

Abstract
Control of transcription in prokaryotes often involves direct contact of regulatory proteins with RNA polymerase from binding sites located adjacent to the target promoter. Alternatively, in the case of genes transcribed by Escherichia coli RNA polymerase holoenzyme containing the alternate sigma factor ς54, regulatory proteins bound at more distally located enhancer sites can activate transcription via DNA looping by taking advantage of the increasing flexibility of DNA over longer distances. While this second mechanism offers a greater possible flexibility in the location of these binding sites, it is not clear how the specificity offered by the proximity of the regulatory protein and the polymerase intrinsic to the first mechanism is maintained. Here we demonstrate that integration host factor (IHF), a protein that induces a sharp bend in DNA, acts both to inhibit DNA-looping-dependent transcriptional activation by an inappropriate enhancer-binding protein and to facilitate similar activation by an appropriate enhancer-binding protein. These opposite effects have the consequence of increasing the specificity of activation of a promoter that is susceptible to regulation by proteins bound to a distal site.
Keywords: Integration host factor, σ54–RNA polymerase, upstream activation sequence, transcriptional activator
The ability of transcriptional regulatory proteins to act at a distance via DNA looping is characteristic of both prokaryotic and eukaryotic promoters (Matthews 1992; Schleif 1992). Over distances less than the persistence length of DNA (∼150 bp), the DNA is relatively stiff, both torsionally and laterally (Wang and Giaever 1988), and looping is typically aided by proteins that bind to specific sequences on the DNA and bend it with a characteristic stereospecificity (Perez-Martin et al. 1994). However, at larger distances, the intrinsic flexibility of the DNA allows the formation of loops in the absence of a DNA-bending protein (Bellomy and Record 1990). These loops, which lack the stereospecificity provided by a specific, protein-induced DNA bend, may result in inappropriate protein–protein contacts. Thus, specific bends may act not only to facilitate correct protein–protein interactions, but also to prevent the stable formation of loops that result in inappropriate protein–protein interactions.
Transcription by prokaryotic RNA polymerase (RNAP) holoenzyme containing the alternate sigma factor, σ54, requires contact between an enhancer-binding protein (EBP) bound at upstream activation sites (UAS) and the holoenzyme (Buck et al. 1986; Reitzer and Magasanik 1986; Ninfa et al. 1987). Through an ATP-hydrolysis-dependent mechanism, activation by the EBP transforms the closed complex formed by σ54–RNAP at the promoter to an open complex permissive for transcriptional initiation (Popham et al. 1989). A DNA loop is thought to facilitate this interaction by increasing the local concentration of the EBP in the vicinity of the RNAP holoenzyme (Buck et al. 1987; Su et al. 1990; Wedel et al. 1990; Rippe et al. 1997).
Many prokaryotic species, including Escherichia coli, have multiple EBPs with distinct regulatory roles (Morett and Segovia 1993; Kaufman and Nixon 1996). Their specificity is thought to be largely a consequence of binding to a particular associated UAS sequence upstream of their target promoter(s) through a carboxy-terminal DNA-binding domain (Morett et al. 1988). While DNA binding is not essential for activation, mutant EBPs lacking the DNA-binding domain require far higher concentrations and show a loss of specificity (Dworkin et al. 1997; North and Kustu 1997).
Integration host factor (IHF), a heterodimeric protein that bends DNA by >160°, (Rice et al. 1996) binds in the promoter region of many σ54-dependent operons at a site typically located between the promoter and the UAS sites (Gralla and Collado-Vides 1996). The sharp bend generated by IHF is thought to facilitate the interaction between the UAS-bound EBP and the σ54-RNAP holoenzyme, particularly at weaker promoters with a lower affinity for the σ54-RNAP holoenzyme (Hoover et al. 1990; Santero et al. 1992). This facilitation depends on the specific geometry of the interaction: If the UAS sites are moved so that the EBP binds on the opposite face of the DNA, then IHF inhibits activation (Claverie-Martin and Magasanik 1992). EBPs can activate from templates missing specific UAS sites; however, this activation is weaker than activation from wild-type templates and it is also inhibited by IHF (Claverie-Martin and Magasanik 1992; Dworkin et al. 1997). By contrast, activation by an EBP lacking a DNA-binding domain is unaffected by IHF (Dworkin et al. 1997); thus, IHF-mediated inhibition is presumably the result of EBPs binding to nonspecific (or cryptic) sites that are not appropriately spaced relative to the IHF bend and the σ54-RNAP bound at the promoter.
The hycA and hypA promoters of the formate hydrogenylase system of E. coli are regulated by IHF and the EBP, FhlA. In the presence of nitrate, expression from these promoters is significantly reduced, and strains lacking IHF (carrying a himA mutation) show an increase (three- to fourfold) in transcription from these promoters over wild-type strains (Hopper et al. 1994). In addition, a Pseudomonas putida strain carrying a himA mutation shows increased basal activation of the σ54-dependent Pu promoter in the absence of the cognate EBP, XylR, and this increase was attributed to activation by heterologous EBPs that might be normally inhibited by IHF (Perez-Martin and de Lorenzo 1995). Similarly, we observed that pspA transcription measured under noninducing conditions (using a pspA–lacZ promoter fusion) in a strain carrying a himA mutation, along with a deletion of the gene encoding the associated EBP, PspF, showed a twofold increase over a strain carrying only the pspF deletion (Dworkin 1997). We, as well as the previous investigators, were unable, however, to attribute this increased basal expression to activation by a particular EBP. We therefore decided to examine whether, in vitro, IHF could inhibit activation by a specific heterologous EBP and whether this inhibition would result in an increase in the specificity of transcriptional activation.
Results
The pspA gene of E. coli is transcribed by σ54–RNAP (Weiner et al. 1991) and is under control of the constitutively active EBP PspF (Jovanovic et al. 1996; Model et al. 1997). The pspA promoter region contains two binding sites for PspF (Jovanovic 1997), as well as a binding site for IHF (Fig. 1A,B) (Weiner et al. 1995). In vitro transcriptional activation by PspF at the pspA promoter is facilitated threefold by IHF (Fig. 2A, lanes 3,4; see also Dworkin et al. 1997). We replaced PspF with the heterologous EBP NRI of E. coli. Under conditions of nitrogen limitation, phosphorylated NRI activates transcription at several σ54-dependent promoters involved in the metabolism of nitrogen-containing compounds (Ninfa and Magasanik 1986). While phosphorylated NRI can activate transcription at the pspA promoter (Fig. 2A, lane 1), IHF inhibits this activation fivefold (lane 2).
Figure 1.

Organization of σ54-dependent promoters and upstream activation sequences. (A) The pspA promoter contains an IHF-binding site (−30 to −65) (Weiner et al. 1995) and two high-affinity binding sites (−89 to −126) for the PspF transcriptional activator (Jovanovic 1997). The glnH promoter contains an IHF-binding site (−33 to −59), two overlapping high-affinity binding sites for the NRI transcriptional activator (−100 to −129), and two lower affinity NRI sites (not shown) (Claverie-Martin and Magasanik 1991). The glnAp2 promoter contains two high affinity NRI sites (−100 to −147) as well as three weaker NRI sites (not shown) (Reitzer and Magasanik 1986). The pspA–ΔUAS promoter is identical to the pspA promoter except that the sequences spanning the PspF binding sites were deleted (Dworkin et al. 1997). (B) The upstream activation sequences (in bold) of the promoters schematized in A. Note that the glnH UAS sequences are overlapping and that, whereas the glnH and glnAp2 UAS sequences are similar, they both differ from the pspA UAS sequences.
Figure 2.
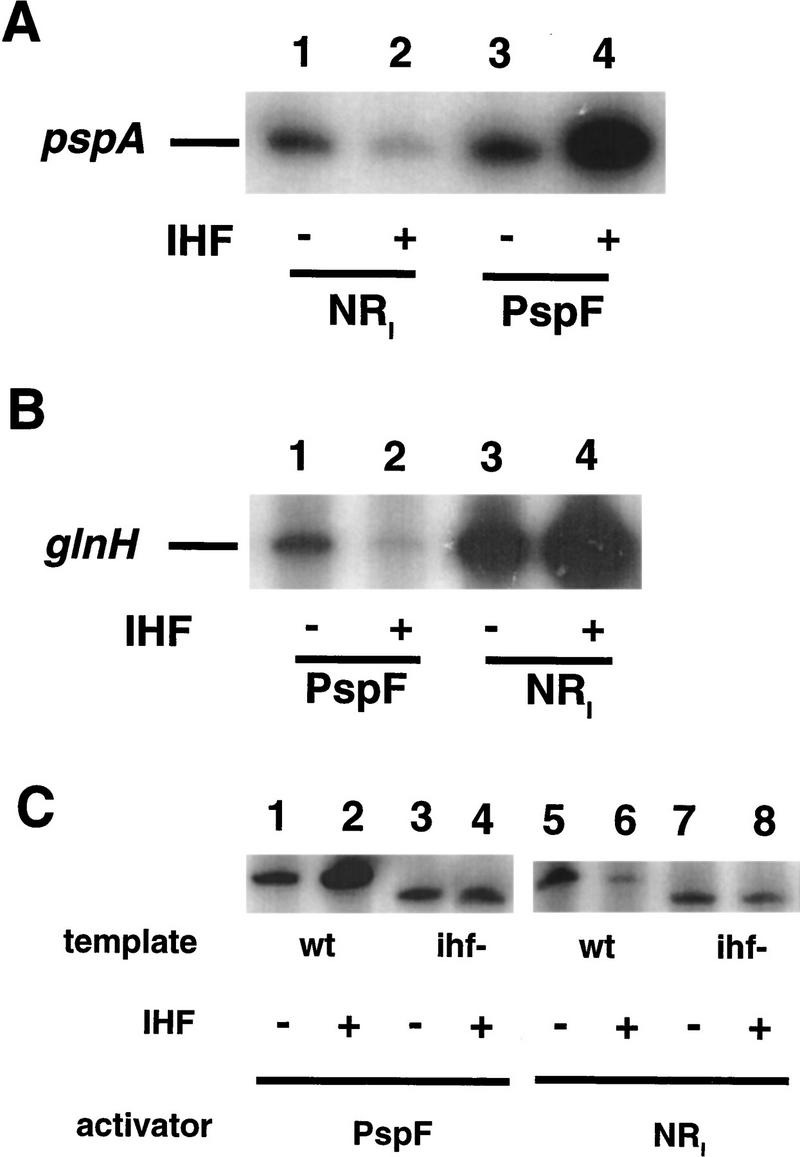
Effect of IHF on PspF- and NRI-dependent in vitro transcription from pspA and glnH promoters. (A) NRI- or PspF-dependent activation assayed on a supercoiled pspA promoter template (pJD10) in the presence or absence of IHF. (B) PspF- or NRI -dependent activation assayed on a supercoiled glnH promoter template (pJD37) in the presence or absence of IHF. In the absence of any EBP, pspA- or glnH-specific transcription is abolished (Claverie-Martin and Magasanik 1991; Dworkin et al. 1997). (C) NRI- or PspF-dependent activation assayed on a supercoiled pspA promoter template (wt; pJD10) or on a supercoiled pspA promoter template lacking the IHF binding site (ihf−; pJD28). Quantified RNA transcripts (in normalized arbitrary units): (A) (Lane 1) 1.0; (lane 2) 0.19; (lane 3) 2.3; (lane 4) 6.4. (B) (Lane 1) 1.0; (lane 2) 0.41; (lane 3) 1.4; (lane 4 5.3.
This inhibition suggests that NRI is bound to site(s) in the pspA promoter region that do not result in an appropriate EBP–IHF–RNAP geometry. The ability of NRI to activate at low concentrations (<100 nm) from templates lacking specific NRI binding sites has been attributed to its ability to bind to the DNA nonspecifically (Weiss et al. 1992). In addition, since EBPs can act at distances of up to 2 kb upstream (or downstream) of their target promoters (Buck et al. 1986; Ninfa et al. 1987), the possible sites are not restricted to those in near proximity to the RNAP bound at the promoter, as is the case for another prokaryotic transcriptional activator, CAP (Busby and Ebright 1994).
In an effort to extend the generality of this observation, we examined the E. coli σ54-dependent glnH promoter. The glnH promoter region contains two strong and two weak binding sites for NRI as well as an IHF site (Fig. 1A,B) (Claverie-Martin and Magasanik 1991). As has been demonstrated (Claverie-Martin and Magasanik 1991), NRI activates transcription from the glnH promoter in vitro and this activation is facilitated fourfold by IHF (Fig. 2B, lanes 3,4). By contrast, PspF-dependent activation of the glnH promoter is inhibited 2.5-fold by IHF (Fig. 2B, lanes 1,2). This inhibition suggests that PspF is binding to site(s) that result in a geometry of the EBP–IHF–RNAP interaction that is unfavorable for activation. The NRI and PspF UAS sites are distinct (Fig. 1B) and neither the pspA nor the glnH promoter region contains sequences matching the heterologous UAS sites.
When the sequence comprising the IHF site in the pspA promoter is replaced with a sequence lacking an IHF site, both the stimulatory effect of IHF on PspF-dependent activation (Fig. 2C, lanes 5–8) and the inhibitory effect of IHF on NRI-dependent activation (Fig. 2C, lanes 1–4) are eliminated. Thus, IHF inhibition of NRI-dependent pspA transcription is not a consequence of binding site competition or protein–protein interaction between IHF and NRI.
The opposite effects of IHF on activation by two different EBPs at a single promoter suggest that IHF could help determine the specificity of activation. That is, in addition to the specificity resulting from EBP binding to a cognate UAS, the bend generated in the promoter region by the binding of IHF would prevent activation by EBPs bound nonspecifically. We examined this question using in vitro transcription reactions containing multiple templates with different promoters. In the absence of IHF, the EBP NRI activates transcription from promoters containing either PspF-specific (pspA) or NRI-specific (glnH and glnA) binding sites comparably (Fig. 3A, lane 1). We take as a measure of specificity the ratio of appropriate to inappropriate transcripts, which, in this case (no IHF), is the ratio glnH : pspA = 1.6 : 1. In the presence of IHF (lane 2), however, this ratio increases tenfold to 15 : 1. This increase is a result of IHF acting both to facilitate NRI-dependent activation of glnH and to inhibit NRI-dependent activation of pspA. The glnA promoter contains no IHF-binding site (Fig. 1C) and thus is unaffected by IHF (Fig. 3A).
Figure 3.

IHF increases specificity of activation by PspF and NRI. (A) NRI-dependent activation assayed on supercoiled templates (3 nm) containing the pspA promoter (pJD10), the glnH promoter (pJD37), and the glnAp2 promoter (pTH8) in the presence or absence of IHF. The specific transcripts generated from each of these templates are identified. (B) PspF-dependent activation assayed on supercoiled templates (3 nm) containing the pspA promoter (pJD10) and the glnH promoter (pJD37) in the presence or absence of IHF. (C) PspF-dependent activation assayed on supercoiled templates (3 nm) containing the pspAΔUAS promoter (pJD12) and the glnH promoter (pJD37) in the presence or absence of IHF. Quantified RNA transcripts (in decreasing order of size and in normalized arbitrary units): (A) (Lane 1) 0.61; 1.0, 0.75; (lane 2) 0.11, 1.6, 0.85; (B) (Lane 1) 1.0, 0.33; (lane 2) 1.8, 0.13; (C) (Lane 1) 1.0, 0.63; (lane 2) 0.29, 0.23.
When the EBP is PspF, a similar effect of IHF is seen. In this case, the appropriate transcript is from the pspA promoter and the inappropriate transcript is from the glnH promoter. Quantification of these transcripts yields the specificity ratio of pspA : glnH is 3 : 1 in the absence of IHF (Fig. 3B, lane 1), and 14 : 1 in its presence (Fig. 3B, lane 2). A comparison of the pspA and glnH transcripts in the presence and absence of IHF demonstrates that IHF acts to increase PspF-dependent activation of pspA while simultaneously inhibiting PspF-dependent activation of glnH.
When the PspF UAS sequences in the pspA promoter are removed, IHF inhibits PspF-dependent transcription (Dworkin et al. 1997). In a transcription reaction containing both this template (pspAΔUAS) and a glnH promoter template (Fig. 3C), there is no change in the specificity ratio pspAΔUAS : glnH in the presence of IHF. Thus, the effect of IHF to increase the specificity of activation is dependent on the presence of a specific UAS.
Discussion
IHF facilitates EBP-dependent activation of the weak, σ54-dependent nifH promoter and at such weak promoters, EBPs that do not have an appropriate binding site properly positioned to take advantage of the IHF-generated bend are unable to activate efficiently (Santero et al. 1992). However, not all promoters regulated by IHF are weak; for example, the two promoters analyzed here, pspA and glnH, are stronger than nifH, and, as predicted, are less dependent on IHF (Fig. 2A,B; threefold) than weaker promoters (e.g., nifH, >20-fold; Hoover et al. 1990). The potential susceptibility of stronger promoters to activation by heterologous activators raises the question of how specificity of activation is maintained (Hoover et al. 1990; Santero et al. 1990). The experiments presented here are consistent with a model in which activation by inappropriate EBPs bound to nonspecific sites is inhibited by IHF.
While binding of proteins to upstream (or downstream) sequences can increase their effective local concentration at the promoter (Mossing and Record 1986; Rippe et al. 1995), we would argue that binding of an EBP to a nonspecific site(s) in the presence of IHF results in a DNA geometry that decreases the local concentration of the EBP near the promoter. Previous explanations of IHF inhibition of background activation have focused on the formation of a specific complex higher-order structure resulting from the relatively unique juxtaposition of several σ54 promoters (Hopper et al. 1994) or on the ability of IHF to occlude access to the RNAP holoenzyme by interposing a segment of DNA that blocks interactions between an EBP acting from solution and the RNAP (de Lorenzo and Perez-Martin 1996; Perez-Martin and de Lorenzo 1995). While the data presented here do not directly evaluate the second model, heterologous EBPs, as DNA-binding proteins, can be assumed from thermodynamic principles to bind DNA nonspecifically (von Hippel et al. 1974; Lin and Riggs 1975). Further, IHF does not negatively regulate a mutant EBP lacking a DNA-binding domain (Dworkin et al. 1997).
A localized DNA bend can have implications for DNA structure beyond its direct effect; a DNA sequence with intrinsic curvature is sufficient to determine the plectonemic structure of a supercoiled plasmid (Laundon and Griffith 1988). In fact, DNA-bending proteins that act to inhibit specific loop formation include the Nac protein of Klebsiella aerogenes, which prevents the interaction of NRI bound at a specific enhancer in the nac promoter region with the σ54–RNAP holoenzyme (Feng et al. 1995), and the CAP protein, which disrupts the repression loop formed by AraC dimers bound simultaneously to two sites in the araBAD promoter region (Lobell and Schleif 1991). Thus, the bend generated by IHF acts both to increase contacts between UAS-bound EBPs and σ54–RNAP at weaker promoters and to reduce contacts between nonspecifically bound heterologous EBPs and the σ54–RNAP at stronger promoters.
These observations may be particularly relevant for the regulation of eukaryotic genes, where it has been suggested that protein–protein interactions between the basal transcriptional machinery and proteins bound to distal enhancers can be modulated by the topology of the intervening DNA (Echols 1986; Ptashne 1986). In the specific case of the chicken βA-globin promoter, enhancer-dependent in vitro transcription requires that the intervening DNA be supercoiled (Barton et al. 1997). Consistent with this observation, Monte Carlo-based simulations of DNA dynamics demonstrate that the effective local concentration of two sites on DNA is far greater on supercoiled DNA than on relaxed DNA (Vologodskii et al. 1992). This increase is largely unaffected by the contour separation between the sites (Vologodskii et al. 1992), suggesting that the basal transcription apparatus could be susceptible to inappropriate contacts from proteins bound at nonspecific sites located far upstream (or downstream). In vivo, where the chromosome is primarily supercoiled [and particularly upstream of promoters where transcription-induced negative supercoiling occurs (Wu et al. 1988)], mechanisms that restrict inappropriate protein–protein interactions must therefore exist.
The data presented here suggest that a general mechanism of negative regulation of loop formation may play an important role in determining the specificity of activation of promoters utilizing DNA looping. It is therefore of interest that the nucleosome appears to block access to promoters by transcription factors not by preventing binding, but instead through a distortion of the DNA structure (Luger et al. 1997).
Materials and methods
DNA manipulation
All DNA manipulations were conducted according to established protocols (Sambrook et al. 1989). All enzymes were obtained from New England Biolabs (Beverly, MA) unless noted. Primers were synthesized by Operon Technologies (Alameda, CA). Template DNA for in vitro transcription reactions was purified by use of either the Wizard Megaprep or Midiprep DNA purification systems (Promega).
Plasmids
pJD10 (pspA) contains a 580-bp PCR-generated fragment encompassing the pspA promoter region (−458 to +122) cloned into the vector pGZ119EH (Lessl et al. 1992) upstream of the rnnBT1 terminator (Dworkin et al. 1997). pJD12 (pspAΔUAS) is identical to pJD10 except that sequences −89 to −126 were replaced with an NdeI site (Dworkin et al. 1997). pTH8 (glnAp2) contains a 600-bp HaeIII fragment spanning the glnA promoter region cloned into the vector pTE103 upstream of the bacteriophage T7 early terminator (Hunt and Magasanik 1985). pJD37 (glnH) was constructed by cloning the EcoRI–HindIII fragment from pFC50 (Claverie-Martin and Magasanik 1991) spanning the glnH promoter into the EcoRI and HindIII sites of pGZ119EH. pJD28 (pspA −ihf) was constructed by PCR mutagenesis, which replaced the sequence −34 to −58 of the pspA promoter in pJD10 with a different sequence (5′-GGATCCTCTAGAGTCGACCTGCAG-3′) of the same length not containing an IHF-binding site. Primers JD54 (5′-GGCTGGTACCTAGCGAGTTCATCAAGAAATA-3′) and JD101 (5′-GGCGGATCCCTGATTGAAGAATCAACAGC-3′) were used in a PCR reaction with Taq polymerase and pBRPS-1 (Brissette et al. 1991) as template. The PCR product was cleaved with BamHI and KpnI and cloned into pGZ119EH cleaved with BamHI and KpnI. This plasmid was cleaved with PstI and HindIII and ligated to a fragment generated by PCR with primers JD102 (5′-GGCGGATCCCTGCAGGATAAAAAATTGGCACGCAAATTG-3′) and JD103 (5′-GGCAAGCTTCAGTTTCTGTGGATCTTCC-3′), Taq polymerase, and pJD10 as template, which was then cleaved with PstI and HindIII. The transcript from pJD28 is 28 nucleotides shorter than that from pJD10 as a result of the cloning strategy employed. The lengths of the RNA transcripts from the respective plasmids are pJD10 (pspA) ∼360 nucleotides; pJD37 (glnH) ∼340 nucleotides; pTH8 (glnAp2) ∼300 nucleotides; pJD12 (pspAΔUAS) ∼360 nucleotides; pJD28 (pspA −ihf) ∼330 nucleotides.
In vitro transcription
Reactions were carried out in a buffer containing 50 mm Tris-HCl (pH 7.5), 50 mm KCl, 10 mm MgCl2, and 0.1 mm EDTA. All reactions also contained 0.5 mm GTP, 0.5 mm UTP, 2 mm ATP, 2 mm DTT, and 0.3 units of recombinant RNAsin (Promega). Core RNAP (Epicentre Technology, Madison WI) and σ54 [purified according to Hunt and Magasanik (1985)] were added at 10 nm and 14 nm, respectively. When used, PspF (gift of G. Jovanovic) was at 4 nm, NRI and NRII [both purified according to Ninfa et al. (1987)] were at 100 nm each, and IHF (gift of H. Nash) was at 15 nm. Typically, all components were incubated with supercoiled template (5 nm) at 37°C for 10 min. Then 0.5 μl of [α-32P]CTP (10 μCi/μl; New England Nuclear) was added [except for reactions containing NRI where heparin (Sigma) was added to 100 μm simultaneously], and the reaction allowed to incubate 10 min more at 37°C. Cold CTP was added to 1 mm and reaction continued at 37°C for 10 min. Reactions then were placed on ice, and an aliquot was treated with phenol–chloroform and precipitated in ethanol with tRNA (50 μg/ml). The pellet was resuspended in RNAse-free glass-distilled H2O, mixed with formamide loading buffer, loaded on 4% polyacrylamide/7 m urea gels, and run at 225 V in 0.5× TBE as described (Sambrook et al. 1989). Gels were subjected to autoradiography or exposed to a Molecular Dynamics Storage Phosphor Screen and analyzed on a Molecular Dynamics PhosphorImager using ImageQuant software. Quantification of PhosphorImager data was as follows: A box was drawn around the band representing the transcription terminating at rrnBT1 and the pixel values were integrated. Background was calculated by integration of a same-sized box below the rrnBT1 band in each lane and subtraction from the rrnBT1 band.
Acknowledgments
We thank Goran Jovanovic and Howard Nash for proteins; Boris Magasanik for plasmids; Goran Jovanovic, Sydney Kustu, David Wilson, and John Marko for discussions; and Marjorie Russel for comments on the manuscript. This work was supported in part by an National Science Foundation (NSF) grant (MCB 93-16625). J.D. held an NSF Graduate Fellowship and was supported by National Institutes of Health Training Grant CA09673-19 and by a Norman and Rosita Winston Foundation Fellowship.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact.
Footnotes
This paper is dedicated to the memory of Brian Laurence Selsky.
E-MAIL dworkin@biosun.harvard.edu; FAX (617) 496-4642.
References
- Barton MC, Madani N, Emerson BM. Distal enhancer regulation by promoter derepression in topologically constrained DNA in vitro. Proc Natl Acad Sci. 1997;94:7257–7262. doi: 10.1073/pnas.94.14.7257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bellomy GR, Record MT., Jr Stable DNA loops in vivo and in vitro: Roles in gene regulation at a distance and in biophysical characterization of DNA. Prog Nucleic Acid Res Mol Biol. 1990;39:81–128. doi: 10.1016/s0079-6603(08)60624-8. [DOI] [PubMed] [Google Scholar]
- Brissette JL, Weiner L, Ripmaster TL, Model P. Characterization and sequence of the Escherichia coli stress-induced psp operon. J Mol Biol. 1991;220:35–48. doi: 10.1016/0022-2836(91)90379-k. [DOI] [PubMed] [Google Scholar]
- Buck M, Miller M, Drummond M, Dixon R. Upstream activator sequences are present in the promoters of nitrogen fixation genes. Nature. 1986;320:374–378. [Google Scholar]
- Buck M, Cannon W, Woodcock J. Transcriptional activation of the Klebsiella pneumoniae nitrogenase promoter may involve DNA loop formation. Mol Microbiol. 1987;1:243–249. doi: 10.1111/j.1365-2958.1987.tb00518.x. [DOI] [PubMed] [Google Scholar]
- Busby S, Ebright RH. Promoter structure, promoter recognition, and transcription activation in prokaryotes. Cell. 1994;79:743–746. doi: 10.1016/0092-8674(94)90063-9. [DOI] [PubMed] [Google Scholar]
- Claverie-Martin F, Magasanik B. Role of integration host factor in the regulation of the glnHp2 promoter of Escherichia coli. Proc Natl Acad Sci. 1991;88:1631–1635. doi: 10.1073/pnas.88.5.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ————— Positive and negative effects of DNA bending on activation of transcription from a distant site. J Mol Biol. 1992;227:996–1008. doi: 10.1016/0022-2836(92)90516-m. [DOI] [PubMed] [Google Scholar]
- de Lorenzo V, Perez-Martin J. Regulatory noise in prokaryotic promoters: how bacteria learn to respond to novel environmental signals. Mol Microbiol. 1996;19:1177–1184. doi: 10.1111/j.1365-2958.1996.tb02463.x. [DOI] [PubMed] [Google Scholar]
- Dworkin J. Mechanisms of transcriptional regulation of the pspA promoter of Escherichia coli. Ph.D. Thesis. New York, New York: Rockefeller University; 1997. [Google Scholar]
- Dworkin J, Jovanovic G, Model P. Role of upstream activation sequences and integration host factor in transcriptional activation by the constitutively active prokaryotic enhancer-binding protein PspF. J Mol Biol. 1997;273:377–388. doi: 10.1006/jmbi.1997.1317. [DOI] [PubMed] [Google Scholar]
- Echols H. Multiple DNA–protein interactions governing high-precision DNA transactions. Science. 1986;233:1050–1056. doi: 10.1126/science.2943018. [DOI] [PubMed] [Google Scholar]
- Feng J, Goss TJ, Bender RA, Ninfa AJ. Repression of the Klebsiella aerogenes nac promoter. J Bacteriol. 1995;177:5535–5538. doi: 10.1128/jb.177.19.5535-5538.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gralla J, Collado-Vides J. Organization and function of transcription regulatory elements. In: Neidhardt F C, et al., editors. Escherichia coli and Salmonella: Cellular and molecular biology. Washington, D.C.: ASM Press; 1996. pp. 1232–1245. [Google Scholar]
- Hoover TR, Santero E, Porter S, Kustu S. The integration host factor stimulates interaction of RNA polymerase with NIFA, the transcriptional activator for nitrogen fixation operons. Cell. 1990;63:11–22. doi: 10.1016/0092-8674(90)90284-l. [DOI] [PubMed] [Google Scholar]
- Hopper S, Babst M, Schlensog V, Fischer HM, Hennecke H, Bock A. Regulated expression in vitro of genes coding for formate hydrogenylase components of Escherichia coli. J Biol Chem. 1994;269:19597–19604. [PubMed] [Google Scholar]
- Hunt TP, Magasanik B. Transcription of glnA by purified Escherichia coli components: Core RNA polymerase and the products of glnF, glnG, and glnL. Proc Natl Acad Sci. 1985;82:8453–8457. doi: 10.1073/pnas.82.24.8453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jovanovic G. Activation of phage shock protein operon transcription in Escherichia coli. Ph.D. Thesis. New York, New York: Rockefeller University; 1997. [Google Scholar]
- Jovanovic G, Weiner L, Model P. Identification, nucleotide sequence, and characterization of PspF, the transcriptional activator of the Escherichia coli stress-induced psp operon. J Bacteriol. 1996;178:1936–1945. doi: 10.1128/jb.178.7.1936-1945.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaufman RI, Nixon BT. Use of PCR to isolate genes encoding sigma54-dependent activators from diverse bacteria. J Bacteriol. 1996;178:3967–3970. doi: 10.1128/jb.178.13.3967-3970.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kustu S, North AK, Weiss DS. Prokaryotic transcriptional enhancers and enhancer-binding proteins. Trends Biochem Sci. 1991;16:397–402. doi: 10.1016/0968-0004(91)90163-p. [DOI] [PubMed] [Google Scholar]
- Laundon CH, Griffith JD. Curved helix segments can uniquely orient the topology of supertwisted DNA. Cell. 1988;52:545–549. doi: 10.1016/0092-8674(88)90467-9. [DOI] [PubMed] [Google Scholar]
- Lessl M, Balzer D, Lurz R, Waters VL, Guiney DG, Lanka E. Dissection of IncP conjugative plasmid transfer: Definition of the transfer region Tra2 by mobilization of the Tra1 region in trans. J Bacteriol. 1992;174:2493–2500. doi: 10.1128/jb.174.8.2493-2500.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin S, Riggs AD. The general affinity of lac repressor for E. coli DNA: Implications for gene regulation in procaryotes and eucaryotes. Cell. 1975;4:107–111. doi: 10.1016/0092-8674(75)90116-6. [DOI] [PubMed] [Google Scholar]
- Lobell RB, Schleif RF. AraC-DNA looping: Orientation and distance-dependent loop breaking by the cyclic AMP receptor protein. J Mol Biol. 1991;218:45–54. doi: 10.1016/0022-2836(91)90872-4. [DOI] [PubMed] [Google Scholar]
- Luger K, Mader AW, Richmond R, Sargent DF, Richmond TJ. Crystal structure of the nucleosome core particle at 2.8 å resolution. Nature. 1997;389:251–260. doi: 10.1038/38444. [DOI] [PubMed] [Google Scholar]
- Matthews KS. DNA looping. Microbiol Rev. 1992;56:123–136. doi: 10.1128/mr.56.1.123-136.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Model P, Jovanovic G, Dworkin J. The Escherichia coli phage shock protein operon. Mol Microbiol. 1997;24:255–261. doi: 10.1046/j.1365-2958.1997.3481712.x. [DOI] [PubMed] [Google Scholar]
- Morett E, Segovia L. The sigma 54 bacterial enhancer-binding protein family: Mechanism of action and phylogenetic relationship of their functional domains. J Bacteriol. 1993;175:6067–6074. doi: 10.1128/jb.175.19.6067-6074.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morett E, Cannon W, Buck M. The DNA-binding domain of the transcriptional activator protein NifA resides in its carboxy terminus, recognises the upstream activator sequences of nif promoters and can be separated from the positive control function of NifA. Nucleic Acids Res. 1988;16:11469–11488. doi: 10.1093/nar/16.24.11469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mossing MC, Record MT., Jr Upstream operators enhance repression of the lac promoter. Science. 1986;233:889–892. doi: 10.1126/science.3090685. [DOI] [PubMed] [Google Scholar]
- Ninfa AJ, Magasanik B. Covalent modification of the glnG product, NRI, by the glnL product, NRII, regulates the transcription of the glnALG operon in Escherichia coli. Proc Natl Acad Sci. 1986;83:5909–5913. doi: 10.1073/pnas.83.16.5909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ninfa AJ, Reitzer LJ, Magasanik B. Initiation of transcription at the bacterial glnAp2 promoter by purified E. coli components is facilitated by enhancers. Cell. 1987;50:1039–1046. doi: 10.1016/0092-8674(87)90170-x. [DOI] [PubMed] [Google Scholar]
- North AK, Kustu S. Mutant forms of the enhancer-binding protein NTRC can activate transcription from solution. J Mol Biol. 1997;267:17–36. doi: 10.1006/jmbi.1996.0838. [DOI] [PubMed] [Google Scholar]
- Perez-Martin J, de Lorenzo V. Integration host factor suppresses promiscuous activation of the sigma 54-dependent promoter Pu of Pseudomonas putida. Proc Natl Acad Sci. 1995;92:7277–7281. doi: 10.1073/pnas.92.16.7277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez-Martin J, Rojo F, de Lorenzo V. Promoters responsive to DNA bending: A common theme in prokaryotic gene expression. Microbiol Rev. 1994;58:268–290. doi: 10.1128/mr.58.2.268-290.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popham DL, Szeto D, Keener J, Kustu S. Function of a bacterial activator protein that binds to transcriptional enhancers. Science. 1989;243:629–635. doi: 10.1126/science.2563595. [DOI] [PubMed] [Google Scholar]
- Ptashne M. Gene regulation by proteins acting nearby and at a distance. Nature. 1986;322:697–701. doi: 10.1038/322697a0. [DOI] [PubMed] [Google Scholar]
- Reitzer LJ, Magasanik B. Transcription of glnA in E. coli is stimulated by activator bound to sites far from the promoter. Cell. 1986;45:785–792. doi: 10.1016/0092-8674(86)90553-2. [DOI] [PubMed] [Google Scholar]
- Rice PA, Yang S, Mizuuchi K, Nash HA. Crystal structure of an IHF–DNA complex: A protein-induced DNA U-turn. Cell. 1996;87:1295–1306. doi: 10.1016/s0092-8674(00)81824-3. [DOI] [PubMed] [Google Scholar]
- Rippe K, von Hippel PH, Langowski J. Action at a distance: DNA-looping and initiation of transcription. Trends Biochem Sci. 1995;20:500–506. doi: 10.1016/s0968-0004(00)89117-3. [DOI] [PubMed] [Google Scholar]
- Rippe K, Guthold M, von Hippel PH, Bustamante C. Transcriptional activation via DNA-looping: Visualization of intermediates in the activation pathway of E. coli RNA polymerase-σ54holoenzyme by scanning force microscopy. J Mol Biol. 1997;270:125–138. doi: 10.1006/jmbi.1997.1079. [DOI] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: A laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Santero E, Hoover T, Kustu S. Mechanism of transcription from nif promoters: Involvement of IHF. In: Gresshoff PH, Roth LE, Stacey G, Newton WE, editors. Nitrogen fixation: Achievements and objectives. New York, NY: Chapman and Hall; 1990. pp. 459–466. [Google Scholar]
- Santero E, Hoover TR, North AK, Berger DK, Porter SC, Kustu S. Role of integration host factor in stimulating transcription from the sigma 54-dependent nifH promoter. J Mol Biol. 1992;227:602–620. doi: 10.1016/0022-2836(92)90211-2. [DOI] [PubMed] [Google Scholar]
- Schleif R. DNA looping. Annu Rev Biochem. 1992;61:199–223. doi: 10.1146/annurev.bi.61.070192.001215. [DOI] [PubMed] [Google Scholar]
- Su W, Porter S, Kustu S, Echols H. DNA-looping and enhancer activity: Association between DNA-bound NtrC activator and RNA polymerase at the bacterial glnA promoter. Proc Natl Acad Sci. 1990;87:5504–5508. doi: 10.1073/pnas.87.14.5504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vologodskii AV, Levene SD, Klenin KV, Frank-Kamenetskii M, Cozzarelli NR. Conformational and thermodynamic properties of supercoiled DNA. J Mol Biol. 1992;227:1224–1243. doi: 10.1016/0022-2836(92)90533-p. [DOI] [PubMed] [Google Scholar]
- von Hippel PH, Revzin A, Gross CA, Wang AC. Non-specific DNA binding of genome regulating proteins as a biological control mechanism: I. The lac operon: Equilibrium aspects. Proc Natl Acad Sci. 1974;71:4808–4812. doi: 10.1073/pnas.71.12.4808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang JC, Giaever GN. Action at a distance along DNA. Science. 1988;240:300–304. doi: 10.1126/science.3281259. [DOI] [PubMed] [Google Scholar]
- Wedel A, Weiss DS, Popham D, Droge P, Kustu S. A bacterial enhancer functions to tether a transcriptional activator near a promoter. Science. 1990;248:486–490. doi: 10.1126/science.1970441. [DOI] [PubMed] [Google Scholar]
- Weiner L, Brissette JL, Model P. Stress-induced expression of the Escherichia coli phage shock protein operon is dependent on σ54 and modulated by positive and negative feedback mechanisms. Genes & Dev. 1991;5:1912–1923. doi: 10.1101/gad.5.10.1912. [DOI] [PubMed] [Google Scholar]
- Weiner L, Brissette JL, Ramani N, Model P. Analysis of cis-acting elements regulating the stress-induced phage shock protein operon. Nucleic Acids Res. 1995;23:2030–2036. doi: 10.1093/nar/23.11.2030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss DS, Klose KE, Hoover TR, North AK, Porter SC, Wedel AB, Kustu S. Prokaryotic transcriptional enhancers. In: McKnight SL, Yamamoto KR, editors. Transcriptional regulation. Cold Spring Harbor, New York: Cold Spring Harbor Press; 1992. pp. 667–726. [Google Scholar]
- Wu HY, Shyy SH, Wang JC, Liu LF. Transcription generates positively and negatively supercoiled domains in the template. Cell. 1988;53:433–440. doi: 10.1016/0092-8674(88)90163-8. [DOI] [PubMed] [Google Scholar]