

Abstract
Smallpox preparedness research has led to development of antiviral therapies for treatment of serious orthopoxvirus infections. Monkeypox virus is an emerging, zoonotic orthopoxvirus which can cause severe and transmissible disease in humans, generating concerns for public health. Monkeypox virus infection results in a systemic, febrile-rash illness closely resembling smallpox. Currently, there are no small-molecule antiviral therapeutics approved to treat orthopoxvirus infections of humans. The prairie dog, using monkeypox virus as a challenge virus, has provided a valuable nonhuman animal model in which monkeypox virus infection closely resembles human systemic orthopoxvirus illness. Here, we assess the efficacy of the antiorthopoxvirus compound ST-246 in prairie dogs against a monkeypox virus challenge of 65 times the 50% lethal dose (LD50). Animals were infected intranasally and administered ST-246 for 14 days, beginning on days 0, 3, or after rash onset. Swab and blood samples were collected every 2 days and analyzed for presence of viral DNA by real-time PCR and for viable virus by tissue culture. Seventy-five percent of infected animals that received vehicle alone succumbed to infection. One hundred percent of animals that received ST-246 survived challenge, and animals that received treatment before symptom onset remained largely asymptomatic. Viable virus and viral DNA were undetected or at greatly reduced levels in animals that began treatment on 0 or 3 days postinfection, compared to control animals or animals treated post-rash onset. Animals treated after rash onset manifested illness, but all recovered. Our results indicate that ST-246 can be used therapeutically, following onset of rash illness, to treat systemic orthopoxvirus infections.
INTRODUCTION
Despite the eradication of smallpox 30 years ago, orthopoxviruses (OPXVs) continue to cause human illness. Variola virus (VARV), the causative agent of smallpox, remains a bioterror agent of concern. Monkeypox virus (MPXV) is endemic to regions of Africa and caused epidemic human disease in the United States following its introduction to North American prairie dogs after importation of, and cohousing with, species from a consignment of infected African animals. Even within the United States, where endemic human OPXVs are not thought to exist, recent phylogenetic analysis describing North American OPXVs suggests that unidentified emerging and/or zoonotic OPXVs may be circulating in wild animals (2, 4, 7, 23). Although severe disease can be associated with human OPXV infection, supportive care is the current mainstay of most treatment approaches. Postexposure administration of smallpox vaccine for prevention of an OPXV infection must be given within 3 days of exposure to provide maximal benefit and within 4 to 7 days to ameliorate disease (17, 18). However, vaccination is restricted due to numerous contraindications for the vaccine, which predispose some recipients to vaccine-related adverse events. People who should not receive the vaccine are those who have weakened immune systems (e.g., HIV-infected subjects, transplant recipients, or those receiving cancer treatment), are pregnant, or suffer from heart or certain skin conditions (e.g., eczema or atopic dermatitis). While most people experience normal, mild inflammatory reactions to the vaccine, some may experience serious to life-threatening reactions such as eczema vaccinatum, progressive vaccinia, or postvaccinial encephalitis. Although vaccinia immune globulin is licensed to treat severe smallpox vaccine adverse events, in order to treat OPXV infections and adverse reactions to smallpox vaccine, it is critical to additionally develop an antiviral compound against OPXVs, especially VARV and MPXV.
Antiviral agents with activity against OPXVs have been extensively studied to provide additional strategies for postexposure and therapeutic benefit. In the United States, recommendations call for the development of two antiviral drugs having different mechanisms of action against poxviruses. ST-246 is one anti-OPXV drug that was identified by a high-throughput screening assay designed to evaluate compounds for their ability to inhibit cytopathic effects (CPE) induced in cell culture by vaccinia virus (VACV) and cowpox virus (CPXV) (27). It targets the F13L gene product of VACV, a 37-kDa palmitoylated peripheral membrane protein required for formation of extracellular virus (EV). In vitro, ST-246 selectively inhibits several members of the OPXV genus, including camelpox virus, CPXV, ectromelia virus (ECTV), MPXV, VACV, and VARV (6, 25, 27). The effective concentration of compound required to inhibit virus-induced cytopathic effect by 50% is less than 0.07 μM and 0.04 μM against six VARV strains and five MPXV strains, respectively (25). The potent, prophylactic or postexposure protective antiviral activity of ST-246 in vivo has been described by numerous investigators using multiple animal models of virulent OPXV illness (1, 8, 10, 19, 20, 24, 27). Although these studies document ST-246 as an effective antiviral drug for prophylactic or postexposure treatment of OPXV infections, they do not address potential human therapeutic use of ST-246, i.e., after definitive rash illness onset.
The current study utilizes wild-caught prairie dogs, Cynomys ludovicianus, inoculated intranasally with MPXV in order to mimic systemic monkeypox or smallpox infection in humans. Human infection with VARV or MPXV has similar clinical disease progression consisting of an asymptomatic period followed by a febrile prodrome prior to development of a generalized rash (2, 3, 13). Lesions initially erupt on surfaces of the oral mucosa such as the mouth, tongue, and oropharynx, followed by systemic spread to the skin of the face and other parts of the body. Lesions evolve through stages of macule, papule, vesicle, pustule, and scab before desquamating, often leaving scars. Prairie dogs were first discovered to be susceptible to infection with MPXV during an outbreak in 2003 in the midwestern United States (21). Prairie dogs that were cohoused with imported African rodents became infected and subsequently transmitted the virus to humans, resulting in 37 laboratory-confirmed and 10 probable cases of monkeypox (22).
Since this outbreak, several studies have been conducted to determine the potential of employing the prairie dog as a surrogate animal model for systemic OPXV infection in humans. Prairie dogs have been infected with MPXV by multiple routes, including intranasal (i.n.), intradermal (i.d.), and intraperitoneal (i.p.), thus confirming their high susceptibility to the virus (12, 9, 26). Animals that were infected i.p. succumbed too quickly for a complete description of disease presentation, but animals infected by other infection routes exhibited similar symptoms. The most common symptoms included lethargy, nasal discharge, inappetence, weight loss, and lesion development between 9 to 12 days postinoculation. More importantly, the illness in prairie dogs followed a course similar to that seen with human monkeypox virus infection, consisting of an asymptomatic period followed by a generalized rash that progressed from macules to pustules. These observations provide clear evidence that MPXV infection in prairie dogs is a valuable surrogate system for monkeypox virus or smallpox virus infection in humans.
In this report, we evaluate the efficacy of ST-246 to protect prairie dogs infected i.n. with 65 times the 50% lethal dose (LD50) of virions from a Central African strain of MPXV from disease and death. ST-246 was administered by oral gavage for a period of 14 days starting on days 0 and 3 or on the day of rash onset to assess its prophylactic, postexposure, and therapeutic potential. Animals were monitored daily for symptom presentation, and blood and swabs were collected every 2 days for viral analysis by tissue culture and real-time PCR.
MATERIALS AND METHODS
Many of the materials and methods have been previously described (12).
Animals.
Black-tailed, juvenile prairie dogs (Cynomys ludovicianus) were caught from the wild in western Kansas. At the time this study began, animals were 3 years old. During the course of study, prairie dogs were housed in large rat cages covered with aerosol filter tops. Cages were maintained inside a Duo-Flow biosafety cabinet (Biochem Lab Products, Seaford, DE) in an animal biosafety level 3 (ABSL-3) room. Animals were cared for in accordance with the CDC Institutional Animal Care and Use Committee (IACUC) under an approved protocol (1563DAMPRAC). Animal handling was performed using biosafety level 3+ personal protective equipment (PPE). In addition to prairie dog chow and hay, animals were provided with monkey biscuits for added dietary enrichment. The average starting weight of animals was 952 g (range, 752 to 1,210 g).
Virus.
MPXV strain ROC-2003-358, was collected from a 2003 outbreak of MPXV in the Republic of Congo (ROC) and has been fully sequenced (16). It had been passaged twice in African green monkey kidney cells (BSC-40) prior to seed pool production. Viral stocks were titrated prior to storage at −80°C. The VACV mutant virus VACV-N267D was provided by SIGA Technologies.
Virus isolates obtained from selected swab samples collected from prairie dogs before (prairie dog 14 [PD14] oral sample, day 8) or posttreatment (PD7 oral, day 22; PD10 oral, day 22; PD11 oral, day 20; PD12 ocular, day 24; PD16 oral, day 18; PD17 oral, day 18) were propagated in tissue culture. Briefly, 100 μl of each original swab eluate was combined with 900 μl of RPMI medium containing 2% fetal bovine serum (FBS) and inoculated onto a T25 flask confluent with BSC-40 cells. Cultures were incubated at 35.5°C until cytopathic effects reached 90%. Monolayers were collected by scraping, and samples were frozen at −80°C. Passage 1 (P1) lysates were thawed, and 1 ml from each isolate was diluted in RPMI medium containing 2% FBS and inoculated onto eight T162 flasks confluent with BSC-40 cells. Cultures were incubated at 35.5°C until CPE reached 90%, at which time P2 lysates were collected in 250-ml Nalgene bottles after monolayers were dislodged with a cell scraper. Lysates were centrifuged at 2,500 rpm for 30 min in a Beckman J14 rotor. Supernatants were removed, and pellets were resuspended in RPMI medium containing 2% FBS. Crude virus preparations were freeze-thawed two times in a dry ice-ethanol bath and 35.5°C incubator, sonicated three times in a cup horn sonicator at 40% output with 1-min intervals, and frozen at −80°C until drug resistance testing. A small aliquot of each P2 crude preparation was removed prior to freezing, and titers were determined on BSC-40 cells.
Animal challenge.
Virus stock was serially diluted 10-fold to obtain an infectious dose of 105 PFU. Inoculum titer was retitrated by plaque counts postinfection (p.i.) and was found to be slightly higher at 3.8 × 105 PFU. All animals were infected by an i.n. route of inoculation while under anesthesia (5% isoflurane). Sixteen prairie dogs were inoculated by use of a micropipettor with a total volume of 10 μl (5 μl in each nostril) of virus suspended in phosphate-buffered saline (PBS). Additionally, two animals were mock infected with the same volume of PBS.
ST-246 treatment.
ST-246, provided by SIGA Technologies (Corvalis, OR), was formulated at 30 mg/ml in a suspension vehicle containing 1% methylcellulose and 0.5% Tween 80. Prairie dogs were administered ST-246 or vehicle alone at 1 ml/kg of body weight (30 mg/kg) daily by oral gavage for a period of 14 days. Prairie dogs were divided into four weight-adjusted groups (range of average group weights, 876 to 976 g) of 4 animals, each containing 2 males and 2 females. Two additional female animals served as uninfected control animals. Three groups were treated with ST-246 either prophylactically (day 0), postexposure (day 3), or therapeutically (post-rash onset). The fourth group received vehicle alone beginning on day 3, and the uninfected control animals received ST-246 also beginning on day 3.
Observations and sampling.
Animals were monitored daily and observations (food consumption, mobility, general symptoms, and disease progression) were recorded for a period of 30 days, with exceptions for uninfected control animals and animals in the therapeutic treatment group, which were extended to day 50 due to one animal having a delayed onset of rash illness. Every 2 days the animals were anesthetized with 5% isoflurane, and samples were collected. Prairie dogs were weighed, and sterile Dacron swabs were used to collect ocular, anal, and oropharyngeal (oral) samples which were stored at −70°C without diluent. Blood was collected from the femoral vein in EDTA-coated tubes (Sarstedt) prior to study onset, on sampling days, and when the animal expired or was euthanized. Swabs and blood were processed and prepared for DNA analysis and virus titration (see below). Lesion counts were conducted on areas of the face, inner hind legs, and genitalia, which have relatively less fur. Additionally, an area approximately 5 cm2 on the back of the animal was shaved prior to the start of the study in order to better visualize the development and progression of lesions on the trunk.
Necropsy and tissue specimen collection.
Animals that survived the challenge were humanely euthanized, and necropsies were performed at 36 days p.i. with exceptions for uninfected control animals and animals in the therapeutic treatment group, which were extended to day 57 due to one animal having a delayed onset of rash illness. Carcasses from animals that did not survive the challenge were frozen at −70°C, and necropsies were performed on day 57. Necropsies on all animals were performed according to IACUC standards in an ABSL-3 laboratory and utilizing full ABSL-3 PPE. Tissues harvested during necropsy included heart, liver, kidney, lung, skin (lesion material or inner leg), spleen, mesenteric lymph node, tongue, and eyelid. Instruments were cleaned and decontaminated with 3% Amphyl and 10% bleach between collections of each tissue. Tissues were frozen at −70°C prior to further processing. Oral, ocular, and anal swabs were collected with sterile Dacron swabs and stored without diluent at −70°C. Serum was separated from whole blood by centrifugation and processed for serology (see below). Tissues and swabs were subsequently processed and further prepared for DNA analysis and virus titration (see below).
Sample preparation.
Sample processing was performed under BSL-2 conditions with BSL-3 work practices. For DNA analysis of blood, 100 μl of sterile water was added to 100 μl of blood in order to achieve a total volume of 200 μl and to prevent coagulation. Samples were incubated at 55°C for 1 h, and genomic DNA was extracted using a BioRobot EZ-1 Workstation (Qiagen). The swab extraction tube system (SETS; Roche) protocol was used to recover sample from swabs. Swabs were hydrated with 400 μl of phosphate-buffered saline (PBS), and eluates were collected by centrifugation. A 100-μl aliquot of the swab eluate was mixed with 90 μl of buffer G2 (Qiagen) and 10 μl of proteinase K and incubated at 55°C for 1 h. DNA was extracted using the BioRobot EZ-1 Workstation. Subsequently, remaining swab eluate was used for virus titration (see below). Tissue samples were suspended in 1 ml of PBS in a 2-ml Sarstedt tube containing a single grinding ball. Tissues were ground using a GenoGrinder 2000 (OPS Diagnostics) at 600 strokes per min for 2 min. Tissue sample aliquots of 100 μl were mixed with 90 μl of buffer G2 (Qiagen) and 10 μl of proteinase K and incubated at 55°C for 1 h. Genomic DNA was extracted using the BioRobot EZ-1 Workstation. The remaining homogenized tissue samples were used for virus titration (see below).
Real-time PCR analysis.
Samples were tested in duplicate by real-time PCR using forward and reverse primers and probe complementary to the conserved OPXV E9L (DNA polymerase) gene (14). Representative samples from each animal were confirmed for MPXV DNA using forward and reverse primers and probe specific to the MPXV cytokine response modifier B gene (15). A standard set of MPXV DNAs in 10-fold dilutions (5 fg to 5 ng) was used as positive controls for both tests. The limit of detection for a positive sample was 5 fg.
Virus titration.
Previous analyses demonstrated that real-time PCR detection of MPXV DNA was significantly more sensitive than detection of plaque-forming virus as an indicator of virus infection (11). Therefore, specimens were first tested for the presence of OPXV DNA by real-time PCR and, if positive, were subsequently evaluated for viable virus by tissue culture propagation. Each swab eluate or tissue sample homogenate was sonicated in a cup horn sonicator set at an output level of 40%. Titrations were made by preparing 10-fold dilutions of swab eluate or tissue homogenate and inoculating confluent monolayers of BSC-40 cells. Monolayers were incubated at 35.5°C and 6% CO2 for 72 h. Plaques were visualized by staining cell monolayers with crystal violet containing formalin. The limit of detection was 10 PFU.
Serologic analysis.
As previously described, a modified enzyme-linked immunosorbent assay (ELISA) was used for analysis of anti-OPXV immunoglobulin types A and G (12). Ninety-six-well plates were divided into two halves, one containing killed VACV and the other containing BSC-40 cell lysate. Serum samples were added identically to both halves of the plates at a dilution of 1:100 in assay diluent. Optical densities (OD) were measured on a spectrophotometer (Victor 3 Multilabel Counter; PerkinElmer, Waltham, MA) at 450 nm. Reported values represent the average of duplicate wells for each sample. Both positive and negative human anti-VACV sera were used as assay controls. The BSC-40 cell lysate half of each plate was used to generate a cutoff value (COV) for each plate by averaging lysate OD values and adding 2 standard deviations. Specimens were considered positive if the test sample's value was above the COV.
Evaluation of drug resistance: plaque assessment and virus comet reduction IHC.
Confluent monolayers of BSC-40 cells were infected with approximately 50 PFU of virus suspended in RPMI medium containing 2% FBS. After a 1-h incubation at 35.5°C, inocula were removed and replaced with 2 ml of RPMI medium containing 2% FBS–0.1% dimethyl sulfoxide (DMSO) with or without 0.005 to 5 μM ST-246. Cultures were incubated at 35.5°C on a fixed incline of approximately 5° for 2 (VACV) or 3 (prairie dog isolates) days. Medium was removed, and monolayers were fixed for 20 min with 2 ml of PBS containing 10% formalin. Immunohistochemistry (IHC) was performed as previously described (5, 25).
Virus isolates obtained from selected swab samples collected from prairie dogs, previously quantified to have greater than 50 PFU/ml, before (PD3 oral swab, day 22), during (PD14 anal swab, day 22), or after (PD11 oral swab, day 18; PD11 oral swab, day 20) treatment were plaqued in the presence or absence of 0, 0.03, 0.3, or 5.0 μM. Control viruses were wild-type MPXV-ROC-358 and a highly ST-246-resistant virus, VACV-N267D.
RESULTS
Prophylactic or 3-day-postexposure treatment with ST-246 protects animals from death and most signs of illness.
Beginning on day 8, infected, sham-treated animals administered vehicle alone displayed general symptoms such as inappetance, facial swelling, nasal discharge, and congestion (Table 1). Individual animal symptoms also included mouth breathing and red, crusty, and/or bloody nose. After initially gaining weight due to daily supplementation of their diet with monkey biscuits, animals began losing weight on days 6 to 8. On day 10 a vesicular rash was observed on PD8 over much of the body including areas of the mouth, tongue, chest, extremities, and groin. Three of the four animals died between days 10 and 12. PD5 and PD15 succumbed to infection by day 10, and PD8 expired during sample collection on day 12. They lost 2 to 10% of their starting weights at the time of death (Fig. 1). Although no papular or pustular lesions were observed at the time of death on PD5 or PD15, blood was present around the nose and mouth, and a petechial rash was present on the chest of PD5. MPXV DNA was initially detected in blood (Fig. 2) and oral swabs on days 4 to 6 for these 3 animals, increased 3 to 5 logs (with the exception of PD15) in oral swabs, and persisted until death (Fig. 3). MPXV DNA was initially detected in ocular and anal swabs on days 6 to 8 for 2 (PD5 and PD8) of the 3 animals and persisted through death. Infectious virus was recovered from oral swabs initially on day 6 (1.7 × 101 PFU/ml), peaked at days 10 to 12 (4.0 × 105 to 4.3 × 107 PFU/ml), and persisted through death; infectious virus from ocular swabs was both initially recovered and peaked on day 8 (4.3 × 102 to 2.8 × 104 PFU/ml), and persisted through death; infectious virus from anal swabs was initially recovered on day 8 (8.8 × 104 PFU/ml), peaked at days 10 to 12 (3.6 × 105 to 1.7 × 106 PFU/ml), and persisted through death (Fig. 4). Viral DNA and viable virus were detected in all tissues collected during necropsy (Fig. 5). Concentrations of virus were highest in different tissues for each animal: for PD5, liver; PD8, tongue; and PD15, lymph node. Anti-OPXV antibodies were detected in one animal, PD8, from this group on day 12 (Fig. 6).
Table 1.
Comparison of disease presentation and molecular findings between prairie dog treatment groupsa
| ST-246 treatment |
Symptoms (no. of PDs affected) | Day of symptom onset | Day of rash onset | Lesion count | No. of surviving PDs | Day(s) of MPXV DNA detection in: |
Days of viable virus detection | Detection of Ig antibody (no. of positive PDs) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Group (n)b | Day(s) of administration | Swabs | Blood | |||||||
| Prophylaxis (4) | 0–13 | NA | NA | NA | NA | 4 | 8–26 | 24 (PD10) | 12–24 | 4 |
| Postexposure (4) | 3–16 | NA | NA | NA | NA | 4 | 6–28 | None | 8–26 | 4 |
| Therapeutic (4) | 10–23, 24–37 (PD3) | Facial swelling, nasal discharge, nasal crust, bloody nose, inappetance, weight loss, pustular lesions (4) | 8 (PD2, PD12, PD14), 24 (PD3) | 10 (PD2, PD12, PD14), 24 (PD3) | <10 (PD2, PD12, PD14), >50 (PD3) | 4 | 4–30 | 6–26 (PD2, PD3, PD14) | 4–30 | 3 (PD2, PD3, PD14) |
| Vehicle control (4) | None | Facial swelling, nasal discharge, nasal crust, bloody nose, inappetance, weight loss, pustular lesions, petechial rash, mouth breathing (3) | 8 (PD5, PD8, PD15) | 10 (PD8) | 10–25 (PD8) | 1 (PD6) | 4–12 (PD5, PD8, PD15) | 4–12 (PD5, PD8, PD15) | 6–12 (PD5, PD8, PD15) | 1 (PD8) |
| Uninfected control (2) | 3–16 | NA | NA | NA | NA | 2 | None | None | None | 0 |
Clinical and molecular observations in prairie dogs (PDs) infected with MPXV and treated with ST-246. Prairie dogs were inoculated intranasally with 3.8 × 105 PFU of MPXV or mock infected with PBS and treated orally with ST-246 or vehicle for a period of 14 days. Observations were recorded for 30 days p.i., with swab and blood sample collections every 2 days for analysis of viral DNA and viable virus. Serum was collected from blood and monitored for antibody production. Specific PDs are indicated in parentheses where appropriate.
Therapeutic, post-rash onset; n, number of animals.
Fig. 1.

Percent fluctuations in weight. Prairie dogs were infected on day 0 with 3.8 × 105 PFU, and weights were measured every 2 days for a period of 30 days. The percent change in weight for each measurement was calculated from the starting weights on day 0 and is plotted as a function of days postinfection for each treatment group. Untreated animals (A) lost weight until death, except for animal PD6. Animals that received ST-246 treatment at rash onset (B) on day 10 (PD2, PD12, and PD14) or day 24 (PD3) lost weight during illness and regained weight upon recovery, except for PD12. Uninfected control animals (not shown) and animals that received ST-246 treatment beginning on day 0 (C) or day 3 (D) steadily increased in weight.
Fig. 2.

Quantitation of viral DNA in blood from prairie dogs infected with MPXV. Treatment with ST-246 reduces MPXV DNA in blood. Prairie dogs were infected on day 0 with 3.8 × 105 PFU, and blood was collected every 2 days over a period of 30 days. Animals began receiving vehicle alone on day 3 (A) or received ST-246 at rash onset (B), on day 0 (C), or on day 3 (D). DNA was extracted, and the E9L gene was amplified and quantitated by real-time PCR. Average values of genome copies per milliliter and standard deviations were plotted on a log scale as a function of day postinfection (A, C, and D) or day post-rash onset (B).
Fig. 3.

Quantitation of viral DNA in secretions from prairie dogs infected with MPXV. Early treatment with ST-246 greatly reduces MPXV DNA in secretions. Prairie dogs were infected on day 0 with 3.8 × 105 PFU, and swab samples were collected every 2 days over a period of 30 days. Animals began receiving vehicle alone on day 3 (A, B, and C) or ST-246 at rash onset (D, E, and F), on day 0 (G, H, and I), or on day 3 (J, K, and L). DNA was extracted from oral swabs (A, D, G, and J), ocular swabs (B, E, H, and K), and anal swabs (C, F, I, and L). The E9L gene was amplified and quantitated by real-time PCR. Average values of genome copies per milliliter and standard deviations were plotted on a log scale as a function of day postinfection (A, B, C, G, H, I, J, K, and L) or day post-rash onset (D, E, and F).
Fig. 4.

Quantitation of infectious virus in secretions from prairie dogs infected with MPXV. Treatment with ST-246 reduces infectious virus in secretions. Prairie dogs were infected on day 0 with 3.8 × 105 PFU, and swab samples were collected every 2 days over a period of 30 days. Animals began receiving vehicle alone on day 3 (A, B, and C) or ST-246 at rash onset (D, E, and F), on day 0 (G, H, and I), or on day 3 (J, K, and L). Infectious virus was recovered from oral swabs (A, D, G, and J), ocular swabs (B, E, H, and K), and anal swabs (C, F, I, and L) by tissue culture. The number of PFU was quantitated by plaque counts, and average values of PFU per milliliter and standard deviations were plotted on a log scale as a function of day postinfection (A, B, C, G, H, I, J, K, and L) or day post-rash onset (D, E, and F).
Fig. 5.

Quantitation of viral DNA and infectious virus in tissues of prairie dogs infected with MPXV. High concentrations of viral DNA and infectious virus were detected in tissues from prairie dogs that received vehicle treatment. Prairie dogs were infected with 3.8 ×105 PFU and administered vehicle daily from day 0 until death. PD5 and PD15 died on day 10, and PD8 died on day 12. Heart, liver, kidney, lung, skin, spleen, lymph node (LN), tongue, and eyelid were harvested for molecular analysis by RT-PCR and tissue culture. Average values for genome copies per milliliter (A) and PFU per gram of tissue (B) along with standard deviations are plotted on log scale as a function of tissue type.
Fig. 6.

Production of anti-OPXV antibodies in prairie dogs infected with MPXV. Initial detection and strength of antibody production in prairie dogs infected with MPXV produced different immune profiles dependent on ST-246 treatment regimen. Prairie dogs were infected with 3.8 × 105 PFU, and serum was collected from blood drawn every 2 days over a period of 30 days. Animals began receiving vehicle alone on day 3 (A) or ST-246 at rash onset (B), on day 0 (C), or on day 3 (D). Serum was diluted 1:100 and tested against whole, inactivated vaccinia virus, and antibody production was measured by ELISA. Test serum was considered positive if its absorbance value was above the COV, which was determined by averaging OD measurements of BSC-40 cell lysates and adding 2 standard deviations.
An outlier of the control vehicle-treated group, PD6, survived infection. The only symptom noted in this animal was a red, puffy nose on days 8 and 9 p.i. Infectious virus was not recovered from any swab samples, and detectable levels of viral DNA were present only on day 10 from an ocular swab, on day 12 from an oral swab, and on day 14 from an anal swab. Viral DNA was not detected in blood, and anti-orthopoxvirus antibodies were not detected in serum.
Prairie dogs treated with ST-246 initiated on day 0 (prophylactic) and administered daily through day 13 p.i. remained visibly asymptomatic for the entire 30 days of observations. No animals lost more than 5% body weight, and all animals survived (Fig. 1). No viable virus was isolated from any swab samples while animals were on treatment except for two oral swabs on day 12. MPXV DNA was detected on day 8 in one animal (PD11) from an oral swab. Interestingly, MPXV DNA was detected in oral swabs from all 4 animals on day 14 (4.2 × 101 to 2.2 × 102 genome copies/ml), and quantitative levels peaked on days 18 to 22 (2.0 × 104 to 4.1 × 105 genome copies/ml) (Fig. 3). MPXV DNA was no longer detected in oral swabs after day 26. Lower concentrations of MPXV DNA were detected in ocular swabs between days 14 and 20 (4.2 × 101 to 5.5 × 102 genome copies/ml) from 3 of 4 animals (PD11, PD16, and PD17), in anal swabs from PD16 on day 26 (6.3 × 101 genome copies/ml), and in blood (Fig. 2) from PD10 on day 24 (5.6 × 101 genome copies/ml). Infectious virus was cultured from oral swabs between days 12 and 24 from all 4 animals, with peak titers (4.6 × 102 to 1.4 × 105 PFU/ml) occurring between days 18 and 22 p.i. (Fig. 4). Viable virus was not recovered from ocular or anal swabs. Antiorthopoxvirus antibodies were first detected on day 14 for PD11, day 16 for PD16 and PD17, and day 20 for PD10. Peak levels (0.403 to 0.551 OD) were measured on day 30 (Fig. 6).
Prairie dogs initially treated with ST-246 at 3 days postexposure, with daily administration through day 16 p.i., also remained visibly asymptomatic for the 30 days of observations. Similar to animals treated prophylactically, no animals lost more than 5% body weight, and all animals survived infection (Fig. 1). Viral DNA was initially detected in oral swabs from all 4 animals on day 8 and peaked between days 8 and 12 (2.3 × 103 to 3.6 × 103 genome copies/ml) for 3 of the animals (PD1, PD4, and PD13) (Fig. 3). Viral DNA was not detected in any of these animals on days 14 and 16, but levels were measured between days 18 and 26 from PD7, peaking on day 24 (2.7 × 104 genome copies/ml), and from PD4 on day 24. Viral DNA was initially detected in ocular swabs on day 8, peaking on day 12 (7.9 × 101 to 2.5 × 102 genome copies/ml) for 3 animals (PD1, PD4, and PD7), and was detected in anal swabs on days 6 and 12 (6.2 × 101 to 2.7 × 102 genome copies/ml) from PD1 and PD4. No viral DNA was detected in blood (Fig. 2). Infectious virus was cultured from oral swabs from 3 animals (PD1, PD4, and PD13) between days 8 and 12 (2.5 × 101 to 3.7 × 103 PFU/ml) and from PD7 between days 12 and 26 (<101 to 4.4 × 102 PFU/ml) (Fig. 4). Viable virus was recovered from one ocular swab on day 12 from PD1 (1.7 × 102 PFU/ml), and no virus was recovered from anal swabs. Anti-orthopoxvirus antibodies were first detected on day 12 for PD7 and PD13 and on day 14 for PD1 and PD4. Peak levels (0.194 to 0.499 OD) were measured on days 28 to 30 (Fig. 6).
Therapeutic use of ST-246, after rash onset, protects animals from death.
Consistent with the vehicle-alone treatment group, prairie dogs in the rash onset (therapeutic) treatment group exhibited the first visible signs of illness at 8 days p.i. Although their symptoms were identical to those of the vehicle control group, they were less severe and were apparent until day 20 p.i. (Table 1). Rash consisting of 5 to 50 lesions was first observed in 3 animals (PD2, PD12, and PD14) on day 10, appearing as macules on areas of the chest, abdomen, back, groin, and tongue. At this time, ST-246 treatment was initiated for these 3 animals. Lesions on PD2 and PD14 developed into pustules by day 12; they dried and scabbed by day 16, and crusts desquamated by day 22. In contrast, the lesions on PD12 did not progress to the pustular stage and, in fact, were no longer visible on day 14. One animal from this group, PD3, was observed as having a vesicular rash on day 24, 2 weeks later than the other animals with rash (Fig. 7). ST-246 treatment was initiated for PD3 on day 24 and continued through day 37. Greater than 50 lesions were counted on areas of the face, mouth, tongue, abdomen, back, extremities, and groin. Lesions progressed classically into pustules before drying and forming scabs by day 28. Scabs had largely desquamated by day 40, at which time the animal appeared to have fully recovered from illness. The 3 prairie dogs (PD2, PD3, and PD14) that developed a pustular rash experienced the most weight loss, after initially gaining weight due to diet supplementation. They lost 10 to 24% of their starting weights, reaching their lowest points 6 days post-rash onset (Fig. 1). Each animal symptomatically recovered at the time lesions had resolved, and weight gain resumed 6 to 8 days following rash onset. PD12, whose rash did not progress beyond the macular stage, exhibited continuous weight gain throughout the study.
Fig. 7.

PD3 developed late-onset rash on day 24. Lesions, numbering greater than 50, were present on areas of the face, mouth, tongue, abdomen, back, extremities, and groin. Pustular lesions present on the tongue on day 24 (A) resolved by day 40 (B), and lesions present on the back formed scabs by day 28 (C) and mostly resolved by day 40 (D).
Since animals treated therapeutically developed rash on different days postinfection, we plotted concentrations of MPXV DNA shed in secretions by these animals as a function of post-rash onset rather than postinfection. Molecular analysis revealed similar trends in shedding among three of the animals (PD2, PD3, and PD14). MPXV DNA was initially detected in oral swabs 4 (PD2), 6 (PD14), or 12 (PD3) days prior to rash onset from these 3 animals, peaked on the day of rash onset or 2 days post-rash onset (8.3 × 105 to 3.0 × 106 genome copies/ml), and persisted until 22 days post-rash onset (Fig. 3). MPXV DNA was detected in ocular swabs 2 to 4 days prior to rash onset, peaked on the day of rash onset or 2 days post-rash onset (9.8 × 104 to 7.5 × 105 genome copies/ml), and persisted until 20 days post-rash onset. Shedding of viral DNA was detected in anal swabs beginning 0 to 2 days prior to rash onset, peaked at 2 to 6 days post rash onset (2.6 × 104 to 1.9 × 106 genome copies/ml), and persisted until 18 days post-rash onset. MPXV DNA was mostly detected in blood from 4 days prior to rash onset to 2 days post-rash onset with concentrations remaining below 103 genome copies/ml (Fig. 2). The other animal from this group, PD12, mostly shed DNA in each of the swab samples much later in the infection than the other 3 animals. In oral swabs from PD12, MPXV DNA was initially detected and peaked 2 days post-rash onset (1.2 × 104 genome copies/ml) but not again until day 14 and cleared by day 22. Similarly, viral DNA was initially detected in ocular swabs 4 days post-rash onset, peaked on day 14 (2.8 × 103 genome copies/ml), and cleared by day 20. Anal swabs from PD12 yielded MPXV DNA only on days 14 and 16 at concentrations below 103 genome copies/ml. No viral DNA was measured in blood (Fig. 2).
Infectious virus recovered from secretions was also analyzed as a function of day post-rash onset rather than day postinfection. Viable virus was initially detected in oral swabs on 6 (PD2 and PD14) or 12 (PD3) days prior to rash onset in 3 prairie dogs and at 2 days post-rash onset from PD12. Virus concentrations peaked between 2 days prior to and 2 days post-rash onset (4.6 × 104 to 1.1 × 107 PFU/ml) and was cleared by 18 days post-rash onset (Fig. 4). Virus was initially recovered from ocular swabs 2 days prior to rash onset (PD14), on the day of rash onset (PD2 and PD3), and 14 days post-rash onset (PD12). Peak levels were between days 0 and 2 (2.1 × 102 to 1.4 × 103 PFU/ml) for PD2, PD3, and PD14 or on day 14 (4.2 × 101 PFU/ml) for PD12, and virus was cleared by 18 days post-rash onset. In anal swabs, virus was initially detected and peaked (3.3 × 101 to 3.1 × 104 PFU/ml) on the day of rash onset to 2 days postonset from 3 animals (PD2, PD3, and PD14) and was cleared by 18 days post-rash onset. Infectious virus was not detected from anal swabs from PD12. Anti-orthopoxvirus antibodies were first detected 0 to 4 days post-rash onset from PD2, PD3, and PD14. Antibody production peaked (0.799 OD) and reached steady-state levels for PD2 by 26 days post-rash, but steady-state endpoint levels had not apparently been reached by the end of the study for PD3 and PD14 after 16 or 30 days post-rash onset, respectively (Fig. 6). Anti-orthopoxvirus antibodies were undetected in serum from PD12.
Uninfected control animals.
PD9 and PD18 uninfected control animals remained asymptomatic throughout the study, and MPXV DNA, infectious virus, and anti-orthopoxvirus antibodies were not detected.
Evaluation for drug resistance was performed by virus comet reduction IHC and plaque reduction assessment.
An unanticipated observation was the demonstration of viable virus in animals after cessation of prophylactic, postexposure, or therapeutic therapy. Six MPXV isolates obtained posttreatment from different animals were compared with both an MPXV isolate obtained from a prairie dog prior to therapeutic treatment and with controls for sensitivity to ST-246. At ST-246 concentrations of 0.5, 0.1, and 0.05 μM, MPXV comet formation was inhibited, and overall plaque size was considerably diminished in all prairie dog-obtained isolates and was equivalent to that in control vaccinia-IHDJ virus (Fig. 8). This assay has comparable sensitivity to a standard 50% effective concentration (EC50) assay (25). An ST-246-resistant VACV provided by SIGA Technologies showed no variation in plaque size at ST-246 concentrations of 0.005 to 10 μM from that seen in the absence of ST-246; interestingly, the mutant VACV-N267D showed no comet formation, even in the absence of ST-246. At lower concentrations of ST-246 (0.01 and 0.005 μM) and in untreated wells, comets were observed in all monkeypox virus (pre- and post-ST-246) isolates from prairie dogs and in the control VACV-IHDJ. Viral resistance was also evaluated by plaquing isolates in the presence of ST-246. Four prairie dog isolates, one prior to onset of therapy, one during therapy, and two postcessation of therapy, along with wild-type MPXV-ROC-358 and ST-246-resistant VACV-N267D, were plaqued in the presence or absence of 0, 0.03, 0.3, and 5.0 μM ST-246. Only the isolates obtained on the 13th day of treatment (PD3 day 22) and on the fifth day postcessation of prophylactic therapy (PD11 day 18) were viable at the time of this experiment, and no plaques were observed when samples from PD3 day 22, PD11 day 18, or MPXV-ROC-358 were grown in the presence of 0.03, 0.3, or 5.0 μM ST-246 (Fig. 9). In contrast, VACV-N267D showed equivalent plaque counts and plaque sizes, at all concentrations of ST-246 in the media, to those observed when virus was plaqued in the absence of ST-246.
Fig. 8.

Evaluation of drug resistance in posttreatment virus isolates from prairie dogs by comet reduction. Approximately 50 PFU from swab isolates, VACV-IHDJ, and ST-246-resistant VACV-N267D was inoculated onto confluent monolayers of BSC-40 cells. Cultures were incubated with or without ST-246 for 2 (IHDJ and N267D) or 3 days (all PD isolates) and fixed with PBS containing formalin. Plaques and comets were visualized by IHC.
Fig. 9.
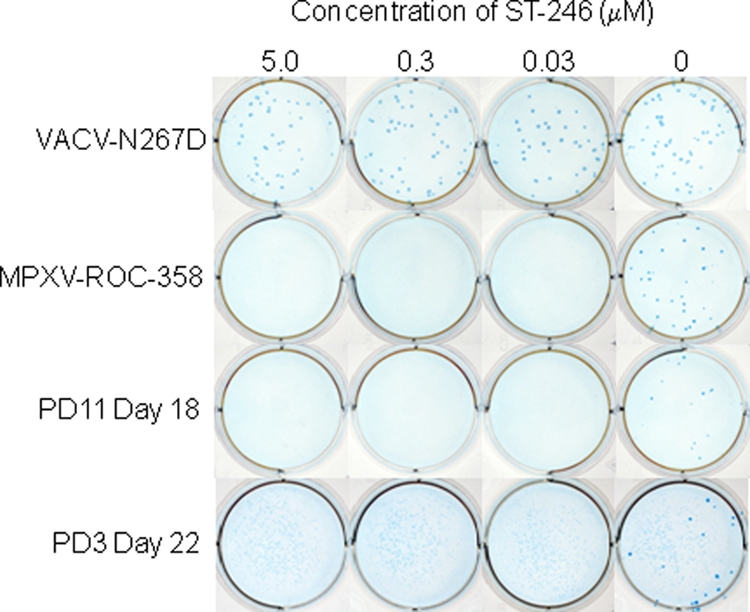
Evaluation of drug resistance in posttreatment virus isolates from prairie dog 5 by plaque reduction. Approximately 50 PFU from oral swab isolates, ST-246-resistant VACV-N267D, and wild-type MPXV-ROC-358 was inoculated onto confluent BSC-40 cells in the presence or absence of ST-246. Cultures were grown for 3 days and fixed with PBS containing formalin. Plaques were visualized by IHC.
DISCUSSION
The data presented in this study clearly demonstrate that ST-246 is efficacious for prophylactic, postexposure, and therapeutic (after rash onset) treatment of monkeypox virus infection in prairie dogs. Animals inoculated with an infectious dose of virus equivalent to 65× the LD50 and administered antiviral treatment initiated on the day of infection or by 3 days postinfection were asymptomatic. Furthermore, all infected animals that began antiviral treatment following rash onset survived infection, whereas three infected animals that did not receive antiviral treatment became severely ill and died. These 3 animals (PD5, PD8, and PD15) underwent an asymptomatic period of 8 days until symptoms consistent with an upper respiratory infection were observed. The disseminated rash observed on PD8 consisted of variously sized, vesicular lesions on multiple areas of the face and body. Although a pustular rash was not observed on PD15, its death on day 10 likely preceded rash eruption. Previous studies by Hutson et al. have shown that rash onset with a Congo Basin strain of MPXV typically occurs on days 9 to 13 p.i. (12, 9). The rash observed on PD5 was a petechial rash and was present in addition to bleeding from the nose. These symptoms are reminiscent of symptoms from human cases of hemorrhagic smallpox, which is a more virulent form of smallpox than ordinary smallpox. Each of these 3 prairie dogs was losing substantial weight at the time of their deaths, and PCR and culture analysis of tissues harvested at necropsy demonstrated that virus had spread to vital target organs, including liver, kidney, and spleen, which probably contributed to their deaths. Unexpectedly, PD6 in this group was asymptomatic; and although random samples were DNA positive, no infectious virus was detected from swab samples, and it did not seroconvert by day 30, suggesting that it was not successfully infected at the time of inoculation.
Similarly, prairie dogs in the rash onset treatment group were asymptomatic for 8 days and then developed upper respiratory symptoms identical to those in the vehicle control group. After antiviral treatment was initiated at rash onset, symptoms generally resolved in 10 days. Specifically, 3 of the 4 prairie dogs (PD2, PD12, and PD14) developed a rash on the same day (10) as the vehicle control “untreated” animal, PD8. Rash progressed as expected in two (PD14 and PD2) of these animals and had resolved by day 16 or 22, respectively. In contrast, the macular rash observed on PD12 did not progress and quickly resolved by day 14. These findings are quite remarkable compared to historic control prairie dogs infected with doses as high as 8 × 105 PFU of the same MPXV strain as used in this study; in these animals lesions did not resolve until 24 to 27 days postinoculation (9). PD12 also did not lose weight or produce measurable levels of antiorthopoxvirus antibodies. Interestingly, PD3 developed late-onset rash on day 24, developed the greatest number of lesions (50+), and produced the highest levels of antibodies. Hutson et al. also observed one prairie dog that experienced a late onset of infection with lesions developing on day 27 p.i. and ultimately succumbing to infection (9). However, for PD3, weight loss and other observable symptoms were not dissimilar from symptoms of the other animals in this group. Moreover, once ST-246 treatment began, the lesions dried and resolved over a period similar to that of the therapeutically treated animals. Differences in rash onset and progression in these animals are difficult to explain. Since these prairie dogs are wild-caught, outbred animals, there is individual variability in their host responses to viral infection and in their responses to drug treatment, both of which could influence disease progression.
Because the prairie dogs in the vehicle-only control group either died early in the study or were not infected, a comparison can be made of the clinical, virological, and molecular data only between therapeutically treated animals and vehicle control animals through day 12. Up to that point, concentrations of MPXV DNA and infectious virus followed the same trends for both groups. In animals treated therapeutically, MPXV DNA and infectious virus reached peak values for all sample types, including blood, at or within 4 days of rash onset and then decreased with initiation of ST-246. Viral DNA was detected from 6 days prior to 20 days post-rash onset, and infectious virus was recovered from 6 days prior to 16 days post-rash onset. The greatest amounts of viral DNA and viable virus were obtained from oral secretions, followed equally by ocular and anal secretions. The absence of an infected survivor from the vehicle control group prevented a comparison between the molecular findings and the duration of recovery for animals treated therapeutically and untreated animals. This analysis would have been beneficial for more adequately describing the impact that ST-246 has on viral clearance and disease recovery in this animal model.
Animals treated prophylactically (day 0) or postexposure (day 3) were protected from observable illness and death. None of the animals in either of these groups presented visible symptoms, including rash development. Instead, they appeared healthy, remained active, and maintained or increased body weight. While on drug treatment, viral DNA was largely undetected in blood, and little to no viral shedding was detected in ocular and anal secretions. Oral secretions yielded the greatest numbers of infectious virus, with peak concentrations measuring 2 to 3 logs less than those of untreated and therapeutically treated animals. Ocular and anal secretions yielded lower or undetectable concentrations of virus, and detection was limited to fewer days postinoculation. These results could be considered consistent with the proposed mechanism that ST-246 inhibits production and release of EV from infected tissues. A reduction in EV, which is considered responsible for spread of the virus within a host, may have limited the infection to localized areas near the intranasal inoculation site, such as the nasal and oral cavities, thereby lessening the duration and morbidity of illness. When prophylactic or postexposure use of drug was discontinued, although animals remained asymptomatic, virus could be measured in secretions and excretions. Interestingly, trends in the range of days that infectious virus was detected differed between the two groups. Detection in animals treated prophylactically began near the end (day 12) of antiviral treatment, increased by 2 to 3 logs following treatment cessation, and persisted until day 24. Detection in animals treated postexposure was generally restricted to days 8 to 12, with the exception of one animal, PD7, which shed virus until day 26. While both prophylactic and postexposure treatments were successful for preventing illness, virus was not ultimately cleared by the end of treatment.
An unanticipated observation was the recovery of infectious virus from animals following cessation of drug exposure. One explanation could have been the presence of drug-resistant viruses. Highly drug-resistant CPXV variants, acquired through passaging of virus in the presence of drug, have been previously described from wild-type CPXV (27). We evaluated for the presence of drug resistance in select, twice-passaged virus isolates from prairie dog samples by a comet reduction assay. ST-246 antiviral activity could be demonstrated by reductions in plaque size and comet formation which are both produced by release of EV. The sensitivity of this assay was shown previously to be approximately equivalent to a standard EC50 assay (25). Additionally, isolates obtained postcessation or near the end of therapy, evaluated for ST-246 resistance without additional passaging, showed drug susceptibility similar to that of wild-type MPXV-ROC-358. Our results do not suggest that significant drug resistance was found in monkeypox viruses isolated postcessation of drug treatment. However, a prophylaxis, postexposure, or therapeutic regimen extending beyond 14 days, such as 21 days, may eliminate posttreatment viral shedding. This will need to be studied in the context of the development of an immunologic response to infection.
Comparison of antibody production between the three treatment groups suggests that timing of treatment initiation and the duration of treatment influence the humoral immune response. Prophylactic, postexposure, and therapeutic treatment resulted in three different antibody response profiles as measured by ELISA. Three of the prairie dogs treated therapeutically began producing measurable levels of Ig within 2 to 4 days following rash onset, and levels sharply increased to peak levels by 16 to 23 days post-rash onset. On the other hand, prairie dogs treated prophylactically produced measurable levels of Ig 4 to 10 days following rash onset, and peak levels were approximately half those of the therapeutic animals. Finally, prairie dogs treated postexposure produced measurable levels of Ig within 2 to 4 days post-rash, and peak levels were approximately a fourth of the levels of the therapeutic animals, with the exception of one animal. Differences in the immune profiles could be related to the availability of immunodominant EV epitopes, such as A33R and B5R; this has not yet been tested.
In several prairie dogs, clearance of infectious virus was temporally associated with antibody production. In the prophylactic group, PD11 and PD16 produced measurable levels of antibodies and eliminated infectious virus (oral swabs) 4 to 6 days earlier than PD10 and PD17. Similarly, in the postexposure group, PD1, PD4, and PD13 produced antibodies 6 to 8 days earlier than PD7 and cleared infectious virus from oral secretions by day 12. Antiorthopoxvirus antibodies were not measured in PD7 until day 20, and virus was not eliminated until day 26. Although cell-mediated responses were not addressed here, the temporal association of clearance of infectious virus and development of anti-orthopoxvirus antibodies suggests that the humoral immune response is important for virus clearance in the prairie dog model of MPXV infection.
Overall, prophylactic and postexposure ST-246 treatment prevented illness, and therapeutic ST-246 treatment prevented death in prairie dogs infected with a virulent strain of MPXV. We used the intranasal route of inoculation in order to more closely resemble a natural route of infection. Disease progression, including an asymptomatic period followed by systemic rash, closely resembled MPXV infection in humans, reaffirming the utility of the prairie dog system as a model for human systemic OPXV infection. These characteristics make the prairie dog a suitable model for investigating and identifying antiviral therapeutics and treatment regimens for monkeypox or smallpox in humans. Our results support future investigations with ST-246 to further characterize postexposure treatment modalities in the prairie dog model. These studies may be important for providing information that can be used for the identification of a safe and efficacious antiviral compound meeting the standards for federal approval.
ACKNOWLEDGMENTS
We thank Victoria A. Olson for discussions, critical analyses, and review of the manuscript and James Gathany for photographing the comet reduction and plaque assay.
For disclosure of competing financial interests, Robert Jordan and Dennis E. Hruby are employed by SIGA Technologies Inc., the company that produced ST-246.
The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention.
Footnotes
Published ahead of print on 22 June 2011.
REFERENCES
- 1. Berhanu A., et al. 2009. ST-246 inhibits in vivo poxvirus dissemination, virus shedding, and systemic disease manifestation. Antimicrob. Agents Chemother. 53:4999–5009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Breman J. G., Henderson D. A. 1998. Poxvirus dilemmas–monkeypox, smallpox, and biologic terrorism. N. Engl. J. Med. 339:556–559 [DOI] [PubMed] [Google Scholar]
- 3. Breman J. G., Henderson D. A. 2002. Diagnosis and management of smallpox. N. Engl. J. Med. 346:1300–1308 [DOI] [PubMed] [Google Scholar]
- 4. Centers for Disease Control and Prevention 2003. Update: multistate outbreak of monkeypox–Illinois, Indiana, Kansas, Missouri, Ohio, and Wisconsin, 2003. MMWR Morb. Mortal. Wkly. Rep. 52:642–646 [PubMed] [Google Scholar]
- 5. Chen Z., et al. 2006. Chimpanzee/human mAbs to vaccinia virus B5 protein neutralize vaccinia and smallpox viruses and protect mice against vaccinia virus. Proc. Natl. Acad. Sci. U. S. A. 103:1882–1887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Duraffour S., et al. 2007. Activity of the anti-orthopoxvirus compound ST-246 against vaccinia, cowpox and camelpox viruses in cell monolayers and organotypic raft cultures. Antivir. Ther. 12:1205–1216 [PubMed] [Google Scholar]
- 7. Emerson G. L., et al. 2009. The phylogenetics and ecology of the orthopoxviruses endemic to North America. PLoS One 4:e7666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Huggins J., et al. 2009. Nonhuman primates are protected from smallpox virus or monkeypox virus challenges by the antiviral drug ST-246. Antimicrob. Agents Chemother. 53:2620–2625 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Hutson C. L., et al. 2010. Dosage comparison of Congo Basin and West African strains of monkeypox virus using a prairie dog animal model of systemic orthopoxvirus disease. Virology 402:72–82 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Hutson C. L., Damon I. K. 2010. Monkeypox virus infections in small animal models for evaluation of anti-poxvirus agents. Viruses 2:2763–2776 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Hutson C. L., et al. 2007. Monkeypox zoonotic associations: insights from laboratory evaluation of animals associated with the multi-state us outbreak. Am. J. Trop. Med. Hyg. 76:757–768 [PubMed] [Google Scholar]
- 12. Hutson C. L., et al. 2009. A prairie dog animal model of systemic orthopoxvirus disease using West African and Congo Basin strains of monkeypox virus. J. Gen. Virol. 90:323–333 [DOI] [PubMed] [Google Scholar]
- 13. Jezek Z., Gromyko A. I., Szczeniowski M. V. 1983. Human monkeypox. J. Hyg. Epidemiol. Microbiol. Immunol. 27:13–28 [PubMed] [Google Scholar]
- 14. Li Y., Olson V. A., Laue T., Laker M. T., Damon I. K. 2006. Detection of monkeypox virus with real-time PCR assays. J. Clin. Virol. 36:194–203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Li Y., Zhao H., Wilkins K., Hughes C., Damon I. K. 2010. Real-time PCR assays for the specific detection of monkeypox virus West African and Congo Basin strain DNA. J. Virol. Methods 169:223–227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Likos A. M., et al. 2005. A tale of two clades: monkeypox viruses. J. Gen. Virol. 86:2661–2672 [DOI] [PubMed] [Google Scholar]
- 17. Massoudi M. S., Barker L., Schwartz B. 2003. Effectiveness of postexposure vaccination for the prevention of smallpox: results of a Delphi analysis. J. Infect. Dis. 188:973–976 [DOI] [PubMed] [Google Scholar]
- 18. Mortimer P. P. 2003. Can postexposure vaccination against smallpox succeed? Clin. Infect. Dis. 36:622–629 [DOI] [PubMed] [Google Scholar]
- 19. Nalca A., et al. 2008. Evaluation of orally delivered ST-246 as postexposure prophylactic and antiviral therapeutic in an aerosolized rabbitpox rabbit model. Antiviral Res. 79:121–127 [DOI] [PubMed] [Google Scholar]
- 20. Quenelle D. C., et al. 2007. Efficacy of delayed treatment with ST-246 given orally against systemic orthopoxvirus infections in mice. Antimicrob. Agents Chemother. 51:689–695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Reed K. D., et al. 2004. The detection of monkeypox in humans in the Western Hemisphere. N. Engl. J. Med. 350:342–350 [DOI] [PubMed] [Google Scholar]
- 22. Reynolds M. G., et al. 2006. Clinical manifestations of human monkeypox influenced by route of infection. J. Infect. Dis. 194:773–780 [DOI] [PubMed] [Google Scholar]
- 23. Rimoin A. W., et al. 2007. Endemic human monkeypox, Democratic Republic of Congo, 2001–2004. Emerg. Infect. Dis. 13:934–937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sbrana E., et al. 2007. Efficacy of the antipoxvirus compound ST-246 for treatment of severe orthopoxvirus infection. Am. J. Trop. Med. Hyg. 76:768–773 [PubMed] [Google Scholar]
- 25. Smith S. K., et al. 2009. In vitro efficacy of ST246 against smallpox and monkeypox. Antimicrob. Agents Chemother. 53:1007–1012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Xiao S. Y., et al. 2005. Experimental infection of prairie dogs with monkeypox virus. Emerg. Infect. Dis. 11:539–545 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Yang G., et al. 2005. An orally bioavailable antipoxvirus compound (ST-246) inhibits extracellular virus formation and protects mice from lethal orthopoxvirus challenge. J. Virol. 79:13139–13149 [DOI] [PMC free article] [PubMed] [Google Scholar]