

Abstract
Missense mutations in the erythroid band 3 protein (Anion Exchanger 1) have been associated with hereditary stomatocytosis. Features of cation leaky red cells combined with functional expression of the mutated protein led to the conclusion that the AE1 point mutations were responsible for Na+ and K+ leak through a conductive mechanism. A molecular mechanism explaining mutated AE1-linked stomatocytosis involves changes in AE1 transport properties that become leaky to Na+ and K+. However, another explanation suggests that point-mutated AE1 could regulate a cation leak through other transporters. This short paper intends to discuss these two alternatives.
1. Introduction
Band 3 or anion exchanger 1 (AE1) is the major red cell membrane protein. It belongs to the Solute Carrier 4A family (SLC4A) grouping bicarbonate transporters [1–3]. This protein catalyzes electroneutral chloride-bicarbonate exchange, and it is also expressed in kidney α-intercalated cells and in cardiomyocytes [4, 5]. In red cells, it is involved in two main tasks: enhancement of carbon dioxide transport and structuration of cell shape. It is found in red cells from all vertebrates except lampreys which naturally do not express erythrocyte AE1 [6]. Besides this exception, its complete absence from mammalian red cells leads to red cell defects whose consequences on health depend on the species. Dyserythropoiesis, severe haemolytic anaemia, and often premature death have been reported in mouse [7], and human [8], whereas cow or zebra fish seems to better withstand red cell AE1 deficiency [9, 10].
In human, many different mutations in SLC4A1 gene coding for AE1 have been reported [11]. Some of them are asymptomatic, whereas some others are associated with red cell pathologies characterized by alteration of red cell shape and rheological properties. As this protein is also expressed in kidney, a renal phenotype can be associated with SLC4A1 mutations. In this paper we will focus on red cell AE1, and the reader interested in kidney AE1 is therefore addressed to very exhaustive recent reviews on this subject [12–15].
When a red cell phenotype is associated with SLC4A1 mutations, the symptoms are hyperhaemolysis and anaemia, icterus, and splenomegaly. However, these symptoms may vary widely in intensity. It appears that the SLC4A1 mutations can be divided into two classes according to the way they impair AE1 function: (1) those that prevent correct folding of the protein so that it is not addressed to plasma membrane. This leads to a lower amount of AE1 in red cell membrane that impairs connection of skeleton and membrane, a feature of hereditary spherocytosis condition [16, 17]; (2) those that are associated with an increased cation permeability of red cell membrane. This latter condition is the hallmark of hereditary stomatocytosis [18, 19].
Since the initial discovery that 5 point mutations in SLC4A1 gene (responsible for L687P, D705Y, S731P, H734R, or R760Q substitutions in AE1) were associated with increased red cell Na+ and K+ leak [20], 4 other point mutations associated with similar red cell phenotype have been reported (G796R, E758K, S762R, and R730C) [21–24]. It has been proposed that the molecular mechanism accounting for cation leaky red cells in these hereditary stomatocytoses was a change in AE1 transport properties induced by the point mutations. The exchanger itself mediates cation leak by a conductive mechanism [25]. However, this interpretation leads to dramatically changing our way of thinking about band 3 transport mechanism. Moreover, the transport features of some of these AE1 mutants lead to another interpretation, that is, cation leak is due to the activation of endogenous Na+ and K+ transporters (or channels) in red cell membrane by mutated AE1 [23, 24, 26]. This short paper intends to discuss the molecular mechanism of red cell cation leak associated with AE1 point mutations in hereditary stomatocytosis.
1.1. Position of Amino Acid Substitutions in AE1 Polypeptide
AE1 polypeptide can be divided into 3 functional domains: a cytoplasmic amino-terminal domain, about 400 amino acids, interacting in red cells with various enzymes, haemoglobin, ankyrin, and band 4.2; a membrane spanning domain where anion exchange takes place and a short carboxy terminal end in the cytoplasm that associates with carbonic anhydrase II [27]. The protein forms part of a macrocomplex, combining membrane, and cytoplasmic proteins that are thought to improve the efficiency of gas transport by red cells [28].
Figure 1 illustrates the position of each of the point mutations that have been identified in patients with cation leaky red cells. The substitutions concern highly conserved amino acids among known electroneutral anion exchangers (SLC4A1, A2, and A3), and they are all located in the membrane spanning domain.
Figure 1.
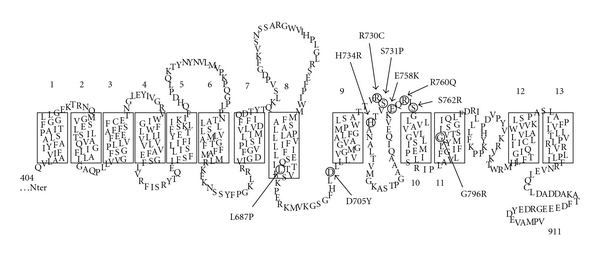
Topology of the membrane spanning domain of human AE1 with 13 α helices, according to Zhu et al. [29]. Aminoacids that are substituted in response to the different point mutations in AE1 gene are labelled by circles. Each of these mutations is associated with hereditary haemolytic anaemia characterized by cation leaky red cells.
1.2. Point-Mutated AE1 and Permeability Features
The permeability of red cells from patients bearing heterozygous mutation on AE1 has been investigated, and the transport features of the point-mutated AE1 have been studied by expression in amphibian oocytes. Thus, in most cases, it was possible to combine data from red cells to data from heterologous expression system. These data are presented here and will be discussed in the third part. Table 1 summarized the main features of cation leaky red cells and mutated AE1.
Table 1.
Features of cation leaky red cells and point-mutated AE1.
| AE1 point mutation | Red cell shape | Abundance of AE1 (band 3) in red cell membrane1 | Red cell cation leak rate at 0°C (multiple of normal)1 | Anion exchange activity of mutated AE12 | Pharmacology of the red cell cation leak | Pharmacology of the cation leak in heterologous expression system (cation conductance or cation flux) |
|---|---|---|---|---|---|---|
| L687P | Stomatocyte | 82% | 7-8 | Abolished | NS1652, SITS, dipyridamol | Cation conductance: SITS, Zn2+, La3+ sensitive |
| D705Y | Spherocyte | 77% | 8 | Abolished | NS1652, SITS, dipyridamol | Cation conductance: SITS, Zn2+, La3+ sensitive |
| R730C | Stomatocyte | Normal | 6 (at 37°C) | Abolished | NT | NT |
| S731P | Stomatocyte | 79% | 30–57-58–87 | Abolished | NS1652, SITS, dipyridamol | Cation conductance: SITS, Zn2+, La3+ sensitive |
| H734R | Stomatocyte | 74–82% | 87–94 | Abolished | NS1652, SITS, dipyridamol | Cation conductance: SITS, Zn2+, La3+ sensitive |
| E758K | Spherostomatocyte | Mild deficiency | NT | Normal with GPA coexpression | NT | Rb flux: DIDS, Zn2+, Gd3+ sensitive. Conductance: Zn2+, SITS and WW-781 sensitive |
| R760Q | Spherocyte | 85–92% | 4–6 | 74% of wt (with GPA) | NS1652, SITS, dipyridamol | NT |
| S762R | Stomatocyte | NT | 7 | Abolished | NT | NT |
| G796R | Stomatocyte | Normal | NT | Abolished | NT | Li uptake insensitive to SITS or H2DIDS |
This table summarized data collected from different publications. For L687P, D705Y, S731P, H734R, [20, 25]. For R730C, [23]. For E758K, [24]. For R760Q, [20, 31]. For S762R, [21]. For G796R, [22]. NT: not tested.
1Each number refers to features of red cells from different patients carrying the same AE1 mutation.
2The anion exchange was assessed in amphibian oocyte expressing the mutated AE1. The loss of anion exchange is confirmed in heterozygote red cells by ≈50% decreased anion flux.
The red cell leaks Na+ and K+ by an ouabain- and bumetanide-resistant mechanism that is temperature dependent; it is increased by temperatures below 37°C. This has been extensively studied in red cells of patients heterozygous for L687P, D705Y, S731P, H734R, R760Q, and S762R AE1 mutants [20, 21]. The diffusion of K+ and Na+ according to their electrochemical gradients leads to osmotic fragility of the red cells. At body temperature, the cation leak can be more or less compensated depending on the mutants. An artefactual rise in plasma K+ (pseudohyperkalaemia) can be observed after cooling blood to room temperature [30]. The shape of cation leak temperature dependence is not identical between the mutants [20], and the morphology of red cells also shows some differences between patients: blood smears exhibit stomatocytes or spherocytes. In red cells with H734R or G796R AE1 mutations, an increased activity of Na+-K+-2Cl− cotransporter, K+-Cl− cotransporter, Na+/H+ exchanger, or K+/Na+/H+ exchanger has been reported [22, 26]. R730C mutant is also associated with increased activity of Na+/H+ exchanger and Na+/K+-pump, whereas the Gardos channel fluxes are reduced [23]. Thus, the red cell permeability results from both the monovalent cation leak induced by AE1 point mutations and the activity of solute carriers that have been stimulated by the initial Na+ and K+ movements. The regulation of these other carriers could differ between patients as well as how the body cope with cation leaky red cells. This could explain variations in patient's phenotypes (Table 1).
For all the studied patients, the abundance of AE1 in red cells is grossly normal. However, the anion permeability of these cells is decreased suggesting a loss of anion exchange function of the protein [20]. Indeed, functional characterization of L687P, D705Y, R730C, S731P, H734R, S762R, and G796R mutants expressed in xenopus oocytes shows that they are no more able to exchange Cl− and HCO3 − [21, 25]. In contrast E758K and R760Q mutants keep an anion exchange activity [24, 31]. Another interesting difference for these two mutants is that their abundance in plasma membrane is highly dependent on glycophorin A (GPA) coexpression. GPA is known to bind AE1 and to act as a chaperone. Moreover this interaction stimulates AE1 transport activity [32–34].
Very few studies are available about conductance of these stomatocytic red cells. Only conductance of red cells from two patients with R730C or H734R mutation on AE1 has been reported. Patch current recordings on 3 red cells from a patient with R730C AE1 mutation do not allow to detect increased cation conductance compared to normal red cells [23]. Similar conclusions have been drawn from conductance analyses on red cells with H734R AE1 mutation [26].
Expression of L687P, D705Y, S731P, H734R, R760Q, S762R, or G796R AE1 mutants in xenopus oocytes induces a reversal in xenopus oocyte Na+ and K+ contents after 3 days in medium with ouabain- and bumetanide. This cation leak is associated with increased ouabain and bumetanide-resistant Rb+ or Li+ uptake which is similar to the red cell cation leak [20–22]. The Na+ and K+ transport associated with AE1 missense mutations shows independent movement of Na+ and K+ with a 1 for 1 stoichiometry. Moreover, when assessed, a conductance has been associated with these cation movements. Thus the molecular mechanism responsible for the observed cation movement is a channel-like transport mechanism [25].
Pharmacology of transport activities of the mutants have been assessed in red cells or in heterologous expression systems. DIDS (4,4′-diisothiocyanatostilbene-2,2′,-disulfonate) has long been known to inhibit anion exchange at micromolar concentrations [35, 36]. Moreover, sulfonate radical of DIDS can link two different lysines (K539 and K851) in putative transmembrane helices (TM) 5 and 12 of AE1 membrane spanning domain [36]. It is thus possible to covalently bind DIDS on each or both of these two lysines. The DIDS derivative SITS (4-acetamido-4′-isothiocyano-2,2′-stilbene disulfonate), flufenamic acid, and niflumic acid are also potent inhibitors of AE1 activity. It is observed that the point mutations impair the protein sensitivity to classical anion exchanger inhibitors. For instance, S731P mutation prevents DIDS covalent binding to the exchanger [20]. For the two mutants keeping anion exchange activity, the DIDS sensitivity of the transport is also impaired. Figure 2 illustrates Cl−/HCO3 − exchange as a function of DIDS concentrations for R760Q mutant compared to wt AE1. The R760Q mutation decreases AE1 DIDS sensitivity as shown by the right shift of the dose-response curve. The anion uptake mediated by E758K mutant is also less sensitive to DIDS than wtAE1 [24].
Figure 2.
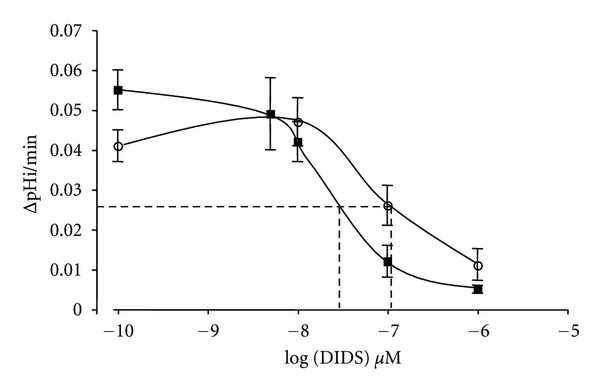
Dose response curves of Cl−/HCO3 − exchange by DIDS. The capacity of oocytes expressing wt AE1 or R760Q mutant (10 ng cRNA coinjected with 2.5 ng GPA-cRNA in both cases) to alkalinize in CO2/HCO3 − buffer without extracellular Cl− (gluconate medium) was assessed in presence of different DIDS concentrations. The initial slope of alkalinization as a function of time was plotted against DIDS concentrations. The method used was described in a previous publication [25]. Data are means +/− s.e.m. of 9 to 20 oocytes from different batches. Black symbols correspond to wt AE1 expressing oocytes, empty circles to R760Q mutant expressing oocytes.
Pharmacology of the cation leak induced by AE1 point mutations has also been investigated. Inhibition of the Na+ and K+ leak induced by L687P, D705Y, S731P, H734R, and R760Q mutations has been observed in red cells with SITS, dipyridamole, and NS1652 also known to block anion exchanger [20]. Pharmacology of the cation leak is difficult to assess in xenopus oocytes since endogenous cation permeabilities could be activated by common AE1 inhibitors such as DIDS or niflumic acid [37, 38]. However, inhibition of the Na+ and K+ conductance induced by expression of mutated AE1 was observed with SITS and plurivalent cations such as Zn2+, La3+, and Gd3+ [24, 25].
Amongst AE1 point mutations associated with cation leaky red cells, two mutants (R730C and E758K) exhibit peculiar transport features in amphibian oocytes. R730C AE1 mutant induces only a weak ouabain- and bumetanide-resistant cation leak in xenopus oocytes. It is not possible to measure a significant increase in ouabain- and bumetanide-resistant 86Rb+ uptake, and only a ≈2.5-fold increase in Li+ uptake is observed (to compare to ≈8-fold increase in oocytes expressing S731P mutant for instance). Moreover expression of R730C mutant is associated with an increased activity of the Na+/K+ ATPase [23]. For E758K, mutant permeability features depend on the expression system. It has been studied in two different amphibian oocytes, xenopus, and ambystoma. In both species, its abundance in plasma membrane is dependent on the coexpression of GPA. The mutant keeps anion exchange activity and induces a 86Rb+ uptake in both systems. However, it appears that this Rb permeability is not correlated to the expression level of the mutant when expressed in ambystoma: the higher number of transporters when coexpressed with GPA does not induce a higher 86Rb uptake. The expression of E758K mutant also slightly increases xenopus oocyte conductance, but this conductance does not account for the observed Rb+ permeability as deduced from differences in pharmacological pattern [24].
1.3. What Is the Pathway for Cations in Cells Expressing Point Mutated AE1?
Point mutations in AE1 are associated with monovalent cation leak in red cells as in heterologous expression systems. This monovalent cation leak could be correlated to a nonselective cation conductance and to elevated activity of endogenous transport systems.
Two possibilities that are not exclusive could be envisioned: the missense mutations in AE1 polypeptide change the transport properties of the protein that becomes leaky to Na+ and K+, or the mutated AE1 stimulates native transporters for Na+ and K+ in red cells as in heterologous expression systems.
The work on trout AE1 has shown that this protein could interact with Na+-K+-2Cl− cotransporter by its carboxy terminal domain, stimulating the activity of this transporter in xenopus oocytes [39]. Reports on E758K and R730C AE1 mutations suggest that the cation leak associated with these mutations likely involves activation of still undefined endogenous transporters.
As AE1 forms part of a macrocomplex, it functionally interacts with carbonic anhydrase [27, 40], and it is also involved in many molecular interactions in red cells, with ankyrin, glycophorin A (GPA), glycolytic enzymes, or haemoglobin, for instance [28, 41–43]. It is thus plausible that point-mutated AE1 could interfere with different endogenous transporters (understood as pumps, channels, or carriers) in red cells as in heterologous expression systems. It could be proposed that point mutations by changing AE1 conformation enable molecular interactions regulating the activity of various endogenous transporters. An AE1 mutated conformation could be envisioned which would not dramatically change AE1 transport features but would activate endogenous monovalent cation permeabilities in red cells as in heterologous expression systems. That should happen with different point mutations in AE1 membrane spanning domain.
Since pioneer work of electrophysiologists on red cells in the 80s [44], numerous cation and anion conductances have been characterized. Anion conductances (maxianion channels [45, 46]) or cation conductances such as nonselective Ca2+ permeable cation channels (L-type Ca2+ channel, voltage-dependent) or nonselective voltage-independent cation channels (NSVCCs) [47–51] and Ca2+ sensitive K+ channel (Gardos channel) [44] are well characterized in human red cells. It has been proposed that the TRPC6, member of the Transient Receptor Potential family proteins, contributes to the nonselective voltage-independent cation current in red cells [52]. However, the molecular identity of channels responsible for most of the electrophysiologically described conductances is unknown. It could be proposed that the nonselective Na+ and K+ leak induced by AE1 point mutations could be mediated by one of these conductances. However, the features of the cation leak associated with AE1 point mutations do not point out any of the red cell channels described so far. In particular, this cation leak is insensitive to amiloride, known to block the NSVCC; it is insensitive to extracellular Cl− concentration, known to stimulate red cell cation channels; it is insensitive to extracellular Ca2+ concentration [25]. Thus, in red cells as in heterologous expression system, the molecular identity of the transporters eventually activated by point-mutated AE1 still remains unknown as their activating mechanism.
Whatever the origin of the cation leak induced by AE1 point mutations, the consecutive alteration of cation permeability will impair red cell homeostasis and modulate the activity of different transporters. The permeability features of red cells with H734R, G796R, or R730C AE1 mutations show that the activity of different transporters could be stimulated: Na+-K+-2Cl− cotransporter, Na+/K+ ATPase, Na+(K+)/H+ exchanger, and K+-Cl+ cotransporter [22, 23, 26]. This could be due to the fact that point mutated AE1 induced a cation leak locally changing cation concentrations. These changes could be responsible for subsequent activation of various cation transporters. As a consequence, a same AE1 mutation could lead to diverse phenotypes in red cells depending on how the endogenous transporters react to the initial cation leak.
In the absence of identified native transporter mediating the cation leak associated with AE1 point mutations, the hypothesis of a cation leaky AE1 is challenging and the simplest to propose. Moreover, it provides an attractive approach to understand the transport mechanism of this protein. Whereas it is depicted as a typical electroneutral anion exchanger, the anion exchange rate through AE1 is extremely fast (10 000 per second) and Cl− slippage occurs occasionally (1 for 10 000 exchange). Crystallographic structures are not available yet with enough resolution to help understand the transport mechanism [53]. This mechanism should allow very rapid conformational changes that could resemble alternately opened gates for instance. Previous work on trout AE1 has shown that this exchanger can behave as an anion conductance permeable to organic solutes (taurine, sorbitol) and to monovalent cations (Na+ and K+) [54, 55]. The work on a truncated human AE1 has also shown that this protein could behave like a conductance when it was deleted of transmembrane segments 6 and 7 [56]. Thus, it appears feasible to convert the electroneutral anion exchanger into a conductive pathway by different manœuvres. Former studies in red cells, based on pharmacological evidence, have also suggested that a monovalent cation leak through AE1 could be induced by decreasing extracellular Cl− concentration [57]. The speculations about a monovalent cation leak through AE1 polypeptide have to combine the following considerations.
The point mutations induce a conformational change of the protein as suggested by changes in pharmacology sensitivity (Figure 2), by impairment of anion exchange capacity, or by requirement of GPA for correct addressing to plasma membrane. Moreover, diverse point mutations are likely to produce a similar AE1 conformation as deduced from similar permeability patterns. The transport features of cells expressing AE1 suggest at least three functional states for AE1: the wt state, a A mutated state where no anion exchange occurs only cation leak exists and a B mutated state where anion exchange and cation leak coexist (Figure 3). Whether or not these functional states are linked to different structural states of AE1 has to be shown but it is likely.
Figure 3.
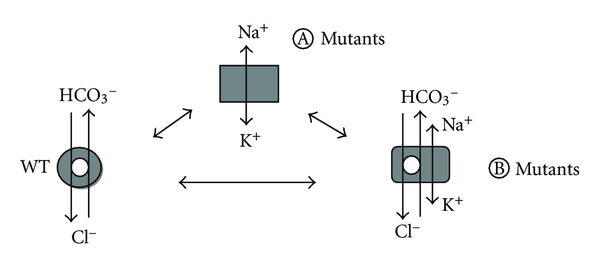
Putative transport states of AE1. Whereas the wt AE1 does only exchange Cl− and HCO3 −, a A mutated state only conducts monovalent cations and a B mutated state shows both transport activities, anion exchange and cation leak. The A state should be obtained with L687P, D705Y, R730C, S731P, H734R, S762R and G796R mutations. The B state should be obtained with E758K and R760Q mutations.
It appears that apparent unrelated positions in AE1 membrane spanning domain are susceptible to impair anion exchange function in the same way: point mutations L687P, D705Y, S731P, H734R, S762R, and G796R abolish anion exchange and induce a similar monovalent cation leak that is also similar to the cation leak observed in cells expressing the mutated AE1 responsible for South East Asian Ovalocytosis (SAO AE1) [21]. SAO AE1 is deleted of 9 amino acids at the junction of cytoplasmic domain and membrane spanning domain of the protein [58]. This mutation is prevalent in South East Asian population where it is proposed to protect against severe forms of malaria [59]. Thus, the cation leaky conformation appears as a complex organization involving distant segments in the membrane spanning domain of the protein, and this conformation could be obtained either by a deletion at the junction of cytoplasmic and transmembrane domain (SAO AE1) or by some specific point mutations. The cytoplasmic domain, per se, is probably not involved in the cation leaky conformation as suggested by the work on a trout AE1 deleted of cytoplasmic domain that kept its conductive transport mechanism [60].
In wtAE1, the anion exchange site involves transmembrane helix 8 (TM 8) and an anion selectivity filter including a region at the top of TM 12 and 13 and amino acids in the loop connecting TM 7 and 8 [61, 62]. Our recent work on the cation leaky H734R mutant has shown that the same TM 8 was also involved in cation movement suggesting a common pathway for anions and cations through AE1 [63]. Moreover, it was shown that amino acids in the intracellular loop connecting TM 8 and TM 9 play an important role in AE1 transport features. For instance, substitution of the charged residues in this loop induces a cation leak and severely impairs anion exchange activity. The position of the point mutations S731P, H734R, E758K, R760Q, and S762R at both extremities of the next loop connecting TM 9 and 10 also suggests an important functional role for this central part of the membrane spanning domain. Amino acid substitutions in these two loops could change the orientation, rotation, or movements of TM 8, 9, and 10 and impair AE1 transport site. The leak could be seen as a broken seal in the transport system that leaks Na+ and K+ for which a high driving force exists. This leads to consider that the transport site is susceptible to structural changes that could be induced by diverse but specific amino acid substitutions. This change unmasks a conductance for monovalent cations that does not seem to interfere with the ability to exchange anions since some mutants exhibit both transport activities.
The possibility for a carrier to function as a channel seems conflicting. Indeed channels are seen as structures that could simultaneously connect intra- and extracellular medium, what should never happen through a carrier. However, there are increasing examples of membrane proteins with ambiguous behaviour between channels and transporters. A historical example of transporter with channel activity is the glutamate transporter which is also a chloride channel [64–66]. Chloride channels, Na+-K+ pump, are other examples of ambiguous transport mechanisms between channels and transporters which strengthen our simple hypothesis of cation leaky AE1 [65–69]. In red cells, monovalent cation leak has also been associated with heterozygous mutations on RhAG (Rhesus Associated Glycoprotein) gene. Two different amino acid substitutions in RhAG could turn on a cation pore through this membrane protein proposed to be a NH3/NH4 + transporter in red cells [70].
2. Conclusion
Whereas specific AE1 mutations are undoubtedly linked to cation leaky red cells responsible for hereditary haemolytic anaemia, it is observed that all the 9 AE1 mutations presented here do not impair AE1 transport features in a similar manner. Moreover, membrane permeability of cells expressing point-mutated AE1 shows some differences suggesting a complicated regulation of this permeability. The proposition of point mutations altering AE1 transport mechanism is an attractive hypothesis supported by experimental evidence. However, this does not exclude the possibility for some mutated AE1 to also regulate the activity of other transporters.
Resolution of the 3D structure of AE1 would greatly help to understand the peculiar transport properties of this surprising protein. It would be of particular interest to know how the studied AE1 point mutations alter AE1 structure, if these different point mutations have a common mechanism of action on the structure. A better understanding of the mechanism of interactions between AE1 and its partners would also help to assign new regulatory function to AE1.
References
- 1.Alper SL. Molecular physiology of SLC4 anion exchangers. Experimental Physiology. 2006;91(1):153–161. doi: 10.1113/expphysiol.2005.031765. [DOI] [PubMed] [Google Scholar]
- 2.Pushkin A, Kurtz I. SLC4 base (HCO3-, CO3 2−) transporters: classification, function, structure, genetic diseases, and knockout models. American Journal of Physiology. 2006;290(3):F580–F599. doi: 10.1152/ajprenal.00252.2005. [DOI] [PubMed] [Google Scholar]
- 3.Romero MF. Molecular pathophysiology of SLC4 bicarbonate transporters. Current Opinion in Nephrology and Hypertension. 2005;14(5):495–501. doi: 10.1097/01.mnh.0000168333.01831.2c. [DOI] [PubMed] [Google Scholar]
- 4.Hamasaki N, Okubo K. Band 3 protein: physiology, function and structure. Cellular and Molecular Biology (Noisy-le-Grand, France) 1996;42(7):1025–1039. [PubMed] [Google Scholar]
- 5.Pucéat M, Korichneva I, Cassoly R, Vassort G. Identification of band 3-like proteins and Cl-/HCO3/- exchange in isolated cardiomyocytes. The Journal of Biological Chemistry. 1995;270(3):1315–1322. doi: 10.1074/jbc.270.3.1315. [DOI] [PubMed] [Google Scholar]
- 6.Hägerstrand H, Danieluk MX, Bobrowska-Hägerstrand MX, et al. Influence of band 3 protein absence and skeletal structures on amphiphile- and Ca2+-induced shape alterations in erythrocytes: a study with lamprey (Lampetra fluviatilis), trout (Onchorhynchus mykiss) and human erythrocytes. Biochimica et Biophysica Acta. 2000;1466(1-2):125–138. doi: 10.1016/s0005-2736(00)00184-x. [DOI] [PubMed] [Google Scholar]
- 7.Southgate CD, Chishti AH, Mitchell B, Yi SJ, Palek J. Targeted disruption of the murine erythroid band 3 gene results in spherocytosis and severe haemolytic anaemia despite a normal membrane skeleton. Nature Genetics. 1996;14(2):227–230. doi: 10.1038/ng1096-227. [DOI] [PubMed] [Google Scholar]
- 8.Ribeiro ML, Alloisio N, Almeida H, et al. Severe hereditary spherocytosis and distal renal tubular acidosis associated with the total absence of band 3. Blood. 2000;96(4):1602–1604. [PubMed] [Google Scholar]
- 9.Inaba M, Yawata A, Koshino I, et al. Defective anion transport and marked spherocytosis with membrane instability caused by hereditary total deficiency of red cell band 3 in cattle due to a nonsense mutation. The Journal of Clinical Investigation. 1996;97(8):1804–1817. doi: 10.1172/JCI118610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Paw BH, Davidson AJ, Zhou Y, et al. Cell-specific mitotic defect and dyserythropoiesis associated with erythroid band 3 deficiency. Nature Genetics. 2003;34(1):59–64. doi: 10.1038/ng1137. [DOI] [PubMed] [Google Scholar]
- 11.Jarolim P. Disorders of band 3. In: Ellory JC, Bernhardt I, editors. Red Cell Membrane Transport in Health and Disease. Berlin, Germany: Springer; 2003. pp. 603–619. [Google Scholar]
- 12.Alper SL. Diseases of mutations in the SLC4A1/AE1 polypeptide. In: Broer S, Wagner C, editors. Membrane Transport Diseases. Boston, Mass, USA: Kluwer Academic; 2003. pp. 39–63. [Google Scholar]
- 13.Alper SL. Familial renal tubular acidosis. Journal of Nephrology. 2010;23(supplement 16):S57–S76. [PubMed] [Google Scholar]
- 14.Bruce LJ, Tanner MJA. Erythroid band 3 variants and disease. Bailliere’s Best Practice and Research in Clinical Haematology. 1999;12(4):637–654. doi: 10.1053/beha.1999.0046. [DOI] [PubMed] [Google Scholar]
- 15.Toye AM. Defective kidney anion-exchanger I (AEI, Band 3) trafficking in dominant distal renal tubular acidosis (dRTA) Biochemical Society Symposium. 2005;72:47–63. doi: 10.1042/bss0720047. [DOI] [PubMed] [Google Scholar]
- 16.Iolascon A, Del Giudice EM, Perrotta S, Alloisio N, Morlé L, Delaunay J. Hereditary spherocytosis: from clinical to molecular defects. Haematologica. 1998;83(3):240–257. [PubMed] [Google Scholar]
- 17.Jarolim P, Murray JL, Rubin HL, et al. Characterization of 13 novel band 3 gene defects in hereditary spherocytosis with band 3 deficiency. Blood. 1996;88(11):4366–4374. [PubMed] [Google Scholar]
- 18.Flatt JF, Bruce LJ. The hereditary stomatocytoses. Haematologica. 2009;94(8):1039–1041. doi: 10.3324/haematol.2009.010041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Stewart GW. Hemolytic disease due to membrane ion channel disorders. Current Opinion in Hematology. 2004;11(4):244–250. doi: 10.1097/01.moh.0000132240.20671.33. [DOI] [PubMed] [Google Scholar]
- 20.Bruce LJ, Robinson HC, Guizouarn H, et al. Monovalent cation leaks in human red cells caused by single amino-acid substitutions in the transport domain of the band 3 chloride-bicarbonate exchanger, AE1. Nature Genetics. 2005;37(11):1258–1263. doi: 10.1038/ng1656. [DOI] [PubMed] [Google Scholar]
- 21.Guizouarn H, Borgese F, Gabillat N, et al. South-east Asian ovalocytosis and the cryohydrocytosis form of hereditary stomatocytosis show virtually indistinguishable cation permeability defects. British Journal of Haematology. 2011;152(5):655–664. doi: 10.1111/j.1365-2141.2010.08454.x. [DOI] [PubMed] [Google Scholar]
- 22.Iolascon A, De Falco L, Borgese F, et al. A novel erythroid anion exchange variant (Gly796Arg) of hereditary stomatocytosis associated with dyserythropoiesis. Haematologica. 2009;94(8):1049–1059. doi: 10.3324/haematol.2008.002873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stewart AK, Kedar PS, Shmukler BE, et al. Functional characterization and modified rescue of novel AE1 mutation R730C associated with overhydrated cation leak stomatocytosis. American Journal of Physiology. 2011;300(5):C1034–C1046. doi: 10.1152/ajpcell.00447.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stewart AK, Vandorpe DH, Heneghan JF, et al. The GPA-dependent, spherostomatocytosis mutant AE1 E758K induces GPA-independent, endogenous cation transport in amphibian oocytes. American Journal of Physiology. 2010;298(2):C283–C297. doi: 10.1152/ajpcell.00444.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Guizouarn H, Martial S, Gabillat N, Borgese F. Point mutations involved in red cell stomatocytosis convert the electroneutral anion exchanger 1 to a nonselective cation conductance. Blood. 2007;110(6):2158–2165. doi: 10.1182/blood-2006-12-063420. [DOI] [PubMed] [Google Scholar]
- 26.Bogdanova A, Goede JS, Weiss E, et al. Cryohydrocytosis: increased activity of cation carriers in red cells from a patient with a band 3 mutation. Haematologica. 2010;95(2):189–198. doi: 10.3324/haematol.2009.010215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Morgan PE, Pastoreková S, Stuart-Tilley AK, Alper SL, Casey JR. Interactions of transmembrane carbonic anhydrase, CAIX, with bicarbonate transporters. American Journal of Physiology. 2007;293(2):C738–C748. doi: 10.1152/ajpcell.00157.2007. [DOI] [PubMed] [Google Scholar]
- 28.Bruce LJ, Beckmann R, Ribeiro ML, et al. A band 3-based macrocomplex of integral and peripheral proteins in the RBC membrane. Blood. 2003;101(10):4180–4188. doi: 10.1182/blood-2002-09-2824. [DOI] [PubMed] [Google Scholar]
- 29.Zhu Q, Lee DWK, Casey JR. Novel topology in C-terminal region of the human plasma membrane anion exchanger, AE1. The Journal of Biological Chemistry. 2003;278(5):3112–3120. doi: 10.1074/jbc.M207797200. [DOI] [PubMed] [Google Scholar]
- 30.Delaunay J. The hereditary stomatocytoses: genetic disorders of the red cell membrane permeability to monovalent cations. Seminars in Hematology. 2004;41(2):165–172. doi: 10.1053/j.seminhematol.2004.02.005. [DOI] [PubMed] [Google Scholar]
- 31.Ellory JC, Guizouarn H, Borgese F, Bruce LJ, Wilkins RJ, Stewart GW. Leaky Cl–HCO3- exchangers: cation fluxes via modified AE1. Philosophical Transactions of the Royal Society B: Biological Sciences. 2009;364(1514):189–194. doi: 10.1098/rstb.2008.0154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Groves JD, Tanner MJA. Glycophorin A facilitates the expression of human band 3-mediated anion transport in Xenopus oocytes. The Journal of Biological Chemistry. 1992;267(31):22163–22170. [PubMed] [Google Scholar]
- 33.Williamson RC, Toye AM. Glycophorin A: band 3 aid. Blood Cells, Molecules, and Diseases. 2008;41(1):35–43. doi: 10.1016/j.bcmd.2008.01.001. [DOI] [PubMed] [Google Scholar]
- 34.Young MT, Tanner MJA. Distinct regions of human glycophorin A enhance human red cell anion exchanger (band 3; AE1) transport function and surface trafficking. The Journal of Biological Chemistry. 2003;278(35):32954–32961. doi: 10.1074/jbc.M302527200. [DOI] [PubMed] [Google Scholar]
- 35.Jennings MJ, Passow H. Anion transport across the erythrocyte membrane, in situ proteolysis of band 3 protein, and cross-linking of proteolytic fragments by 4,4’-diisothiocyano dihydrostilbene-2,2-disulfonate. Biochimica et Biophysica Acta. 1979;554(2):498–519. doi: 10.1016/0005-2736(79)90387-0. [DOI] [PubMed] [Google Scholar]
- 36.Okubo K, Kang D, Hamasaki N, Jennings ML. Red blood cell band 3. Lysine 539 and lysine 851 react with the same H2DIDS (4,4′-diisothiocyanodihydrostilbene-2,2’-disulfonic acid) molecule. The Journal of Biological Chemistry. 1994;269(3):1918–1926. [PubMed] [Google Scholar]
- 37.Bielfeld-Ackermann A, Range C, Korbmacher C. Maitotoxin (MTX) activates a nonselective cation channel in Xenopus laevis oocytes. Pflugers Archiv European Journal of Physiology. 1998;436(3):329–337. doi: 10.1007/PL00008085. [DOI] [PubMed] [Google Scholar]
- 38.Diakov A, Koch JP, Ducoudret O, Müller-Berger S, Frömter E. The disulfonic stilbene DIDS and the marine poison maitotoxin activate the same two types of endogenous cation conductance in the cell membrane of Xenopus laevis oocytes. Pflugers Archiv European Journal of Physiology. 2001;442(5):700–708. doi: 10.1007/s004240100593. [DOI] [PubMed] [Google Scholar]
- 39.Guizouarn H, Gabillat N, Borgese F. Evidence for Up-regulation of the Endogenous Na-K-2Cl Co-transporter by Molecular Interactions with the Anion Exchanger tAE1 Expressed in Xenopus Oocyte. The Journal of Biological Chemistry. 2004;279(12):11513–11520. doi: 10.1074/jbc.M311920200. [DOI] [PubMed] [Google Scholar]
- 40.Sterling D, Reithmeier RAF, Casey JR. A transport metabolon: functional interaction of carbonic anhydrase II and chloride/bicarbonate exchangers. The Journal of Biological Chemistry. 2001;276(51):47886–47894. doi: 10.1074/jbc.M105959200. [DOI] [PubMed] [Google Scholar]
- 41.Campanella ME, Chu H, Wandersee NJ, et al. Characterization of glycolytic enzyme interactions with murine erythrocyte membranes in wild-type and membrane protein knockout mice. Blood. 2008;112(9):3900–3906. doi: 10.1182/blood-2008-03-146159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chu H, Low PS. Mapping of glycolytic enzyme-binding sites on human erythrocyte band 3. Biochemical Journal. 2006;400(1):143–151. doi: 10.1042/BJ20060792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chétrite G, Cassoly R. Affinity of hemoglobin for the cytoplasmic fragment of human erythrocyte membrane band 3. Equilibrium measurements at physiological pH using matrix-bound proteins: the effects of ionic strength, deoxygenation and of 2,3-diphosphoglycerate. Journal of Molecular Biology. 1985;185(3):639–644. doi: 10.1016/0022-2836(85)90076-2. [DOI] [PubMed] [Google Scholar]
- 44.Grygorczyk R, Schwarz W. Properties of the Ca2+-activated K+ conductance of human red cells as revealed by the patch-clamp technique. Cell Calcium. 1983;4(5-6):499–510. doi: 10.1016/0143-4160(83)90025-8. [DOI] [PubMed] [Google Scholar]
- 45.Decherf G, Bouyer G, Egée S, Thomas SLY. Chloride channels in normal and cystic fibrosis human erythrocyte membrane. Blood Cells, Molecules, and Diseases. 2007;39(1):24–34. doi: 10.1016/j.bcmd.2007.02.014. [DOI] [PubMed] [Google Scholar]
- 46.Glogowska E, Dyrda A, Cueff A, et al. Anion conductance of the human red cell is carried by a maxi-anion channel. Blood Cells, Molecules, and Diseases. 2010;44(4):243–251. doi: 10.1016/j.bcmd.2010.02.014. [DOI] [PubMed] [Google Scholar]
- 47.Bennekou P, Barksmann TL, Kristensen BI, Jensen LR, Christophersen P. Pharmacology of the human red cell voltage-dependent cation channel. Part II: inactivation and blocking. Blood Cells, Molecules, and Diseases. 2004;33(3):356–361. doi: 10.1016/j.bcmd.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 48.Bennekou P, Barksmann TL, Jensen LR, Kristensen BI, Christophersen P. Voltage activation and hysteresis of the non-selective voltage-dependent channel in the intact human red cell. Bioelectrochemistry. 2004;62(2):181–185. doi: 10.1016/j.bioelechem.2003.08.006. [DOI] [PubMed] [Google Scholar]
- 49.Christophersen P, Bennekou P. Evidence for a voltage-gated, non-selective cation channel in the human red cell membrane. Biochimica et Biophysica Acta. 1991;1065(1):103–106. doi: 10.1016/0005-2736(91)90017-3. [DOI] [PubMed] [Google Scholar]
- 50.Kaestner L, Bernhardt I. Ion channels in the human red blood cell membrane: their further investigation and physiological relevance. Bioelectrochemistry. 2002;55(1-2):71–74. doi: 10.1016/s1567-5394(01)00164-5. [DOI] [PubMed] [Google Scholar]
- 51.Kaestner L, Christophersen P, Bernhardt I, Bennekou P. The non-selective voltage-activated cation channel in the human red blood cell membrane: reconciliation between two conflicting reports and further characterisation. Bioelectrochemistry. 2000;52(2):117–125. doi: 10.1016/s0302-4598(00)00110-0. [DOI] [PubMed] [Google Scholar]
- 52.Föller M, Kasinathan RS, Koka S, et al. TRPC6 contributes to the Ca2+ leak of human erythrocytes. Cellular Physiology and Biochemistry. 2008;21(1–3):183–192. doi: 10.1159/000113760. [DOI] [PubMed] [Google Scholar]
- 53.Yamaguchi T, Fujii T, Abe Y, et al. Helical image reconstruction of the outward-open human erythrocyte band 3 membrane domain in tubular crystals. Journal of Structural Biology. 2010;169(3):406–412. doi: 10.1016/j.jsb.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 54.Guizouarn H, Gabillat N, Motais R, Borgese F. Multiple transport functions of a red blood cell anion exchanger, tAE1: its role in cell volume regulation. Journal of Physiology. 2001;535(2):497–506. doi: 10.1111/j.1469-7793.2001.t01-1-00497.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Martial S, Guizouarn H, Gabillat N, Pellissier B, Borgese F. Consequences of point mutations in trout anion exchanger 1 (tAE1) transmembrane domains: evidence that tAE1 can behave as a chloride channel. Journal of Cellular Physiology. 2006;207(3):829–835. doi: 10.1002/jcp.20631. [DOI] [PubMed] [Google Scholar]
- 56.Parker MD, Young MT, Daly CM, Meech RW, Boron WF, Tanner MJA. A conductive pathway generated from fragments of the human red cell anion exchanger AE1. Journal of Physiology. 2007;581(1):33–50. doi: 10.1113/jphysiol.2007.128389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jones GS, Knauf PA. Mechanism of the increase in cation permeability of human erythrocytes in low-chloride media. Journal of General Physiology. 1985;86(5):721–738. doi: 10.1085/jgp.86.5.721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jarolim P, Palek J, Amato D, et al. Deletion in erythrocyte band 3 gene in malaria-resistant Southeast Asian ovalocytosis. Proceedings of the National Academy of Sciences of the United States of America. 1991;88(24):11022–11026. doi: 10.1073/pnas.88.24.11022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Allen SJ, O’Donnell A, Alexander NDE, et al. Prevention of cerebral malaria in children in Papua New Guinea by Southeast Asian ovalocytosis band 3. American Journal of Tropical Medicine and Hygiene. 1999;60(6):1056–1060. doi: 10.4269/ajtmh.1999.60.1056. [DOI] [PubMed] [Google Scholar]
- 60.Fiévet B, Gabillat N, Borgese F, Motais R. Expression of band 3 anion exchanger induces chloride current and taurine transport: structure-function analysis. The EMBO Journal. 1995;14(21):5158–5169. doi: 10.1002/j.1460-2075.1995.tb00200.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Tang XB, Kovacs M, Sterling D, Casey JR. Identification of residues lining the translocation pore of human AE1, plasma membrane anion exchange protein. The Journal of Biological Chemistry. 1999;274(6):3557–3564. doi: 10.1074/jbc.274.6.3557. [DOI] [PubMed] [Google Scholar]
- 62.Zhu Q, Casey JR. The substrate anion selectivity filter in the human erythrocyte Cl−/HCO3 − exchange protein, AE1. The Journal of Biological Chemistry. 2004;279(22):23565–23573. doi: 10.1074/jbc.M401380200. [DOI] [PubMed] [Google Scholar]
- 63.Barneaud-Rocca D, Borgese F, Guizouarn H. Dual transport properties of anion exchanger 1: the same transmembrane segment is involved in anion exchange and in a cation leak. The Journal of Biological Chemistry. 2011;286(11):8909–8916. doi: 10.1074/jbc.M110.166819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Larsson HP, Picaud SA, Werblin FS, Lecar H. Noise analysis of the glutamate-activated current in photoreceptors. Biophysical Journal. 1996;70(2):733–742. doi: 10.1016/S0006-3495(96)79613-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ryan RM, Vandenberg RJ. A channel in a transporter. Clinical and Experimental Pharmacology and Physiology. 2005;32(1-2):1–6. doi: 10.1111/j.1440-1681.2005.04164.x. [DOI] [PubMed] [Google Scholar]
- 66.Vandenberg RJ, Huang S, Ryan RM. Slips, leaks and channels in glutamate transporters. Channels. 2008;2(1):51–58. doi: 10.4161/chan.2.1.6047. [DOI] [PubMed] [Google Scholar]
- 67.DeFelice LJ, Goswami T. Transporters as channels. Annual Review of Physiology. 2007;69:87–112. doi: 10.1146/annurev.physiol.69.031905.164816. [DOI] [PubMed] [Google Scholar]
- 68.Miller C. ClC chloride channels viewed through a transporter lens. Nature. 2006;440(7083):484–489. doi: 10.1038/nature04713. [DOI] [PubMed] [Google Scholar]
- 69.Takeuchi A, Reyes N, Artigas P, Gadsby DC. The ion pathway through the opened Na+,K+-ATPase pump. Nature. 2008;456(7220):413–416. doi: 10.1038/nature07350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bruce LJ, Guizouarn H, Burton NM, et al. The monovalent cation leak in overhydrated stomatocytic red blood cells results from amino acid substitutions in the Rh-associated glycoprotein. Blood. 2009;113(6):1350–1357. doi: 10.1182/blood-2008-07-171140. [DOI] [PubMed] [Google Scholar]