

Abstract
Abstract
Theories concerning the role of the climbing fibre system in motor learning, as opposed to those addressing the olivocerebellar system in the organization of motor timing, are briefly contrasted. The electrophysiological basis for the motor timing hypothesis in relation to the olivocerebellar system is treated in detail.
Rodolfo R. Llinás is currently the Thomas and Suzanne Murphy Professor of Neuroscience and Chairman of the department of Physiology and Neuroscience at the NYU School of Medicine. Interested in electrophysiology he has contributed studies in squid giant synapse, the cerebellum and inferior olive, the cerebral and entorhinal cortex, the vestibular system and spinal cord. He has also studied human brain function using magnetoencephalography (MEG) on the basis of which he introduced the concept of thalamocortical dysrhythmia.
 |
Introduction
Over the last 40 years, following the discovery of climbing fibre activation of Purkinje cells (Eccles et al. 1964, 1966a), many questions have been posed regarding the functional significance of this extraordinarily powerful synaptic input. Two hypotheses were proposed in the next few years following their discovery. The simpler was that the climbing fibre's role had to do with rapid (phasic) motor timing (see Llinas, 1974) while the second posited that climbing fibre function had to do with motor learning (Marr, 1969; Ito & Kano, 1982).
The common theme of both had to do with the bizarre distribution of the synaptic input over the dendrite of the Purkinje cell. Indeed it was pointed out that a synaptic distribution that simultaneously activated most of the basic dendritic tree would result in a negation of dendritic integration (Llinas, 1974) as the dendrite membrane potential becomes close to isopotential during this activation. Simply stated then, one idea was that ‘climbing fibre input has to be strong to override whatever the cell is doing and generate a well timed activation resulting in a well timed inhibition of the cerebellar nuclear neurons (Llinas, 1974).’ Indeed, the statement implied that the Purkinje cell, a glorified inhibitory interneuron, serves as a feed-forward inhibitor of the olivocerebellar input to the cerebellar nuclei.
The learning hypothesis proposed that the climbing fibre had no real role in the olivocerebellar circuit, as the Purkinje cells being over-depolarized probably would not fire due to sodium conductance inactivation. This being the case, the climbing fibre input should have another function. And the proposed function, on this assumption, was that the climbing fibre system had evolved to change the gain of the parallel fibre–Purkinje cell synapse and so serve as ‘instructors’ that would modify parallel synaptic strength (Ito, 2001).
Initially it was suggested that long-term potentiation (Marr, 1969) would be the mechanism for such plasticity, but experimental studies found that upon artificially pairing climbing fibre activation with parallel fibre activation by electrical stimulation, a reduction, rather than a potentiation, and actual long-term depression (LTD) of the parallel fibre amplitude was encountered (Ito & Kano, 1982). This led to theoretical papers suggesting that LTD (Albus, 1971) was the mechanism for cerebellar learning.
At this time most experiments were actually implemented in vitro and only a few valiant experiments were implemented in vivo, with great technical skill (Jörntell & Ekerot, 2002). The problem with such experiments was that LTD could only be evoked in the living preparation following local inhibitory blockers (Jörntell & Ekerot, 2002), and so the results had a difficult bias to contend with. By contrast, the timing hypothesis remained in the shadow, as the last decades of the twentieth century were the time of the flourishing of synaptic plasticity while non-learning research was considered to be of diminished interest in neuroscience.
Currently several papers have demonstrated that motor learning proceeds in the absence of LTD. Indeed, the experiments based both in pharmacological (Welsh et al. 2005) and genetic manipulations that impede LTD in Purkinje cells (Schonewille et al. 2011) demonstrate no modification of motor learning under such conditions. This being the case I would like to address the timing hypothesis, as some of us see it these days.
Still fundamental to the initial discussion of timing is role of the inferior olive in the genesis of a timing signal, and equally interesting, why it should be that inferior olive (IO) rhythmicity is so constant in its intrinsic frequency among individuals with so different a motor ‘plant’ as a mouse and a giraffe. Indeed, consider the consequences to our motricity and our ability to cope with our cognitive requirements on a planet with twice the mass (not to consider an order of magnitude larger mass). Indeed just from the speed of fall alone our reflex response speed and our motricity, in general, would become woefully inadequate.
The mass of earth and the speed of motricity
And so why do we humans (as well as other mammals) implement a 10-cycle rhythm to pace motricity? Indeed, if we look at the speed with which movements are implemented they most likely fall within the approximately 10 Hz generated by the inferior olivary–cerebellar nuclei circuit and its control via the olivo–Purkinje–cerebellar nuclei loop. The arguments fall on the tetanic fusion frequency of striate muscle (Cavagna et al. 1976), resonant properties of limbs (Vallbo & Wessberg, 1993), optimization of energy consumption (Biewener, 2006) and/or the computational advantages of discontinuous motricity (Welsh & Llinas, 1997). Perhaps, by considering mostly the optimization of ‘body plant’ dynamics we disregard the most likely variable forging the limits of motion speed, the mass of earth. Indeed, considering that earth's mass (and thus its gravity) has been quite constant since the evolution of multicellular life forms (in the Cambrian) the speed of falling objects within the range of usual heights (tall trees) is ancient. It is, thus, not surprising that the speed of motricity is quite similar among most species.
Basically one must move fast enough to avoid being hit by a falling object (be it a brick or a tiger). Roughly, except for power flight in some birds, e.g. the peregrine falcon diving down on its prey 91.7 mph (Clark, 1995), maximum quadruped horizontal displacement velocity, the cheetah at 65 mph (Sharp, 1997), is actually half that of the speed of freefall (120 mph for skydiving in the belly-to-earth position).
Consider, by contrast, the motor problems we would encounter on the surface of a planet having twice, or even higher mass, than earth.
Motor coordination and timing
Concerning motor coordination and timing, several general issues are evident in the electrophysiology of both the olivocerebellar system and the mossy fibre granule cell–Purkinje cell systems.
Concerning the olivocerebellar system, (1) the system generates a timing signal that is inscribed in the intrinsic electrical properties of single neurons, (2) the organization of the nucleus via electrical coupling allows for synchronous multicellular temporal coherence that generates a close to simultaneous neuronal cluster activation, and (3) due to the remarkable property of conduction isochronicity the timing signal does not disperse against distance as it is conducted along the pathways carrying it to the final integration sites at the cerebellar nuclear level.
Concerning the mossy fibre system afferent system, the timing component must of necessity be deeply involved with the mossy fibre–granule cell–Purkinje cell activation system (D'Angelo et al. 2009) in conjunction with the mossy fibre collateral afferents terminating at cerebellar nuclear level (D'Angelo, 2010). Added to this rather larger system is the feedback inhibitory system generated by the inhibitory Golgi cell system (Eccles et al. 1966b). The actual mechanism of how such crucial input must interact in a temporal framework with its olivocerebellar counterpart is at present not clear. Nevertheless, new information concerning this system and its important role in the cerebellar timing function has been treated in detail by D'Angelo et al. (2009). Also of importance here is the fact that while plasticity as it relates to the climbing fibre Purkinje cell activation and the generation of LTD has been demonstrated to be bogus (see above), the possibility remains that plasticity within the mossy fibre afferent system may be an important parameter optimizing temporal coincidence between the two forms of Purkinje cell activation. In particular the possibility that the glomerular synapse, at granule cell level, which brings together mossy fibre input and feedback inhibition onto the granule cell dendrites, may be the site of temporal coincidence modulation must be seriously considered (D'Angelo, 2010). As this aspect of cerebellar timing will be covered in Professor D'Angelo's paper I will limit my remarks to the olivocerebellar system.
The olivocerebellar system
The climbing fibre system, shown morphologically as originating from the inferior olive nucleus (Szentagothai & Rajkovits, 1959), is one of the two major afferent pathways to the cerebellar cortex (Cajal, 1888). Climbing fibres innervate Purkinje cells (PCs) directly in the cerebellar cortex, generating the largest synaptic junction in the vertebrate central nervous system. Indeed, activation of a climbing fibre is followed by an all or none burst of spikes in all of its postsynaptic PCs (Eccles et al. 1966b). On average, each IO neuron generates 10 or so climbing fibres (Armstrong & Schild, 1970) that distribute rostrocaudally over many folia of the cerebellar cortex. In addition to contacting PCs, climbing fibres also produce collateral branches that terminate in all cerebellar nuclei (Palay & Chan-Palay, 1974).
IO neurons fire spontaneously at 1–10 Hz and can exhibit rhythmic oscillatory activity near 10 Hz (Crill, 1970; Llinas & Yarom, 1981). Likewise, the resulting complex spikes in the PCs are also rhythmic and have an average frequency of 10 Hz (Bell & Kawasaki, 1972). The fact that IO neurons are electrically coupled and tend to fire in groups (Llinas, 1974; Sotelo et al. 1974) lends support to the proposal that climbing fibres may perform a timing function in motor coordination (Llinas et al. 1975). Furthermore, simultaneous recordings from multiple PCs have shown that complex spikes occur synchronously within groups of PCs (Bower & Woolston, 1983; Llinas & Sasaki, 1989; Lang et al. 1996; De Zeeuw et al. 1998; Fukuda et al. 2001a). Such synchronous activation of multiple PCs by the IO nucleus suggests that clusters of IO neurons have the ability to fire rhythmically and in unison.
This spatial organization would be the actual product of dynamic plasticity established during development (Nicholson & Freeman, 2003). These synchronous IO oscillations have been proposed to be important in determining the timing and spatial organization of motor sequences (Llinas, 1988; Lampl & Yarom, 1997; Welsh & Llinas, 1997). According to this hypothesis, IO activity functions as a motor timing signal by generating synchronous and rhythmic activation of cerebellar nuclear and descending vestibular nuclear neurons. The timing of the activation of these nuclei is controlled additionally by the synchronized inhibitory PC barrage originated by the climbing fibre activation of PCs. Evidence in support of this motor timing proposal includes the synchronous firing of a population of PCs in awake, anaesthetized animals (Llinas & Sasaki, 1989; Lang et al. 1999; Fukuda et al. 2001b); recordings of rhythmic inhibitory potentials in the deep cerebellar nuclei (Llinas & Muhlethaler, 1988); and a temporal correlation between the firing of the olivocerebellar system and the execution of movements (Welsh et al. 1995).
At a more global level, the IO has been shown to have a deep and significant role in the maintenance of the dynamic properties of the cerebellar control of motricity by regulating the firing properties of PCs. Thus, in the absence of the climbing fibre input PCs fire constantly, indicating a dis-regulation of their intrinsic electrical properties (Strata & Montarolo, 1982).
In summary, the functioning of the olivocerebellar system has been viewed (1) as one that addresses the time coherence of muscle activation that is required to generate the massively parallel event known as motor coordination, and (2) as one that responds to errors in motor execution. These are considered in turn.
Single cell electrophysiology
The ability of the olivocerebellar system to generate synchronous rhythmic activity has been attributed to the intrinsic oscillatory properties of the IO neurons (Llinas & Yarom, 1981a,b; Benardo & Foster, 1986; Bal & McCormick, 1997) and their electrotonic coupling (Llinas, 1974; Sotelo et al. 1974; Llinas & Yarom, 1981b; Lampl & Yarom, 1997; Makarenko & Llinas, 1998; Yarom & Cohen, 2002). In particular, several types of voltage-dependent calcium and potassium conductances, in addition to those involved in action potential generation, enable IO cells to oscillate and fire rhythmically at 1–10 Hz. These conductances include a high-threshold Ca2+ conductance, a low-threshold Ca2+ conductance, a Ca2+-activated K+ conductance, and a hyperpolarization-activated cationic conductance (Llinas & Yarom, 1981a,b, 1986; Bal & McCormick, 1997).
Concerning the electrical coupling, as in other CNS structures (Bennett, 2000), gap junctions constitute the main communication pathway between the IO neurons (Sotelo et al. 1974; De Zeeuw et al. 1996). Such electrotonic coupling has been assumed to play a crucial role in synchronizing IO oscillations and in generating groups of concurrently oscillating neurons (Llinas & Yarom, 1986).
This coupling was also assumed to be controlled by return glomerular inhibition (Llinas, 1974). IO afferents were, in fact, found to modulate the efficiency of electrotonic coupling via inhibition shunting directly at the glomerulus. The pathway turned out to be supported by a large set of cerebellar nuclear GABAergic neurons (Sotelo et al. 1986; de Zeeuw et al. 1989; Fredette & Mugnaini, 1991; De Zeeuw et al. 1996; Medina et al. 2002) that, surprisingly, represent almost 50% of the total neuronal population in such nuclei, giving some measure of the importance of this feedback inhibitory pathway.
Indeed, it was determined experimentally that such input can control the degree and distribution of synchronous oscillatory activity in the IO nucleus (Leznik et al. 2002) and the cerebellar cortex (Lang et al. 1996; Lang, 2001, 2002). Moreover, dynamic groups of IO neurons oscillating in-phase can synchronously activate a population of PCs and thereby control patterns of synchronous activity in the cerebellum during motor coordination (Welsh et al. 1995).
Visualization of IO cluster activity
Although synchronized IO oscillations are a neuronal ensemble event, they have been studied primarily on a single-cell level and no information has been available about their spatial profiles. Thus, an attempt was made to address this issue by utilizing voltage-sensitive dye optical imaging (Leznik et al. 2002; Leznik & Llinas, 2005). This technique is at present the methodology of choice in studying the geometrical distribution of activity in a large neuronal ensemble (see, for instance, Ebner & Chen, 1995).
We have shown that ensemble oscillations in the IO emanate from clusters of synchronized activity, where each cluster is a localized functional event composed of hundreds of cells. Given the distribution of complex spike activity in the cerebellum cortex, we have proposed that these clusters are very likely to be responsible for the synchronized activation of the PCs observed in previous in vivo multielectrode experiments (Lang et al. 1996). Furthermore, when comparing our experimental results with those obtained by computational modelling of IO neuronal ensembles endowed with oscillatory electrical properties and electrotonic coupling (Makarenko & Llinas, 1998; Velarde et al. 2002), we could show that neuronal oscillatory clustering is a direct consequence of the combined electrotonic/intrinsic properties of coupled IO neurons (Leznik et al. 2002).
While electrical recording of IO neurons in vitro had indicated the possibility that electrically coupled IO cells could actually cluster into synchronized ensemble neuronal groupings, there was no direct demonstration of such dissipative structures. In searching for such dissipative events, voltage-sensitive dye imaging of oscillatory activity was attempted and successfully implemented in rodent IO slices (Leznik et al. 2002). Thus, spatio-temporal profiles of ensemble IO oscillations were unambiguously observed following IO electrical stimuli.
This direct stimulation served to both reset the phase of subthreshold oscillation and to entrain a large proportion of neurons to in-phase oscillations. Indeed, synchronization of oscillatory activity over the IO network increased the amplitude of the optical signal to a level that could be easily detected with our imaging approach. Such oscillatory reset was also simultaneously observed with intracellular recordings from IO neurons (Fig. 1, upper panel intracellular trace (black) compared with the simultaneously recorded optical signal (red)). The optically recorded oscillatory clusters have a dynamic spatial organization, and their amplitude depends on the oscillation phase such that they embraced the largest area during the upward phase of the oscillations. Each cluster consisted of a core region and the adjoining area. The core region demonstrated a close to constant size, but the extent of the adjoining area was found to be phase dependent.
Figure 1. IO oscillations in voltage-sensitve imaging.
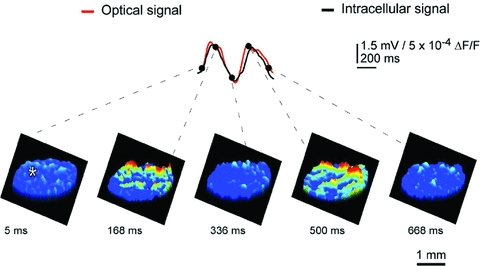
Spontaneous oscillatory electrical activity was acquired simultaneously using optical voltage imaging and intracellular recording. The beginning of oscillatory sequence was defined as 0 ms. The upper panel superimposes the optical signal in red and intracellular voltage recording (asterisk) in black and demonstrates temporal waveform coherence. The lower panels illustrate the spatial distribution of voltage imaging at five different time points indicated by dots in the upper panel from two successive oscillation cycles averaged three times over the oscillatory sequence. Note that ensemble oscillations emanated from several fluorescent clusters of coherent activity, and that the spatial and temporal structure of the IO cluster activity is discernable directly from cluster distribution and size. Time voltage and spatial distribution are as indicated by the calibration bars. (Modified from Leznik & Llinas, 2005.)
We calculated the core area and maximum area (i.e. the core region plus the adjoining area at its utmost extent) for several representative clusters in each experiment. The mean core area and the mean maximum area of a cluster occupied an area of several hundred square micrometres. IO clusters are three-dimensional structures and measurements suggest that they comprise hundreds of cells. Thus, our optical data indicate that at the network level, the IO nucleus is organized in functionally coupled activity clusters. Each cluster is comprised of several hundred cells, which may act in unison to activate groups of thousands of cerebellar PCs simultaneously in agreement with the multiple electrode recordings observed previously.
In conclusion, the dimensions of clusters are probably determined by the IO electrical coupling coefficient, and thus by the magnitude and distribution of the return inhibition from the cerebellar nuclear feedback, which has been demonstrated in previous in vivo experiments (Ruigrok & Voogd, 1995; Lang et al. 1996) and supported with mathematical modelling (Leznik et al. 2002; Velarde et al. 2002).
The climbing fibre conduction isochronicity
From another perspective, while the temporal distribution of activity is well demonstrated at the olivary level, one may wonder about the time dispersion produced by the olivocerebellar pathway given the different distances between the IO axons and their target PCs. However, if isochronicity is present, then the conduction time between an IO neuron and its PC should be close to uniform and independent of the distance such a signal had to travel.
This issue is particularly significant given that the folded nature of such a cortex can increase the path length to the PCs by more than 50%. Furthermore, the correction of the conduction velocity required to insure synchronicity should be related linearly to distance. This was, in fact, the case. The time dispersion for a nearly 4 ms conduction time was plus or minus 500 μs to any regions of the cerebellar mantle, regardless of the distance between the IO and the cerebellar cortex at the bottom or top of the deep cerebellar folia or at any point in between (Sugihara et al. 1993). The results were based on complex spike latency from 660 different PCs from 12 rats (Fig. 2).
Figure 2. Olivocerebellar conduction time is constant throughout the extent of the cerebellar cortex.
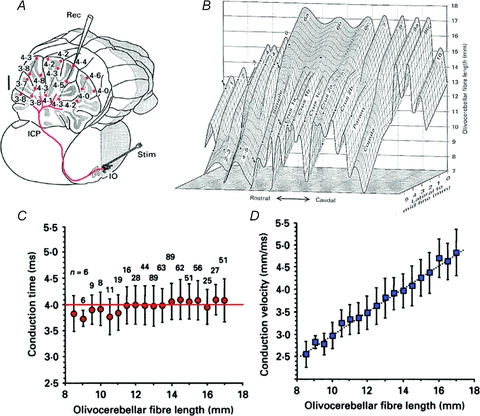
A, diagram of the path for a single climbing fibre (red). Microelectrode recorded Purkinje cell complex spike latency (in ms) at different depths (red dots) is shown following IO electrical stimulation. B, tridimensional representation of climbing fibre length. The X and Z coordinates indicate, respectively, rostrocaudal and mediolateral climbing fibre localization in the different folia (indicated) and the Y coordinate the climbing fibre length. C, conduction time is plotted against climbing fibre length (4 ms (variance ± 500 μs)). D, conduction velocity related linearly to length. (Modified from Sugihara et al. 1993.)
Since our original demonstration, this isochrony has been confirmed in further experiments with other cerebellar systems (Ariel, 2005, Brown & Ariel, 2009). A similar finding concerning conduction isochronicity has also been observed in the thalamocortical system and has been interpreted, as in the case of the olivocerebellar system, as a mechanism for temporal coherence. In this case, such timing has been related to the temporal coherence associated with cognitive binding (Engel et al. 1997; Salami et al. 2003; Chomiak et al. 2008; Vicente et al. 2008).
Therefore, the results indicated that the cerebellar cortex, while being deeply folded anatomically, behaves functionally as an isochronous sphere as far as the olivocerebellar system is concerned. Further, such isochronicity is actually related to the onset time and duration required for proper motor execution (Welsh et al. 1995).
The olivocerebellar system and error sensing
Finally, the issue of error sensing, which was previously of great interest to cerebellar physiologists, has been treated in detail in excellent reviews concerning IO function (Simpson et al. 1996). My personal view is that the error-sensing signal that is often observed in climbing fibre responses – while being a very important functional phenotype – is not the central cerebellar function as some authors claim. From my perspective, the high probability of complex spike activation in relation to unexpected error signals correlates well with such events simply because it is easy to detect. This is the case because climbing fibre activation is massive both when a large reset of the oscillatory phase occurs (Makarenko & Llinas, 1998; Leznik et al. 2002; Chorev et al. 2007; Khosrovani et al. 2007; see also Van der Giessen et al. 2008 for the connexin 36 role in this reset) and when a massive temporal reorganization of motor pattern activity is required.
Experimental findings
This “reset” question was addressed in studies of rodent brainstem slices. In agreement with previous intracellular results (Llinas & Yarom, 1986), an extracellular stimulation given at the dorsal border of the IO nucleus generates a full action potential followed by a membrane hyperpolarization in nearby neurons. These results also demonstrate that if the cell was oscillating at the time of the stimulus, its oscillations are stopped momentarily, but resumed with a different phase shortly after the stimulation (Leznik E & Llinas R 2005).
Moreover, in later experiments, it was also determined that such extracellular stimulation may reset the phase without affecting the amplitude or frequency of the subthreshold oscillation (Leznik et al. 2002), and that for most cells recorded, this phase reset could be observed repeatedly with subsequent stimuli (Fig. 3). However, the most surprising property discovered was the fact that the oscillation phase shift was remarkably constant (Fig. 3C (insert) and D) and independent of the original phase moment at which the stimulus was delivered.
Figure 3. Intracellular in vitro recording of spontaneous subthreshold oscillations and phase reset by an electrical extracellular stimulus.
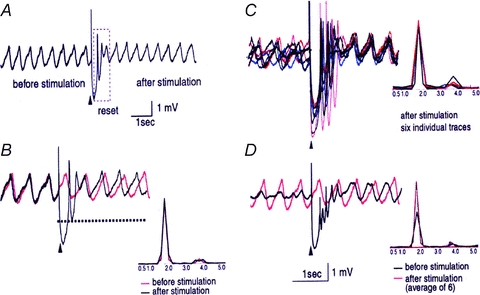
A, following an extracellular stimulation (marked with an arrowhead), the oscillations disappeared for 750 ms (boxed area) and then resumed. The membrane potential was –60 mV. B, superposition of intracellular recordings of spontaneous (dashed black line) and stimulus-evoked (continuous black line) oscillations in the same cell. Their corresponding power spectra are shown below. Note that extracellular stimulation only modified the phase of the spontaneous oscillations without affecting their amplitude or frequency. C, superposition of six individual intracellular traces of stimulus-evoked oscillations from the same cell. Each trace is shown in a different colour. Their corresponding power spectra are displayed below. Note that in each trace, the stimulation-induced shift in the cell's oscillatory rhythm is remarkably similar. Oscillations are seen clearly after the stimulus-induced reset but can barely be detected before the stimulation. Calibration bar: 1 mV 1 s. (Leznik, Makarenko & Llinas, 2002.)
This constant phase shift is of central importance in defining IO function, as it gives a clear time constraint to the functional states generated by the neuronal ensemble. The reset property of the IO circuit can thus be considered as the main component in the large correction that must be generated when a movement error occurs. This is best illustrated by the fast recovery that we all experience when tripping during locomotion and the fact that we do not fall, while robots do, under similar circumstances.
The issue of error correction has been studied elegantly under conditions where random stimuli require temporal resting under circumstances of robust activation of the cerebellar system (Schweighofer et al. 2004); however, this issue must be addressed further as other views are also clearly present (Horn et al. 2004). The image one has is of the activation of a very large population of Purkinje cells that mediate a rapid inhibition of the inhibitory cells of the nucleo-olivary pathway, resulting in increased coupling at the olivary level. This event will produce a large and coherent activation of IO neurons; thus, an increased probability of PC complex spike activation ensues. In short, then, error correction is one mode, but not the main mode of IO function.
Modelling of IO function
From a motor control perspective, a previous study (Makarenko & Llinas, 1998) demonstrated that the dynamics of IO subthreshold oscillations are robust but sensitive with respect to their interactions between themselves (‘long-term’ stimulation by electrotonic coupling) and external stimuli (‘short-term’ stimulation by synaptic input). From that study, it was determined that IO dynamics exhibit weakly chaotic properties allowing almost regular periodic oscillations, and a non-linear sensitivity to the initial conditions, which results in the fast renewal of the system's memory (at 4 ms). Indeed, the phase plane portrait of the oscillations demonstrates radial trajectory divergence (stretching and folding) with little azimuthally divergence as the result of the kinetics of the channels responsible for the membrane potential oscillations.
These experimental findings were supported by a formal model of subthreshold oscillations in IO neurons using a Rössler-type non-linear system (Makarenko & Llinas, 1998). A more recent study using a non-linear dynamical system (Kazantsev et al. 2003) to model the self-referential phase reset demonstrated an excellent fit to the experimental data (Llinas & Yarom, 1986; Lang et al. 1999).
Moreover, this network dynamic has true non-trivial attributes concerning the type of global motor control function implemented by the olivocerebellar network. Since its response is independent of the IO oscillatory phase when the stimulus arrives, the system demonstrates extraordinary flexibility in organizing a given motor intention and in modifying its activity according to sensory feedback. The system does not need an operational memory.
This makes it very reliable and prevents ‘computational overloads’ that appear when memorizing the states. In fact, the speed of the operation (on the order of an oscillation period) is limited only by the oscillatory frequency, and thus it can operate many times faster than the actuators it controls.
Artificial control systems
The motor control property of the IO oscillator can be very clearly demonstrated in artificial control systems. Indeed, the oscillatory network embodies a phase controller that can set and maintain a required phase by delivering an appropriate stimulus pulse to the oscillator. If a system is developed where the oscillatory phase is associated with a given physical parameter (e.g. position, velocity, angle, or temperature) such a phase controller can maintain a pre-specified posture or motor sequence. In contrast with standard control systems, the controlling principle deals with a ‘motor recovery response’ (Pikovsky et al. 2001).
Within limits, a walking animal may stumble without falling and recovers its walking rhythm promptly irrespective of when an obstacle is encountered during the stride. Such synchronization of a large oscillator array requires only a synchronous reset stimulus. A fundamental aspect concerns the possibility of phase encoding when the map has both periodic orbits and chaotic attractors. In this case, the phase resets associated with these orbits are naturally transformed into neuronal spiking. In turn, neurons fire spike trains with inter-spike intervals that are correlated directly with the orbits.
Finally, the effect of the phase reset can be viewed as an effective tool to represent and/or store information in the form of oscillatory clusters. In contrast to Hopfield gradient networks (Hopfield, 1982), cluster reorganization can be extremely fast because the oscillator frequency may be moved up to the limits of the constituent materials. This is compared with Kuramoto-like oscillatory systems (Kuramoto, 2003), where a complex coupling matrix can work directly with information converted to the stimulus template. In fact, a recent paper (Bandyopadhyay et al. 2009) describes an actual biologically inspired underwater vehicle using olivocerebellar dynamics as a motor control system, which demonstrates that an analog IO-based circuit allows better control per unit time than a numerically controlled system.
Conclusions and implications
Three main issues have been addressed in this short paper concerning the functional organization of the olivocerebellar system. (1) The olivocerebellar system seems to be related centrally to the control of motor timing. Its exceptional neuronal characteristics, and the network properties that it supports, make the olivocerbellar system a unique control system, where timing seems to be a central theme. (2) The combination of strong and rather stereotyped intrinsic electrical properties with electrical coupling among the neuronal elements allows the synchronous activation of clusters of neurons. Further, feedback inhibition provides the dynamic variance of the membership of such coupled clusters. (3) And finally, the very fundamental property of the resetting of the phase of groups of neurons by a stimulus, such that the new phase is coherent and independent from the original phase, makes this event truly spectacular. These three elements give the IO a very powerful set of network properties allowing not only the temporal control of many variables simultaneously, as occurs during motor control, but also the possibility of rapid correction in the presence of unexpected events that require rapid global motor correction.
Finally, nature has evolved a mechanism by which this very elaborate cluster dynamic generating system can transmit the timing sequences into a folded cortical geometry, without differential conduction time aberrations, and terminate its path by generating the most powerful synapse in the CNS. If this were not sufficient, the neurons it activates are the largest in the brain, they receive just one such climbing fibre afferent, and its output is inhibitory (Ito & Yoshida, 1966).
And so, nature has evolved the cerebellar circuit, one of its most conserved neuronal systems (Llinas, 1974) to control motricity by inhibition, a very fitting attribute given that selection, via inhibition, is ubiquitous as the mechanism for neuronal pattern generation in the CNS.
References
- Albus JS. A theory of cerebellar function. Math Biosci. 1971;10:25–61. [Google Scholar]
- Ariel M. Latencies of climbing fiber inputs to turtle cerebellar cortex. J Neurophysiol. 2005;93:1042–1054. doi: 10.1152/jn.00132.2004. [DOI] [PubMed] [Google Scholar]
- Armstrong DM, Schild RF. A quantitative study of the Purkinje cells in the cerebellum of the albino rat. J Comp Neurol. 1970;139:449–456. doi: 10.1002/cne.901390405. [DOI] [PubMed] [Google Scholar]
- Bal T, McCormick DA. Synchronized oscillations in the inferior olive are controlled by the hyperpolarization-activated cation current Ih. J Neurophysiol. 1997;77:3145–3156. doi: 10.1152/jn.1997.77.6.3145. [DOI] [PubMed] [Google Scholar]
- Bandyopadhyay P, Singh S, Thivierge D, Annaswamy A, Leinhos H, Fredette A, Beal D. Synchronization of animal-inspired multiple high-lift fins in an underwater vehicle using olivo-cerebellar dynamics. Oceanic Eng IEEE J. 2009;33:563–578. [Google Scholar]
- Bell C, Kawasaki T. Relations among climbingfibre responses of nearby Purkinje cells. J Neurophysiol. 1972;35:155–169. doi: 10.1152/jn.1972.35.2.155. [DOI] [PubMed] [Google Scholar]
- Benardo LS, Foster RE. Oscillatorybehaviour in inferior olive neurons: mechanism, modulation, cell aggregates. Brain Res Bull. 1986;17:773–784. doi: 10.1016/0361-9230(86)90089-4. [DOI] [PubMed] [Google Scholar]
- Bennett M. Electrical synapses, a personal perspective (or history) Brain Res Brain Res Rev. 2000;32:16–28. doi: 10.1016/s0165-0173(99)00065-x. [DOI] [PubMed] [Google Scholar]
- Biewener AA. Patterns of mechanical energy change in tetrapod gait: pendula, springs and work. J Exp Zool A Comp Exp Biol. 2006;305:899–911. doi: 10.1002/jez.a.334. [DOI] [PubMed] [Google Scholar]
- Bower JM, Woolston DC. Congruence of spatial organization of tactile projections to granule cell and Purkinje cell layers of cerebellar hemispheres of the albino rat: vertical organization of cerebellar cortex. J Neurophysiol. 1983;49:745–766. doi: 10.1152/jn.1983.49.3.745. [DOI] [PubMed] [Google Scholar]
- Brown M, Ariel M. Topography and response timing of intact cerebellum stained with absorbance voltage-sensitive dye. J Neurophysiol. 2009;101:474–490. doi: 10.1152/jn.90766.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cajal R. Sobre las fibras nerviosas de la capa molecular del cerebelo. Trimestr Histol. 1888;2:33–41. [Google Scholar]
- Cavagna GS, Thys H, Zamboni A. The sources of external work in level walking and running. J Physiol. 1976;262:639–657. doi: 10.1113/jphysiol.1976.sp011613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chomiak T, Peters S, Hu B. Functional architecture and spike timing properties of corticofugal projections from rat ventral temporal cortex. J Neurophysiol. 2008;100:327–335. doi: 10.1152/jn.90392.2008. [DOI] [PubMed] [Google Scholar]
- Chorev E, Yarom Y, Lampl I. Rhythmic episodes of subthreshold membrane potential oscillations in the rat inferior olive nuclei in vivo. J Neurosci. 2007;27:5043–5052. doi: 10.1523/JNEUROSCI.5187-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark WS. How fast is the fastest bird? Wild Bird. 1995;9:42–43. [Google Scholar]
- Crill W. Unitary multiple-spiked responses in cat inferior olive nucleus. J Neurophysiol. 1970;33:199–209. doi: 10.1152/jn.1970.33.2.199. [DOI] [PubMed] [Google Scholar]
- D'Angelo ES, Koekkoek KE, Lombardo P, Solinas S, Ros E, Garrido J, Schonewille M, De Zeeuw CI. Timing in the cerebellum: oscillations and resonance in the granular layer. Neuroscience. 2009;162:805–815. doi: 10.1016/j.neuroscience.2009.01.048. [DOI] [PubMed] [Google Scholar]
- D'Angelo ES. A realistic large-scale model of the cerebellum granular layer predicts circuit spatio-temporal fltering properties. Front Cell Neurosci. 2010;4:12. doi: 10.3389/fncel.2010.00012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Zeeuw C, Simpson J, Hoogenraad C, Galjart N, Koekkoek S, Ruigrok T. Microcircuitry and function of the inferior olive. Trends Neurosci. 1998;21:391–400. doi: 10.1016/s0166-2236(98)01310-1. [DOI] [PubMed] [Google Scholar]
- de Zeeuw CI, Holstege JC, Ruigrok TJ, Voogd J. Ultrastructural study of the GABAergic, cerebellar, and mesodiencephalic innervation of the cat medial accessory olive: anterograde tracing combined with immunocytochemistry. J Comp Neurol. 1989;284:12–35. doi: 10.1002/cne.902840103. [DOI] [PubMed] [Google Scholar]
- De Zeeuw CI, Lang EJ, Sugihara I, Ruigrok TJ, Eisenman LM, Mugnaini E, Llinas R. Morphological correlates of bilateral synchrony in the rat cerebellar cortex. J Neurosci. 1996;16:3412–3426. doi: 10.1523/JNEUROSCI.16-10-03412.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebner TJ, Chen G. Use of voltage-sensitive dyes and optical recordings in the central nervous system. Progr Neurobiol. 1995;46:463–506. doi: 10.1016/0301-0082(95)00010-s. [DOI] [PubMed] [Google Scholar]
- Eccles JC, Llinas R, Sasaki K. Excitation of cerebellar Purkinje cells by the climbing fibres. Nature 203. 1964:245–246. doi: 10.1038/203245a0. [DOI] [PubMed] [Google Scholar]
- Eccles JC, Llinas R, Sasaki K. The excitatory synaptic action of climbing fibres on the Purkinje cells of the cerebellum. J Physiol. 1966a;182:268–296. doi: 10.1113/jphysiol.1966.sp007824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eccles JC, Llinas R, Sasaki K. The mossy fibre-granule cell relay of the cerebellum and its inhibitory control by Golgi cells. Exp Brain Res. 1966b;1:82–101. doi: 10.1007/BF00235211. [DOI] [PubMed] [Google Scholar]
- Engel AK, Roelfsema PR, Fries P, Brecht M, Singer W. Role of the temporal domain for response selection and perceptual binding. Cereb Cortex. 1997;7:571–582. doi: 10.1093/cercor/7.6.571. [DOI] [PubMed] [Google Scholar]
- Fredette BJ, Mugnaini E. The GABAergic cerebello-olivary projection in the rat. Anat Embryol. 1991;184:225–243. doi: 10.1007/BF01673258. [DOI] [PubMed] [Google Scholar]
- Fukuda M, Yamamoto T, Llinas R. The isochronic band hypothesis and climbing fibre regulation of motricity: an experimental study. Eur J Neurosci. 2001a;13:315–326. doi: 10.1046/j.0953-816x.2000.01394.x. [DOI] [PubMed] [Google Scholar]
- Fukuda M, Yamamoto Y, Llinas R. The isochronic band hypothesis and climbingfibre regulation of motricity: an experimental study. Eur J Neurosci. 2001b;13:315–326. doi: 10.1046/j.0953-816x.2000.01394.x. [DOI] [PubMed] [Google Scholar]
- Hopfield J. Neural networks and physical systems with emergent collective computational abilities. Proc Natl Acad Sci U S A. 1982;79:2549–2535. doi: 10.1073/pnas.79.8.2554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horn KM, Pong M, Gibson AR. Discharge of inferior olive cells during reaching errors and perturbations [erratum appears in Brain Res 1008, 137–138] Brain Res. 2004;996:148–158. doi: 10.1016/j.brainres.2003.10.021. [DOI] [PubMed] [Google Scholar]
- Ito M. Cerebellar long-term depression: characterization, signal transduction and functional roles. Physiol Rev. 2001;81:1143–1195. doi: 10.1152/physrev.2001.81.3.1143. [DOI] [PubMed] [Google Scholar]
- Ito M, Yoshida M. The origin of cerebellar-induced inhibition of Deiters neurons. Exp Brain Res. 1966;2:330–349. doi: 10.1007/BF00234779. [DOI] [PubMed] [Google Scholar]
- Ito M, Kano M. Long-lasting depression of parallel fibre-Purkinje cell transmission induced by conjunctive stimulation of parallel fibres and climbing fibres in the cerebellar cortex. Neurosci Lett. 1982;33:253–258. doi: 10.1016/0304-3940(82)90380-9. [DOI] [PubMed] [Google Scholar]
- Jorntell H, Ekerot CF. Reciprocal bidirectional plasticity of parallel fiber receptive fields in cerebellar Purkinje cells and their afferent interneurons. Neuron. 2002;34:797–806. doi: 10.1016/s0896-6273(02)00713-4. [DOI] [PubMed] [Google Scholar]
- Kazantsev VB, Nekorkin VI, Makarenko VI, Llinas R. Olivo-cerebellar cluster-based universal control system. Proc Natl Acad Sci U S A. 2003;100:13064–13068. doi: 10.1073/pnas.1635110100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khosrovani S, Van Der Giessen RS, De Zeeuw CI, De Jeu MTG. In vivo mouse inferior olive neurons exhibit heterogeneous subthreshold oscillations and spiking patterns. Proc Natl Acad Sci U S A. 2007;104:15911–15916. doi: 10.1073/pnas.0702727104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuramoto Y. Chemical Oscillations, Waves, and Turbulence. Courier Dover Publications, Mineola; 2003. [Google Scholar]
- Lampl I, Yarom Y. Subthreshold oscillations and resonantbehaviour: two manifestations of the same mechanism. Neuroscience. 1997;78:325–341. doi: 10.1016/s0306-4522(96)00588-x. [DOI] [PubMed] [Google Scholar]
- Lang E. Organization of olivocerebellar activity in the absence of excitatory glutamatergic input. J Neurosci. 2001;21:1663–1675. doi: 10.1523/JNEUROSCI.21-05-01663.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang E. GABAergic and glutamatergic modulation of spontaneous and motor-cortex-evoked complex spike activity. J Neurophysiol. 2002;87:1993–2008. doi: 10.1152/jn.00477.2001. [DOI] [PubMed] [Google Scholar]
- Lang EJ, Sugihara I, Llinas R. GABAergic modulation of complex spike activity by the cerebellar nucleoolivary pathway in rat. J Neurophysiol. 1996;76:255–275. doi: 10.1152/jn.1996.76.1.255. [DOI] [PubMed] [Google Scholar]
- Lang EJ, Sugihara I, Welsh JP, Llinas R. Patterns of spontaneous purkinje cell complex spike activity in the awake rat. J Neurosci. 1999;19:2728–2739. doi: 10.1523/JNEUROSCI.19-07-02728.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leznik E, Llinas R. Role of gap junctions in synchronized neuronal oscillations in the inferior olive. J Neurophysiol. 2005;94:2447–2456. doi: 10.1152/jn.00353.2005. [DOI] [PubMed] [Google Scholar]
- Leznik E, Makarenko V, Llinas R. Electrotonically mediated oscillatory patterns in neuronal ensembles: an in vitro voltage-dependent dye-imaging study in the inferior olive. J Neurosci. 2002;22:2804–2815. doi: 10.1523/JNEUROSCI.22-07-02804.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas R. Eighteenth Bowditch Lecture: Motor aspects of cerebellar control. Physiologist. 1974;17:19–46. [PubMed] [Google Scholar]
- Llinas RR. The intrinsic electrophysiological properties of mammalian neurons: insights into central nervous system function. Science. 1988;242:1654–1664. doi: 10.1126/science.3059497. [DOI] [PubMed] [Google Scholar]
- Llinas R, Muhlethaler M. An electrophysiological study of the in vitro, perfused brain stem-cerebellum of adult guinea-pig. J Physiol. 1988;404:215–240. doi: 10.1113/jphysiol.1988.sp017287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas R, Sasaki K. The functional organization of the olivo-cerebellar system as examined by multiple Purkinje cell recordings. Eur J Neurosci. 1989;1:587–602. doi: 10.1111/j.1460-9568.1989.tb00365.x. [DOI] [PubMed] [Google Scholar]
- Llinas R, Walton K, Hillman DE, Sotelo C. Inferior olive: its role in motor learning. Science. 1975;190:1230–1231. doi: 10.1126/science.128123. [DOI] [PubMed] [Google Scholar]
- Llinas R, Yarom Y. Electrophysiology of mammalian inferior olivary neurones in vitro. Different types of voltage-dependent ionic conductances. J Physiol. 1981;315:549–567. doi: 10.1113/jphysiol.1981.sp013763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Llinas R, Yarom Y. Oscillatory properties of guinea-pig inferior olivary neurones and their pharmacological modulation: an in vitro study. J Physiol. 1986;376:163–182. doi: 10.1113/jphysiol.1986.sp016147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marr D. A theory of the cerebellar cortex. J Physiol. 1969;202:437–470. doi: 10.1113/jphysiol.1969.sp008820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makarenko V, Llinas R. Experimentally determined chaotic phase synchronization in a neuronal system. Proc Natl Acad Sci U S A. 1998;95:15747–15752. doi: 10.1073/pnas.95.26.15747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medina JF, Nores WL, Mauk MD. Inhibition of climbing fibres is a signal for the extinction of conditioned eyelid responses. Nature. 2002;416:330–333. doi: 10.1038/416330a. [DOI] [PubMed] [Google Scholar]
- Nicholson DA, Freeman JH., Jr Developmental changes in evoked Purkinje cell complex spike responses [erratum appears in J Neurophysiol 91, 1108] J Neurophysiol. 2003;90:2349–2357. doi: 10.1152/jn.00481.2003. [DOI] [PubMed] [Google Scholar]
- Palay S, Chan-Palay V. Cerebral Cortex Cytology and Organization. New York: Springer-Verlag; 1974. [Google Scholar]
- Pikovsky A, Rosenblum M, Kurths J. Synchronization: A Universal Concept in Nonlinear Sciences. Cambridge: Cambridge University Press; 2001. [Google Scholar]
- Ruigrok TJ, Voogd J. Cerebellar influence on olivary excitability in the cat. Eur J Neurosci. 1995;7:679–693. doi: 10.1111/j.1460-9568.1995.tb00672.x. [DOI] [PubMed] [Google Scholar]
- Salami M, Itami C, Tsumoto T, Kimura F. Change of conduction velocity by regional myelination yields constant latency irrespective of distance between thalamus and cortex. Proc Natl Acad Sci U S A. 2003;100:6174–6179. doi: 10.1073/pnas.0937380100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schweighofer N, Doya K, Fukai H, Chiron JV, Furukawa T, Kawato M. Chaos may enhance information transmission in the inferior olive. Proc Natl Acad Sci U S A. 2004;101:4655–4660. doi: 10.1073/pnas.0305966101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schonewille M, Gao Z, Boele HJ, Vinueza Veloz MF, Amerika WE, Simek AA, De Jeu MT, Steinberg JP, Takamiya K, Hoebeek FE, Linden DJ, Huganir RL, De Zeeuw CL. Reevaluating the role of LTD in cerebellar motor learning. Neuron. 2011;70:43–50. doi: 10.1016/j.neuron.2011.02.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson J, Wylle D, De Zeeuw CI. On climbingfibre signals and their consequences. Behav Brain Sci. 1996;19:496–527. [Google Scholar]
- Sharp NNC. Timed running speed of a cheetah (Acinonyx jubatus) J Zool. 1997;241:493–494. [Google Scholar]
- Sotelo C, Gotow T, Wassef M. Localization of glutamic-acid-decarboxylase-immunoreactive axon terminals in the inferior olive of the rat, with special emphasis on anatomical relations between GABAergic synapses and dendrodendritic gap junctions. J Comp Neurol. 1986;252:32–50. doi: 10.1002/cne.902520103. [DOI] [PubMed] [Google Scholar]
- Sotelo C, Llinas R, Baker R. Structural study of inferior olivary nucleus of the cat: morphological correlates of electrotonic coupling. J Neurophysiol. 1974;37:541–559. doi: 10.1152/jn.1974.37.3.541. [DOI] [PubMed] [Google Scholar]
- Strata P, Montarolo PG. Functional aspects of the inferior olive. Archives Italiennes de Biologie. 1982;120:321–329. [PubMed] [Google Scholar]
- Sugihara I, Lang EJ, Llinas R. Uniform olivocerebellar conduction time underlies Purkinje cell complex spike synchronicity in the rat cerebellum. J Physiol. 1993;470:243–271. doi: 10.1113/jphysiol.1993.sp019857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Szentagothai J, Rajkovits K. Ueber den Ursprung der Kletterfasern des kleinhirns. Z Anat Entwicklungsgeschichte. 1959;121:130–141. [Google Scholar]
- Vallbo AB, Wessberg J. Organization of motor output in slow finger movements in man. J Physiol. 1993;469:673–691. doi: 10.1113/jphysiol.1993.sp019837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Der Giessen RS, Koekkoek SK, van Dorp S, Jornt R, De Gruijl JR, Cupido A, Khosrovani S, Dortland B, Wellershaus K, Degen J, Deuchar J, Fuchs EC, Monyer H, Willecke K, De Jeu MTG, De Zeeuw CI. Role of olivary electrical coupling in cerebellar motor learning. Neuron. 2008;58:599–612. doi: 10.1016/j.neuron.2008.03.016. [DOI] [PubMed] [Google Scholar]
- Velarde MG, Nekorkin VI, Kazantsev VB, Makarenko VI, Llinas R. Modeling inferior olive neuron dynamics. Neural Networks. 2002;15:5–10. doi: 10.1016/s0893-6080(01)00130-7. [DOI] [PubMed] [Google Scholar]
- Vicente R, Gollo LL, Mirasso CR, Fischer I, Pipa G. Dynamical relaying can yield zero time lag neuronal synchrony despite long conduction delays. Proc Natl Acad Sci U S A. 2008;105:17157–17162. doi: 10.1073/pnas.0809353105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welsh JP, Lang EJ, Suglhara I, Llinas R. Dynamic organization of motor control within the olivocerebellar system. Nature. 1995;374:453–457. doi: 10.1038/374453a0. [DOI] [PubMed] [Google Scholar]
- Welsh JP, Llinas R. Some organizing principles for the control of movement based on olivocerebellar physiology. Progr Brain Res. 1997;114:449–461. doi: 10.1016/s0079-6123(08)63380-4. [DOI] [PubMed] [Google Scholar]
- Welsh JP, Yamaguchi H, Zeng XH, Kojo M, Nakada Y, Takagi, Sugimori M, Llinas RR. Normal motor learning during pharmacological prevention of Purkinje cell long-term depression. Proc Natl Acad Sci USA. 2005;102:17166–17171. doi: 10.1073/pnas.0508191102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yarom Y, Cohen D. The olivocerebellar system as a generator of temporal patterns. Ann N Y Acad Sci. 2002;978:122–134. doi: 10.1111/j.1749-6632.2002.tb07561.x. [DOI] [PubMed] [Google Scholar]