

Abstract
The endocannabinoid system is implicated in a variety of physiological and pathological conditions (inflammation, immunomodulation, analgesia, cancer and others). The main active ingredient of cannabis, Δ9-tetrahydrocannabinol (Δ9-THC), produces its effects through activation of CB1 and CB2 receptors. CB1 receptors are expressed at high levels in the central nervous system (CNS), whereas CB2 receptors are concentrated predominantly, although not exclusively, in cells of the immune system. Endocannabinoids are endogenous lipid-signalling molecules that are generated in the cell membrane from phospholipid precursors. The two best characterized endocannabinoids identified to date are anandamide (AEA) and 2-arachidonoylglycerol (2-AG). Here we review the relationship between the endocannabinoid system and anti-tumour actions (inhibition of cell proliferation and migration, induction of apoptosis, reduction of tumour growth) of the cannabinoids in different types of cancer. This review will focus on examining how activation of the endocannabinoid system impacts breast, prostate and bone cancers in both in vitro and in vivo systems. The therapeutic potential of cannabinoids for cancer, as identified in clinical trials, is also discussed. Identification of safe and effective treatments to manage and improve cancer therapy is critical to improve quality of life and reduce unnecessary suffering in cancer patients. In this regard, cannabis-like compounds offer therapeutic potential for the treatment of breast, prostate and bone cancer in patients. Further basic research on anti-cancer properties of cannabinoids as well as clinical trials of cannabinoid therapeutic efficacy in breast, prostate and bone cancer is therefore warranted.
LINKED ARTICLES
This article is part of a themed issue on Cannabinoids in Biology and Medicine. To view the other articles in this issue visit http://dx.doi.org/10.1111/bph.2011.163.issue-7
Keywords: 2-arachidonoylglycerol, anandamide, bone cancer, breast cancer, cannabinoid, anti-cancer, clinical trial, endocannabinoid, prostate cancer, tumour growth, apoptosis, proliferation, migration
Introduction
Cancer resulted in approximately 7.6 million deaths worldwide in 2008 (Ferlay et al., 2010). An estimated 12.7 million new cancer cases were diagnosed in 2008 alone (Ferlay et al., 2010). Worldwide, an estimated 1.38 million women and 914 000 men were diagnosed with breast and prostate cancer, respectively, in 2008 (Ferlay et al., 2010). If this trend continues, cancer will overtake heart disease and become the predominant cause of death (Heron et al., 2009). Effective treatment and management of cancer is critical for cancer patients but the development of safe and effective treatments that improve cancer therapy remains an unmet need. Cannabinoids and modulators of the endocannabinoid system have recently been shown to produce anti-tumour actions (reduction of inflammation, cell proliferation and cell survival properties) in different models of cancer. The present review focuses on breast, prostate and bone cancer in which links to the endocannabinoid system have been studied. More work is necessary to determine whether pharmacotherapies targeting the endocannabinoid system improve the treatment of cancer in patients.
Cannabinoids are implicated in a variety of physiological and pathological conditions including inflammation, immunomodulation, analgesia, cancer and others (for reviews Pacher et al., 2006; Di Marzo, 2008). The main active ingredient of cannabis, Δ9-tetrahydrocannabinol (Δ9-THC), produces its effects through activation of G-protein-coupled CB1 (Matsuda et al., 1990; Ledent et al., 1999; Zimmer et al., 1999) and CB2 (Munro et al., 1993; Buckley et al., 2000) receptors. Endocannabinoids are endogenous lipid-signalling molecules that are generated in the cell membrane from phospholipid precursors. They bind and activate one or more cannabinoid receptor subtypes, thus producing cannabimimetic properties (for reviews Piomelli, 2005; Di Marzo, 2006). The two best studied endocannabinoids isolated to date are anandamide (AEA) and 2-arachidonoylglycerol (2-AG). Anandamide is hydrolysed by the enzyme fatty-acid amide hydrolase (FAAH) whereas 2-AG is degraded predominantly, although not exclusively, by monoacylglycerol lipase (MGL) (for reviews Pacher et al., 2006; Jhaveri et al., 2007; Guindon and Hohmann, 2009; Wang and Ueda, 2009). Thus, enzymes catalysing endocannabinoid hydrolysis represent potential new targets for cancer pharmacotherapies. In this review, drug and molecular target nomenclature conforming to British Journal of Pharmacology Guide to Receptors and Channels (Alexander et al., 2009) has been employed. Here, we will refer to effects of cannabinoids blocked by CB1 antagonists (SR141716A, AM251) as being CB1-mediated and effects blocked by CB2 antagonists (SR144528, AM630) as CB2-mediated.
Cancer is marked by uncontrolled cell division and cell death emerging from cumulative damage of important regulatory genes. Multiple genes likely need to be damaged in order for a cancer to grow and develop the ability to spread (i.e. metastasize). Cancers may be hereditary and/or caused by external (tobacco, chemicals, radiation, infectious organisms and others) and/or internal (inherited mutations, hormones, mutations from metabolism and others) factors. This review will focus on uncovering connections between the endocannabinoid system and breast, prostate and bone cancers with an emphasis on understanding how these connections could be exploited for their therapeutic anti-cancer potential.
Animal models of cancer have been developed to experimentally assess pathophysiological mechanisms implicated in the analogous clinical syndrome. These models provide insight into pathophysiological processes of the disease state and elucidate mechanisms of action that may be targeted by drug discovery efforts aimed at identifying novel therapeutics. These models thus permit preclinical evaluation and validation of therapeutic efficacy of new pharmacotherapies.
Anti-proliferative properties of cannabis compounds were first identified 35 years ago. Here, it was first shown that administration of Δ9-THC inhibits lung adenocarcinoma cell growth in vivo (i.e. after oral administration in mice) and in vitro (Munson et al., 1975; Carchman et al., 1976). It took more that two decades before the potential anti-tumour properties associated with a cannabinoid were investigated further. In the past 14 years, an emerging body of research, primarily employing in vitro models of different cancers, has helped elucidate the mechanisms through which cannabinoids and the endocannabinoid tumour system influences cancer cell proliferation, migration and apoptosis (i.e. programmed cell death).
The mechanisms through which cannabinoids/cannabinoids receptors impact proliferation, migration and apoptosis of cancer cells are quite complex and our understanding of these processes remain incomplete. Moreover, these mechanisms differ in different types of cancer, and both pro- and anti-apoptotic effects of cannabinoids have been reported. A schematic representation of the major signalling pathways that are implicated in the activation of different cannabinoid receptor subtypes through their agonists and their involvement in these processes is summarized in Figure 1.
Figure 1.
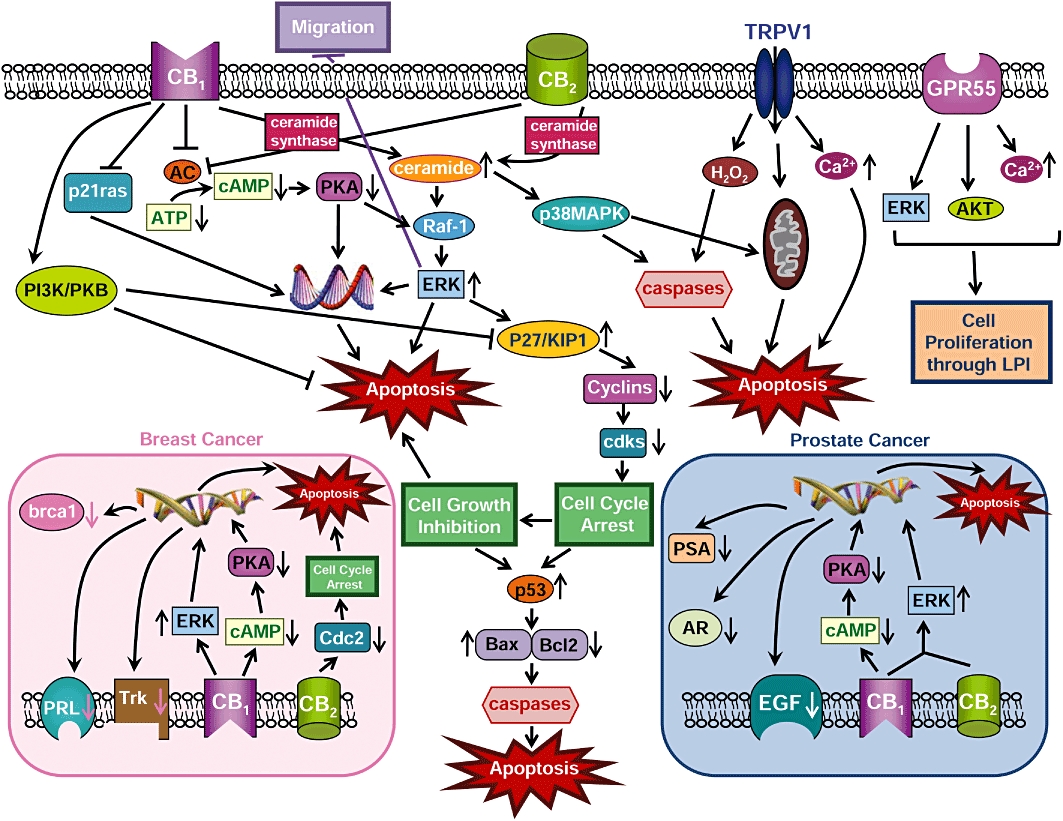
Schematic representation of different mechanisms/signalling pathways through which cannabinoids impact apoptosis, proliferation and migration. AC, adenylyl cyclase; AKT, protein kinase B; AR, androgen receptor; ATP, adenosine triphosphate; Bax, pro-apoptotic protein; Bcl2, antiapoptotic protein; brca1, breast cancer susceptibility gene product; cAMP, cyclic adenosine monophosphate; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; Cdc2, p34 cyclin-dependent kinase 1; CDK, cyclin-dependent kinases; EGF, epidermal growth factor; ERK, extracellular regulated kinase; GPR55, G protein-coupled receptor 55; H2O2, hydrogen peroxide; p27/KIP1, cyclin kinase inhibitor; PRL, prolactin receptor; p53, p53 protein; p21ras, p21 ras protein; PI3K, phosphatidyl inositol 3 kinase; PKA, protein kinase A; PKB, protein kinase B; PSA, prostate-specific antigen; Raf-1, protein Raf-1; Trk, high-affinity nerve growth factor receptor; TRPV1, transient receptor potential cation channel V1.
Several mechanisms are likely to underline the pro-apoptotic effects of cannabinoids and explain their anti-cancer effects. Cannabinoids induce de novo synthesis of ceramides, a family of lipid molecules composed of sphingosine and a fatty acid, found in the cell membrane. Synthesis of ceramide occurs via activation of the enzyme ceramide synthase and leads to downstream activation of an extracellular regulated kinase (ERK) signalling cascade. This process results in cell cycle arrest and apoptosis. Activation of either CB1 or CB2 receptors triggers the ceramide-ERK signalling pathway to promote apoptosis (Kogan, 2005; Sarfaraz et al., 2006, 2008) (Figure 1). The increase in ceramide can also activate the p38 mitogen-activated protein kinase (p38MAPK) pathway which can lead to apoptosis through multiple mechanisms (i.e. through activation of cysteine proteases (i.e. caspases) or through cytochrome C release from mitochondria). The sustained activation of ERK also promotes the induction of cyclin kinase inhibitor (p27/KIP1) which modulates regulatory molecules of the cell cycle (cyclins, cdks) resulting in cell cycle arrest and apoptosis (Kogan, 2005; Sarfaraz et al., 2006, 2008) (Figure 1). Cell cycle arrest involves the up-regulation of the p53 protein which will differentially alter levels of pro- and anti-apoptotic proteins (i.e. increase the levels of the pro-apoptotic protein Bax and lower the levels of the anti-apoptotic protein Bcl2, respectively, thereby shifting the ratio towards Bax) which ultimately leads to activation of caspases that play an essential role in triggering apoptosis (Sarfaraz et al., 2006). Activation of either CB1 or CB2 receptors also inhibits adenylyl cyclase (AC) activity and lowers both cyclic adenosine monophosphate (cAMP) levels and protein kinase A (PKA) activity, thereby causing down-regulation of gene transcription, leading to apoptosis (Guzmán, 2003; Kogan, 2005; Bifulco et al., 2008; Sarfaraz et al., 2008). Activation of transient receptor potential cation channel V1 (TRPV1) receptors also leads to increases in intracellular levels of both hydrogen peroxide (H2O2) and/or calcium or the release of cytochrome C from mitochondria, causing apoptosis through both distinct and overlapping mechanisms (Figure 1) (Maccarrone et al., 2000).
Cannabinoid receptor 1 ligands have pro-apoptotic properties, in part, through inhibition of the Ras protein (p21ras) which is involved in inducing DNA synthesis (Bifulco et al., 2001). However, CB1 receptor activation can also trigger activation of different tumour cascades that are linked to promotion of cancer cell survival and inhibition of apoptosis. Indeed, CB1 ligands also stimulate the PI3K/PKB tumour pathway, an essential tumour pathway implicated in cell survival. Activation of PI3K/PKB downstream of CB1 activation may promote apoptosis either directly or through inhibition of p27/KIP1 (Gómez del Pulgar et al., 2000; Sarfaraz et al., 2008). Thus, both pro- and anti-apoptotic effects of cannabinoids have been reported, raising the possibility that a dysregulation of the endocannabinoid system may also contribute to cancer/tumour development. However, activation of ERK downstream of cannabinoid receptor activation also promotes anti-cancer effects through inhibition of cancer cell migration (Blazquez et al., 2003; Kogan, 2005). More work is necessary to determine the balance between these various mechanisms, and how they modulate cancer in vivo.
Phytocannabinoids may also interfere with the ability of lysophosphatidylinositol (LPI), a putative endogenous ligand for G-protein-coupled receptor 55 (GPR55), to promote cancer cell proliferation through activation of the putative novel cannabinoid receptor subtype (Figure 1). Pretreatment of breast and prostate cancer cells with cannabidiol, a major component of marijuana, or with SR141716A (a CB1 antagonist that also possesses agonist/antagonist properties at GPR55), blocks the ability of LPI to induce cell proliferation through GPR55, thereby producing anti-cancer effects (Piñeiro et al., 2011). Activation of GPR55 by LPI promotes cancer cell proliferation following activation of distinct intracellular tumour cascades [i.e. ERK, AKT and calcium mobilization (Piñeiro et al., 2011)]; these anti-proliferative effects were also blocked using siRNA for GPR55.
Breast cancer
Breast cancer is the most common cancer among women (Ferlay et al., 2010). Genetics, lack of child bearing/breastfeeding, higher hormone levels and iodine deficiency have all been identified as risk factors for developing breast cancer (Madigan et al., 1995). Breast cancer, also known as malignant breast neoplasm, originates from breast tissue, usually from the milk ducts (ductal carcinomas) or the lobules (lobular carcinomas) that supply the ducts with milk (Glass et al., 2007). Breast cancer cells may spread to other organs such as the bones, lungs and lymph nodes (Guise et al., 2010; Pantel and Alix-Panabières, 2010). Recent research identifies a role for the endocannabinoid system in the regulation of tumour growth, induction of apoptosis (programmed cell death) and control of tumour vascularization (angiogenesis) in breast cancer (Grimaldi et al., 2006; Ligresti et al., 2006; Qamri et al., 2009; Caffarel et al., 2010). The phenomenon of angiogenesis is required for tumours to transition from a dormant to malignant state (Fontanini, 2000; Ribatti and Crivellato, 2010).
Expression of cannabinoid receptors in different breast cancer tissue/cell lines has been described. CB1 expression by immunohistochemistry was detected in 14% of human breast cancer tumour tissue expressing a member of the epidermal growth factor (EGF) family referred to as the ErbB2 tyrosine kinase receptor. No correlation between CB1 expression and ErbB2 expression was found (Caffarel et al., 2010). CB1 immunoreactivity was also expressed in 28% of human breast carcinoma (Qamri et al., 2009). CB1 receptors are also present in different breast cancer cell lines (MCF-7, T-47D, MDA-MB-231, TSA-E1, MDA-MB-468) and in human breast tissues using RT-PCR, immunofluorescence and/or Western blot (Melck et al., 2000; Di Marzo et al., 2001; McKallip et al., 2005; Sarnataro et al., 2005, 2006; Caffarel et al., 2006; Grimaldi et al., 2006; Ligresti et al., 2006; Qamri et al., 2009). By contrast, CB2 immunoreactivity was detected in 72% of human breast tumour tissue (Caffarel et al., 2010). Interestingly, CB2 receptors were found in 91% of ErbB2-positive tumour tissue, suggesting a link between CB2 and ErbB2-expression, but not between CB1 and ErbB2-expression (Caffarel et al., 2010). In another study, CB2 receptors immunoreactivity was observed in 35% of human breast carcinoma (Qamri et al., 2009). However, CB2 receptors are also expressed in different breast carcinoma cell lines (MCF-7, T-47D, MDA-MB-231, MDA-MB-468, EVSA-T, SkBr3) and human breast tissues using RT-PCR, immunofluorescence and/or Western blot (Melck et al., 2000; Di Marzo et al., 2001; McKallip et al., 2005; Ligresti et al., 2006; Qamri et al., 2009; Caffarel et al., 2006, 2010). The putative novel cannabinoid receptor subtype GPR55 was highly expressed in a MDA-MB-231 cell line using RT-PCR, but it is expressed at lower (30-fold) levels in MCF-7 breast carcinoma cell lines using RT-PCR (Ford et al., 2010). Lysophosphatidylinositol (LPI), the putative endogenous ligand for GPR55, also stimulates cell migration and invasion in a MDA-MB-231 cell line and this LPI effect on migration is blocked by pretreatment with cannabidiol (CBD) (Ford et al., 2010). Furthermore, it was also demonstrated that LPI stimulate proliferation and this effect was blocked by CBD (Piñeiro et al., 2011). Moreover, the anandamide hydrolysing enzyme (FAAH) is found in EFM-19 and MCF-7 cancer cell lines using Northern blot analyses (Bisogno et al., 1998) or RT-PCR (Takeda et al., 2008). No study has evaluated whether the 2-AG hydrolysing enzyme MGL is similarly present, or whether changes in enzymes catalysing endocannabinoid synthesis or degradation accompany anti-cancer effects. Nonetheless, the literature suggests that multiple cannabinoid receptor subtypes as well as enzymes catalysing endocannabinoid hydrolysis (i.e. FAAH) show an anatomical distribution appropriate to regulate breast cancer cell proliferation, migration and/or apoptosis.
In vitro studies show that endocannabinoids and cannabinoid-like compounds inhibit proliferation and/or migration and/or induce apoptosis in different breast carcinoma (MCF-7, EFM-19, T-47D, MDA-MB-231, MDA-MB-468, MDA-MB-436, 4T1, TSA-E1, EVSA-T, SkBr3, HTB-126) cell lines. It is important to note that the MDA-MB-231 cell line represents a small proportion of all types of breast cancer; it is highly metastatic and lacks expression of other receptors (oestrogen, progesterone). The phytocannabinoid CBD inhibits cell proliferation (Ruh et al., 1997; Ligresti et al., 2006; McAllister et al., 2007, 2010), increases apoptosis (Ligresti et al., 2006) and reduces migration (McAllister et al., 2007, 2010; Ford et al., 2010) in different cancer cell lines (Table 1). Anti-proliferative effects of CBD are partially reversed by SR144528 (Ligresti et al., 2006), although a possible role for CB1 cannot be ruled out because its possible contribution was not evaluated. The mechanism of action of CBD in producing these effects is not fully understood and needs to be investigated further. Δ9-THC also possesses anti-proliferative properties (McAllister et al., 2007; Caffarel et al., 2006, 2008; von Bueren et al., 2008), increases apoptosis (Caffarel et al., 2006) and decreases cancer cell migration (McAllister et al., 2007). Δ9-THC-induced anti-proliferative and pro-apoptotic properties are mediated by CB2, but not CB1 receptors (Caffarel et al., 2006). The lack of effect of Δ9-THC on CB1 receptors in EVSA-T cells could be explained by the lack of CB1 receptors in this cell line. By contrast, some studies have shown that Δ9-THC failed to inhibit cell proliferation (Ruh et al., 1997) or induce apoptosis (McKallip et al., 2005). Cannabinol (CBN), a phytocannabinoid that is both a metabolite of Δ9-THC and a weak agonist at CB1 and CB2 receptors, also inhibits cell proliferation (McAllister et al., 2007). Another study provided findings contradictory to these observations; CBD, Δ9-THC and CBN all stimulated proliferation at low (1 µM) or high (up to 20 µM) concentrations in MCF-7 cells, but there were limitations in the studies which affect data interpretation. For example, these groups failed to demonstrate the expression of CB1 or CB2 receptors (Watanabe et al., 2005; Takeda et al., 2008), a finding which conflicts with other studies that show CB1 and CB2 receptor expression in the same cell line (Movsesyan et al., 2004; McKallip et al., 2005). Nonetheless, the possibility remains that the combination of phytocannabinoids in cannabis may offer greater therapeutic potential compared with Δ9-THC or CBD alone.
Table 1.
Breast cancer in vitro and ex vivo studies
| Apoptosis/proliferation/migration mediated by | |||||||
|---|---|---|---|---|---|---|---|
| Cell line/tissue | Expressing | Apoptosis | Proliferation | Migration | CB1 | CB2 | Reference |
| MCF-7 | – | – | ↓ by DALN, CBD | – | – | – | Ruh et al., 1997 |
| EFM-19 | FAAH | – | ↓ by AEA | – | SR1 | – | Bisogno et al., 1998 |
| EFM-19 | FAAH | – | ↓ by Oleamide | – | SR1 | – | Bisogno et al., 1998 |
| MCF-7 | – | – | ↓ by AEA | – | SR1 | – | De Petrocellis et al., 1998 |
| EFM-19 | |||||||
| EFM-19 | – | – | ↓ by 2-AG, MET, HU-210 | – | – | – | De Petrocellis et al., 1998 |
| MCF-7 | – | – | ↓ by AEA | – | – | – | Melck et al., 1999 |
| EFM-19 | |||||||
| MCF-7 | CB1, CB2 | – | ↓ by AEA, arvanil | – | SR1 | Not SR2 | Melck et al., 2000 |
| T-47D | |||||||
| MCF-7 | CB1, CB2 | – | ↓ by 2-AG | – | – | – | Melck et al., 2000 |
| T-47D | |||||||
| MCF-7 | CB1, CB2 | – | PEA enhance AEA ↓ | – | – | Not SR2 | Di Marzo et al., 2001 |
| MCF-7 | CB1, CB2 | – | PEA enhance arvanil ↓ PEA enhance HU-210 ↓ | – | – | – | Di Marzo et al., 2001 |
| EFM-19 | – | – | PEA enhance olvanil ↓ | – | – | Not SR2 | De Petrocellis et al., 2002 |
| MCF-7 | CB1 low, not CB2 | Δ9-THC no Δ | – | – | – | – | McKallip et al., 2005 |
| 4T1* | Not CB1 nor CB2 | Δ9-THC no Δ | – | – | – | – | McKallip et al., 2005 |
| MDA-MB-231 | CB1 | – | – | – | – | – | Sarnataro et al., 2005 |
| MCF-7 | – | – | ↑ by Δ9-THC, CBD, CBN | – | – | – | Watanabe et al., 2005 |
| EVSA-T | CB2 | ↑ by Δ9-THC | ↓ by Δ9-THC | – | Not SR1 pro | SR2 pro, apo | Caffarel et al., 2006 |
| MCF-7 | – | – | ↓ by Δ9-THC | – | – | – | Caffarel et al., 2006 |
| T-47D | |||||||
| MDA-MB-231 | |||||||
| MDA-MB-468 | |||||||
| SkBr3 | |||||||
| Breast tumour | CB1, CB2 | – | – | – | – | – | Caffarel et al., 2006 |
| MDA-MB-231 | – | MET no Δ | ↓ by MET | ↓ by MET | SR1 mig | – | Grimaldi et al., 2006 |
| T-47D | CB1 | – | MET no Δ | – | – | – | Grimaldi et al., 2006 |
| TSA-E1* | CB1 | – | ↓ by MET | ↓ by MET | SR1 mig | – | Grimaldi et al., 2006 |
| MCF-7 | CB1 weak, CB2 weak | – | ↓ by CBD, CBG, CBC, CBD acid, THC acid | – | – | – | Ligresti et al., 2006 |
| MDA-MB-231 | CB1 weak, CB2 medium | ↑ by CBD | ↓ by CBD, CBG, CBC, CBD acid, THC acid | – | – | Partially by SR2 with CBD for pro | Ligresti et al., 2006 |
| MCF-7 | CB1 | – | ↓ by SR1 | – | – | – | Sarnataro et al., 2006 |
| T-47D | |||||||
| MDA-MB-231 | CB1 | SR1 no Δ | ↓ by SR1 | – | – | – | Sarnataro et al., 2006 |
| MDA-MB-231 | – | – | ↓ by Δ9-THC, CBN, WIN-2, CP55,940, CBD | ↓ by Δ9-THC, CBD, WIN-2 | – | – | McAllister et al., 2007 |
| MDA-MB-436 | – | – | ↓ by Δ9-THC, CBN, WIN-2, CP55,940, CBD | ↓ by CBD | – | – | McAllister et al., 2007 |
| HTB-126 | – | – | ↓ by NPT, NPD | – | SR1 with NPT | – | Burstein and Salmonsen, 2008 |
| EVSA-T | – | – | ↓ by Δ9-THC | – | – | – | Caffarel et al., 2008 |
| MDA-MB-231 | – | – | – | ↓ by MET | SR1 | – | Laezza et al., 2008 |
| MCF-7 | FAAH | – | ↑ by Δ9-THC, CBD, CBN | – | Not SR1 nor AM251 with Δ9-THC | – | Takeda et al., 2008 |
| Not CB1 nor CB2 | |||||||
| MCF7 | – | – | ↓ by Δ9-THC | – | – | – | von Bueren et al., 2008 |
| MCF7-AR1 | |||||||
| MDA-MB-231 | CB1, CB2 | ↑ by JWH-133, WIN-2 | ↓ by JWH-133, WIN-2 | ↓ by JWH-133, WIN-2 | – | siRNA and WB | Qamri et al., 2009 |
| MDA-MB-468 | CB1, CB2 | – | ↓ by JWH-133, WIN-2 | ↓ by JWH-133, WIN-2 | – | – | Qamri et al., 2009 |
| MCF-7 | – | – | ↑ by Δ9-THC | – | – | – | Takeda et al., 2009 |
| Breast tumour | CB1, CB2 | – | – | – | – | – | Caffarel et al., 2010 |
| MDA-MB-231 | GPR55 | – | – | ↓ by CBD | – | – | Ford et al., 2010 |
| MCF-7 | – | – | ↓ by MET | – | – | – | Laezza et al., 2010 |
| MDA-MB-231 | |||||||
| MCF-7 | – | – | ↓ by CBD | ↓ by CBD | – | – | McAllister et al., 2010 |
| MDA-MB-231 | |||||||
Human and murine* cell line/tissue.
↑, increase; ↓, decrease; –, not tested; 2-AG, 2-arachidonoyl glycerol; apo, apoptosis; AEA, anandamide; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; CBC, cannabichromene; CBD, cannabidiol; CBD acid; cannabidiol acid; CBG, cannabigerol; CBN, cannabinol; Δ9-THC, delta 9-tetrahydrocannabinol; DALN, desacetyllevonantradol; FAAH, fatty-acid amide hydrolase; MET, methanandamide; mig, migration; no Δ, no change; NPD, N-palmitoyl dopamine; NPT, N-palmitoyl tyrosine; PEA, palmitoylethanolamide; pro, proliferation; siRNA, silencing RNA; SR1, SR141716A; SR2, SR144528; THC acid, tetrahydrocannabinol acid; WB, Western blot; WIN-2, WIN55,212-2.
Endocannabinoids exhibit anti-proliferative properties in vitro. Indeed, AEA inhibits cell proliferation (Bisogno et al., 1998; De Petrocellis et al., 1998; Melck et al., 1999, 2000) which is mediated by CB1 (Bisogno et al., 1998; De Petrocellis et al., 1998; Melck et al., 2000), but not by CB2 (Melck et al., 2000) receptors (Table 1). However, CB2-mediated effects of AEA cannot be discounted because only one study evaluated their possible contribution (Melck et al., 2000). Moreover, 2-AG, oleamide and arvanil all inhibit cell proliferation. The anti-proliferative effects of oleamide and arvanil are inhibited by CB1 (Bisogno et al., 1998; Melck et al., 2000), but not CB2 (Melck et al., 2000) receptor antagonists. Interestingly, palmitoylethanolamide (PEA), a fatty-acid amide that does not bind to cannabinoid receptors, enhances the anti-proliferative effects of AEA, arvanil, olvanil and HU-210 (Di Marzo et al., 2001; De Petrocellis et al., 2002), which is suggestive of synergism or an entourage effect (Ben-Shabat et al., 1998; De Petrocellis et al., 2002). Moreover, the enhancement of AEA and olvanil anti-proliferative effects does not involve CB2 receptors (Di Marzo et al., 2001; De Petrocellis et al., 2002). Methanandamide inhibits both cell proliferation (De Petrocellis et al., 1998; Grimaldi et al., 2006; Laezza et al., 2010) and cell migration (Grimaldi et al., 2006; Laezza et al., 2008). The methanandamide-induced inhibition of cell migration involves CB1 receptors (Grimaldi et al., 2006). By contrast, the mixed CB1/CB2 agonist WIN55,212-2 and CB2 agonist JWH-133 inhibits both cell proliferation (McAllister et al., 2007; Qamri et al., 2009) and migration (McAllister et al., 2007; Qamri et al., 2009) through a mechanism that requires CB2 receptor activation, although a possible role for CB1 was not assessed (Qamri et al., 2009) (Table 1). Surprisingly, both cannabinoid agonists such as CP55 940 (McAllister et al., 2007) and HU-210 (De Petrocellis et al., 1998) and cannabinoid CB1 antagonists such as SR141716A (Sarnataro et al., 2006) have all been shown to inhibit cell proliferation. Amide derivatives such as N-palmitoyl tyrosine and N-palmitoyl dopamine also possess anti-proliferative properties; in the case of N-palmitoyl tyrosine, effects are mediated by CB1 receptors (Burstein and Salmonsen, 2008). Cannabigerol, cannabichromene, cannabidiol acid and THC acid (Ligresti et al., 2006) as well as desacetyllevonantradol (Ruh et al., 1997) also inhibit cell proliferation in different breast cancer cell lines (Table 1).
In breast cancer cells, cannabinoid receptor agonists inhibit breast cancer cell proliferation by a down-regulation of high-affinity nerve growth factor (Trk) and prolactin (PRL) receptors as well as down-regulation of breast cancer susceptibility gene product (brca1) through the cAMP-PKA/MAPK/Raf-ERK signalling pathways (De Petrocellis et al., 1998; Melck et al., 1999, 2000; Bifulco et al., 2008) (Figure 1, left inset). Furthermore, it has been demonstrated that Δ9-THC decreased the levels of Cdc2 (major cyclin dependent kinase controlling the entrance of cells in mitosis), thereby causing cell cycle arrest and subsequent apoptosis (Caffarel et al., 2006) (Figure 1, left inset). Other signalling pathways could be involved or discovered in future studies.
In vivo studies demonstrate that cannabinoids reduce tumour growth and metastasis as well as cell proliferation and angiogenesis in mice injected with different breast cancer cell lines. For example, Δ9-THC decreases tumour size as well as the number of tumour and lung metastases and inhibits both cell proliferation and angiogenesis in an engineered animal model of ErbB2 (tyrosine kinase receptor)-driven metastatic breast cancer (Caffarel et al., 2010) (see Table 2 for more details). This inhibition of cell proliferation involves CB2, but not CB1 receptors (Caffarel et al., 2010). Further evidence for a role for CB2 receptors in these anti-cancer properties is based upon the ability of the CB2 agonist JWH-133 to decrease size and number of tumours, reduce the number and size of lung metastases, inhibit cell proliferation and decrease angiogenesis in mice injected with different breast cancer cell lines (Qamri et al., 2009; Caffarel et al., 2010). These effects were mediated by CB2, but not CB1 receptors (Qamri et al., 2009; Caffarel et al., 2010). In CB-17 immunodeficient mice injected with MDA-MB-231 cells, the mixed CB1/CB2 agonist WIN55,212-2 also reduces tumour size, decreases the number and size of lung metastases, inhibits proliferation and reduces angiogenesis; these effects were mediated by CB1 and CB2 receptors (Qamri et al., 2009) (Table 2). Furthermore, the phytocannabinoid cannabidiol (CBD) also reduces tumour growth (size) and decreases the number of lung metastases in mice injected with MDA-MB-231(Ligresti et al., 2006) or 4T1 (McAllister et al., 2010) breast cancer cell lines (see Table 2). Moreover, the anandamide analogue methanandamide also reduces the number and size of lung tumour nodules in mice injected with TSA-1 mammary carcinoma cell line through a CB1 receptor mechanism (Grimaldi et al., 2006) (Table 2). Strikingly, the CB1 antagonist SR141716A, administered alone, has also been reported to decrease tumour size in mice injected with MDA-MB-231 cancer cells (Sarnataro et al., 2006); more work is necessary to determine whether effects of SR141716A observed here can be attributed to direct activation of CB1 receptors or to other receptor mechanisms (e.g. GPR55). However, conflicting data are reported in the literature in this regard because systemic administration of Δ9-THC has been reported to increase the local tumour size and the number/size of metastasis in mice injected with 4T1 tumour cells into the rear foot-pads (McKallip et al., 2005). These unusual findings are potentially explained by the fact that Δ9-THC suppresses the anti-tumour immune response which involves CB2, but not the CB1 receptors (McKallip et al., 2005). Moreover, SCID-NOD mice, which are devoid of anti-tumour immune responses, do not exhibit increases in tumour size or metastasis following Δ9-THC administration (McKallip et al., 2005). None of these models of breast cancer is actually injecting tumour cells into the milk ducts or lobules of the breast which diminishes the translation relevance to human breast cancer and reinforces the need to develop new animal models that better reproduce the disease state.
Table 2.
Breast cancer in vivo studies
| Tumour | Metastasis | Proliferation (pro)/apoptosis (apo)/angiogenesis (ang) | Mediated by | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mouse strain | Tumour induction (cell line) | Drugs | Dose and route | Size | Number | Size | Number | CB1 | CB2 | Reference | |
| BALB/c | 4T1 paw | Δ9-THC | 12.5–50 mg·kg−1 i.p. | ↑ by Δ9-THC | – | ↑ by Δ9-THC | ↑ by Δ9-THC | – | – | – | McKallip et al., 2005 |
| BALB/c | 4T1 paw | Δ9-THC | 25–50 mg·kg−1 i.p. | ↑ by Δ9-THC | – | – | ↑ by Δ9-THC | – | – | – | McKallip et al., 2005 |
| SCID-NOD | 4T1 paw | Δ9-THC | 25 mg·kg−1 i.p. | Δ9-THC no Δ | – | – | Δ9-THC no Δ | – | – | – | McKallip et al., 2005 |
| C57BL/6N | TSA-E1 paw | MET | 0.5 mg·kg−1 i.p. | – | – | ↓ by MET | ↓ by MET | – | SR1 | – | Grimaldi et al., 2006 |
| Athymic | MDA-MB-231 dorsal side | CBD | 5–6.5 mg·kg−1 s.c. | ↓ by CBD | – | – | – | – | – | – | Ligresti et al., 2006 |
| BalB/c | MDA-MB-231 paw | CBD | 5–6.5 mg·kg−1 s.c. | – | – | – | ↓ by CBD | – | – | – | Ligresti et al., 2006 |
| CD1 | MDA-MB-231 flank | SR1 | 0.7 mg·kg−1 s.c. | ↓ by SR1 | – | – | – | – | – | – | Sarnataro et al., 2006 |
| CB-17 immuno | MDA-MB-231 flank | JWH-133 | 5 mg·kg−1 p.t. | ↓ by JWH-133 | – | – | – | JWH-133 ↓ pro, ↓ ang | – | SR2 | Qamri et al., 2009 |
| CB-17 immuno | MDA-MB-231 flank | WIN-2 | 5 mg·kg−1 p.t. | ↓ by WIN-2 | – | – | – | WIN-2 ↓ pro, ↓ ang | AM251 | SR2 | Qamri et al., 2009 |
| CB-17 immuno | MDA-MB231-luc-D3H2LN lateral vein | JWH-133 | 5 mg·kg−1 i.p. | – | – | ↓ by JWH-133 | ↓ by JWH-133 | – | – | SR2 | Qamri et al., 2009 |
| CB-17 immuno | MDA-MB231-luc-D3H2LN lateral vein | JWH-133, WIN-2 | 5 mg·kg−1 i.p. | – | – | ↓ by WIN-2 | ↓ by WIN-2 | – | AM251 | SR2 | Qamri et al., 2009 |
| PyMT | NA | JWH-133 | 5 mg·kg−1 i.p. | ↓ by JWH-133 | – | – | – | JWH-133 ↓ pro, ↓ ang | – | – | Qamri et al., 2009 |
| MMTV-neu | NA | Δ9-THC | 0.5 mg p.t. | ↓ by Δ9-THC | ↓ by Δ9-THC | – | ↓ by Δ9-THC | Δ9-THC ↓ pro, ↑ apo, ↓ ang | Not SR1 pro | SR2 pro | Caffarel et al., 2010 |
| MMTV-neu | NA | JWH-133 | 0.05 mg p.t. | ↓ by JWH-133 | ↓ by JWH-133 | ↓ by JWH-133 | – | JWH-133 ↓ pro, ↑ apo, ↓ ang | Not SR1 pro | SR2 pro | Caffarel et al., 2010 |
| BalB/c | 4T1 fourth major nipple | CBD | 1–5 mg·kg−1 s.c. | ↓ by CBD | – | ↓ by CBD | ↓ by CBD | – | – | – | McAllister et al., 2010 |
↑, increase; ↓, decrease; –, not tested; ang, angiogenesis; apo, apoptosis; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; CBD, cannabidiol; Δ9-THC, deltat 9-tetrahydrocannabinol; immuno, immunodeficient; i.p., intraperitoneal; MET, methanandamide; MMTV-neu mice, genetically engineered mice of ErbB2 (tyrosine kinase receptor)-driven metastatic breast cancer; NA, not applicable; no Δ, no change; pro, proliferation; p.t., peritumoral; PyMT mice, transgenic mice developing mammary gland tumours; s.c., subcutaneous; SCID-NOD, devoid of anti-tumour immune response; SR1, SR141716A; SR2, SR144528; WIN-2, WIN55,212-2.
Prostate cancer
Prostate cancer is the second most frequently diagnosed malignancy in men (Bray et al., 2010; Ferlay et al., 2010). Genetics, diet, medical exposure and viral infection are all factors implicated in occurrence or incidence of the disease (Djulbegovic et al., 2010). Prostate cancer develops in the prostate, a gland in the male reproductive system. Most prostate cancers grow slowly although cases of aggressive prostate cancers are also observed. These cancer cells may metastasize from the prostate to other parts of the body, particularly to the bones and lymph nodes (Parkin et al., 2001; Schroder et al., 2009; Djulbegovic et al., 2010).
Several studies have also evaluated the expression of cannabinoid receptors in different prostate cancer tissue/cell lines. Indeed, it was shown that high CB1 receptor immunoreactivity score in prostate cancer tissue is associated with prostate cancer severity and outcome (Chung et al., 2009). It was also demonstrated that CB1 expression is up-regulated in prostate cancer tissue (Czifra et al., 2009). Moreover, multiple prostate cancer cell lines (i.e. PC-3, DU-145, LNCaP, CWR22Rv1, CA-HPV-10) and human prostate cancer tissues express CB1 receptors using RT-PCR, immunofluorescence, and Western blot (Ruiz et al., 1999; Melck et al., 2000; Sánchez et al., 2003a; Nithipatikom et al., 2004; Sarfaraz et al., 2005; Chung et al., 2009; Czifra et al., 2009; Brown et al., 2010). CB2 receptors are also expressed in different prostate cancer cell lines (PC-3, DU-145, LNCaP, CWR22Rv1, CA-HPV-10) using RT-PCR, immunofluorescence and Western blot (Melck et al., 2000; Sánchez et al., 2003a; Nithipatikom et al., 2004; Sarfaraz et al., 2005; Brown et al., 2010). Moreover, expression of FAAH is demonstrated in prostate cancer cell lines (PC-3, DU-145, LNCaP) and human prostate cancer tissue using Western blot, immunohistochemistry and RT-PCR (Ruiz-Llorente et al., 2004; Endsley et al., 2008; Takeda et al., 2008; Brown et al., 2010; Thors et al., 2010; Wang et al., 2008) (Table 3). Furthermore, the putative cannabinoid receptor GPR55 is also expressed in PC-3 and DU-145 prostate carcinoma cell lines using Western blot and RT-PCR (Piñeiro et al., 2011). Thus, multiple cannabinoid receptor subtypes and endocannabinoid hydrolysing enzymes are localized to prostate tissue and synthetic cannabinoids, endocannabinoids and related compounds inhibit prostate cancer cell proliferation and produce apoptosis through CB1 and/or CB2 receptor mechanisms.
Table 3.
Prostate cancer in vitro and ex vivo studies
| Apoptosis/proliferation mediated by | ||||||
|---|---|---|---|---|---|---|
| Human cell line/tissue | Expressing | Apoptosis | Proliferation | CB1 | CB2 | Reference |
| PC-3 | CB1 | ↑ by Δ9-THC | – | Not AM251 | – | Ruiz et al., 1999 |
| DU-145 | CB1, low CB2 | – | ↓ by AEA, 2-AG, MET, HU-210, arvanil | SR1 for AEA, arvanil | Not SR2 for AEA, arvanil | Melck et al., 2000 |
| DU-145 | – | – | ↓ by AEA, arvanil, HU-210 enhanced by PEA | – | – | Di Marzo et al., 2001 |
| PC-3 | – | – | ↓ by Δ9-THC | Not SR1 | – | Velasco et al., 2001 |
| PC-3 | – | – | ↑ by Δ9-THC | SR1 | – | Velasco et al., 2001 |
| PC-3 | – | ↑ by AEA | ↓ by AEA | SR1 for apo | SR2 for apo; not SR2 for pro | Mimeault et al., 2003 |
| LNCaP | – | ↑ by AEA | ↓ by AEA | SR1 for pro | Not SR2 for pro | Mimeault et al., 2003 |
| DU-145 | ||||||
| PC-3 | CB1, CB2 | – | ↑ by Δ9-THC, MET | – | – | Sánchez et al., 2003a |
| LNCaP | – | – | ↑ by Δ9-THC, MET | SR1 for MET | SR2 for MET | Sánchez et al., 2003b |
| LNCaP | – | – | ↓ by JWH-015 | – | – | Sánchez et al., 2003b |
| PC-3 | CB1, CB2 | – | ↓ by Noladin ether, WIN-2, MET | – | – | Nithipatikom et al., 2004 |
| DU-145 | ||||||
| PC-3 | CB1, CB2 | – | ↓ by endocannabinoid hydrolysis blockade (MAFP) | SR1 | – | Nithipatikom et al., 2004 |
| DU-145 | ||||||
| LNCaP | CB1, CB2 | – | – | – | – | Nithipatikom et al., 2004 |
| PC-3 | FAAH | – | – | – | – | Ruiz-Llorente et al., 2004 |
| PC-3 | – | – | ↓ by 2-AG hydrolysis blockade (OTFP) | SR1 | – | Nithipatikom et al., 2005 |
| DU-145 | – | – | ↓ by 2-AG hydrolysis blockade (OTFP) | – | – | Nithipatikom et al., 2005 |
| LNCaP | CB1, CB2 | ↑ by WIN-2 | ↓ by WIN-2 | SR1 | SR2 | Sarfaraz et al., 2005 |
| DU-145 | CB1, CB2 | – | – | – | – | Sarfaraz et al., 2005 |
| PC-3 | ||||||
| CWR22Rv1 | ||||||
| CA-HPV-10 | ||||||
| DU-145 | – | CBD no Δ | ↓ by CBD, CBG, CBC, CBD acid, THC acid | – | – | Ligresti et al., 2006 |
| LNCaP | – | ↑ by WIN-2 | – | – | – | Sarfaraz et al., 2006 |
| PC-3 | – | – | ↓ by 2-AG hydrolysis blockade (OTFP) | – | – | Endsley et al., 2007 |
| Prostate tumour | FAAH high | – | – | – | – | Endsley et al., 2008 |
| LNCaP | FAAH high | – | ↓ by FAAH blockade (CAY10401) | – | – | Endsley et al., 2008 |
| DU-145 | FAAH medium | – | – | – | – | Endsley et al., 2008 |
| PC-3 | FAAH low | – | – | – | – | Endsley et al., 2008 |
| LNCaP | FAAH | – | – | – | – | Wang et al., 2008 |
| PC-3 | ||||||
| DU-145 | ||||||
| Prostate tumour | CB1 high in severe cancer | – | – | – | – | Chung et al., 2009 |
| Prostate tumour | CB1 high | – | – | – | – | Czifra et al., 2009 |
| PC-3 | – | ↑ by MET, JWH-015 | ↓ by MET, JWH-015 | Not SR1 | SR2 apo | Olea-Herrero et al., 2009a |
| LNCaP | – | ↑ by MET, JWH-015 | ↓ by MET, JWH-015 | – | – | Olea-Herrero et al., 2009a |
| DU-145 | ||||||
| PC-3 | – | ↑ by MET | ↓ by MET | Not SR1 | SR2 partially | Olea-Herrero et al., 2009b |
| LNCaP | CB1, CB2, FAAH | – | ↓ by EPEA | – | – | Brown et al., 2010 |
| PC-3 | CB1, CB2 | – | ↓ by EPEA | AM281 | AM630 | Brown et al., 2010 |
↑, increase; ↓, decrease; –, not tested; 2-AG, 2-arachidonoyl glycerol; AEA, anandamide; apo, apoptosis; CAY10401, fatty-acid amide hydrolase inhibitor; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; CBC, cannabichromene; CBD, cannabidiol; CBD acid, cannabidiol acid; CBG, cannabigerol; Δ9-THC, delta-9 tetrahydrocannabinol; EPEA, eicospentaenoyl ethanolamide; FAAH, fatty-acid amide hydrolase; MAFP, methyl arachidonyl fluorophosphonate; MET, methanandamide; no Δ, no change; OTFP (3-octylthio-1,1,1-trifluoropropan-2-one; PEA, palmitoylethanolamide; pro, proliferation; SR1, SR141716A; SR2, SR144528; THC acid, tetrahydrocannabinol; WIN-2, WIN55,212-2.
In vitro studies have extensively evaluated the ability of different endocannabinoids or cannabis-like compounds to inhibit prostate cancer cell proliferation and/or induce apoptosis in different prostate carcinoma (PC-3, DU-145, LNCaP, CWR22Rv1, CA-HPV-10) cell lines (Table 3). Δ9-THC possesses anti-proliferative (Velasco et al., 2001) and pro-apoptotic properties in prostate cancer cell lines (Ruiz et al., 1999). The pro-apoptotic effects of Δ9-THC are not mediated by CB1 receptors, although a possible role for CB2 receptors was not assessed (Ruiz et al., 1999). Methanandamide has also been shown to inhibit prostate cancer cell proliferation (Melck et al., 2000; Nithipatikom et al., 2004; Olea-Herrero et al., 2009a,b;) and induce apoptosis (Olea-Herrero et al., 2009a); these effects are mediated by CB2, but not CB1 receptors. Both Δ9-THC and methanandamide induce cell proliferation at nanomolar concentrations (Velasco et al., 2001; Sánchez et al., 2003a,b;), suggesting that the observed effects are likely to be physiologically relevant. The mixed cannabinoid agonist WIN55,212-2 also inhibits cell proliferation (Nithipatikom et al., 2004; Sarfaraz et al., 2005) and induces apoptosis (Sarfaraz et al., 2005, 2006); these effects are mediated by both CB1 and CB2 receptors (Sarfaraz et al., 2005). Furthermore, the CB2-preferring agonist JWH-015 has anti-proliferative (Sánchez et al., 2003b; Olea-Herrero et al., 2009a) and pro-apoptotic properties (Olea-Herrero et al., 2009a) and these effects are mediated by CB2, but not CB1 receptors (Olea-Herrero et al., 2009a). HU-210 also inhibits cell proliferation (Melck et al., 2000), although pharmacological specificity was not evaluated. Finally, synthetic endocannabinoids also exhibit anti-proliferative properties. In fact, AEA inhibits cell proliferation through CB2, but not CB1 receptors (Melck et al., 2000; Mimeault et al., 2003), whereas induction of apoptosis involves both CB1 and CB2 receptors (Mimeault et al., 2003). Inhibition of endocannabinoid hydrolysis by methyl arachidonyl fluorophosphonate (Nithipatikom et al., 2004), which targets FAAH and MGL, and by CAY10401 (Endsley et al., 2008), which aims FAAH, suggests that elevation of endocannabinoids also inhibits cell proliferation. Inhibition of 2-AG hydrolysis by OTFP (3-octylthio-1,1,1-trifluoropropan-2-one), a compound containing a trifluoromethylketone moiety, also inhibited cell proliferation (Nithipatikom et al., 2005; Endsley et al., 2007) through a CB1-dependent mechanism (Nithipatikom et al., 2005). More work is necessary to demonstrate the specificity of OTFP for MGL in the model system. Phytocannabinoids including CBD, cannabigerol and cannabichromene, cannabidiol acid and THC acid (Ligresti et al., 2006) as well as putative endocannabinoids such as arvanil (Melck et al., 2000) and noladin ether (Nithipatikom et al., 2004) all inhibit cell proliferation in different prostate cancer cell lines (Table 3). Anti-proliferative effects of arvanil are mediated by CB1, but not CB2 receptors (Melck et al., 2000). Interestingly, palmitoylethanolamide (PEA) enhances the anti-proliferative effects of AEA, arvanil and HU-210 in a prostate cancer cell line (Di Marzo et al., 2001), suggestive of an entourage effect. Omega-3 fatty-acid ethanolamides such as eicosapenta enoyl ethanolamide (EPEA) also exhibit anti-proliferative properties which involve both CB1 and CB2 receptors (Brown et al., 2010).
In prostate cancer cells, cannabinoids, following receptor binding, inhibit cell proliferation and induce cell cycle arrest and apoptosis through cAMP-PKA/Raf-ERK signalling pathways. Indeed, treatment with anandamide produced an inhibition of epiderdmal growth factor (EGF)-induced proliferation via cell cycle arrest in prostate cancer cells and a down-regulation of EGF receptors levels (Mimeault et al., 2003; Bifulco et al., 2008) (Figure 1, right inset). Another study showed that treatment with WIN55,212-2 decreased androgen receptor (AR) expression and prostate specific antigen (PSA) levels in prostate cancer cells and also induced apoptosis (Sarfaraz et al., 2005, 2006) (Figure 1, right inset). However, other signalling pathways may also be involved or implicated in the modulation of apoptosis, cell cycle arrest and/or proliferation by cannabinoids.
One in vivo model of prostate cancer has evaluated the ability of a cannabinoid to inhibit tumour growth. In this model, subcutaneous injection of PC-3 cells (prostate carcinoma cells) in the right flank of athymic nude male mice induced the development of tumours (Olea-Herrero et al., 2009a). Interestingly, direct peritumoral administration of the CB2 preferring agonist JWH-015 reduced tumour growth in the mice and this reduction of growth was inhibited by the CB2 receptor antagonist SR144528 (Olea-Herrero et al., 2009a). Injection of canine prostate carcinoma (ACE-1) cells into the mouse femur produces bone pain and bone remodelling, sprouting of calcitonin gene-related peptide (CGRP) and of sensory nerve fibres (Jimenez-Andrade et al., 2010a), changes which impact processes implicated in bone metastasis observed following prostate cancer. The development of new animal models which better reproduce prostate cancer observed clinically may improve prostate cancer treatment; injection of prostate tumour cells into the flank limits translational relevance of the model and does not truly reproduce cancer originating in the prostate gland.
Bone cancer
Primary bone cancer identified as sarcomas arises in the cartilage. Chondrosarcoma and osteosarcoma are the most frequent primary bone cancers (Bovée et al., 2010). Secondary bone cancer is more frequent as it is associated with the wide spread of other cancers through bone metastases from lung or other solid tumours (Mercadante, 1997; Portenoy et al., 1999).
In mice injected with NCTC-2472 sarcoma cell lines, CB1 (Khasabova et al., 2008) or CB2 (Hald et al., 2008) receptors are up-regulated in DRG ipsilateral to cancer bearing limb. However, no change in the expression of CB1 (Hald et al., 2008; Furuse et al., 2009) or CB2 (Hald et al., 2008; Curto-Reyes et al., 2010) receptors was observed in the spinal cord, suggesting that the observed up-regulation was restricted to the periphery. Another study failed to observe up-regulation of CB2 in DRG (Curto-Reyes et al., 2010). Furthermore, FAAH activity is also increased in DRG and plantar skin of mice injected with NCTC-2472 sarcoma cells (Khasabova et al., 2008).
In vitro studies demonstrate that multiple cannabinoid or cannabis-derived compounds induce apoptosis and/or reduce bone resorption in different bone sarcoma (i.e. NCTC-2472, B16-F10) cell lines (Table 4). WIN55,212-2 induces apoptosis in the NCTC-2472 sarcoma cell line (Hald et al., 2008). Furthermore, the CB2 agonist AM1241 produced a reduction in bone loss in the femur of mice injected with NCTC-2472 sarcoma cell line (Lozano-Ondoua et al., 2010). Thus, regulatory changes in the endocannabinoid system (in DRG and plantar skin) are observed in models of bone cancer pain and activation of cannabinoid CB1 or CB2 receptors produces anti-nociception and apoptosis. Activation of CB2 receptors additionally reduces bone loss in tumour-treated mice, suggesting that many aspects of cannabinoid pharmacology may promote anti-cancer effects.
Table 4.
Bone cancer in vitro and ex vivo studies
| Murine cell line\tissue | Tumour induction (cell line) | Expressing | Apoptosis | Bone loss | Reference |
|---|---|---|---|---|---|
| NCTC-2472 | NA | CB1, CB2 | WIN-2 | – | Hald et al., 2008 |
| Femur | NCTC-2472 | – | – | No Δ by WIN-2 | Hald et al., 2008 |
| DRG | NCTC-2472 | ↑ by CB2 ipsi; no Δ CB1 | – | – | Hald et al., 2008 |
| Spinal cord | NCTC-2472 | No Δ CB1, CB2 | – | – | Hald et al., 2008 |
| DRG | NCTC-2472 | ↑ by CB1 ipsi | – | – | Khasabova et al., 2008 |
| DRG and plantar skin | NCTC-2472 | ↑ by FAAH activity | – | – | Khasabova et al., 2008 |
| Spinal cord | NCTC-2472 | No Δ CB1 | – | – | Furuse et al., 2009 |
| Spinal cord and DRG | NCTC-2472 | No Δ CB2 | – | – | Curto-Reyes et al., 2010 |
| Spinal cord and DRG | B16-F10 | No Δ CB2 | – | – | Curto-Reyes et al., 2010 |
| Femur | NCTC-2472 | – | – | ↓ by AM1241 | Lozano-Ondoua et al., 2010 |
↑, increase; ↓, decrease; –, not tested; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; DRG, dorsal root ganglion; FAAH, fatty-acid amide hydrolase; ipsi, ipsilateral side; no Δ, no change; NA, not applicable; WIN-2, WIN55,212-2.
In vivo bone cancer models differ from breast and prostate cancer models in that they more readily lend themselves to direct assessments of therapeutic effects of cannabinoids such as anti-nociception (Jimenez-Andrade et al., 2010b). These studies have evaluated the efficacies of cannabinoids in suppressing tumour-evoked hyperalgesia and/or reductions of tumour growth and metastasis. Injection of different cancer (fibrosarcoma NCTC-2472 or melanoma B16-F10) cells into the calcaneous, tibial or femur bones of mice produces bone tumours (Table 5). Systemic injection of CP55 940 produced anti-nociceptive properties in the tail flick test and suppressed mechanical hyperalgesia (to von Frey stimulation) in this model (Hamamoto et al., 2007). These anti-nociceptive effects were mediated by CB1, but not CB2 receptors (Hamamoto et al., 2007) (Table 5). Effects of subcutaneously administered WIN55,212-2 on weight bearing and mechanical hyperalgesia were consistent with cannabinoid receptor-mediated anti-nociception (Hald et al., 2008). WIN55,212-2 also attenuates tumour-evoked mechanical hyperalgesia following local (intraplantar) administration through activation of CB1 and CB2 receptors (Potenzieri et al., 2008). Endocannabinoids and modulators of the endocannabinoid system also attenuate tumour-evoked pain. Indeed, intraplantar administration of AEA reduces mechanical hyperalgesia and URB597, a potent inhibitor of FAAH, increases AEA levels and decreases hyperalgesia in a model of calcaneous bone cancer pain (Khasabova et al., 2008). These effects of AEA and URB597 are mediated by CB1, but not CB2 receptors (Khasabova et al., 2008). However, intrathecal administration of either FAAH (URB597) or MGL (URB602) inhibitors failed to produce anti-nociception when tested for spontaneous flinches, limb use and weight bearing (Furuse et al., 2009). Moreover, the CB1 agonist arachidonoyl-2-chloroethylamide (ACEA) produces anti–nociceptive properties following intrathecal administration in this model; ACEA suppressed spontaneous flinches and increased limb use and weight bearing through CB1, but not CB2 receptor mechanisms (Furuse et al., 2009) (Table 5). Furthermore, systemic and intrathecal administration of the CB2 agonist AM1241 produces anti-nociception measured with multiple dependent measures (i.e. flinches, limb use, hot plate, von Frey) (Curto-Reyes et al., 2010; Lozano-Ondoua et al., 2010). These anti-nociceptive effects, assessed with multiple different testing methods, are all mediated by CB2, but not CB1 receptors (Curto-Reyes et al., 2010; Lozano-Ondoua et al., 2010). Animal models of bone cancer closely reproduce human bone cancer because tumour cells are injected directly into the bone, where the cancer originates in humans. New techniques for injection of mammary rat metastasis tumour cells in the femur have also recently been developed in rats (Doré-Savard et al., 2010).
Table 5.
Bone cancer in vivo studies
| Mediated by | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Mouse strain | Tumour induction (cell line) | Drugs | Dose and route | Anti-nociception | Mechanical anti-allodynia | Catalepsy | CB1 | CB2 | Reference |
| C3H/HeNCr | NCTC-2472 calcaneous | CP55,940 | 0.1–3 mg·kg−1 i.p. | CP55,940 TF | CP55,940 VF | CP55,940 BT | SR1 TF, VF, BT | Not SR2 TF, VF, BT | Hamamoto et al., 2007 |
| C3H/HeN | NCTC-2472 femur | WIN-2 | 0.5–5 mg·kg−1 s.c. | WIN-2 WB, OF | WIN-2 VF | – | – | – | Hald et al., 2008 |
| C3H/HeN | NCTC-2472 calcaneous | AEA | 1 µg i.pl. | – | AEA VF | – | AM281 VF | Not AM630 VF | Khasabova et al., 2008 |
| C3H/HeN | NCTC-2472 calcaneous | URB597 | 9 µg i.pl. | – | URB597 VF | – | AM281 VF | Not AM630 VF | Khasabova et al., 2008 |
| C3H/He | NCTC-2472 calcaneous | WIN-2 | 1.5–10 µg i.pl. | – | WIN-2 VF | No WIN-2 | AM251 VF | AM630 VF | Potenzieri et al., 2008 |
| C57BL/6J | NCTC-2472 femur | ACEA | 1 nmol i.t. | ACEA SF, LU, WB | – | No ACEA | AM251 SF, LU, WB | Not AM630 SF, LU, WB | Furuse et al., 2009 |
| C57BL/6J | NCTC-2472 femur | URB597 | 0.3–3 nmol i.t. | No SF, LU, WB | – | – | – | – | Furuse et al., 2009 |
| C57BL/6J | NCTC-2472 femur | URB602 | 0.17–1.7 nmol i.t. | No SF, LU, WB | – | – | – | – | Furuse et al., 2009 |
| C3H/He | NCTC 2472 tibia | AM1241 | 1–10 mg·kg−1 i.p. | AM1241 HP | AM1241 VF | – | Not AM251 HP, VF | SR2 HP, VF | Curto-Reyes et al., 2010 |
| C3H/He | NCTC 2472 tibia | AM1241 | 0.1–1 µg i.t. | AM1241 HP | AM1241 VF | – | – | SR2 HP, VF | Curto-Reyes et al., 2010 |
| C57BL/6 | B16-F10 tibia | AM1241 | 1–10 mg·kg−1 i.p. | AM1241 HP | AM1241 VF | – | Not AM251 HP, VF | SR2 HP, VF | Curto-Reyes et al., 2010 |
| C3H/HeJ | NCTC-2472 femur | AM1241 | 6 mg·kg−1 i.p. | AM1241 SF, LU | AM1241 VF | – | – | SR2 SF, LU, VF | Lozano-Ondoua et al., 2010 |
↑, increase; ↓, decrease; –, not tested; AEA, anandamide; ACEA, arachidonyl-2′-chloroethylamide; BT, bar test; CB1, cannabinoid receptor 1; CB2, cannabinoid receptor 2; HP, hot plate; i.p., intraperitoneal; i.pl., intraplantar; i.t., intrathecal; LU, limb use; no Δ, no change; OF, open field; s.c., subcutaneous; SF, spontaneous flinches; SR1, SR141716A; SR2 SR144528; TF, tail flick; VF, von Frey; WIN-2, WIN55,212-2; WB, weight bearing.
Cannabinoids in clinical cancer studies
Cannabinoids have been evaluated in cancer patients for their anti-emetic, anti-nociceptive and orexigenic properties. Numerous studies have been published documenting the anti-emetic properties of cannabis-like compounds (Davis, 2008; Navari, 2009; Parker et al., 2011 for reviews). Fewer clinical trials have evaluated pain relief following cannabinoid administration in cancer patients. Indeed, the role of cannabinoids in relieving pain associated with cancer has been evaluated in only five clinical studies. Four of these studies were performed more than 32 years ago (Noyes et al., 1975a,b; Jochimsen et al., 1978; Staquet et al., 1978; Johnson et al., 2010). In the initial study, oral THC, at doses of 15 and 20 mg, produced analgesic effects in patients experiencing cancer pain (Noyes et al., 1975a). The same study showed that oral administration of single lower dose of Δ9-THC (10 mg) to patients with cancer pain was well-tolerated and produced a mild analgesic effect, whereas higher doses of Δ9-THC (20 mg) also produced adverse side effects (Noyes et al., 1975b). Another study, using a nitrogen analogue of Δ9-THC, showed that pain relief was superior to placebo in patients with cancer (Staquet et al., 1978). Another group, also using a nitrogen analogue of Δ9-THC, showed that the cannabinoid (2 or 4 mg) was not effective as an analgesic, compared with placebo and even appeared to augment pain perception in patients with chronic pain due to malignancies (Jochimsen et al., 1978). A more recent study demonstrated that the administration of Sativex (Δ9-THC: CBD in a 1:1 ratio) reduced pain scores when compared with placebo, whereas effects of Δ9-THC administration alone on pain did not reach significance (Johnson et al., 2010).
The present studies suggest that additional clinical trials evaluating the therapeutic efficacy of cannabinoids in cancer pain are warranted, particularly in light of other aspects of cannabinoid receptor pharmacology that hold considerable therapeutic potential (i.e. anti-emetic and anti-tumour properties).
Conclusion and limitations
The available literature suggests that the endocannabinoid system may be targeted to suppress the evolution and progression of breast, prostate and bone cancer as well as the accompanying pain syndromes. Although this review focuses on these three types of cancer, activation of the endocannabinoid signalling system produces anti-cancer effects in other types of cancer including skin, brain (gliomas) and lung (Velasco et al., 2007; Bíróet al., 2009; Pacher and Mechoulam, 2011 for reviews). Interestingly, cannabis trials in population-based studies failed to show any evidence for increased risk of respiratory symptoms/chronic obstructive pulmonary disease (Tan et al., 2009; Hancox et al., 2010) or lung cancer (Tashkin, 2005) associated with smoking cannabis. Moreover, synthetic cannabinoids and the endocannabinoid system play a role in inhibiting cancer cell proliferation and angiogenesis, reducing tumour growth and metastases and inducing apoptosis in all three types of cancers reviewed here. These observations raise the possibility that a dysregulation of the endocannabinoid system may promote cancer, by fostering physiological conditions that allow cancer cells to proliferate, migrate and grow. These observations also raise the exciting possibility that enhancing cannabinoid tone through cannabinoid-based pharmacotherapies may attenuate these harmful processes to produce anti-cancer effects in humans. However, the basic research findings are far from being completely understood and further research is warranted to better understand the complexity of dynamic changes in the endocannabinoid system in cancer. One of the reasons for this complexity is likely attributable to the highly interactive nature of lipid signalling pathways which recruit different signalling pathways and mechanisms of action. Indeed, endocannabinoids are known to interact with the cyclooxgenase enzyme, inhibit the transcription of genes implicated in metastasis processes, induce cell cycle arrest, activate the formation of reactive oxygen species and ensure the integrity of raft/caveolae needed for anti-proliferative properties. However, other mechanisms are also likely to be involved and interact with the endocannabinoid system in ways that are yet to be discovered.
Many in vitro and in vivo studies have shown that cannabinoids are efficacious in reducing cancer progression (i.e. inhibition of tumour growth and metastases as well as induction of apoptosis and other anti-cancer properties) in breast, prostate and bone cancer. However, further research is needed because the complexity of the effects of cannabinoids and their interaction with other mechanisms and signalling pathways remain to be elucidated. The need for further study is particularly crucial in the case of prostate cancer; only one study, performed in mice, has evaluated in vivo effects of a cannabinoid (JWH-015) on tumour growth. The paucity of in vivo preclinical and clinical data is striking given the large number of compounds that have been tested in vitro in different types of prostate cancer cell lines.
Despite the need for further in vitro and in vivo studies, the literature is nearly unanimous in suggesting that cannabinoids and endocannabinoids reduce the progression of cancer in both in vivo preclinical and in vitro model systems. The need for additional clinical trials of cannabinoid therapeutic efficacy in cancer appears beyond doubt; only few studies have evaluated the effects of cannabinoid in alleviating cancer pain, in contrast to the extensive literature supporting efficacy of cannabinoids as anti-emetics. Furthermore, future research needs to explore the therapeutic potential of multimodal analgesic strategies that combine cannabinoids with other commonly used medications (opioids) or employ multiple phytocannabinoids in combination. The use of different pharmacotherapies in combination may increase the likelihood of synergistic interactions between compounds with multiple distinct mechanisms of action; such combinations may produce a more beneficial therapeutic ratio in cancer patients compared with conventional analgesics, resulting in improved pain relief and anti-cancer effects with fewer adverse side effects. Moreover, because cannabinoids attenuate neuropathy produced by cancer chemotherapy through CB1 and CB2-dependent mechanisms (Rahn and Hohmann, 2009 for review), the possibility remains that cannabinoids in combination with chemotherapy may enhance both anti-tumour actions of chemotherapy and attenuate unwanted iatrogenic side effects (e.g. emesis, neuropathy). Further basic research on cannabinoid anti-cancer properties as well as clinical trials evaluating cannabinoid efficacy in cancer are required before cannabinoid use can be established and accepted as effective adjuncts to cancer therapy.
Acknowledgments
Dedicated to my mother, Manon Marcotte, who passed away early 2011 of lymphoma (JG) and all the people who have lost their battles against cancer and to all those who are still fighting this dreadful disease. JG is supported by a Fonds de la recherche en santé du Québec (FRSQ) postdoctoral fellowship. AGH is supported by DA021644 and DA028200.
Glossary
Abbreviations
- 2-AG
2-arachidonoylglycerol
- AC
adenylyl cyclase
- ACEA
arachidonyl-2′-chloroethylamide
- AEA
anandamide
- AKT
protein kinase B
- AR
androgen receptor
- ATP
adenosine triphosphate
- Bax
pro-apoptotic protein
- Bcl2
anti-apoptotic protein
- brca1
breast cancer susceptibility gene product
- cAMP
cyclic adenosine monophosphate
- CBD
cannabidiol
- CB
cannabinoid
- CB1
cannabinoid receptor 1
- CB2
cannabinoid receptor 2
- CBN
cannabinol
- Cdc2
p34 cyclin-dependent kinase 1
- CDK
cyclin-dependent kinases
- CNS
central nervous system
- DALN
desacetyllevonantradol
- Δ9-THC
delta 9-tetrahydrocannabinol
- DRG
dorsal root ganglion
- EGF
epidermal growth factor receptor
- EPEA
eicosapentaenoyl ethanolamide
- ERK
extracellular regulated kinase
- FAAH
fatty-acid amide hydrolase
- GPR55
G-protein-coupled receptor 55
- H2O2
hydrogen peroxide
- p27/KIP1
cyclin kinase inhibitor
- p38MAPK
p38 mitogen-activated protein kinase
- PRL
prolactin receptor
- MET
methanandamide
- MGL
or MAGL, monoacylglycerol lipase
- OTFP
3-octylthio-1,1,1-trifluoropropan-2-one
- p53
p53 protein
- p21ras
p21 ras protein
- PEA
palmitoylethanolamide
- PI3K
phosphatidyl inositol 3 kinase
- PKA
protein kinase A
- PKB
protein kinase B
- PRL
prolactin receptor
- PSA
prostate-specific antigen
- Raf-1
protein Raf-1
- RT-PCR
reverse transcriptase polymerase chain reaction
- Trk
high-affinity nerve growth factor receptor
- TRPV1
transient receptor potential cation channel V1
Supporting Information
Teaching Materials; Fig 1 as PowerPoint slide.
References
- Alexander SPH, Mathie A, Peters JA. Guide to Receptors and Channels (GRAC), 4th Edition. Br J Pharmacol. 2009;158(Suppl 1):S1–S254. doi: 10.1111/j.1476-5381.2009.00499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-Shabat S, Fride E, Sheskin T, Tamiri T, Rhee MH, Vogel Z, et al. An entourage effect: inactive endogenous fatty acid glycerol esters enhance 2-arachidonoyl-glycerol cannabinoid activity. Eur J Pharmacol. 1998;353:23–31. doi: 10.1016/s0014-2999(98)00392-6. [DOI] [PubMed] [Google Scholar]
- Bifulco M, Laezza C, Portella G, Vitale M, Orlando P, De Petrocellis L, et al. Control by the endogenous cannabinoid system of ras oncogene-dependent tumor growth. FASEB J. 2001;15:2745–2747. doi: 10.1096/fj.01-0320fje. [DOI] [PubMed] [Google Scholar]
- Bifulco M, Malfitano AM, Pisanti S, Laezza C. Endocannabinoids in endocrine and related tumours. Endocr Relat Cancer. 2008;15:391–408. doi: 10.1677/ERC-07-0258. [DOI] [PubMed] [Google Scholar]
- Bíró T, Tóth BI, Haskó G, Paus R, Pacher P. The endocannabinoid system of the skin in health and disease: novel perspectives and therapeutic opportunities. Trends Pharmacol Sci. 2009;30:411–420. doi: 10.1016/j.tips.2009.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bisogno T, Katayama K, Melck D, Ueda N, De Petrocellis L, Yamamoto S, et al. Biosynthesis and degradation of bioactive fatty acid amides in human breast cancer and rat pheochromocytoma cells – implications for cell proliferation and differentiation. Eur J Biochem. 1998;254:634–642. doi: 10.1046/j.1432-1327.1998.2540634.x. [DOI] [PubMed] [Google Scholar]
- Blazquez C, Casanova ML, Planas A, Gomez Del Pulgar T, Villanueva C, Fernandez-Acenero MJ, et al. Inhibition of tumor angiogenesis by cannabinoids. FASEB J. 2003;17:529–531. doi: 10.1096/fj.02-0795fje. [DOI] [PubMed] [Google Scholar]
- Bovée JV, Hogendoorn PC, Wunder JS, Alman BA. Cartilage tumours and bone development: molecular pathology and possible therapeutic targets. Nat Rev Cancer. 2010;10:481–488. doi: 10.1038/nrc2869. [DOI] [PubMed] [Google Scholar]
- Bray F, Lortet-Tieulent J, Ferlay J, Forman D, Auvinen A. Prostate cancer incidence and mortality trends in 37 European countries: an overview. Eur J Cancer. 2010;46:3040–3052. doi: 10.1016/j.ejca.2010.09.013. [DOI] [PubMed] [Google Scholar]
- Brown I, Cascio MG, Wahle KW, Smoum R, Mechoulam R, Ross RA, et al. Cannabinoid receptor-dependent and -independent anti-proliferative effects of omega-3 ethanolamides in androgen receptor-positive and -negative prostate cancer cell lines. Carcinogenesis. 2010;31:1584–1591. doi: 10.1093/carcin/bgq151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckley NE, McCoy KL, Mezey E, Bonner T, Zimmer A, Felder CC, et al. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB2 receptor. Eur J Pharmacol. 2000;396:141–149. doi: 10.1016/s0014-2999(00)00211-9. [DOI] [PubMed] [Google Scholar]
- von Bueren AO, Schlumpf M, Lichtensteiger W. Delta(9)-tetrahydrocannabinol inhibits 17beta-estradiol-induced proliferation and fails to activate androgen and estrogen receptors in MCF7 human breast cancer cells. Anticancer Res. 2008;28:85–89. [PubMed] [Google Scholar]
- Burstein S, Salmonsen R. Acylamido analogs of endocannabinoids selectively inhibit cancer cell proliferation. Bioorg Med Chem. 2008;16:9644–9651. doi: 10.1016/j.bmc.2008.10.015. [DOI] [PubMed] [Google Scholar]
- Caffarel MM, Sarrió D, Palacios J, Guzmán M, Sánchez C. Delta9-tetrahydrocannabinol inhibits cell cycle progression in human breast cancer cells through Cdc2 regulation. Cancer Res. 2006;66:6615–6621. doi: 10.1158/0008-5472.CAN-05-4566. [DOI] [PubMed] [Google Scholar]
- Caffarel MM, Moreno-Bueno G, Cerutti C, Palacios J, Guzman M, Mechta-Grigoriou F, et al. JunD is involved in the antiproliferative effect of Delta9-tetrahydrocannabinol on human breast cancer cells. Oncogene. 2008;27:5033–5044. doi: 10.1038/onc.2008.145. [DOI] [PubMed] [Google Scholar]
- Caffarel MM, Andradas C, Mira E, Pérez-Gómez E, Cerutti C, Moreno-Bueno G, et al. Cannabinoids reduce ErbB2-driven breast cancer progression through Akt inhibition. Mol Cancer. 2010;9:196–206. doi: 10.1186/1476-4598-9-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carchman RA, Harris LS, Munson AE. The inhibition of DNA synthesis by cannabinoids. Cancer Res. 1976;36:95–100. [PubMed] [Google Scholar]
- Chung SC, Hammarsten P, Josefsson A, Stattin P, Granfors T, Egevad L, et al. A high cannabinoid CB(1) receptor immunoreactivity is associated with disease severity and outcome in prostate cancer. Eur J Cancer. 2009;45:174–182. doi: 10.1016/j.ejca.2008.10.010. [DOI] [PubMed] [Google Scholar]
- Curto-Reyes V, Llames S, Hidalgo A, Menéndez L, Baamonde A. Spinal and peripheral analgesic effects of the CB2 cannabinoid receptor agonist AM1241 in two models of bone cancer-induced pain. Br J Pharmacol. 2010;160:561–573. doi: 10.1111/j.1476-5381.2009.00629.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Czifra G, Varga A, Nyeste K, Marincsák R, Tóth BI, Kovács I, et al. Increased expressions of cannabinoid receptor-1 and transient receptor potential vanilloid-1 in human prostate carcinoma. J Cancer Res Clin Oncol. 2009;135:507–514. doi: 10.1007/s00432-008-0482-3. [DOI] [PubMed] [Google Scholar]
- Davis MP. Oral nabilone capsules in the treatment of chemotherapy-induced nausea and vomiting and pain. Expert Opin Investig Drugs. 2008;17:85–95. doi: 10.1517/13543784.17.1.85. [DOI] [PubMed] [Google Scholar]
- De Petrocellis L, Melck D, Palmisano A, Bisogno T, Laezza C, Bifulco M, et al. The endogenous cannabinoid anandamide inhibits human breast cancer cell proliferation. Proc Natl Acad Sci USA. 1998;95:8375–8380. doi: 10.1073/pnas.95.14.8375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Petrocellis L, Bisogno T, Ligresti A, Bifulco M, Melck D, Di Marzo V. Effect on cancer cell proliferation of palmitoylethanolamide, a fatty acid amide interacting with both the cannabinoid and vanilloid signalling systems. Fundam Clin Pharmacol. 2002;16:297–302. doi: 10.1046/j.1472-8206.2002.00094.x. [DOI] [PubMed] [Google Scholar]
- Di Marzo V. Endocannabinoids: synthesis and degradation. Rev Physiol Biochem Pharmacol. 2006;160:1–24. doi: 10.1007/112_0505. [DOI] [PubMed] [Google Scholar]
- Di Marzo V. Targeting the endocannabinoid system: to enhance or reduce? Nat Rev Drug Discov. 2008;7:438–455. doi: 10.1038/nrd2553. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Melck D, Orlando P, Bisogno T, Zagoory O, Bifulco M, et al. Palmitoylethanolamide inhibits the expression of fatty acid amide hydrolase and enhances the anti-proliferative effect of anandamide in human breast cancer cells. Biochem J. 2001;358:249–255. doi: 10.1042/0264-6021:3580249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Djulbegovic M, Beyth RJ, Neuberger MM, Stoffs TL, Vieweg J, Djulbegovic B, et al. Screening for prostate cancer: systematic review and meta-analysis of randomised controlled trials. BMJ. 2010;341:4543–4551. doi: 10.1136/bmj.c4543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doré-Savard L, Otis V, Belleville K, Lemire M, Archambault M, Tremblay L, et al. Behavioral, medical imaging and histopathological features of a new rat model of bone cancer pain. Plos One. 2010;5:13774–13788. doi: 10.1371/journal.pone.0013774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endsley MP, Aggarwal N, Isbell MA, Wheelock CE, Hammock BD, Falck JR, et al. Diverse roles of 2-arachidonoylglycerol in invasion of prostate carcinoma cells: Location, hydrolysis and 12-lipoxygenase metabolism. Int J Cancer. 2007;121:984–991. doi: 10.1002/ijc.22761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endsley MP, Thill R, Choudhry I, Williams CL, Kajdacsy-Balla A, Campbell WB, et al. Expression and function of fatty acid amide hydrolase in prostate cancer. Int J Cancer. 2008;123:1318–1326. doi: 10.1002/ijc.23674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127:2893–2917. doi: 10.1002/ijc.25516. [DOI] [PubMed] [Google Scholar]
- Fontanini G. Angiogenesis and cancer. Surg Technol Int. 2000;9:25–32. [PubMed] [Google Scholar]
- Ford LA, Roelofs AJ, Anavi-Goffer S, Mowat L, Simpson DG, Irving AJ, et al. A role for L-alpha-lysophosphatidylinositol and GPR55 in the modulation of migration, orientation and polarization of human breast cancer cells. Br J Pharmacol. 2010;160:762–771. doi: 10.1111/j.1476-5381.2010.00743.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuse S, Kawamata T, Yamamoto J, Niiyama Y, Omote K, Watanabe M, et al. Reduction of bone cancer pain by activation of spinal cannabinoid receptor 1 and its expression in the superficial dorsal horn of the spinal cord in a murine model of bone cancer pain. Anesthesiology. 2009;111:173–186. doi: 10.1097/ALN.0b013e3181a51e0d. [DOI] [PubMed] [Google Scholar]
- Glass AG, Lacey JV, Jr, Carreon JD, Hoover RN. Breast cancer incidence, 1980-2006: combined roles of menopausal hormone therapy, screening mammography, and estrogen receptor status. J Natl Cancer Inst. 2007;99:1152–1161. doi: 10.1093/jnci/djm059. [DOI] [PubMed] [Google Scholar]
- Gómez del Pulgar T, Velasco G, Guzmán M. The CB1 cannabinoid receptor is coupled to the activation of protein kinase B/Akt. Biochem J. 2000;347:369–373. doi: 10.1042/0264-6021:3470369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimaldi C, Pisanti S, Laezza C, Malfitano AM, Santoro A, Vitale M, et al. Anandamide inhibits adhesion and migration of breast cancer cells. Exp Cell Res. 2006;312:363–373. doi: 10.1016/j.yexcr.2005.10.024. [DOI] [PubMed] [Google Scholar]
- Guindon J, Hohmann AG. The endocannabinoid system and pain. CNS Neurol Disord Drug Targets. 2009;8:403–421. doi: 10.2174/187152709789824660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guise TA, Brufsky A, Coleman RE. Understanding and optimizing bone health in breast cancer. Curr Med Res Opin. 2010;26:3–20. doi: 10.1185/03007995.2010.533162. [DOI] [PubMed] [Google Scholar]
- Guzmán M. Cannabinoids: potential anticancer agents. Nat Rev Cancer. 2003;3:745–755. doi: 10.1038/nrc1188. [DOI] [PubMed] [Google Scholar]
- Hald A, Ding M, Egerod K, Hansen RR, Konradsen D, Jørgensen SG, et al. Differential effects of repeated low dose treatment with the cannabinoid agonist WIN 55,212-2 in experimental models of bone cancer pain and neuropathic pain. Pharmacol Biochem Behav. 2008;91:38–46. doi: 10.1016/j.pbb.2008.04.021. [DOI] [PubMed] [Google Scholar]
- Hamamoto DT, Giridharagopalan S, Simone DA. Acute and chronic administration of the cannabinoid receptor agonist CP 55,940 attenuates tumor-evoked hyperalgesia. Eur J Pharmacol. 2007;558:73–87. doi: 10.1016/j.ejphar.2006.11.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hancox RJ, Poulton R, Ely M, Welch D, Taylor DR, McLachlan CR, et al. Effects of cannabis on lung function: a population-based cohort study. Eur Respir J. 2010;35:42–47. doi: 10.1183/09031936.00065009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heron M, Hoyert DL, Murphy SL, Xu J, Kochanek KD, Tejada-Vera B. Deaths: final data for 2006. National Vital Statistics Reports. 2009;57:1–135. [PubMed] [Google Scholar]
- Jhaveri MD, Richardson D, Chapman V. Endocannabinoid metabolism and uptake: novel targets for neuropathic and inflammatory pain. Br J Pharmacol. 2007;152:624–632. doi: 10.1038/sj.bjp.0707433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimenez-Andrade JM, Bloom AP, Stake JI, Mantyh WG, Taylor RN, Freeman KT, et al. Pathological sprouting of adult nociceptors in chronic prostate cancer-induced bone pain. J Neurosci. 2010a;30:14649–14656. doi: 10.1523/JNEUROSCI.3300-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jimenez-Andrade JM, Mantyh WG, Bloom AP, Ferng AS, Geffre CP, Mantyh PW. Bone cancer pain. Ann N Y Acad Sci. 2010b;1198:173–181. doi: 10.1111/j.1749-6632.2009.05429.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jochimsen PR, Lawton RL, VerSteeg K, Noyes R. Effect of benzopyranoperidine, a delta-9-THC congener, on pain. Clin Pharmacol Ther. 1978;24:223–227. doi: 10.1002/cpt1978242223. [DOI] [PubMed] [Google Scholar]
- Johnson JR, Burnell-Nugent M, Lossignol D, Ganae-Motan ED, Potts R, Fallon MT. Multicenter, double-blind, randomized, placebo-controlled, parallel-group study of the efficacy, safety, and tolerability of THC:CBD extract and THC extract in patients with intractable cancer-related pain. J Pain Symptom Manage. 2010;39:167–179. doi: 10.1016/j.jpainsymman.2009.06.008. [DOI] [PubMed] [Google Scholar]
- Khasabova IA, Khasabov SG, Harding-Rose C, Coicou LG, Seybold BA, Lindberg AE, et al. A decrease in anandamide signaling contributes to the maintenance of cutaneous mechanical hyperalgesia in a model of bone cancer pain. J Neurosci. 2008;28:11141–11152. doi: 10.1523/JNEUROSCI.2847-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kogan NM. Cannabinoids and Cancer. Mini Rev Med Chem. 2005;5:941–952. doi: 10.2174/138955705774329555. [DOI] [PubMed] [Google Scholar]
- Laezza C, Pisanti S, Malfitano AM, Bifulco M. The anandamide analog, Met-F-AEA, controls human breast cancer cell migration via the RHOA/RHO kinase signaling pathway. Endocr Relat Cancer. 2008;15:965–974. doi: 10.1677/ERC-08-0030. [DOI] [PubMed] [Google Scholar]
- Laezza C, Malfitano AM, Proto MC, Esposito I, Gazzerro P, Formisano P, et al. Inhibition of 3-hydroxy-3-methylglutaryl-coenzyme A reductase activity and of Ras farnesylation mediate antitumor effects of anandamide in human breast cancer cells. Endocr Relat Cancer. 2010;17:495–503. doi: 10.1677/ERC-10-0009. [DOI] [PubMed] [Google Scholar]
- Ledent C, Valverde O, Cossu G, Petitet F, Aubert JF, Beslot F, et al. Unresponsiveness to cannabinoids and reduced addictive effects of opiate in CB1 receptor knockout mice. Science. 1999;283:401–404. doi: 10.1126/science.283.5400.401. [DOI] [PubMed] [Google Scholar]
- Ligresti A, Moriello AS, Starowicz K, Matias I, Pisanti S, De Petrocellis L, et al. Antitumor activity of plant cannabinoids with emphasis on the effect of cannabidiol on human breast carcinoma. J Pharmacol Exp Ther. 2006;318:1375–1387. doi: 10.1124/jpet.106.105247. [DOI] [PubMed] [Google Scholar]
- Lozano-Ondoua AN, Wright C, Vardanyan A, King T, Largent-Milnes TM, Nelson M, et al. A cannabinoid 2 receptor agonist attenuates bone cancer-induced pain and bone loss. Life Sci. 2010;86:646–653. doi: 10.1016/j.lfs.2010.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McAllister SD, Christian RT, Horowitz MP, Garcia A, Desprez PY. Cannabidiol as a novel inhibitor of Id-1 gene expression in aggressive breast cancer cells. Mol Cancer Ther. 2007;6:2921–2927. doi: 10.1158/1535-7163.MCT-07-0371. [DOI] [PubMed] [Google Scholar]
- McAllister SD, Murase R, Christian RT, Lau D, Zielinski AJ, Allison J, et al. Pathways mediating the effects of cannabidiol on the reduction of breast cancer cell proliferation, invasion, and metastasis. Breast Cancer Res Treat. 2010 doi: 10.1007/s10549-010-1177-4. DOI: 10.1007/S10549-010-1177-4. PMID: 20859676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKallip RJ, Nagarkatti M, Nagarkatti PS. Delta-9-tetrahydrocannabinol enhances breast cancer growth and metastasis by suppression of the antitumor immune response. J Immunol. 2005;174:3281–3289. doi: 10.4049/jimmunol.174.6.3281. [DOI] [PubMed] [Google Scholar]
- Maccarrone M, Lorenzon T, Bari M, Melino G, Finazzi-Agro A. Anandamide induces apoptosis in human cells via vanilloid receptors. Evidence for a protective role of cannabinoid receptors. J Biol Chem. 2000;275:31938–31945. doi: 10.1074/jbc.M005722200. [DOI] [PubMed] [Google Scholar]
- Madigan MP, Ziegler RG, Benichou J, Byrne C, Hoover RN. Proportion of breast cancer cases in the United States explained by well-established risk factors. J Natl Cancer Inst. 1995;87:1681–1685. doi: 10.1093/jnci/87.22.1681. [DOI] [PubMed] [Google Scholar]
- Matsuda LA, Lolait SJ, Brownstein MJ, Young AC, Bonner TI. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature. 1990;346:561–564. doi: 10.1038/346561a0. [DOI] [PubMed] [Google Scholar]
- Melck D, Rueda D, Galve-Roperh I, De Petrocellis L, Guzmán M, Di Marzo V. Involvement of the cAMP/protein kinase A pathway and of mitogen-activated protein kinase in the anti-proliferative effects of anandamide in human breast cancer cells. FEBS Lett. 1999;463:235–240. doi: 10.1016/s0014-5793(99)01639-7. [DOI] [PubMed] [Google Scholar]
- Melck D, De Petrocellis L, Orlando P, Bisogno T, Laezza C, Bifulco M, et al. Suppression of nerve growth factor Trk receptors and prolactin receptors by endocannabinoids leads to inhibition of human breast and prostate cancer cell proliferation. Endocrinology. 2000;141:118–126. doi: 10.1210/endo.141.1.7239. [DOI] [PubMed] [Google Scholar]
- Mercadante S. Malignant bone pain: pathophysiology and treatment. Pain. 1997;69:1–18. doi: 10.1016/s0304-3959(96)03267-8. [DOI] [PubMed] [Google Scholar]
- Mimeault M, Pommery N, Wattez N, Bailly C, Hénichart JP. Anti-proliferative and apoptotic effects of anandamide in human prostatic cancer cell lines: implication of epidermal growth factor receptor down-regulation and ceramide production. Prostate. 2003;56:1–12. doi: 10.1002/pros.10190. [DOI] [PubMed] [Google Scholar]
- Movsesyan VA, Stoica BA, Yakovlev AG, Knoblach SM, Lea PM, 4th, Cernak I, et al. Anandamide-induced cell death in primary neuronal cultures: role of calpain and caspase pathways. Cell Death Differ. 2004;11:1121–1132. doi: 10.1038/sj.cdd.4401442. [DOI] [PubMed] [Google Scholar]
- Munro S, Thomas KL, Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature. 1993;365:61–65. doi: 10.1038/365061a0. [DOI] [PubMed] [Google Scholar]
- Munson AE, Harris LS, Friedman MA, Dewey WL, Carchman RA. Antineoplastic activity of cannabinoids. J Natl Cancer Inst. 1975;55:597–602. doi: 10.1093/jnci/55.3.597. [DOI] [PubMed] [Google Scholar]
- Navari RM. Antiemetic control: toward a new standard of care for ematogenic chemotherapy. Expert Opin Pharmacother. 2009;10:629–644. doi: 10.1517/14656560902731894. [DOI] [PubMed] [Google Scholar]
- Nithipatikom K, Endsley MP, Isbell MA, Falck JR, Iwamoto Y, Hillard CJ, et al. 2-arachidonoylglycerol: a novel inhibitor of androgen-independent prostate cancer cell invasion. Cancer Res. 2004;64:8826–8830. doi: 10.1158/0008-5472.CAN-04-3136. [DOI] [PubMed] [Google Scholar]
- Nithipatikom K, Endsley MP, Isbell MA, Wheelock CE, Hammock BD, Campbell WB. A new class of inhibitors of 2-arachidonoylglycerol hydrolysis and invasion of prostate cancer cells. Biochem Biophys Res Commun. 2005;332:1028–1033. doi: 10.1016/j.bbrc.2005.05.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noyes R, Brunk SF, Baram DA, Canter A. Analgesic effect of delta-9-tetrahydrocannabinol. J Clin Pharmacol. 1975a;15:139–143. doi: 10.1002/j.1552-4604.1975.tb02348.x. [DOI] [PubMed] [Google Scholar]
- Noyes R, Brunk SF, Avery DA, Canter AC. The analgesic properties of delta-9-tetrahydrocannabinol and codeine. Clin Pharmacol Ther. 1975b;18:84–89. doi: 10.1002/cpt197518184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olea-Herrero N, Vara D, Malagarie-Cazenave S, Díaz-Laviada I. Inhibition of human tumour prostate PC-3 cell growth by cannabinoids R(+)-Methanandamide and JWH-015: involvement of CB2. Br J Cancer. 2009a;101:940–950. doi: 10.1038/sj.bjc.6605248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olea-Herrero N, Vara D, Malagarie-Cazenave S, Díaz-Laviada I. The cannabinoid R+ methanandamide induces IL-6 secretion by prostate cancer PC3 cells. J Immunotoxicol. 2009b;6:249–256. doi: 10.3109/15476910903241696. [DOI] [PubMed] [Google Scholar]
- Pacher P, Mechoulam R. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Prog Lipid Res. 2011;50:193–211. doi: 10.1016/j.plipres.2011.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacher P, Batkai S, Kunos G. The endocannabioid system as an emerging target of pharmacotherapy. Pharmacol Rev. 2006;58:389–462. doi: 10.1124/pr.58.3.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pantel K, Alix-Panabières C. Circulating tumour cells in cancer patients: challenges and perspectives. Trends Mol Med. 2010;16:398–406. doi: 10.1016/j.molmed.2010.07.001. [DOI] [PubMed] [Google Scholar]
- Parker LA, Rock E, Limebeer C. Regulation of nausea and vomiting by cannabinoids. Br J Pharmacol. 2011;163:1411–1422. doi: 10.1111/j.1476-5381.2010.01176.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parkin D, Bray F, Devesa S. Cancer burden in the year 2000. The global picture. Eur J Cancer. 2001;37:S4–S66. doi: 10.1016/s0959-8049(01)00267-2. [DOI] [PubMed] [Google Scholar]
- Piñeiro R, Maffucci T, Falasca M. The putative cannabinoid receptor GPR55 defines a novel autocrine loop in cancer cell proliferation. Oncogene. 2011;30:142–152. doi: 10.1038/onc.2010.417. [DOI] [PubMed] [Google Scholar]
- Piomelli D. The endocannabinoid system: a drug discovery perspective. Curr Opin Investig Drugs. 2005;6:672–679. [PubMed] [Google Scholar]
- Portenoy RK, Payne D, Jacobsen P. Breakthrough pain: characteristics and impact in patients with cancer pain. Pain. 1999;81:129–134. doi: 10.1016/s0304-3959(99)00006-8. [DOI] [PubMed] [Google Scholar]
- Potenzieri C, Harding-Rose C, Simone DA. The cannabinoid receptor agonist, WIN 55, 212-2, attenuates tumor-evoked hyperalgesia through peripheral mechanisms. Brain Res. 2008;1215:69–75. doi: 10.1016/j.brainres.2008.03.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qamri Z, Preet A, Nasser MW, Bass CE, Leone G, Barsky SH, et al. Synthetic cannabinoid receptor agonists inhibit tumor growth and metastasis of breast cancer. Mol Cancer Ther. 2009;8:3117–3129. doi: 10.1158/1535-7163.MCT-09-0448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rahn EJ, Hohmann AG. Cannabinoids as pharmacotherapies for neuropathic pain: from the bench to the bedside. Neurotherapeutics. 2009;6:713–737. doi: 10.1016/j.nurt.2009.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribatti D, Crivellato E. Mast cells, angiogenesis and tumour growth. Biochim Biophys Acta. 2010 doi: 10.1016/j.bbadis.2010.11.010. DOI: 10.1016/j.bbadi3.2010.11.010. PMID: 21130163. [DOI] [PubMed] [Google Scholar]
- Ruh MF, Taylor JA, Howlett AC, Welshons WV. Failure of cannabinoid compounds to stimulate estrogen receptors. Biochem Pharmacol. 1997;53:35–41. doi: 10.1016/s0006-2952(96)00659-4. [DOI] [PubMed] [Google Scholar]
- Ruiz L, Miguel A, Díaz-Laviada I. Delta9-tetrahydrocannabinol induces apoptosis in human prostate PC-3 cells via a receptor-independent mechanism. FEBS Lett. 1999;458:400–404. doi: 10.1016/s0014-5793(99)01073-x. [DOI] [PubMed] [Google Scholar]
- Ruiz-Llorente L, Ortega-Gutiérrez S, Viso A, Sánchez MG, Sánchez AM, Fernández C, et al. Characterization of an anandamide degradation system in prostate epithelial PC-3 cells: synthesis of new transporter inhibitors as tools for this study. Br J Pharmacol. 2004;141:457–467. doi: 10.1038/sj.bjp.0705628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sánchez MG, Ruiz-Llorente L, Sánchez AM, Díaz-Laviada I. Activation of phosphoinositide 3-kinase/PKB pathway by CB(1) and CB(2) cannabinoid receptors expressed in prostate PC-3 cells. Involvement in Raf-1 stimulation and NGF induction. Cell Signal. 2003a;15:851–859. doi: 10.1016/s0898-6568(03)00036-6. [DOI] [PubMed] [Google Scholar]
- Sánchez MG, Sánchez AM, Ruiz-Llorente L, Díaz-Laviada I. Enhancement of androgen receptor expression induced by (R)-methanandamide in prostate LNCaP cells. FEBS Lett. 2003b;555:561–566. doi: 10.1016/s0014-5793(03)01349-8. [DOI] [PubMed] [Google Scholar]
- Sarfaraz S, Afaq F, Adhami VM, Mukhtar H. Cannabinoid receptor as a novel target for the treatment of prostate cancer. Cancer Res. 2005;65:1635–1641. doi: 10.1158/0008-5472.CAN-04-3410. [DOI] [PubMed] [Google Scholar]
- Sarfaraz S, Afaq F, Adhami VM, Malik A, Mukhtar H. Cannabinoid receptor agonist-induced apoptosis of human prostate cancer cells LNCaP proceeds through sustained activation of ERK1/2 leading to G1 cell cycle arrest. J Biol Chem. 2006;281:39480–39491. doi: 10.1074/jbc.M603495200. [DOI] [PubMed] [Google Scholar]
- Sarfaraz S, Adhami VM, Syed DN, Afaq F, Mukhtar H. Cannabinoids for cancer treatment: progress and promise. Cancer Res. 2008;68:339–342. doi: 10.1158/0008-5472.CAN-07-2785. [DOI] [PubMed] [Google Scholar]
- Sarnataro D, Grimaldi C, Pisanti S, Gazzerro P, Laezza C, Zurzolo C, et al. Plasma membrane and lysosomal localization of CB1 cannabinoid receptor are dependent on lipid rafts and regulated by anandamide in human breast cancer cells. FEBS Lett. 2005;579:6343–6349. doi: 10.1016/j.febslet.2005.10.016. [DOI] [PubMed] [Google Scholar]
- Sarnataro D, Pisanti S, Santoro A, Gazzerro P, Malfitano AM, Laezza C, et al. The cannabinoid CB1 receptor antagonist rimonabant (SR141716) inhibits human breast cancer cell proliferation through a lipid raft-mediated mechanism. Mol Pharmacol. 2006;70:1298–1306. doi: 10.1124/mol.106.025601. [DOI] [PubMed] [Google Scholar]
- Schroder FH, Hugosson J, Roobol MJ, Tammela TL, Ciatto S, Nelen V, et al. Screening and prostate-cancer mortality in a randomized European study. N Engl J Med. 2009;360:1320–1328. doi: 10.1056/NEJMoa0810084. [DOI] [PubMed] [Google Scholar]
- Staquet M, Gantt C, Machin D. Effect of a nitrogen analog of tetrahydrocannabinol on cancer pain. Clin Pharmacol Ther. 1978;23:397–401. doi: 10.1002/cpt1978234397. [DOI] [PubMed] [Google Scholar]
- Takeda S, Yamaori S, Motoya E, Matsunaga T, Kimura T, Yamamoto I, et al. Delta(9)-Tetrahydrocannabinol enhances MCF-7 cell proliferation via cannabinoid receptor-independent signaling. Toxicology. 2008;245:141–146. doi: 10.1016/j.tox.2007.12.019. [DOI] [PubMed] [Google Scholar]
- Takeda S, Yamamoto I, Watanabe K. Modulation of Delta9-tetrahydrocannabinol-induced MCF-7 breast cancer cell growth by cyclooxygenase and aromatase. Toxicology. 2009;259:25–32. doi: 10.1016/j.tox.2009.01.024. [DOI] [PubMed] [Google Scholar]
- Tan WC, Lo C, Jong A, Xing L, Fitzgerald MJ, Vollmer WM, et al. Marijuana and chronic obstructive lung disease: a population-based study. CMAJ. 2009;180:814–820. doi: 10.1503/cmaj.081040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tashkin DP. Smoked marijuana as a cause of lung injury. Monaldi Arch Chest Dis. 2005;63:93–100. doi: 10.4081/monaldi.2005.645. [DOI] [PubMed] [Google Scholar]
- Thors L, Bergh A, Persson E, Hammarsten P, Stattin P, Egevad L, et al. Fatty acid amide hydrolase in prostate cancer: association with disease severity and outcome, CB1 receptor expression and regulation by IL-4. Plos One. 2010;5:12275–12286. doi: 10.1371/journal.pone.0012275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Velasco L, Ruiz L, Sánchez MG, Díaz-Laviada I. Delta(9)-Tetrahydrocannabinol increases nerve growth factor production by prostate PC-3 cells. Involvement of CB1 cannabinoid receptor and Raf-1. Eur J Biochem. 2001;268:531–535. doi: 10.1046/j.1432-1327.2001.01884.x. [DOI] [PubMed] [Google Scholar]
- Velasco G, Carracedo A, Blázquez C, Lorente M, Aguado T, Haro A, et al. Cannabinoids and gliomas. Mol Neurobiol. 2007;36:60–67. doi: 10.1007/s12035-007-0002-5. [DOI] [PubMed] [Google Scholar]
- Wang J, Ueda N. Biology of the endocannabinoid synthesis system. Prostaglandins Other Lipid Mediat. 2009;89:112–119. doi: 10.1016/j.prostaglandins.2008.12.002. [DOI] [PubMed] [Google Scholar]
- Wang J, Zhao LY, Uyama T, Tsuboi K, Wu XX, Kakehi Y, et al. Expression and secretion of N-acylethanolamine-hydrolysing acid amidase in human prostate cancer cells. J Biochem. 2008;144:685–690. doi: 10.1093/jb/mvn122. [DOI] [PubMed] [Google Scholar]
- Watanabe K, Motoya E, Matsuzawa N, Funahashi T, Kimura T, Matsunaga T, et al. Marijuana extracts possess the effects like the endocrine disrupting chemicals. Toxicology. 2005;206:471–478. doi: 10.1016/j.tox.2004.08.005. [DOI] [PubMed] [Google Scholar]
- Zimmer A, Zimmer AM, Hohmann AG, Herkenham M, Bonner TI. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1 receptor knockout mice. Proc Natl Acad Sci USA. 1999;96:5780–5785. doi: 10.1073/pnas.96.10.5780. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.