motives: self-protection and mate acquisition.
Self-Protective Social Cognition
Social environments—crowded parties, campus malls, or city streets—are informationally complex, populated by people who vary along numerous dimensions (sex, size, demeanor, emotional expression, etc.) that may or may not have implications for the perceiver’s well-being. Yet perceptual resources are also limited, so they must be selectively allocated to a small subset of features on a small subset of people in any situation. From an evolutionary perspective, these precious resources should be allocated selectively to individuals bearing features that, historically, were likely to have had implications for survival or reproduction. Allocation of cognitive resources should be especially selective when self-protection goals are aroused.
A self-protection motive can be temporarily activated by the perception of specific features in other people (e.g., angry facial expressions) or by awareness of specific events (e.g., recent acts of violence). In some people, self-protection motives may be chronically accessible (e.g., because of a dispositional belief that the world is dangerous). Once activated, a self-protection motive leads to heightened attention to functionally specific categories—and specific features—of social situations.
For example, people for whom a self-protection goal is chronically active are especially likely to have their attention held by angry faces (Fox, Russo, Bowles, & Dutton, 2001). An angry expression indicates a potential adaptive problem, especially if the angry person has the inclination and the means to do harm. Consistent with this logic, people are generally able to identify an angry face within a fraction of a second, and they are especially quick and accurate when the angry expression is displayed by a man rather than a woman (Becker, Kenrick, Neuberg, Blackwell, & Smith, 2007).
Self-protection motives also have implications for memory. For example, there is a well-known recognition-memory bias whereby White perceivers are good at distinguishing among faces of racial in-group members but relatively poor at distinguishing among faces of out-group members. This out-group homogeneity effect in recognition memory disappears, however—and sometimes even reverses—when perceiving angry faces. In the perceptual context of angry male faces, White perceivers are sometimes actually more (not less) accurate when distinguishing between the faces of Black men than they are when distinguishing the faces of White men (Ackerman et al., 2006).
The arousal of a self-protection motive also activates stereotypes and prejudices. Although encounters with members of stereotyped groups automatically trigger stereotypes, the specific content of a stereotype differs depending on which fundamental motives are currently active. Being in the dark arouses self-protection motives. Consequently, when White people—especially those with a dispositional tendency toward self-protection—view Black men under conditions of ambient darkness, threat-connoting stereotypes (“criminal,” “untrustworthy”) are likely to be activated into working memory (Schaller, Park, & Faulkner, 2003). Importantly, no such stereotype-activation effect is found on equally negative stereotypic traits irrelevant to threat and self-protection (e.g., “lazy,” “ignorant”). This functionally specific pattern emerges also on explicit measures of intergroup prejudice: When in a dark room, Canadians are especially likely to evaluate Iraqis unfavorably compared to fellow Canadians; again, this bias is specific to threat-relevant traits (Schaller et al., 2003; see Fig. 1).
Fig. 1.
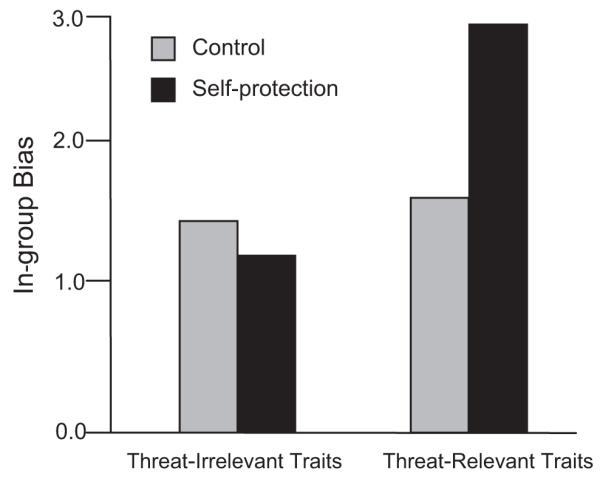
In-group bias on threat-relevant and threat-irrelevant traits, as a function of self-protective concerns triggered by ambient darkness. Canadians in a darkened room were especially likely to evaluate Iraqis less favorably than they evaluated fellow Canadians. This effect was specific to prejudicial perceptions on traits highly relevant to safety threats (“hostile,” “trustworthy”) but not to perceptions of equally evaluative but threat-irrelevant traits (“ignorant,” “open-minded”). Results based on Schaller et al. (2003).
Self-protection motives also exert functional biases on social inference. For example, people tend to “functionally project” specific adaptively relevant emotions onto another person’s objectively expressionless face (Maner et al., 2005). When a self-protection motive is aroused, people tend to erroneously perceive anger (but not other emotional states) in faces of ethnic out-group members (but not in-group members). This is not due to simple semantic priming, in which case one would expect people feeling fear to perceive fear in others—they don’t. Instead, fearful people selectively see anger, and they selectively see it in faces of individuals stereotypically expected to pose a threat.
Mate Acquisition and Social Cognition
Whereas self-protection motives direct attention to other people believed to pose dangers to physical safety, mating motives direct attention in qualitatively different ways. For instance, although self-protection motives generally have similar effects on both male and female perceivers, mating motives tend to have sex-specific effects (e.g., Griskevicius, Cialdini, & Kenrick, 2006). These sex-specific effects fit an extensive nomological network of findings on mating strategies, much of which fits with the biological principles of sexual selection and differential parental investment. Across a broad range of species, there is a positive relationship between parental investment and selectivity in mating: When there are sex differences in investment within a species, the sex investing less in offspring typically competes to mate with the more invested—and more selective—sex. (Females typically have higher investment in offspring, but in some species males care for offspring; in these high-male-investing species, males are more selective and females compete to mate with males.) In humans, the implication is that women (who have intrinsically high obligatory parental investment) are highly selective in their approach to mating; in contrast, men are motivationally inclined to be less selective, particularly with regard to short-term (i.e., low-involvement) mating opportunities (Li & Kenrick, 2006).
One implication is that, whereas women are likely to be attentive to qualities of men that cannot easily be assessed from mere physical appearance (such as relative social status compared to other local males), men are more likely to allocate perceptual and cognitive resources to superficial physical features connoting fertility—such as physical attractiveness. Consistent with this prediction, men (but not women) show enhanced encoding of and memory for the faces of attractive opposite-sex others (Becker, Kenrick, Guerin, & Maner, 2005). Moreover, among perceivers for whom short-term mating motives are chronically aroused, men (but not women) show enhanced visual attention to the faces of attractive opposite-sex others; this sex difference does not occur among individuals for whom mating motives are relatively inactive (Duncan et al., 2007).
Another implication is that, compared with women, men are more likely to take behavioral risks with potentially attractive mates—especially when mating motives are temporarily aroused. This sex differences shows up in a predictable misperception of others’ facial emotions. In comparison to the inferential consequences of self-protection motives (which lead both sexes to project anger onto faces of stereotypically threatening out-group members; described above), mating motives lead men (but not women) to erroneously perceive sexual arousal (but not other emotional states) in the faces of physically attractive (but not unattractive) members of the opposite sex (Maner et al., 2005).
Functionally Specific Effects on Downstream Social Behaviors
Just as fundamental motives influence initial attention, encoding, and retrieval of information, they also orchestrate subsequent social behavior in functionally sensible ways. Because successful self-protection and mate acquisition are influenced by a wide variety of behaviors (many of which are not overt acts of self-defense or sexual reproduction), fundamental motives can influence a remarkably diverse range of behavioral outcomes, including aggression, altruism, conformity, creativity, and economic decision making.
Consider, for example, the different effects of functionally distinct motivational states on conformity. Consistent with a safety-in-numbers strategy observed across many species, self-protection motives inspire both men and women to conform more to group pressure. In contrast, mating motives have dramatically different effects on male and female conformity, and these sex differences are exactly in line with evolved differences in mating strategies (Griskevicius, Goldstein, et al., 2006; see Fig. 2). Mating motives lead women to become more agreeable and conforming—a characteristic men find especially desirable in a relationship partner. In contrast, mating motives incline men toward anticonformity—a behavioral display that can signal dominance and help an individual man stand out from the crowd of other men competing for favor among highly selective females.
Fig. 2.
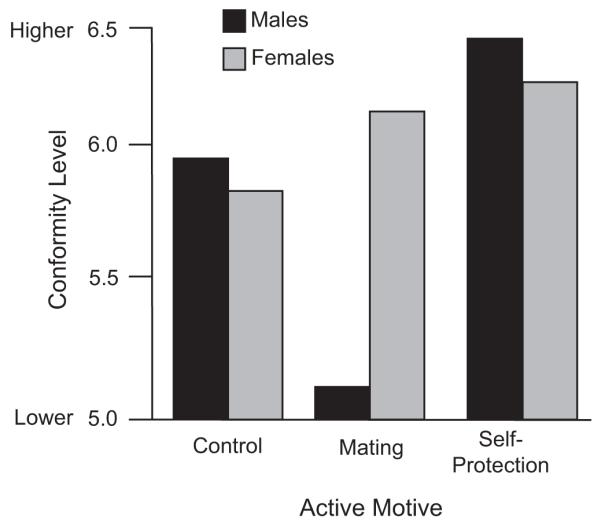
Conformity as a function of activated motive. Both men and women are more conforming when self-protection motives are active (compared to a control condition). In contrast, activating mating motives leads men to become less conforming and, as discussed in the text, more creative and showy in other ways (results based on Griskevicius, Goldstein, Mortensen, Cialdini, & Kenrick, 2006).
Mating motives also predictably increase a man’s willingness to spend money on specific kinds of consumer products. Whereas mating motives lead men to splurge on conspicuous luxuries such as cars and watches (purchases that can advertise wealth and status to selective females), the same mating motives incline men to be penny pinchers when it comes to inconspicuous items such as tissues and tile cleaner (Griskevicius et al., 2007).
Additional research reveals parallel consequences for creativity. Whether aroused by thoughts of short-term relationships (e.g., sexual flings) or long-term relationships (e.g., marriages), mating motives lead men to display greater levels of creativity and narrative flair—characteristics especially attractive to potential female mates. In contrast, women display higher levels of creativity only when motivated to pursue a long-term mate (Griskevicius, Cialdini, & Kenrick, 2006). This is consistent with the expectation that, because of their greater obligatory parental investment, women are more judicious in advertising their attractive qualities.
Discussion and Future Directions
When activated, fundamental motives dramatically alter how people think about and respond to social situations. We have focused on self-protection and mating motives, but these are just illustrative cases. A wealth of research on human evolution implies the existence of a broader set of fundamental motives designed to solve a variety of fitness-relevant problems that require specific kinds of behavioral solutions (e.g., avoiding infectious diseases, maintaining coalitional affiliations, rearing offspring). These motivational states cannot be reduced simply to approach or avoidance inclinations. Instead, each motivational state is conceptually distinct and has functionally specific consequences for perception, cognition, and behavior.
This burgeoning body of research builds theoretically interesting bridges between contemporary cognitive science, evolutionary biology, and psychological theories of emotion and motivation. This conceptual synergy provides a mechanism through which classic theories of historical interest (such as those offered by William James, William McDougall, and Abraham Maslow) may be revisited to make them more pertinent to contemporary research in the psychological sciences (e.g., see Kenrick et al., in press).
Fundamental motives have effects across a wide range of cognitive and behavioral domains, with important implications for economic decision making, intergroup conflict, and other phenomena of real-world significance (e.g., Ackerman & Kenrick, 2008; Kenrick et al., 2009; Schaller & Neuberg, 2008). Thus, an understanding of evolutionarily fundamental human motives (and their consequences) is not only of substantial theoretical interest but also of potentially profound practical importance.
Acknowledgments
Funding
The preparation of this article was facilitated by National Institute of Mental Health Grant MH064734 to Douglas Kenrick and Steven Neuberg; by National Science Foundation Grant BCS-0642873 to D. Vaughn Becker, Douglas Kenrick, and Steven Neuberg; and by a grant from the Social Sciences and Humanities Research Council of Canada to Mark Schaller.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the authorship and/or publication of this article.
References
- Ackerman JM, Kenrick DT. The costs of benefits: Help-refusals highlight key trade-offs of social life. Personality & Social Psychology Review. 2008;12:118–140. doi: 10.1177/1088868308315700. [DOI] [PubMed] [Google Scholar]
- Ackerman J, Shapiro JR, Neuberg SL, Kenrick DT, Schaller M, Becker DV, et al. They all look the same to me (unless they’re angry): From out-group homogeneity to out-group heterogeneity. Psychological Science. 2006;17:836–840. doi: 10.1111/j.1467-9280.2006.01790.x. [DOI] [PubMed] [Google Scholar]
- Becker DV, Kenrick DT, Guerin S, Maner JM. Concentrating on beauty: Sexual selection and sociospatial memory. Personality and Social Psychology Bulletin. 2005;12:1643–1652. doi: 10.1177/0146167205279583. [DOI] [PubMed] [Google Scholar]
- Becker DV, Kenrick DT, Neuberg SL, Blackwell KC, Smith DM. The confounded nature of angry men and happy women. Journal of Personality and Social Psychology. 2007;92:179–190. doi: 10.1037/0022-3514.92.2.179. [DOI] [PubMed] [Google Scholar]
- Duncan LA, Park JH, Faulkner J, Schaller M, Neuberg SL, Kenrick DT. Adaptive allocation of attention: Effects of sex and sociosexuality on visual attention to attractive opposite-sex faces. Evolution and Human Behavior. 2007;28:359–364. doi: 10.1016/j.evolhumbehav.2007.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox E, Russo R, Bowles R, Dutton K. Do threatening stimuli draw or hold visual attention in subclinical anxiety? Journal of Experimental Psychology: General. 2001;130:681–700. [PMC free article] [PubMed] [Google Scholar]
- Griskevicius V, Cialdini RB, Kenrick DT. Peacocks, Picasso, and parental investment: The effects of romantic motives on creativity. Journal of Personality and Social Psychology. 2006;91:63–76. doi: 10.1037/0022-3514.91.1.63. [DOI] [PubMed] [Google Scholar]
- Griskevicius V, Goldstein NJ, Mortensen CR, Cialdini RB, Kenrick DT. Going along versus going alone: When fundamental motives facilitate strategic (non)conformity. Journal of Personality and Social Psychology. 2006;91:281–294. doi: 10.1037/0022-3514.91.2.281. [DOI] [PubMed] [Google Scholar]
- Griskevicius V, Goldstein NJ, Mortensen CR, Sundie JM, Cialdini RB, Kenrick DT. Fear and loving in Las Vegas: Evolution, emotion, and persuasion. Journal of Marketing Research. 2009;46:384–395. doi: 10.1509/jmkr.46.3.384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griskevicius V, Tybur JM, Sundie JM, Cialdini RB, Miller GF, Kenrick DT. Blatant benevolence and conspicuous consumption: When romantic motives elicit strategic costly signals. Journal of Personality and Social Psychology. 2007;93:85–102. doi: 10.1037/0022-3514.93.1.85. [DOI] [PubMed] [Google Scholar]
- Kenrick DT, Griskevicius V, Neuberg SL, Schaller M. Renovating the pyramid of needs: Contemporary extensions built upon ancient foundations. Perspectives on Psychological Science. doi: 10.1177/1745691610369469. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenrick DT, Griskevicius V, Sundie JM, Li NP, Li YJ, Neuberg SL. Deep rationality: The evolutionary economics of decision-making. Social Cognition. 2009;27:764–778. doi: 10.1521/soco.2009.27.5.764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenrick DT, Li NP, Butner J. Dynamical evolutionary psychology: Individual decision-rules and emergent social norms. Psychological Review. 2003;110:3–28. doi: 10.1037/0033-295x.110.1.3. [DOI] [PubMed] [Google Scholar]
- Kenrick DT, Shiota MN. Approach and Avoidance Motivation(s): An Evolutionary Perspective. In: Elliot AJ, editor. Handbook of Approach and Avoidance Motivation. Psychology Press; New York: 2008. pp. 273–288. [Google Scholar]
- Li NP, Kenrick DT. Sex similarities and differences in preferences for short-term mates: What, whether, and why. Journal of Personality and Social Psychology. 2006;90:468–489. doi: 10.1037/0022-3514.90.3.468. [DOI] [PubMed] [Google Scholar]
- Maner JK, Kenrick DT, Becker DV, Robertson TE, Hofer B, Neuberg, et al. Functional projection: How fundamental social motives can bias interpersonal perception. Journal of Personality and Social Psychology. 2005;88:63–78. doi: 10.1037/0022-3514.88.1.63. [DOI] [PubMed] [Google Scholar]
- Schaller M, Neuberg SL. Intergroup prejudices and inter-group conflicts. In: Crawford C, Krebs DL, editors. Foundations of evolutionary psychology: Ideas, applications, and applications. Erlbaum; Mahwah, NJ: 2008. pp. 401–414. [Google Scholar]
- Schaller M, Park JH, Faulkner J. Prehistoric dangers and contemporary prejudices. European Review of Social Psychology. 2003;14:105–137. [Google Scholar]
- Sherry DF, Schacter DL. The evolution of multiple memory systems. Psychological Review. 1987;94:439–454. [Google Scholar]
Recommended Reading
- Kenrick DT, Delton AW, Robertson T, Becker DV, Neuberg SL. How the mind warps: A social evolutionary perspective on cognitive processing disjunctions. In: Forgas JP, Haselton MG, Hippel W, editors. The Evolution of the Social Mind: Evolution and Social Cognition. Psychology Press; New York: 2007. pp. 49–68. A chapter reviewing findings indicating that different motivational states produce theoretically meaningful discrepancies between attention, encoding, and memory.
- Kenrick, D.T., Griskevicius, V., Neuberg, S.L., & Schaller, M. (in press). (See References.) An article revisiting Abraham Maslow’s famous “pyramid of needs” approach to human motivation and presenting an updated hierarchy of fundamental human motives.
- Maner JK, DeWall CN, Baumeister RF, Schaller M. Does social exclusion motivate interpersonal reconnection? Resolving the ‘porcupine problem.’. Journal of Personality and Social Psychology. 2007;92:42–55. doi: 10.1037/0022-3514.92.1.42. An article describing how the arousal of an affiliation motive has functionally specific consequences for social perception and social behavior.
- Tooby J, Cosmides L. Conceptual foundations of evolutionary psychology. In: Buss DM, editor. Handbook of Evolutionary Psychology. John Wiley & Sons; Hoboken, NJ: 2005. pp. 5–67. A chapter providing an excellent overview of the assumptions underlying an evolutionary approach to human motivation and cognition.