

Abstract
Guanosine 3′-di-5′-(tri)di-phosphate nucleotides [(p)ppGpp], synthesized in response to amino acid limitation, induce early gene expression leading to multicellular fruiting body formation in Myxococcus xanthus. A mutant (DK527) that fails to accumulate (p)ppGpp in response to starvation was found to be blocked in development prior to aggregation. By use of a series of developmentally regulated Tn5lac transcriptional fusion reporters, the time of developmental arrest in DK527 was narrowed to within the few hours of development, the period of starvation recognition. The mutant is also defective in the production of A-factor, an early extracellular cell-density signal. The relA gene from Escherichia coli, which encodes a ribosome-dependent (p)ppGpp synthetase, rescues this mutant. We also demonstrate that inactivation of the M. xanthus relA homolog blocks development and the accumulation of (p)ppGpp. Moreover, the wild-type allele of Myxococcus relA rescues DK527. These observations support a model in which accumulation of (p)ppGpp, in response to starvation, initiates the program of fruiting body development, including the production of A-factor.
Keywords: Guanosine tetraphosphate, Myxococcus xanthus development, RelA, stringent response, A-factor
When Myxobacteria are deprived of essential nutrients, they undergo a developmental program in which ∼100,000 cells aggregate to form a multicellular fruiting body. Individual cells within the fruiting bodies differentiate into environmentally resistant and metabolically quiescent myxospores. Previous work has demonstrated that carbon, nitrogen, or phosphate deprivation (Dworkin 1962; Wireman et al. 1977; Manoil and Kaiser 1980a; Manoil 1982; Shimkets 1984), but not purine or pyrimidine starvation (Kimsey and Kaiser 1991; Singer and Kaiser 1995) initiates fruiting body development. M. xanthus is bacteriolytic and feeds on the proteins, peptides, and amino acids of prey bacteria but is unable to utilize the sugars liberated because of the absence of certain glycolytic enzymes such as pyruvate kinase (Watson and Dworkin 1968). Studies probing the nutritional requirements of M. xanthus have demonstrated this organism’s preference for amino acids and small molecular weight carbon compounds such as pyruvate, acetate (Bretscher and Kaiser 1978), and to a lesser extent certain Kreb’s cycle intermediates (Watson and Dworkin 1968).
M. xanthus is unable to synthesize leucine, isoleucine, or valine. Starvation for any of these three essential amino acids, or of an auxotroph for its newly essential amino acid, or addition of a tRNA charging inhibitor (such as serine hydroxamate or tyrosinol) elicits a developmental response (Manoil and Kaiser 1980a,b). All of these conditions lead to a decrease in the corresponding charged tRNAs, suggesting a connection between the initiation of fruiting body development and a stringent response. The stringent response couples the availability of aminoacylated (charged) tRNA molecules to the rate of protein synthesis through the signaling molecules guanosine-5′-(tri)di-3′-diphosphate [(p)ppGpp] (for review, see Cashel et al. 1996). In Escherichia coli, for example, ribosomal and tRNA synthesis is immediately inhibited in response to amino acid limitation (Cashel 1969; Cashel and Gallant 1969; Stent and Brenner 1961). Accumulation of (p)ppGpp arrests synthesis of stable RNA, activates amino acid biosynthetic operons, increases proteolysis, and inhibits the synthesis of DNA, membranes, and cell walls (Cashel et al. 1996). In addition, recent work by Chakraburtty and Bibb (1997) has implicated (p)ppGpp as a positive regulator of antibiotic, pigment, and aerial mycelium formation in Streptomyces coelicolor A3(2).
Two independent lines of evidence support the hypothesis that (p)ppGpp is a general starvation signal for M. xanthus. First, Manoil and Kaiser (1980a,b) found that (p)ppGpp accumulates rapidly after transfer of growing cells to nutrient-limited conditions. Furthermore, all known conditions that initiate fruiting body development have been found to elicit an increase in intracellular (p)ppGpp concentration (Manoil and Kaiser 1980a,b). Second, we recently demonstrated that expression of the E. coli relA+ gene in M. xanthus, even in the presence of ample nutrients, resulted in a simultaneous increase in (p)ppGpp accumulation and activation of early developmentally specific gene expression (Singer and Kaiser 1995). The finding that ectopic expression of E. coli relA and subsequent (p)ppGpp accumulation induces development implies that this nucleotide is sufficient to initiate the developmental program.
Before this work, the question remained whether (p)ppGpp is also necessary for development. We have taken two approaches to address this question. First, we have further examined the developmental block associated with a known mutant, DK527, which has lost the ability to accumulate (p)ppGpp in response to starvation (Manoil and Kaiser 1980a,b). Second, we initiated a search for the M. xanthus homolog of relA, which is responsible in E. coli and Salmonella typhimurium for ribosome-dependent (p)ppGpp synthetase activity, and have characterized the phenotype of a disruption in this gene with respect to development.
Results
Mutant DK527, which fails to develop, uncouples RNA synthesis from amino acid availability
DK527 was derived from DK101 (Table 1), which is capable of responding to starvation by accumulating (p)ppGpp, and subsequently forming fruiting bodies with spores (Manoil and Kaiser 1980b). The failure of DK527 to accumulate (p)ppGpp after starvation parallels a similar failure in the relA− (relaxed) mutants of E. coli (Borek et al. 1956; Stent and Brenner 1961; Fiil and Friesen 1968; Cashel and Gallant 1969). If DK527 is the Myxococcus version of a relaxed mutant, its stable RNA synthesis should be uncoupled from amino acid availability. To test this deduction, the synthesis of RNA by M. xanthus DK101 and the DK527 mutant strain were compared with that of wild-type E. coli and an E. coli relA− mutant. As shown in Figure 1A, when wild-type M. xanthus cells are abruptly deprived of seryl-tRNA by the addition of serine hydroxamate, a competitive inhibitor of seryl-tRNA charging in E. coli (Tosa and Pizer 1971), within 30 min total RNA synthesis decreases to ∼50% of the pretreatment rate. Apparently, wild-type M. xanthus, like wild-type E. coli (Fig. 1B), interrupts stable RNA synthesis when seryl-tRNA is no longer available for protein synthesis. The response of M. xanthus is more gradual than that of E. coli, consistent with the ∼10-fold lower rate of cell growth and RNA synthesis in M. xanthus as compared with E. coli. Serine hydroxamate induces an increase in the intracellular level of (p)ppGpp in M. xanthus (Manoil and Kaiser 1980a; Singer and Kaiser 1995). Thus, it appears that wild-type M. xanthus, like E. coli, couples stable RNA synthesis to aminoacyl tRNA availability. In contrast, when the DK527 mutant was exposed to serine hydroxamate, total RNA synthesis continued at the pretreatment rate (Fig. 1A), like the E. coli relA mutant (Fig. 1B).
Table 1.
Strains and plasmids
| Plasmid
|
Construction or relevant markers
|
Source
|
|---|---|---|
| G4 | M. xanthus cosmid | R. Gill (University of Colorado, Health Sciences, Denver) |
| D10 | M. xanthus cosmid | R. Gill |
| pBGS18 | cloning vector carrying Kmr | Spratt et al. (1986) |
| pBluescript SK+ | cloning vector carrying Apr | Stratagene |
| pTTQ18 | Ptac expression vector | Amersham |
| pPLH323 | cloning vector carrying Kmr and Mx8attP site | P Hartzell (University of Idaho, Moscow) |
| pSWU29 | cloning vector carrying Tcr and Mx8attP site | S. Wu (Stanford University, CA) |
| pPCR-A | 311-bp PCR fragment cloned into pBGS18 | this study |
| pMS131 | control for pMS131, lacks E. coli relA | Singer and Kaiser (1995) |
| pMS132 | E. coli relA gene under Pcar control | Singer and Kaiser (1995) |
| pMS300 | 1.2-kb SacI fragment from pMS302 in pBGS18 | this study |
| pMS302 | 4.8-kb PstI fragment from G4 in pBGS18 | this study |
| pMS320 | 4-kb PstI–NcoI of pMS302 | this study |
| pMS321 | 4-kb PstI–HindIII fragment from pMS320 | this study |
| cloned into pSWU29 (Tcr) | ||
| pMS323 | 1-kb SacI–NcoI fragment from pMS320 into pBGS18 | this study |
| pMS325 | 480-bp EagI (filled in)–XhoI fragment cloned into HindIII (filled in)–XhoI sites of pMS321 (Tcr) | this study |
| pMS350 | 4.8-kb PstI fragment from G4 into pPLH343 | this study |
| pMS379 | 3.2-kb XhoI–PstI fragment into pBGS18 | this study |
| pMS380 | 267-bp BamHI–XhoI PCR fragment | this study |
| cloned into pMS379 | ||
| pMS381 | 3.4-kb BamHI–PstI fragment from pMS380 | this study |
| cloned into pTTQ18–RelA under Ptac control. | ||
| Strains | Relevant genotype | |
| M. xanthus | ||
| DK101 | sglA1 | D. Kaiser (Stanford University, CA) |
| DK527 | DK101 relA527 | Manoil and Kaiser (1980a) |
| DK10528 | DK527 pMS132 | this study |
| DK10529 | DK101 pMS132 | this study |
| DK480 | DK101asgB480 | Kuspa et al. (1986) |
| DK4324 | DK101asgB480 Ω4521 | Kuspa et al. (1986) |
| DK4521 | Ω4521 | Kroos et al. (1986) |
| DK4400 | Ω4400 | Kroos et al. (1986) |
| DK4408 | Ω4408 | Kroos et al. (1986) |
| DK4455 | Ω4455 | Kroos et al. (1986) |
| MS8 | DK101 relA∷pMS302 (tandom duplication of relA region) | this study |
| MS9 | DK527relA∷pMS302 (tandom duplication of relA region) | this study |
| MS10 | DK101 relA∷pMS300 | this study |
| MS11 | DK101 pMS321 | this study |
| MS12 | MS10 pMS321 | this study |
| MS13 | DK527 pMS321 | this study |
| MS15 | DK527 pMS325 | this study |
| MS16 | DK101 pMS325 | this study |
| E. coli | ||
| MG1655 | wild type | B. Bachman (Yale University, New Haven, CT) |
| CF1651 | relAΔ251∷kan | M. Cashel (National Institutes of Health, Bethesda, MD) |
| MS-Ec12 | MG1655 pMS381 | this study |
| MS-Ec51 | CF1651 pMS381 | this study |
Figure 1.
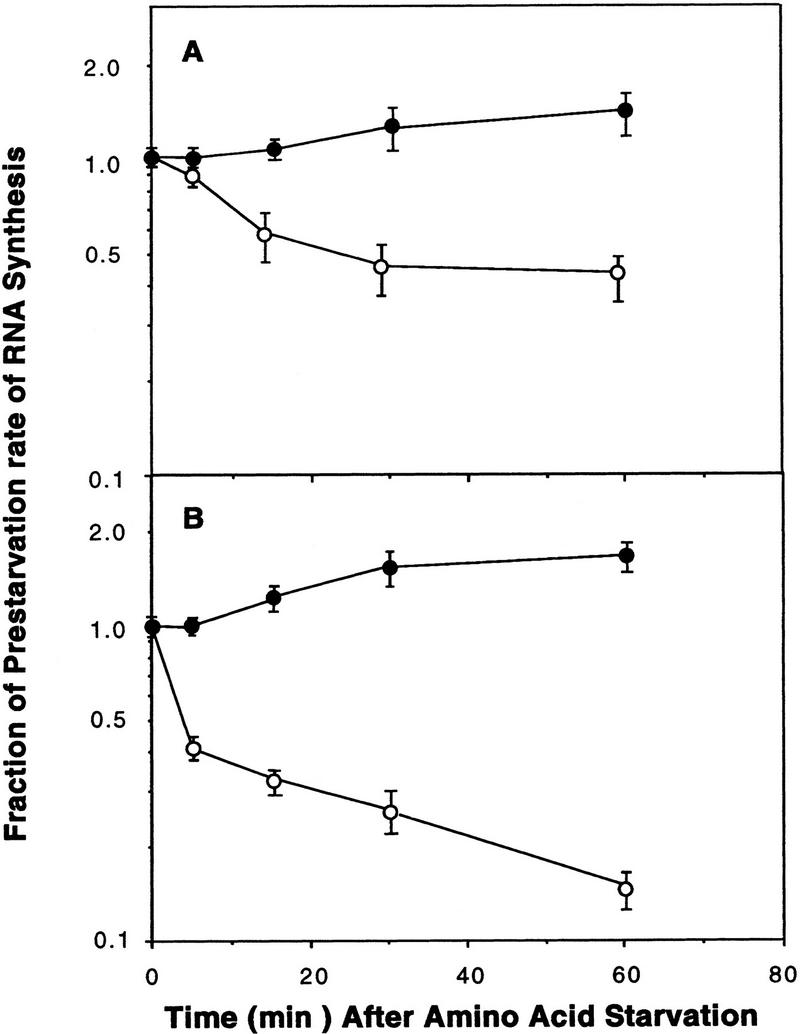
Effects of serine hydroxamate-induced serine starvation on RNA synthesis in M. xanthus, DK101 (○) or DK527 (•) (A), and E. coli, wild-type (○) or relA− (•) (B). Strains were grown at 30°C in either M9–glucose (E. coli) to an OD450 of 0.2 to 0.3 or in M1 medium (M. xanthus) at 33°C to a density of 60–100 Klett units, as described in Materials and Methods. Samples were pulse-labeled at the indicated times (in min) with [3H]uridine, and RNA synthesis rates were determined as described in Materials and Methods. Each time point records an average of three independent experiments.
The E. coli relA gene rescues the developmental defect of DK527
The failure of DK527 to develop fruiting bodies in response to starvation accompanies its failure to accumulate (p)ppGpp. To determine whether the failure to develop results from the inability to accumulate (p)ppGpp, the E. coli relA gene was tested for its ability to restore development in DK527. M. xanthus DK101 and DK527 strains were transformed with plasmid pMS132, which carries the E. coli relA gene under the transcriptional control of the light-inducible M. xanthus carQRS promoter (Singer and Kaiser 1995). To provide negative controls, these same strains were transformed with plasmid pMS131, which is identical to pMS132 but lacks the E. coli relA gene (Singer and Kaiser 1995). Both plasmids (pMS131 and pMS132) have the myxophage Mx8 attachment site Mx8attP, which provides high-efficiency, site-specific integration at the chromosomal Mx8 attB site (Orndorff et al. 1984), and the nptII gene encoding kanamycin resistance. Following transformation, the presence of a single integrated copy of pMS131 or pMS132 in the M. xanthus genome was confirmed by Southern blot (data not shown). Inducible expression of E. coli RelA protein was demonstrated by Western blotting cultures with or without exposure to light (Fig. 2).
Figure 2.

Expression of E. coli RelA in M. xanthus strains DK101 and DK527 under starvation in the dark (time 0) and after 5 hr of exposure to light. Western blotting is described in Materials and Methods. Arrows indicate the expected mobility of E. coli RelA protein.
Plasmid-carrying derivatives of DK101 and DK527 were also examined for the initiation of fruiting body development when bacteria were starved and expression of E. coli RelA was induced by exposure to light. The M. xanthus DK527 strain containing the E. coli relA+ gene regained the ability to form mounds when placed on starvation medium in the presence of light; plasmid-containing DK527 cells lacking E. coli relA+ were, like DK527 itself, unable to form mounds. DK101 strains, with or without the E. coli relA+ gene, were unaffected with respect to mound formation. It should be mentioned that under these conditions, development is inhibited at the mound stage (Li et al. 1992; Singer and Kaiser 1995).
Western Blot analysis (Fig. 2) shows that cultures before exposure to light (0 hr) were expressing a small amount of E. coli RelA protein; this lower level of E. coli RelA protein was sufficient to rescue fruiting body development of DK527 under starvation conditions. Cultures carrying the E. coli relA+ gene were starved in the dark; they too had regained the ability to form fruiting bodies and to sporulate. DK527 strains carrying the control plasmid pMS131 (no E. coli relA+) remained unable to form fruiting bodies. In addition to regaining the ability to develop in response to starvation, DK527 strains carrying E. coli relA+ regain the ability to accumulate (p)ppGpp in response to starvation. This experiment implies that a relatively small amount of E. coli RelA protein is sufficient to produce the (p)ppGpp needed to rescue DK527.
The DK527 mutant grows slowly, even in complete medium. Long term cultures accumulate strains—presumably with suppressor mutations—that can grow faster than DK527. Therefore, it was necessary to test whether the recovery of development in the strains that carried the E. coli relA+ gene might be the result of a second-site suppressor. To check this possibility, the plasmid pMS132 in strain DK10528 (DK527 pMS132) was replaced with pMS133, which lacks the E. coli relA+ gene but encodes tetracycline resistance (Tcr) (Singer and Kaiser 1995). Replacement was achieved by transduction and homologous recombination. Ten Tcr transductants of DK10528 were then screened with an E. coli relA+ gene-specific probe by Southern blot (data not shown). All ten transductants had lost the E. coli relA gene. Seven of the ten transductants had lost the ability of DK10528 to develop, returning to the phenotype of DK527. Because seven of the ten transductants of DK10528 lost the ability to develop following exchange of pMS133 for pMS132, it is clear that the developmental competence of DK10528 was associated with the relA+ gene and not with a second-site suppressor elsewhere in the genome. The three Tcr transductants that retained the Agg+ Fb+ phenotype may be caused by suppressor(s) that had arisen while those transductants grew on their Tc selection plate. These experiments confirm that rescue of the DK527 mutation by the E. coli relA+ gene is the result of complementation by the gene and its protein product.
DK527 fails to express early developmental markers
DK527 cannot undergo aggregation (Manoil and Kaiser 1980b), but wild-type cells (DK101) show their first morphological signs of aggregation at ∼6 hr postinitiation. To define more precisely the stage at which the mutation in DK527 causes it to deviate from the normal developmental program, four well-characterized Tn5lac transcriptional fusion reporters, Ω4408, Ω4455, Ω4521, and Ω4400, representative of the early stages of development were examined in the DK527 genetic background. These reporters begin their β-galactosidase expression at different fusion-specific times in the 0–6 hr interval. No increase in β-galactosidase expression from any of these four reporter fusions was observed in DK527, as compared with DK101 (Fig. 3). Thus, the mutation in DK527 prevents cells from initiating early gene expression as defined by these Tn5lac reporters. These experiments confine the temporal block to within the first few hours of the normal developmental program. Because (p)ppGpp is synthesized then, this is expected to be the time when starvation is recognized and the decision to develop fruiting bodies is made.
Figure 3.

β-Galactosidase expression from Tn5lac fusions 4455, 4400, 4408, and 4521 after starvation in strains DK101 (○) and DK527 (•). Measurement of β-galactosidase specific activity is described in Materials and Methods; each point is an average of at least three independent experiments.
Cloning the M. xanthus RelA homolog
On the basis of the phenotype of DK527 and its complementation by the E. coli relA gene, a search for the M. xanthus relA homolog was initiated by use of a PCR-based approach. Taking advantage of regions of amino acid identity between the known amino acid sequences of RelA proteins from several different Eubacteria (Metzger et al. 1988; Sarubbi et al. 1989; Chakraburtty et al. 1996), PCR primers were designed to amplify DNA sequences encoding the M. xanthus relA homolog. A single 311-bp PCR product was detected by use of primers R-1750 and X-1125 (see Materials and Methods). DNA sequence analysis of this fragment predicted a protein coding sequence that was 49% identical and 75% similar to both the E. coli RelA and SpoT proteins, which are very similar to each other (Chakraburtty et al 1996).
The 311-bp PCR-generated fragment was used as a probe to screen an M. xanthus DNA cosmid library (generously provided by R. Gill, University of Colorado Health Sciences, Denver). This screen identified two cosmid clones, G4 and D10. Subsequent restriction endonuclease mapping, and Southern blotting of these cosmids revealed that they contained overlapping DNA and that the M. xanthus relA homolog resided on a common 4.8-kb PstI fragment (data not shown). This 4.8-kb PstI fragment was cloned into pBGS18; a restriction map of this region is diagrammed in Figure 4A. The sequence of the fragment has been submitted to Genbank, accession no. AF025847.
Figure 4.



Analysis of the M. xanthus relA locus. (A) Restriction map of the M. xanthus relA region. Arrows above the boxes represent orientation of each gene and putative ORF. DNA sequences contained in plasmids used in this work are designated by lines below the restriction map followed by the plasmid designation (see Materials and Methods for construction details). The black box in pMS381 designates the Ptac promoter region used to drive the relA gene. Forked arrowheads represent primers used for primer extensions and for PCR construction of pPCR-A. (B) (See facing page.) Sequence comparison between M. xanthus RelA and six other species. Alignments were performed by use of Macaw v. 2.0.5 from the National Center for Biotechnology Information, National Library of Medicine. Black boxes represent identical matches in all seven proteins; shaded boxes represent either five of seven matches or similar amino acid substitutions. (M.x.) M. xanthus; (E.c.) E. coli; (V.s.) Vibrio sp.; (H.i.) H. influenzae; (Sn.) Synechocystis sp.; (S.c.) S. coelicolor; (S.e.) S. equi.
The 4.8-kb PstI fragment is predicted to encode three ORFs, designated ORF 1–3 (Fig. 4A). ORF 2 exhibits strong similarity to RelA and SpoT; Figure 4B shows sequence similarity between ORF 2 and several bacterial RelA proteins. ORF 3 shows similarity to a group of small acid-soluble proteins implicated in translation inhibition, the YER057c/YJGF family (Oka et al. 1995; Ceciliani et al. 1996; Schmiedeknecht et al. 1996). Both putative ORFs would be transcribed in the same direction. Sequence analysis of DNA 5′ to the putative relA/spoT gene (which includes the putative ORF 1) shows no significant similarity to anything in the database when translated in all six potential reading frames.
Primer extension analysis was used to identify the transcriptional start site for the relA/spoT gene; a single 5′ end was identified (Fig. 5A). On the basis of the results of these experiments, on sequence similarity, and on the availability of a promising Shine–Dalgarno site nearby, we have placed the putative translational start at a GTG located 106 bp downstream from the transcriptional start site (Fig. 5A); this relA gene would encode a protein of 757 amino acids with a formal molecular mass of 85 kD. Sequence analysis also revealed a potential 8-bp DNA stem and 3-bp loop structure located between relA and dfrA (ΔG = −18.3 kcal), followed by a run of three T residues (Fig. 5B). This putative stem-loop structure is located 15 bp from the presumed translational start of dfrA and could represent a transcriptional terminator for the relA gene.
Figure 5.

(A) Nucleotide sequence of the 5′-region of relA. Transcriptional start site for relA is indicated by +1 and bold caps. The −10 and −35 regions are underlined, and the putative ribosome binding site is indicated by bold caps. (PE) Primer extension product; (T, G, C, A) DNA sequence reactions with the same primer used in the primer extension analysis as indicated in Materials and Methods; (Control) tRNA control primer extension reaction. (B) Nucleotide sequence of the intergenic region between relA and dfrA. A putative 9-bp stem 3-bp loop structure, which may act as a transcriptional terminator, is shown.
Locating the relA gene on the M. xanthus physical map
By use of the 311-bp PCR-generated fragment described above as a probe for Southern blot analysis of M. xanthus chromosomal DNA, two unique restriction fragments were identified. This implies that M. xanthus, like E. coli, may have one relA gene and one spoT gene. In E. coli, the relA and spoT genes are physically separated and map at 60 and 82 min, respectively. To determine whether the two putative M. xanthus genes were physically separated, their locations were mapped to the physical M. xanthus genome map (Shimkets 1993). The PCR-generated fragment was used as a probe to determine the position of each putative gene by use of pulsed-field electrophoresis (Chen et al. 1990). This analysis placed one of the genes at position 1–1.3Mb (AseI fragment N and SpeI fragment U′) and the other at position 4.8–5Mb (AseI fragment P and SpeI fragment S) on the 9.5-Mb M. xanthus physical map (Fig. 6). To determine the precise location of the cloned gene, the entire 4.8-kb PstI fragment was used to probe DNA isolated from an ordered M. xanthus YAC library (Kuspa et al. 1989). Hybridization results indicated that the cloned gene resides on YACs 1372 and 1334, placing it between tgl and 442 on the physical map (Fig. 6).
Figure 6.

Physical mapping of the relA gene. (A) Slot blots using total YAC DNA from an ordered YAC library, probed with DNA fragment harboring the M. xanthus relA gene. (B) Schematic of the 3.6- to 4.8-Mb region of the physical map of M. xanthus. YACs covering this region are indicated and labeled at the bottom. The position of relA is designated by the shaded box. The top line represents AseI and SpeI fragments as designated by Chen et al. (1990); the second line represents the position of previously mapped markers (Shimkets 1993).
The cloned gene has RelA function
Because ORF 2 displays sequence similarity to both RelA and SpoT proteins, a functional assay was required to indicate its role. For this purpose, we tested the M. xanthus clone for its capacity to complement an E. coli relA deletion strain to produce (p)ppGpp. Plasmid pMS381, which contains the M. xanthus gene under the transcriptional control of the IPTG inducible tac promoter, was introduced into a ΔrelA E. coli strain, CF165 (Xiao et al. 1991). Cells carrying either pMS381 or pTTQ18 were assayed for their ability to produce (p)ppGpp in response to IPTG. Cells carrying pMS381 were able to produce (p)ppGpp in an IPTG-dependent fashion (Fig. 7), demonstrating that expression of the M. xanthus gene correlates with an increase in the production of this nucleotide. Strains carrying the control plasmid pTTQ18 showed no increase in (p)ppGpp levels (Fig. 7). Previously, the overproduction of E. coli relA in both E. coli (Schreiber et al. 1991) and M. xanthus (Singer and Kaiser 1995) have been shown to increase the level of (p)ppGpp.
Figure 7.

Expression of M. xanthus relA in E. coli leads to (p)ppGpp production. One-dimensional thin layer chromatography autoradiographs of extracts from E. coli strains carrying either pMS381 (M. xanthus relA) or pTTQ18 (vector), with equal radioactivity loaded at the origin in each lane.
Disruption of relA prevents the accumulation of (p)ppGpp in response to amino acid starvation and causes an early developmental arrest
To assess the role of relA in (p)ppGpp synthesis and development in M. xanthus, the gene was disrupted by insertional mutagenesis. An internal 1.2-kb SacI fragment (Fig. 4A) cloned to the Kmr plasmid pBGS18, designated pMS300, was integrated at the chromosomal locus by homologous recombination. Integration of the plasmid at the chromosomal relA locus would result in a tandem duplication of the 1.2-kb region and generate two truncated genes. The structure of the expected tandem duplication was confirmed by Southern blot analysis (data not shown) and the resulting strain was designated MS10.
This relA insertion mutant, MS10, along with its isogenic parent, DK101, were assayed for their ability to synthesize (p)ppGpp in response to amino acid deprivation. Under these conditions DK101 accumulates (p)ppGpp within the first 15 min of starvation and the levels slowly decay (Fig. 8), consistent with previous studies (Manoil and Kaiser 1980a; Singer and Kaiser 1995). In contrast, strain MS10 does not accumulate (p)ppGpp even after 60 min (Fig. 9).
Figure 8.

Disruption of the M. xanthus relA gene prevents the accumulation of (p)ppGpp. One dimensional thin layer chromatography autoradiograph of extracts from M. xanthus strains harboring either pMS300 (MS10) or no insertion (wild type, DK101), with equal radioactivity loaded at the origin.
Figure 9.

Rescue of M. xanthus DK527 fruiting body formation by wild-type M. xanthus relA. Strains DK101 and DK527 with no plasmid (A,B) or with pMS325 (M. xanthus relA integrated at the Mx8att site) (C,D) were spotted on TPM agar as described in Materials and Methods and photographed at 72 hr poststarvation.
Previously, we have proposed that for the purpose of initiating fruiting body development, M. xanthus senses its nutritional status by measuring its intracellular levels of (p)ppGpp and thereby monitoring its capacity for protein synthesis. It follows that cells unable to produce (p)ppGpp will be unable to initiate development. DK101 and MS10 strains were examined for the ability to form fruiting bodies and spores. No fruiting bodies, mounds, or aggregates were obtained from the MS10 relA insertion strain, even after 5 days. Wild-type cells form fruiting bodies within 24 hr. Furthermore, when spore assays were performed after 24, 72, and 120 hr, the relA insertion strain produced <0.01% of the wild-type viable spore level.
Because dfrA lies 72-bp downstream to relA, it waspossible that these two genes constituted a single operon and that the insertion into the upstream relA gene also inactivated dfrA. Two approaches were taken to demonstrate that the developmental phenotype associated with MS10 is caused by a disruption of relA and not the result of a polar effect on the downstream dfrA gene. In the first approach, we disrupted the intercistronic region between dfrA and relA genes by inserting plasmid pMS323 into DK101. This plasmid contains a 1-kb SacI–NcoI fragment carrying the 3′-end of relA and 5′-end of the dfrA gene (Fig. 4A). Integration of this plasmid would duplicate this region, producing one complete relA gene and one complete dfrA gene that are separated by vector sequences. If dfrA is expressed from the relA promoter, the only intact dfrA gene would now be separated from its promoter. Alternatively, if dfrA is being expressed from its own promoter or is not essential for development, no developmental phenotype should be observed. The resulting strain, designated MS11, was assayed for development along with strains MS10 and DK101. MS11 developed and sporulated normally (unlike MS10), suggesting that either dfrA is not required for development or dfrA expression is independent of relA expression.
The second approach was to demonstrate that the M. xanthus relA gene was sufficient to rescue the developmental defect of strain MS10 when expressed in trans. For this purpose, we introduced a second copy of the relA gene into strains MS10 and DK101 and designated these strains MS12 and MS13, respectively. Plasmid pMS321 contains the intact M. xanthus relA gene and the first 130 bases of the dfrA gene cloned into the tetracycline resistance Mx8att vector, pSWU29. When assayed for development, strain MS13 regained the ability to form fruiting bodies and could sporulate to 80% of wild-type levels, demonstrating that the defect of MS10 is caused by the disruption of relA and not caused by polar effects on dfrA.
The M. xanthus relA gene rescues the DK527 mutant
Previously, we demonstrated that the E. coli relA gene could rescue the developmental defect of DK527. To determine if the developmental block of strain DK527 could be rescued by the M. xanthus relA gene, two plasmids were constructed containing the M. xanthus relA gene. Plasmid pMS325 also contains Mx8attP, allowing us to test complementation when the M. xanthus relA+ gene is introduced at the Mx8attB site. Plasmid pMS302 lacks an Mx8attP site, but can integrate by homologous recombination in the relA gene. This would result in a tandem duplication of the relA region. When these plasmids were introduced into DK527, both plasmid-carrying strains were found to regain the ability to produce fruiting bodies (the pMS325-carrying strain is shown in Fig. 9) and sporulate based on direct spore count (data not shown) and viable spore assay (Table 2). Control plasmids lacking the M. xanthus relA gene were unable to rescue the developmental defect of DK527. These data strongly suggest that DK527 has a mutant allele of relA.
Table 2.
Sporulation in DK527 rescue strains
| Percent DK101 viable spores (± s.d.)
|
|||
|---|---|---|---|
| Strain
|
72 hr (3 days)
|
120 hr (5 days)
|
168 hr (7 days)
|
| DK101 | 100 (±14.0) | 100 (±2.7) | 100 (±1.7) |
| (wild type) | |||
| DK527 | 0.0007 (±0.0002) | 0.0002 (±0.00009) | 0.0002 (±0.0001) |
| (relA527 mutant) | |||
| MS16 | 84.8 (±10.4) | 71.9 (±5.6) | 78.6 (±2.5) |
| (DK101 pMS325) | |||
| MS17 | 109.4 (±15.3) | 107.1 (±5.7) | 102.1 (±7.0) |
| (DK527 pMS325) | |||
(p)ppGpp production is required for A-factor production
One of the earliest responses to starvation is the production of an extracellular population density signal, Myxococcus A-factor (Kuspa et al. 1986). Because the DK527 mutant fails to express the A-factor-dependent fusions Ω4521 or Ω4400 (Fig. 3), we suspected that DK527 might be deficient in A-factor production. To examine the relationship between (p)ppGpp accumulation and A-factor production, we measured the amount of A-factor produced by DK527 and the relA insertion mutant. Two methods were used to collect A-factor: extracellular complementation (an in situ method in which whole cells are the A-factor producers), and collection of conditioned medium that serves as the source of A-factor.
The bioassay for A-factor employs an A-signaling defective tester strain (DK4324, asgB Ω4521, Kuspa et al. 1986). Six different A-factor donor strains—DK101 (wild-type, positive control), DK527, DK10528 (E. coli relA+), DK10529, MS10 (the relA insertion null), and asgB mutant DK480 (negative control) were each mixed with DK4324. In accordance with earlier experiments (Kuspa and Kaiser 1989; Kuspa et al. 1986), DK101 (asgB+) was able to rescue the A-signaling defect of strain DK4324, whereas the asgB480 (A-factor defective) mutant strain could not (Table 3). Strains DK527 and MS10 had significantly less ability to rescue the defect of the tester strain (Table 3), estimated as 16% and 7% of wild-type A-factor activity, respectively. The E. coli relA+ plasmid restored to DK527 the ability to produce A-factor to 78% of the wild-type level. DK10529 produced the same level of A-factor as DK10528 (within experimental error).
Table 3.
A-factor production
| Donor strain
|
In situ productiona
|
Production in suspensiona
|
|||
|---|---|---|---|---|---|
| U/ml
|
%W.T.
|
timeb
|
U/ml
|
%W.T.c
|
|
| DK101 | 480 ± 86 | 100 | 3 (hr) | 36 ± 3.8 | 100 |
| (wild type) | 6 (hr) | 29 ± 3.2 | 100 | ||
| 9 (hr) | 22 ± 3.1 | 100 | |||
| DK480 | 28 ± 12 | 6 | 3 (hr) | 0.8 ± 0.3 | 2 |
| (DK101 asgB) | 6 (hr) | 1.1 ± 0.4 | 4 | ||
| 9 (hr) | 1.2 ± 0.4 | 5 | |||
| DK527 | 76 ± 22 | 16 | 3 (hr) | 2.2 ± 0.8 | 8 |
| (DK101relA527) | 6 (hr) | 2.3 ± 0.9 | 7 | ||
| 9 (hr) | 2.2 ± 0.8 | 10 | |||
| MS10 | 35 ± 08 | 7 | 3 (hr) | 1.1 ± 0.2 | 3 |
| (DK101relA∷pMS300) | 6 (hr) | 0.9 ± 0.4 | 3 | ||
| 9 (hr) | 0.8 ± 0.4 | 4 | |||
| DK10529d | 425 ± 73 | 88 | 3 (hr) | 39 ± 4.8 | 108 |
| (DK101 pMS132) | 6 (hr) | 32 ± 4.2 | 110 | ||
| 9 (hr) | 21 ± 3.1 | 95 | |||
| DK10528d | 375 ± 76 | 78 | 3 (hr) | 30 ± 4.6 | 83 |
| (DK527 pMS132) | 6 (hr) | 26 ± 4.8 | 89 | ||
| 9 (hr) | 23 ± 4.8 | 104 | |||
Values are shown as units per milliliter of cells (or per milliliter of supernatant recovered from shaken suspensions) at a cell density of 1000 Klett units, prepared as described in Materials and Methods. The values given are an average of three independent experiments.
Time values indicate that the duration cells were cultured in MC7 buffer prior to bioassay of A-factor.
%W.T. activity was calculated by comparing the A-factor activity found in DK480 and DK527 to the amount found in the wild-type strain. Wild-type activity was considered 100% for each time point.
Because levels of (p)ppGpp adequate for fruiting body development are produced by these strains, experiments were performed in the dark, under conditions that lead to fruiting body formation for these strains.
A-factor production was also tested by bioassay of the conditioned medium: MC7 suspension buffer after cells had been agitated and starved therein. MC7 buffer from each of the strains described above—DK101, DK527, DK10528, MS10, and DK480—was assayed for A-factor activity at 3, 6, and 9 hr postinitiation (Table 3). Consistent with the in situ assays, DK101 produced and released A-factor, whereas DK480, DK527, and MS10 produced only 2%, 8%, and 3% of wild-type A-factor activity after 3 hr. Not only were DK527 and MS10 deficient for A-factor production at 3 hr, normally the time of peak level of A-factor production (Kuspa et al. 1992), but there was no evidence for delayed production of A-factor at 6 or 9 hr postinitiation (Table 3). Finally, the DK10528 strain recovered the ability to produce A-factor and to release it into the medium. The data of Table 3 shows that DK527 is defective in the production of A-factor and that the defect can be rescued by expression of the E. coli relA+ gene in DK527.
Discussion
When faced with starvation, M. xanthus has a choice between two responses to that stress: It can either slow its rate of growth to one that can be sustained at the nutrient level of its surroundings, or it can initiate fruiting body development. What role might (p)ppGpp have in that decision? As shown previously, expression of the E. coli relA+ gene in M. xanthus leads to accumulation of (p)ppGpp and activation of early developmental gene expression, even in the presence of a nutrient level sufficient to support growth (Singer and Kaiser 1995). Thus, accumulation of (p)ppGpp appears to be sufficient to initiate the developmental process and represents nutrient limitation.
Previously, we showed that ectopic production of (p)ppGpp in M. xanthus is sufficient to activate the A-factor-dependent Ω4521 fusion in the presence of nutrients (Singer and Kaiser 1995). However, (p)ppGpp cannot activate the asgB480 mutant. Thus both (p)ppGpp and asgB+ are required for activation of Ω4521. Because neither DK527 nor the relA insertion strain is able to produce (p)ppGpp, (p)ppGpp may work through asgB (and/or asgA and asgC) to initiate A-factor production. Were there a second or alternative starvation sensor as well as (p)ppGpp, then this hypothetical sensor might allow starvation-induced A-factor production even in the absence of (p)ppGpp accumulation. However, little A-factor was produced by either strain unable to produce (p)ppGpp, whereas restoration of (p)ppGpp production by the E. coli relA+ largely restored A-factor production. These data are consistent with a model in which individual cells monitor their nutritional status by means of changes in the intracellular concentration of (p)ppGpp, such that accumulation of this nucleotide initiates the developmental program. One output of the early parts of the program would be the activation of A-factor production through the asgA, asgB, and asgC gene products.
Consistent with the suggestion that (p)ppGpp works through asg gene products is the recent work of Plamann and colleagues, showing that asgC is the M. xanthus homolog of rpoD, the E. coli gene that encodes the σ70 subunit of its RNA polymerase (Davis et al. 1995). The asgC767 mutation, which almost completely eliminates A-factor production (Kuspa and Kaiser 1989), results from a glutamic acid to lysine change at amino acid position 598 in the M. xanthus σ70 homolog (Davis et al. 1995). This homolog is called SigA in M. xanthus (Inouye 1990). Glutamate 598 is adjacent to an equivalent amino acid position in the E. coli σ70 protein, at which suppressors of a ΔrelA ΔspoT double-null mutation arise (Hernandez and Cashel 1995). Experiments with E. coli mutants carrying either the rpoD(P504L) allele or the rpoD(S506) mutation suggest that region 3 of the E. coli σ70 subunit may be directly involved in (p)ppGpp-mediated gene regulation (Hernandez and Cashel 1995).
Experiments reported here combined with those of Singer and Kaiser (1995) imply that the accumulation of (p)ppGpp in response to nutrient deprivation is both necessary and sufficient to activate the developmental program of M. xanthus. Moreover, intracellular production of (p)ppGpp precedes, and is necessary for, extracellular production of A-factor. The next challenge is to understand how (p)ppGpp and A-factor jointly regulate Ω4521 and other genes that are both starvation and A-factor dependent, and how the critical choice between slow growth and initiation of fruiting body development is made.
Materials and methods
Bacterial strains, phage, and plasmids
All strains and plasmids used are listed in Table 1. The reference strain used for these studies was DK101 (Manoil and Kaiser 1980a), a developmentally competent M. xanthus strain that contains an sglA1 allele, allowing cells to grow well dispersed in liquid culture (Hodgkin and Kaiser 1979). For simplicity, DK101 has been referred to as wild-type throughout the paper. The isolation of DK527 from DK101 has been described (Manoil and Kaiser 1980a). The transducing mxyophages Mx4ts18ts27hrm (Campos et al. 1978) and Mx8clp2 (Martin et al. 1978) have been described, as well as the E. coli strain MG1655 and the isogenic ΔrelA+ strain (Singer et al. 1991). The Tn5lac transcriptional fusions Ω4408, Ω4455, Ω4400, and Ω4521 have also been described previously (Kroos et al. 1986). DK101 and DK527 derivatives harboring each fusion were constructed by Mx8- or Mx4-mediated transduction. The presence of a single Tn5 element in the M. xanthus chromosome was confirmed by Southern blotting.
Plasmid construction
The 4.8-kb PstI fragment encoding the M. xanthus relA was cloned from cosmid G4 into the PstI sites of pBGS18, giving rise to pMS302, and into the Mx8attP containing vector pPLH343, giving rise to pMS350. Subclones containing small fragments of pMS302 were cloned into either pBGS18 or pBluescriptSK (Stratagene, La Jolla, CA) by use of standard cloning protocols (Sambrook et al. 1989) and are listed in Table 1.
Plasmid pMS321 was constructed by digesting pMS302 with NcoI and blunting the end with T4 DNA polymerase, then digesting with BamHI. This fragment was then cloned into the BamHI–SmaI sites of pSWU29, a plasmid that carries the Mx8attP region and tetracycline resistance. This plasmid removes the downstream dfrA ORF from the original insert.
Plasmid pMS325 was constructed by digesting pMS321 with HindIII and blunting the end with DNA polymerase Klenow fragment, then by cutting with XhoI. This procedure removed all M. xanthus sequences upstream to the internal XhoI site of relA. Next, a 480-bp fragment containing 5′ sequences from the upstream EagI site (blunted by DNA polymerase Klenow fragment) to the internal XhoI site was inserted. The final construct, which removes upstream and downstream ORFs, was verified by sequencing with an internal primer.
Plasmid pMS381, containing the M. xanthus relA gene under the control of the E. coli tac Promoter (Ptac), was constructed in a two-step procedure. First, a 477-bp PCR fragment was isolated with primers RELA2nd06 (5′-AGTTCTCCGCCTGCTTCTCC-3′) and RelABam-1 (5′-TAGGATCCGGGTCAACGAAAGCGAACGCA-3′), placing a BamHI restriction site 20 bp from the GTG translational start. This fragment contains the 5′ end of the relA gene without its promoter. The resulting 477-bp PCR product was then digested with XhoI and BamHI, giving rise to a 267-bp fragment encoding the 5′ end of the relA gene. The 267-bp XhoI–BamHI fragment was then cloned into pMS302 and designated pMS380. The construct was verified by sequencing with an internal primer. To construct pMS381, the 3.3-kb BamHI–HindIII fragment from pMS380 was cloned into the BamHI–HindIII site of pTTQ18 (Amersham, Arlington Heights, IL), placing M. xanthus relA under the control of Ptac. The junction fragment was confirmed by DNA sequencing.
Growth and development
M. xanthus cells were grown in either CTT liquid [1% casitone (Difco Laboratories), 10 mm (Tris-H)ydrochloride (pH 7.6), 1 mm KHPO4 (pH 7.8), 8 mm MgSO4], M1 liquid (Zusman et al. 1971), or A1 liquid (Bretscher and Kaiser 1978) at 33°C with vigorous agitation. For solid support, agar to a final concentration of 1.5% (wt/vol) (CTT and M1), or agarose to a final concentration of 0.8% (wt/vol) (A1) was added as described (Bretscher and Kaiser 1978). Where indicated, medium was supplemented with oxytetracycline (12 μg/ml) or kanamycin (40 μg/ml). Cell growth in liquid cultures was monitored with a Klett–Summerson photoelectric colorimeter (model 800-3) equipped with a red filter; 100 Klett units corresponds to 5 × 108 cells/ml. E. coli cells were grown in M9 complete medium (Singer et al. 1991). For RNA synthesis experiments with serine hydroxamate and for labeling with [3H]uridine, E. coli cells were grown in M9 complete medium lacking serine and uracil (Singer et al. 1991).
Development was initiated in M. xanthus by starvation as described previously (Kroos et al. 1986). Developmental β-galactosidase assays were performed as described (Kroos et al. 1986). The time of initial β-galactosidase expression (expression time) was determined by extrapolating the curve of increasing enzyme activity to its intercept with a line from the t = 0 value prolonged parallel to the time axis (Kroos et al. 1986). Fruiting body development was monitored visually with a dissecting microscope (Wild-Heerbrug, Switzerland). Two methods were used to examine sporulation efficiency. First, spore viability assays were performed as described previously (Thony-Meyer and Kaiser 1993). Second, direct spore counts were performed with a Petroff–Hausser counting chamber and phase contrast microscopy.
Light induction of E. coli RelA in M. xanthus
M. xanthus strains were grown in liquid or on solid support medium and irradiated as described previously (Singer and Kaiser 1995).
PCR analysis
Two synthetic primers, R972 [5′-AAAAAGAATTCAACGG(C/G) TACCAG(T/A)(C/G)(C/G)ATCCACAC-3′] and X1122 [5′AAAAAATCTAGA(C/G) CCCTT(C/G)GG(C/G)GTGAA(C/G)ACGTA-3′], were used to amplify an internal segment of the M. xanthus relA homolog from strain DK101 by the polymerase chain reaction. Reaction mixtures contained 10 mm Tris-HCl (pH 8.5), 50 mm KCl, 2 mm MgCl2, 200 μm of each of the four dNTPs, 2.5 units of Taq polymerase, 40 pmole of each primer, and 50 ng of chromosomal DNA in a final reaction volume of 100 μl. Samples were denatured at 95°C for 2 min and subjected to 30 cycles of denaturation (95°C for 1 min), annealing (57°C for 30 sec), and extension (72°C for 1 min) with a final extension incubation for 10 min, at 72°C. PCR products were then analyzed by agarose gel electrophoresis (1.5% wt/vol). The resulting 311-bp fragment was isolated by use of the QIAEX II gel extraction kit (Qiagen Inc., Chatsworth, CA) and cloned into the EcoRI and BamHI sites of pBGS18.
Colony hybridization and radioactive probe protocols
Nitrocellulose filters containing an ordered array of DNA from an M. xanthus cosmid library, were provided by R. Gill. Filters were probed by use of the 311-bp PCR-generated DNA fragment labeled with 32P. Radioactive probes were made by the random primer method (Sambrook et al. 1989) and [α-32P]GTP (NEN) at 3000 Ci/mmole, 10 mCi/ml.
Total RNA synthesis
RNA synthesis rates in vivo were measured as described previously (Singer et al. 1991) with the following modifications. Cells were grown in either M9 glucose (E. coli) or M1 (M. xanthus) medium to a density of 5 × 108 cells/ml, then serine hydroxamate (final concentration 2.5 mg/ml) was added to inhibit the charging of seryl-tRNA. Portions of 100 μl were removed for analysis: two portions prior to addition of serine hydroxamate (t=0), followed by experimental portions (performed in duplicate) at the time intervals indicated in Figure 1. All samples were exposed for 2 min to 10 μCi/ml [5,6-3H]uridine (Amersham TRK. 410; 40 Ci/mmole) plus 2 μg/ml uridine. Incorporation was stopped by addition of 2 ml of ice-cold 5% trichloroacetic acid (TCA) for 30 min. Samples were washed three times with 5 ml of ice-cold 2% TCA, rinsed with 70% ethanol, air-dried, and the 3H-incorporated into TCA insoluble material was determined in a Beckman scintillation counter (model LS 1801). Incorporation of radioactivity was linear for up to 30 min under these conditions.
Measurement of guanosine nucleotides
Guanosine nucleotides were isolated and measured as described previously (Manoil and Kaiser 1980a; Singer and Kaiser 1995).
Blotting and physical mapping
Southern blotting was performed as described previously (Sambrook et al. 1989). Physical mapping was performed by the Bio-Rad CHEF Mapping DRII system with a 1% agarose gel in 0.5% TBE buffer at 10°C for 15:10 hr at 6V/cm (200 V) with switch times of 70 and 120 sec. M. xanthus DNA was prepared and digested in agarose plugs as described previously (Chen et al. 1990, 1991). CHEF gels were then used for Southern blot analysis.
Gene locations were further localized on the M. xanthus physical map by performing Southern blot analysis by use of an ordered set of M. xanthus YAC clones (Kuspa et al. 1989; Chen et al. 1990, 1991). Whole yeast DNA (containing the YAC DNA) was purified from strains carrying M. xanthus YAC clones (Coulson et al. 1988). Samples were denatured with 0.4 m NaOH at 65°C for 15 min, and 25 μg of DNA was transferred to a Nytran filter (Amersham, Arlington Heights, IL) by use of a slot blot vacuum apparatus (Bio-Rad, Hercules, CA). The DNA was then cross-linked to the membrane with a UV transilluminator and subjected to Southern hybridization procedures as described above.
Western blots
Western blots were performed as described previously (Straus et al. 1987; Singer and Kaiser 1995) Anti-E. coli RelA antibody was provided by G. Glaser (Hadassa Medical School, Jerusalem, Israel).
Sequence analysis
DNA sequence analysis was performed by the Division of Biological Studies Automated DNA Sequencing Facility, at the University of California, Davis, with the ABI Prism Dye Terminator Cycle Sequencing Ready Reaction Kit using AmpliTaq DNA polymerase. Reactions were run on a 4.25% acryl/bisacrylamide gel with an ABI Prism 377 DNA Sequencer. Sequence data was analyzed by use of the ABI Prism Sequencing 2.1.1 software (Perkin Elmer, Foster City, CA) and contigs assembled with Sequencher v3.0 (GeneCodes Corporation, Madison, WI).
Primer extension
Vegetative RNA was isolated from DK101 cells as described previously (Kaplan et al. 1991). Primer extensions were performed as per Mirel and Chamberlin (1989) with the following modifications: 25 μg of vegetative RNA or tRNA was used in each reaction, by use of the synthetic oligonucleotides RelA2nd05 (5′-CCTTCTTGATGATGTCCAG-3′) and RelA2nd06 (5′-AGTTCTCCGCCTGCTTCTCC-3′). Samples were run on a 5% Long Ranger (J.T. Baker, Phillipsburg, NJ) urea acrylamide gel alongside DNA sequence reactions. Sequence reactions were performed with the fmol DNA cycle sequencing kit (Promega, Madison, WI), as described by the manufacturer. Primers used for primer extension were simultaneously used for fmol cycle sequencing reactions.
A-factor assays
A-factor production in situ was assayed as described previously (Kuspa et al. 1986). A-factor released into the extracellular medium by the donor cells was assayed as described (Kuspa and Kaiser 1989).
Acknowledgments
We thank A. Garza for his generous assistance and E. Wickstrom for helpful discussions of this work, S. Hoover and D. Lavell in the University of California at Davis sequencing facilities for technical support, and R. Gill for supplying his cosmid library. In addition, we thank G. Holland, M. Igo, C. Price, and L. Plamann for critically reading this manuscript. This work was supported in part by National Institutes of Health grants GM54592 to M.S. and GM23441 to D.K.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact.
Footnotes
E-MAIL mhsinger@ucdavis.edu; FAX (530) 752-9014.
References
- Borek E, Rockenbach J, Ryan A. Studies on a mutant of Escherichia coli with unbalanced ribonucleic acid synthesis. J Bacteriol. 1956;71:318–323. doi: 10.1128/jb.71.3.318-323.1956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bretscher A, Kaiser D. Nutrition of Myxococcus xanthus, a fruiting myxobacterium. J Bacteriol. 1978;133:763–768. doi: 10.1128/jb.133.2.763-768.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campos J, Geisselsoder J, Zusman D. Isolation of bacteriophage MX4, a generalized transducing phage for Myxococcus xanthus. J Mol Biol. 1978;119:167–178. doi: 10.1016/0022-2836(78)90431-x. [DOI] [PubMed] [Google Scholar]
- Cashel M. The control of ribonucleic acid synthesis in Escherichia coli. IV. Relevance of unusual phosphorylated compounds from amino acid starved stringent strains. J Biol Chem. 1969;244:3133–3141. [PubMed] [Google Scholar]
- Cashel M, Gallant J. Two compounds implicated in the function of the RC gene of Escherichia coli. Nature (London) 1969;221:838–841. doi: 10.1038/221838a0. [DOI] [PubMed] [Google Scholar]
- Cashel M, Gentry D, Hernandez J, Vinella D. The stringent response. In: Neidhart FC, editor. Escherichia coli and Salmonella: Cellular and molecular biology. 2nd ed. Vol. 1. Washington DC.: ASM Press; 1996. pp. 1458–1496. [Google Scholar]
- Ceciliani F, Faotto L, Negri A, Colombo I, Berra B, Bartorelli A, Ronchi S. The primary structure of UK114 tumor antigen. FEBS Lett. 1996;393:147–150. doi: 10.1016/0014-5793(96)00850-2. [DOI] [PubMed] [Google Scholar]
- Chakraburtty R, Bibb M. The ppGpp synthetase gene (relA) of Streptomyces coelicolor A3(2) plays a conditional role in antibiotic production and morphological differentiation. J Bacteriol. 1997;179:5854–5861. doi: 10.1128/jb.179.18.5854-5861.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakraburtty R, White J, Takano E, Bibb M. Cloning, characterization and disruption of a (p)ppGpp synthetase gene (relA) of Streptomyces coelicolor A3(2) Mol Microbiol. 1996;19:357–368. doi: 10.1046/j.1365-2958.1996.390919.x. [DOI] [PubMed] [Google Scholar]
- Chen H, Keseler I, Shimkets L. Genome size of Myxococcus xanthus determined by pulsed-field gel electrophoresis. J Bacteriol. 1990;172:4206–4213. doi: 10.1128/jb.172.8.4206-4213.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen H, Kuspa A, Keseler I, Shimkets L. Physical map of the Myxococcus xanthus chromosome. J Bacteriol. 1991;173:2109–2115. doi: 10.1128/jb.173.6.2109-2115.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coulson A, Waterston R, Kiff J, Aulston J, Kohara Y. Genome linking with yeast artificial chromosomes. Nature. 1988;335:184–186. doi: 10.1038/335184a0. [DOI] [PubMed] [Google Scholar]
- Davis J, Mayor K, Plamann L. A missense mutation in rpoD results in an A-signaling defect in Myxococcus xanthus. Mol Microbiol. 1995;18:943–952. doi: 10.1111/j.1365-2958.1995.18050943.x. [DOI] [PubMed] [Google Scholar]
- Dworkin M. Nutritional requirements for vegetative growth of Myxococcus xanthus. J Bacteriol. 1962;84:250. doi: 10.1128/jb.84.2.250-257.1962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiil NP, Friesen JD. Isolation of relaxed mutants of Escherichia coli. J Bacteriol. 1968;95:769–786. doi: 10.1128/jb.95.2.729-731.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernandez J, Cashel M. Changes in conserved region 3 of Escherichia coli σ70 mediate ppGpp-dependent function in vivo. J Mol Biol. 1995;252:536–549. doi: 10.1006/jmbi.1995.0518. [DOI] [PubMed] [Google Scholar]
- Hodgkin J, Kaiser D. Genetics of gliding motility in M. xanthus (Myxobacterales): Two gene systems control movement. Mol & Gen Genet. 1979;171:177–191. [Google Scholar]
- Inouye S. Cloning and DNA sequence of the gene coding for the major sigma factor from Myxococcus xanthus. J Bacteriol. 1990;172:80–85. doi: 10.1128/jb.172.1.80-85.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan H, Kuspa A, Kaiser D. Suppressors that permit A-signal independent developmental gene expression in Myxococcus xanthus. J Bacteriol. 1991;173:1460–1470. doi: 10.1128/jb.173.4.1460-1470.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimsey H, Kaiser D. Targeted disruption of the Myxococcus xanthus orotidine-5′-monophosphate decarboxylase gene: Effects on growth and fruiting body development. J Bacteriol. 1991;173:6790–6797. doi: 10.1128/jb.173.21.6790-6797.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroos L, Kuspa A, Kaiser D. A global analysis of developmentally regulated genes in Myxococcus xanthus. Dev Biol. 1986;117:252–266. doi: 10.1016/0012-1606(86)90368-4. [DOI] [PubMed] [Google Scholar]
- Kuspa A, Kaiser D. Genes required for developmental signaling in Myxococcus xanthus: Three asg loci. J Bacteriol. 1989;171:2762–2772. doi: 10.1128/jb.171.5.2762-2772.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuspa A, Kroos L, Kaiser D. Intercellular signaling is required for developmental gene expression in Myxococcus xanthus. Dev Biol. 1986;117:267–276. doi: 10.1016/0012-1606(86)90369-6. [DOI] [PubMed] [Google Scholar]
- Kuspa A, Vollrath D, Cheng Y, Kaiser D. Physical mapping of the Myxococcus xanthus genome by random cloning in yeast artificial chromosomes. Proc Natl Acad Sci. 1989;86:8917–8921. doi: 10.1073/pnas.86.22.8917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuspa A, Plamann L, Kaiser D. Identification of heat-stable A-factor from Myxococcus xanthus. J Bacteriol. 1992;174:3319–3326. doi: 10.1128/jb.174.10.3319-3326.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li S, Lee B, Shimkets L. csgA expression entrains Myxococcus xanthus development. Genes & Dev. 1992;6:401–410. doi: 10.1101/gad.6.3.401. [DOI] [PubMed] [Google Scholar]
- Manoil C. “Initiation of the development of Myxococcus xanthus.” Ph.D. thesis. Stanford, CA: Stanford University; 1982. [Google Scholar]
- Manoil C, Kaiser D. Accumulation of guanosine tetraphosphate and guanosine pentaphosphate in Myxococcus xanthus during starvation and myxospore formation. J Bacteriol. 1980a;141:297–304. doi: 10.1128/jb.141.1.297-304.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ————— Guanosine pentaphosphate and guanosine tetraphosphate accumulation and induction of Myxococcus xanthus fruiting body development. J Bacteriol. 1980b;141:1062–1065. doi: 10.1128/jb.141.1.305-315.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin S, Sodergren E, Masuda T, Kaiser D. Systematic isolation of transducing phages for Myxococcus xanthus. Virology. 1978;88:44–53. doi: 10.1016/0042-6822(78)90108-3. [DOI] [PubMed] [Google Scholar]
- Metzger S, Dror IB, Aizenman E, Schreiber G, Toone M, Friesen JD, Cashel M, Glaser G. The nucleotide sequence and characterization of the relA gene of Escherichia coli. J Biol Chem. 1988;263:15699–15704. [PubMed] [Google Scholar]
- Mirel D, Chamberlin M. The Bacillus subtilis fagellin gene (hag) is transcribed by the σ28 form of RNA polymerase. J Bacteriol. 1989;171:3095–3101. doi: 10.1128/jb.171.6.3095-3101.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oka T, Tsuji H, Noda C, Sakai K, Hong Y, Suzuki I, Muñoz S, Natori Y. Isolation and characterization of a novel perchloric acid-soluble protein inhibiting cell-free protein synthesis. J Biol Chem. 1995;270:30060–30067. doi: 10.1074/jbc.270.50.30060. [DOI] [PubMed] [Google Scholar]
- Orndorff P, Stellwag E, Starich T, Dworkin M, Zissler J. Genetic and physical characterization of lysogeny by bacteriophage Mx8 in Myxococcus xanthus. J Bacteriol. 1983;154:772–779. doi: 10.1128/jb.154.2.772-779.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Fritsch E, Maniatis T. Molecular cloning: A laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Sarubbi E, Rudd KE, Xiao H, Ikehara K, Kalman M, Cashel M. Characterization of the spoT gene of Escherichia coli. J Biol Chem. 1989;264:15074–15082. [PubMed] [Google Scholar]
- Schmiedeknecht G, Kerkhoff C, Orsó E, Stöhr J, Aslandis C, Nagy G, Kneuchel R, Schmitz G. Isolation and characterization of a 14.5-kDa trichloroacetic-acid-soluble translational inhibitor protein from human monocytes that is upregulated upon cellular differentiation. Eur J Biochem. 1996;242:339–351. doi: 10.1111/j.1432-1033.1996.0339r.x. [DOI] [PubMed] [Google Scholar]
- Schreiber G, Metzger S, Aizenman E, Roza S, Cashel M, Glaser G. Overexpression of the relA gene in Escherichia coli. J Biol Chem. 1991;266:3760–3767. [PubMed] [Google Scholar]
- Shimkets L. Nutrition, metabolism and the initiation of development. In: Rosenberg E, editor. Myxobacteria, development and cell interactions. New York: Springer-Verlag; 1984. pp. 92–108. [Google Scholar]
- ————— . The myxobacterial genome. In: Dworkin M, Kaiser D, editors. Myxobacteria II, Washington, DC.: American Society of Microbiology; 1993. [Google Scholar]
- Singer M, Kaiser D. Ectopic production of guanosine penta- and tetraphosphate can initiate early developmental gene expression in Myxococcus xanthus. Genes & Dev. 1995;9:1633–1644. doi: 10.1101/gad.9.13.1633. [DOI] [PubMed] [Google Scholar]
- Singer M, Walter W, Cali B, Liebke H, Gourse R, Gross C. Physiological effects of the fructose-1,6-diphosphate aldolase ts8 mutation on stable RNA synthesis in Escherichia coli. J Bacteriol. 1991;173:6249–6257. doi: 10.1128/jb.173.19.6249-6257.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spratt B, Hedge P, Heesen S, Edelman A, Broome-Smith J. Kanomycin-resistant vectors that are analogues of plasmids pUC8, pUC9, pEMBL8, and pEMBL9. Gene. 1986;41:337–342. doi: 10.1016/0378-1119(86)90117-4. [DOI] [PubMed] [Google Scholar]
- Stent G, Brenner S. A genetic locus for the regulation of ribonucleic acid synthesis. Proc Natl Acad Sci. 1961;47:2005–2014. doi: 10.1073/pnas.47.12.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straus D, Walter W, Gross C. The heat shock response of E. coli is regulated by changes in the concentration of σ32. Nature. 1987;329:348–351. doi: 10.1038/329348a0. [DOI] [PubMed] [Google Scholar]
- Thony-Meyer L, Kaiser D. devRS, an autoregulated and essential genetic locus for fruiting body development in Myxococcus xanthus. J Bacteriol. 1993;175:7450–7462. doi: 10.1128/jb.175.22.7450-7462.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tosa T, Pizer L. Biochemical bases for the antimetabolite action of L-serine hydroxamate. J Bacteriol. 1971;106:972–982. doi: 10.1128/jb.106.3.972-982.1971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watson B, Dworkin M. Comparative intermediary metabolism of vegetative cells and microcysts of Myxococcus xanthus.. J Bacteriol. 1968;96:1456–1473. doi: 10.1128/jb.96.5.1465-1473.1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wireman J, Dworkin M. Developmentally induced autolysis during fruiting body formation by Myxococcus xanthus. J Bacteriol. 1977;129:796–802. doi: 10.1128/jb.129.2.798-802.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao H, Kalman M, Ikehara K, Aemel S, Glaser G, Cashel M. Residual guanosine 3′5′-bispyrophosphate synthetic activity of relA null mutants can be eliminated by spoT null mutations. J Biol Chem. 1991;266:5980–5990. [PubMed] [Google Scholar]
- Zusman D, Rosenberg E. Division cycle of Myxococcus xanthus II. Kinetics of cell growth and protein synthesis. J Bacteriol. 1971;105:811–819. doi: 10.1128/jb.105.3.811-819.1971. [DOI] [PMC free article] [PubMed] [Google Scholar]