

Abstract
GATA transcription factors are required for the differentiation of diverse cell types in several species. Recent evidence suggests that their biologic activities may be modulated through interaction with multitype zinc finger proteins, such as Friend of GATA-1 (FOG) and U-shaped (Ush). In cell culture, FOG cooperates with the hematopoietic transcription factor GATA-1 to promote erythroid and megakaryocytic differentiation. We show here that mice lacking FOG die during mid-embryonic development with severe anemia. FOG−/− erythroid cells display a marked, but partial, blockage of maturation, reminiscent of GATA-1− erythroid precursors. In contrast to GATA-1 deficiency, however, megakaryocytes fail to develop in the absence of FOG. Although the FOG−/− erythroid phenotype supports the proposed role of FOG as a GATA-1 cofactor in vivo, the latter finding points to a pivotal, GATA-1-independent requirement for FOG in megakaryocyte development from the bipotential erythroid/megakaryocytic progenitor. We speculate that FOG and other FOG-like proteins serve as complex cofactors that act through both GATA-dependent and GATA-independent mechanisms.
Keywords: Friend of GATA-1 (FOG), GATA-1, cofactor, gene targeting, hematopoiesis
GATA transcription factors have emerged as central regulators of diverse developmental processes in both vertebrate and invertebrate species. Members of the GATA family are distinguished by a DNA-binding domain comprised of one or two highly conserved, C4-type zinc fingers that mediate sequence-specific binding to the consensus motif (A/T)GATA(A/G) (Evans and Felsenfeld 1989; Tsai et al. 1989). In vertebrates, six GATA factors (GATA-1 through GATA-6) have been identified, each serving distinct roles in development (Pevny et al. 1991; Laverriere et al. 1994; Tsai et al. 1994; Ting et al. 1996; Kuo et al. 1997; Molkentin et al. 1997). GATA factors have also been isolated from Drosophila (Ramain et al. 1993), Caenorhabditis elegans (Spieth et al. 1991; Zhu et al. 1997), and fungi such as Aspergillus (Kudla et al. 1990).
GATA-1, the founding member of the GATA factor family, is perhaps the most extensively studied of the vertebrate GATA factors. A hematopoietic-restricted transcription factor, GATA-1 is expressed at high levels in erythroid, megakaryocytic, mast, and eosinophilic cells and at lower levels in multipotential progenitors (for review, see Orkin 1998). Consistent with the finding that virtually all characterized erythroid- and megakaryocytic-specific genes contain GATA motifs in critical cis-regulatory elements (Orkin 1992; Shivdasani 1997), gene targeting experiments in embryonic stem (ES) cells and mice have established an essential role for GATA-1 in both the erythroid and megakaryocytic lineages. Loss of GATA-1 in erythroid cells leads to a developmental arrest at the proerythroblast stage and apoptosis (Pevny et al. 1991, 1995; Weiss et al. 1994; Weiss and Orkin 1995), resulting in embryonic lethality by day 11.5 of gestation (Fujiwara et al. 1996). Megakaryocytes lacking GATA-1 also arrest in their maturation, but undergo unrestrained proliferation rather than apoptosis (Shivdasani et al. 1997). In addition to driving terminal maturation of erythroid and megakaryocytic cells, GATA-1 is also capable of influencing the cellular phenotype of recipient hematopoietic progenitors. Specifically, forced expression of GATA-1 induces differentiation of mouse myeloid 416B cells along the megakaryocytic lineage (Visvader et al. 1992) and myb–ets-transformed chicken myeloblasts along erythroid, thromboblastic (megakaryocytic), and eosinophilic lineages (Kulessa et al. 1995).
Fundamental to understanding how GATA-1 functions in differentiation and development is the elucidation of the mechanisms by which it acts in transcription. Although early studies implicated the amino-terminal activation domain of GATA-1 as critical for function based on its transactivation potential in fibroblastic cells (Martin and Orkin 1990), more recent studies have suggested that the biological activity of GATA-1 actually resides in its zinc finger DNA-binding domain, as assayed in 416B cells (Visvader et al. 1995), GATA-1− embryoid bodies (Blobel et al. 1995), and GATA-1− erythroid precursor cells (Weiss et al. 1997). Moreover, the amino-terminal zinc finger of GATA-1, which is largely dispensable for DNA binding and for transactivation in fibroblasts, is absolutely required for terminal erythroid differentiation (Weiss et al. 1997). These findings led to the proposal that a transcriptional cofactor might be required that would associate with DNA-bound GATA-1 and link it to larger transcriptional complexes (Weiss et al. 1997).
In a yeast two-hybrid screen for GATA-1-interacting proteins, we recently identified a novel, multitype zinc finger protein Friend of GATA-1 (FOG) (Tsang et al. 1997). Several lines of evidence support its proposed role as a GATA-1 cofactor. First, FOG interacts specifically with the amino finger of GATA-1 in vitro and in vivo. Second, the expression of FOG parallels that of GATA-1 during early embryonic development. Within adult hematopoietic lineages, both are highly expressed in erythroid and megakaryocytic cells. Third, FOG synergizes with GATA-1 in activating transcription from a hematopoietic-specific regulatory region, and cooperates with GATA-1 in cell culture to promote erythroid and megakaryocytic differentiation. Finally, FOG is structurally similar to the recently characterized Drosophila cofactor U-shaped (Ush) (Cubadda et al. 1997; Haenlin et al. 1997). Like FOG, Ush is a complex zinc finger protein that interacts with the amino finger of the Drosophila GATA factor Pannier (Ramain et al. 1993). However, in contrast to FOG, Ush negatively regulates Pannier’s transcriptional activity. Taken together, these results suggest that FOG and Ush define a novel class of cofactors that modulate, either positively or negatively, the transcriptional activities of GATA factors in diverse developmental settings.
To test this paradigm in mammalian development, we have disrupted the FOG gene by homologous recombination in ES cells. We show here that mice lacking FOG die between embryonic (E) days 10.5 and 12.5 with severe anemia. FOG−/− erythroid cells exhibit a marked, but partial, arrest in development at the proerythroblast stage, reminiscent of GATA-1− erythroid precursors. In contrast to GATA-1 deficiency, however, loss of FOG leads to the specific ablation of the megakaryocytic lineage. These findings demonstrate an in vivo requirement for FOG that supports its proposed role as a GATA-1 cofactor in erythroid cells and, furthermore, establish a pivotal, GATA-1 independent role for FOG during the earliest stages in megakaryocyte development.
Results
Targeted disruption of the murine FOG gene results in early embryonic lethality
To disrupt the FOG gene, we constructed two different targeting vectors, each of which replaced coding regions by selectable markers (Fig. 1A). Two independently targeted ES cell clones, one derived from each targeting vector (Fig. 1B), were injected into C57BL/6 blastocysts and gave germ-line transmission. Heterozygous mice (FOG+/−) appeared normal and were interbred to generate homozygous mutants (FOG−/−). Among 90 liveborn offspring of FOG+/− crosses, none was homozygous mutant (Table 1), indicating that FOG−/− embryos die in utero. To determine the developmental stage at which the FOG gene mutation is lethal, E10.5–E13.5 embryos were examined. Whereas at E10.5 ∼26% of viable embryos were of the FOG−/− genotype (Fig. 1C), at E11.5 only ∼14% of viable embryos were FOG−/−, and none survived to E12.5 (Table 1). Thus, homozygous FOG−/− embryos die between E10.5 and E12.5. No phenotypic differences were observed between the two independently targeted mouse lines. Absence of FOG expression in mutant embryos was confirmed by RT–PCR analysis for intact FOG transcripts (Fig. 1D) and by immunofluorescence analysis using antibodies directed against the amino terminus of the FOG protein (data not shown).
Figure 1.




Targeted disruption of the mouse FOG gene. (A) Partial restriction map of the mouse FOG locus (top) and structures of the two FOG targeting vectors (middle and bottom). The first targeting construct (pPNT FOG) contains the HSV–tk and neomycin resistance (neor) genes, both under the control of the mouse phosphoglycerate kinase (PGK) promoter; the second construct (pTKLNCL FOG) contains HSV–tk and neor genes, as well as the cytosine deaminase gene. Homologous recombination results in disruption of the coding sequence of FOG and removal of exons encoding zinc fingers (z) 1–z7 or z1–z4 of FOG, respectively. FOG-coding exons are depicted as empty boxes; the numbers (1–9) below these boxes denote the positions of the nine zinc fingers of the FOG protein. Small half-arrows above FOG exons indicate positions of RT–PCR primers. The flanking probes used for Southern blot analysis are shown as black bars. Thin black boxes in pTKLNCL FOG represent loxP sites. (B) BamHI; (Bg) BglII; (Bs) BstEII; (E) EcoRI; (Hd) HindIII; (K) KpnI; (N) NotI; (X) XbaI. (B) Southern blot analysis of G418- and Gancyclovir-resistant ES cell clones. Note that one targeted ES clone was derived from each construct and used to generate germ-line transmitting chimeras. (wt) wild-type allele; (mut) mutant allele. (C) Southern blot analysis of E10.5 embryos resulting from an intercross of FOG+/− mice, demonstrating the presence of all expected genotypes, including homozygous mutants. (D) RT–PCR analysis of total RNA from peripheral blood of viable E11.5 embryos. RNA was amplified with FOG-specific primers that bracket the 5′ position of the neoR cassette insertion (see A). As a control, RT–PCR was also performed for HPRT RNA.
Table 1.
Genotypes of viable progeny derived from heterozygous parents
| Stage |
Total |
Genotype
|
||
|---|---|---|---|---|
| +/+
|
+/−
|
−/−
|
||
| Postnatal | 90 | 30 | 60 | 0 |
| E13.5 | 12 | 4 | 8 | 0 |
| E12.5 | 22 | 10 | 12 | 0 |
| E11.5 | 156 | 43 | 91 | 22 |
| E10.5 | 111 | 26 | 56 | 29 |
Genotyping of offspring of FOG+/− intercrosses was done by Southern blot analysis using tail DNA at the postnatal stage and embryo or yolk sac DNA at embryonic stages. Viable embryos were defined as those with beating hearts. Data shown are for mice derived from one of the two independently targeted ES cell clones. Similar data were obtained for mice derived from the second ES clone.
At E11.5, viable FOG−/− embryos were comparable in overall development to wild-type and heterozygous littermates, but were readily distinguished by their smaller size and marked pallor (Fig. 2). In contrast to the normal, easily visualized pattern of yolk sac vasculature in littermates (Fig. 2A), the yolk sac vessels of E11.5 FOG−/− embryos appeared very pale and thin (Fig. 2B). Upon closer inspection, the small capillaries of mutant yolk sacs appeared dilated to variable extents, presumably as a secondary consequence of anemia (see below). The fetal livers of mutant embryos were four to five times smaller than those of normal embryos and were also very pale (Fig. 2C,D), suggesting a significant reduction in effective fetal liver (definitive) erythropoiesis.
Figure 2.
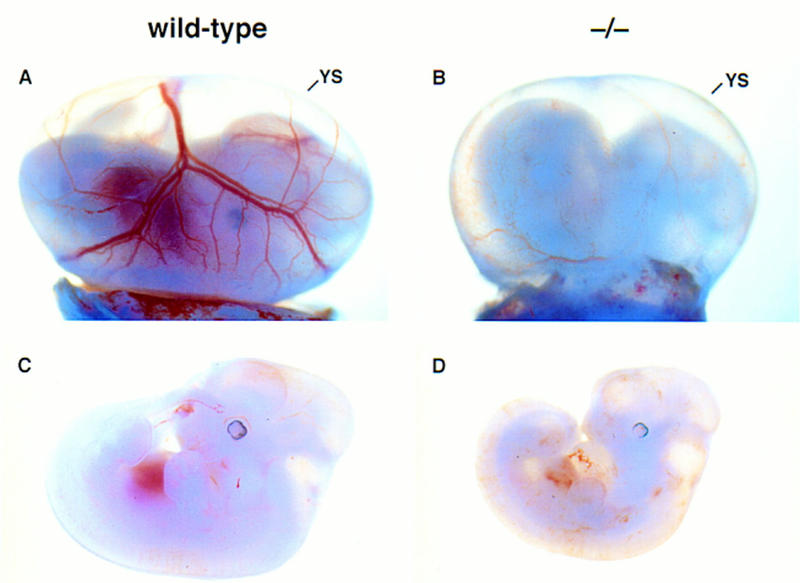
Phenotypic comparison of wild-type and FOG−/− embryos at E11.5. (A,B) Wild-type and FOG−/− E11.5 embryos with intact visceral yolk sacs (YS). (C,D) Wild-type and FOG−/− E11.5 embryos dissected free of the yolk sacs. Mutant embryos appear morphologically normal, except for the pale, slightly orange coloring of their fetal livers and circulating blood.
The phenotype of FOG−/− embryos was studied further by histological examination of E11.5 embryo sections. The fetal liver was of particular interest, as blood formation shifts from the yolk sac to the fetal liver at around E11.5–E12.5 of murine embryonic development. As shown in Figure 3A,B, the fetal livers of homozygous mutant embryos were smaller than those of wild-type embryos, yet contained dark-stained hematopoietic cells within the liver parenchyma, indicating that precursors for the definitive erythroid lineage were present at E11.5. Although smaller than wild-type, other organ systems in FOG−/− embryos also appeared to be normal. Cardiac development, for example, was not obviously impaired in mutant embryos (Fig. 3C,D). Of particular note, no differences were discernible between wild-type and mutant yolk sacs by histological analysis, and endodermal and endothelial cells lining the capillaries of mutant yolk sacs appeared histologically normal (Fig. 3E,F). In addition, the intraembryonic vasculature of mutants, including the aorta (Fig. 3G,H), intersomitic vessels (Fig. 3I,J), and branching head veins (data not shown), also appeared histologically normal.
Figure 3.
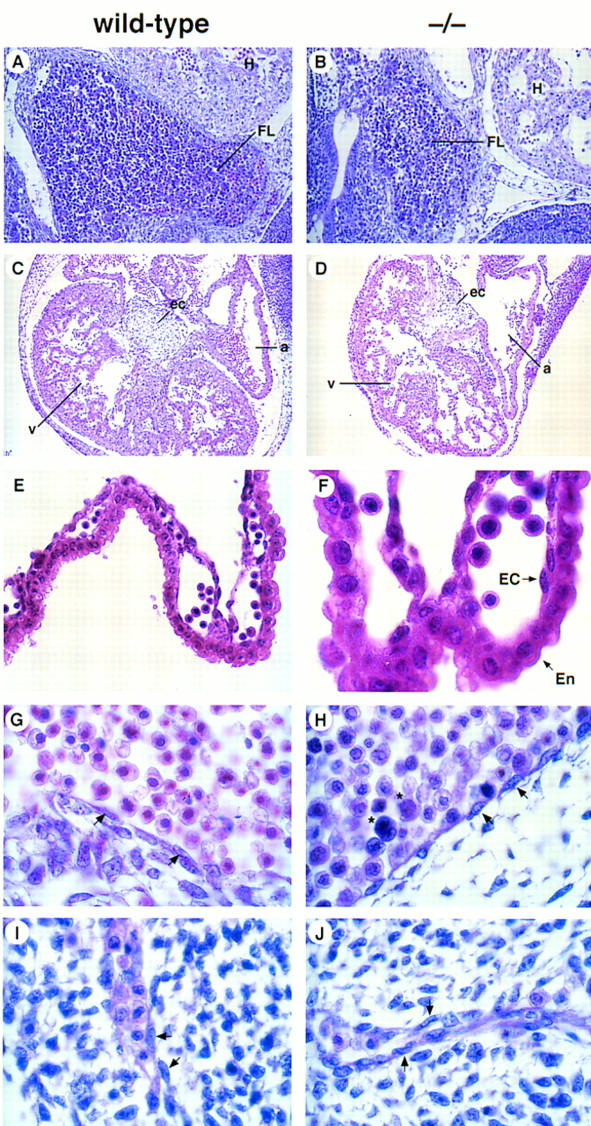
Histological analysis of wild-type and FOG−/− embryos. (A,B) Hematoxylin and eosin (H &E)-stained sagittal sections of wild-type and FOG−/− E11.5 embryos, revealing dark-stained hematopoietic elements within vessels and sinusoids of the fetal liver. Note that at this early stage of fetal liver hematopoiesis only nucleated definitive erythoblasts are observable in wild-type and mutant fetal livers. (FL) fetal liver; (H) heart. Original magnification, 200×. (C,D) H&E-stained transverse sections through the fetal hearts of wild-type and FOG−/− E11.5 embryos. Although mutant hearts are smaller than those of wild-type embryos, cardiac development does not appear to be impaired. (a) atrium; (v) ventricle; (ec) endocardial cushion. Original magnification, 100×. (E,F) H&E-stained sections of yolk sacs from wild-type and FOG−/− E10.5 embryos. No obvious defects in the vasculature of mutant yolk sacs are apparent by histological analysis. Note the presence of primitive blood cells in sections of mutant yolk sacs. Although not evident here because of H&E overstaining, these mutant cells are morphologically abnormal (see Fig. 4A). (En) extraembryonic endoderm; (EC) endothelial cell. Original magnifications, 400× and 1000×, respectively. (G–J) H&E-stained sagittal sections showing the presence of endothelial cell-lined aortas (G,H) and intersomitic vessels (I,J) in both wild-type and mutant embryos. Note the abnormal morphology of mutant blood cells (asterisks). Arrows indicate endothelial cells. Original magnification, 1000×.
Primitive hematopoiesis is defective in FOG-deficient embryos
The absence of gross morphological or histological abnormalities in FOG−/− embryos pointed to a specific failure of the primitive hematopoietic system in the absence of FOG. To address this, we first examined the peripheral blood of E10.5 and E11.5 embryos. At this stage of development, peripheral blood contains predominantly large primitive erythroid cells of yolk sac origin. These cells are nucleated and express primarily embryonic globin chains as well as small quantities of adult globins (Whitelaw et al. 1990). In contrast to the maturing primitive erythroblasts evident in normal blood (Fig. 4A, left), the primitive erythroid cells of mutant embryos appeared to be partially arrested, with many of the cells resembling proerythroblasts, the first morphologically identifiable erythroid precursors (Fig. 4A, right). In addition, a significant percentage of mutant red cells exhibited a prominent delay in nuclear versus cytoplasmic maturation (megaloblastosis) (Fig. 4A, right).
Figure 4.
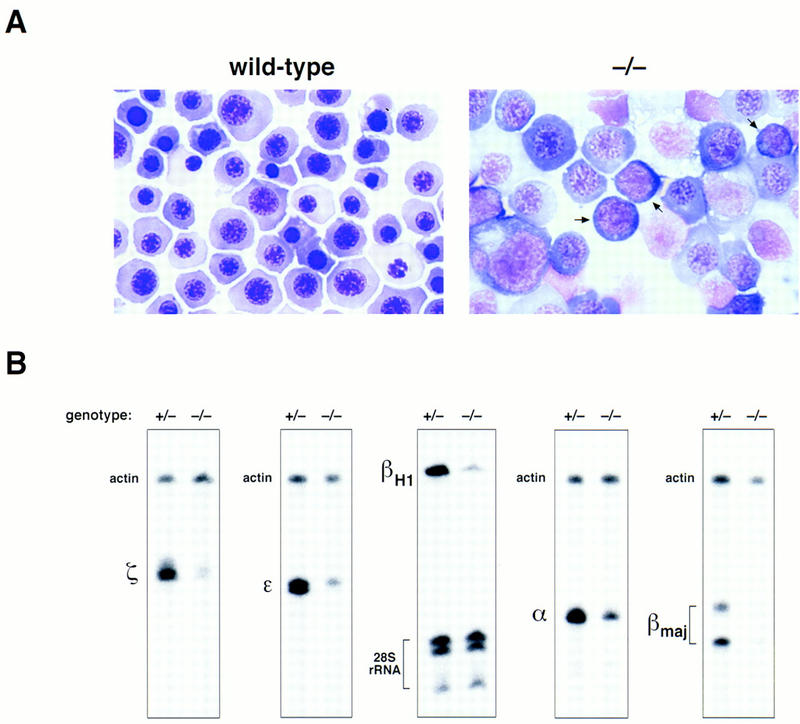
Defective primitive erythropoiesis in mice lacking FOG. (A) May–Grunwald–Giemsa staining of blood cells from wild-type and FOG−/− yolk sacs. Proerythroblast-like cells (arrows), characterized by large nuclei and basophilic cytoplasm, as well as large, megaloblastic cells are abundant in the peripheral blood of mutant embryos. Original magnification, 1000×. (B) RNase protection analysis of embryonic (ζ, ε, βH1) and adult (α, βmaj) globin transcripts in E11.5 peripheral blood from heterozygous (+/−) and homozygous mutant (−/−) embryos. To establish relative amounts of RNAs, each sample was simultaneously hybridized with an internal standard probe (mouse β-actin RNA or human 28S rRNA).
To determine the effect of this maturational defect on globin gene expression, we analyzed E11.5 peripheral blood RNA for embryonic and adult globin transcripts. As shown in Figure 4B, all embryonic globin RNAs (ζ, ε, and βH1) as well as adult globin RNAs (α and β) examined were significantly decreased (four- to fivefold) in mutant compared to heterozygous blood samples. In contrast, transcripts for actin (Fig. 4B) as well as GATA-1 (data not shown) were present at comparable levels. From these findings, we conclude that FOG is essential for the normal development of primitive erythrocytes.
Definitive erythropoiesis is severely impaired in the absence of FOG
As FOG−/− embryos die at or before the onset of the fetal liver stage of hematopoiesis, we examined a requirement for FOG in definitive erythropoiesis by in vitro hematopoietic colony-forming assays (Wong et al. 1986). Individual yolk sacs and fetal livers from mutant and control embryos were disaggregated into single cell suspensions and cultured in methylcellulose media supplemented with hematopoietic growth factors. Under assay conditions optimal for the differentiation of definitive hematopoietic progenitor cells, wild-type and heterozygous cells generated numerous, hemoglobinized colony-forming unit–erythroid (CFU-E) and burst-forming unit–erythroid (BFU-E) colonies when grown in erythropoietin (Epo) and erythropoietin plus kit ligand (Epo-KL), respectively. Staining of cytospin preparations revealed the presence of definitive erythroid cells at all stages of differentiation, including terminally differentiated, enucleated erythrocytes (Fig. 5A,C). In contrast, mutant progenitor cells yielded only pale CFU-E colonies that contained developmentally arrested proerythroblasts (Fig. 5B). Similarly, BFU-E colonies of FOG−/− origin were pale, stained weakly with benzidine, and contained morphologically aberrant erythroid precursors (Fig. 5D). In addition to large proerythroblasts, mutant colonies also contained cells with fragmented nuclei and clumped chromatin, as well as an abundance of other cell types, particularly mast and mast-like cells (Fig. 5D).
Figure 5.
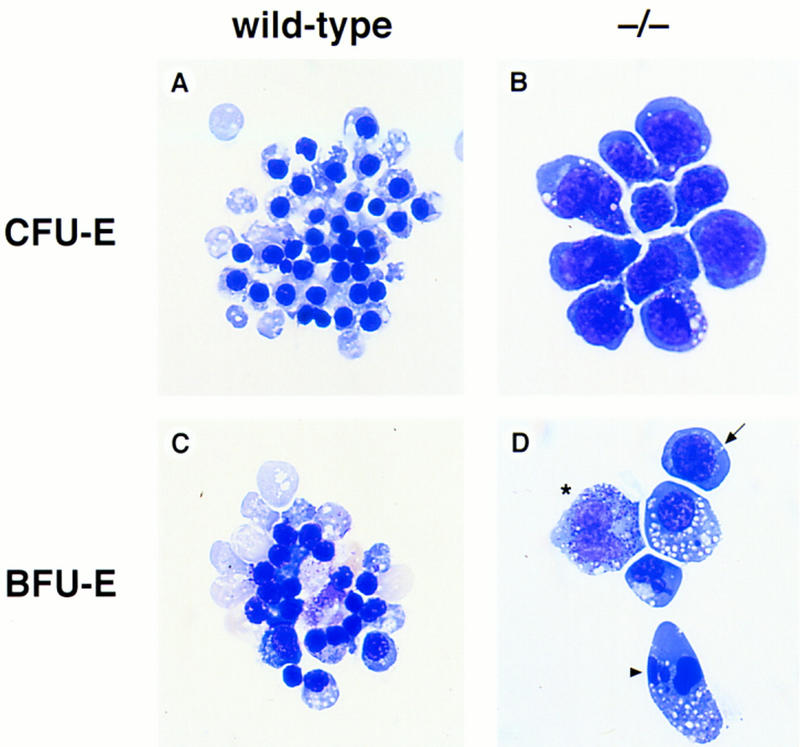
Severe impairment of in vitro erythroid differentiation in the absence of FOG. (A–D) Hematopoietic progenitor colony assays from yolk sacs and fetal livers of wild-type and mutant (−/−) embryos. (A,B) May–Grunwald–Giemsa staining of cells from wild-type and mutant CFU-Es. Colonies were grown in the presence of Epo for 2–3 days. (C,D) May–Grunwald–Giemsa staining of cells from wild-type and mutant BFU-Es. Colonies were grown in the presence of Epo-KL for 4–6 days. Mutant colonies contained proerythroblasts (arrow), cells with fragmented nuclei (arrowhead), and other morphologically abnormal cell types such as mast-like cells (asterisk).
To evaluate the consequences of loss of FOG on definitive erythrocytes in vivo, FOG−/− ES cells were generated (see Materials and Methods) and injected into wild-type host blastocysts. The animals obtained were estimated to be ∼50% chimeric based on agouti coat color. To assess contribution of FOG−/− ES cells to mature erythroid cells, we examined the hemoglobin in peripheral blood of adult chimeras. Host blastocysts of strain C57BL/6 are homozygous for the Hbbs β-globin haplotype, whereas CJ-7 ES cells of strain 129/Sv are homozygous for the Hbbd haplotype (Pevny et al. 1991). As shown in Figure 6, chimeras generated with FOG+/− ES cells showed ES cell contribution, as evidenced by the presence of Hbbd (lane 3). In contrast, chimeras made with FOG−/− ES cells showed only host-derived Hbbs (lane 2), indicating no ES cell contribution. Thus, in accordance with the in vitro findings, FOG is required for production of mature red blood cells in vivo. We propose that the embryonic lethal phenotype of FOG−/− embryos is attributable to the combined failure of both primitive and definitive erythropoiesis.
Figure 6.

Contribution of ES-derived cells to mature red blood cells of chimeras. Hemoglobin was analyzed in the peripheral blood of chimeras made with FOG+/− and FOG−/− ES cells. The Hbbs haplotype (H) is specific for host blastocysts of strain C57BL/6; Hbbd (ES) is specific for ES cells of strain 129/Sv. The figure shown is representative for four chimeras generated from two independently targeted FOG−/− ES cell clones. (C) control.
Loss of FOG leads to the specific ablation of the megakaryocytic lineage at a very early stage
Previously, we have shown that FOG is highly expressed in the megakaryocytic lineage and, furthermore, cooperates with GATA-1 during megakaryocytic cell differentiation (Tsang et al. 1997). Because FOG−/− embryos die before the appearance of detectable numbers of megakaryocytes and platelets in the fetal liver and peripheral blood, respectively, we examined the role of FOG in the megakaryocytic lineage, as well as in other cell types, by cultivating yolk sac and fetal liver cells in the presence of hematopoietic growth factors known to stimulate formation of specific nonerythroid colonies. In the presence of thrombopoietin (Tpo), wild-type and heterozygous cells generated numerous megakaryocyte colonies (Fig. 7A), defined as clusters of >20–50 large cells. As shown in Figure 7B, these colonies contained largely mature megakaryocytes, as evidenced by their characteristic morphology and intense staining for acetylcholinesterase (AChE) activity, a specific marker of mouse megakaryocytes. Under identical assay conditions, no mature megakaryocyte colonies or cells were obtained from either the yolk sacs or fetal livers of viable FOG−/− embryos (Fig. 7A). Staining for AChE activity confirmed the complete absence of mature megakaryocytes, revealing only nonspecific staining of nuclei or, rarely, small, morphologically indistinct cells with weak AChE activity (Fig. 7B). Immunofluorescence staining with rabbit anti-mouse platelet antiserum (McDonald and Jackson 1990) yielded similar results as AChE staining (data not shown). In addition, the combination of Tpo with other growth factors, such as KL, interleukin (IL)-6, and IL-11, failed to enhance the proliferation or differentiation of utant progenitors with megakaryocytic potential (data not shown).
Figure 7.
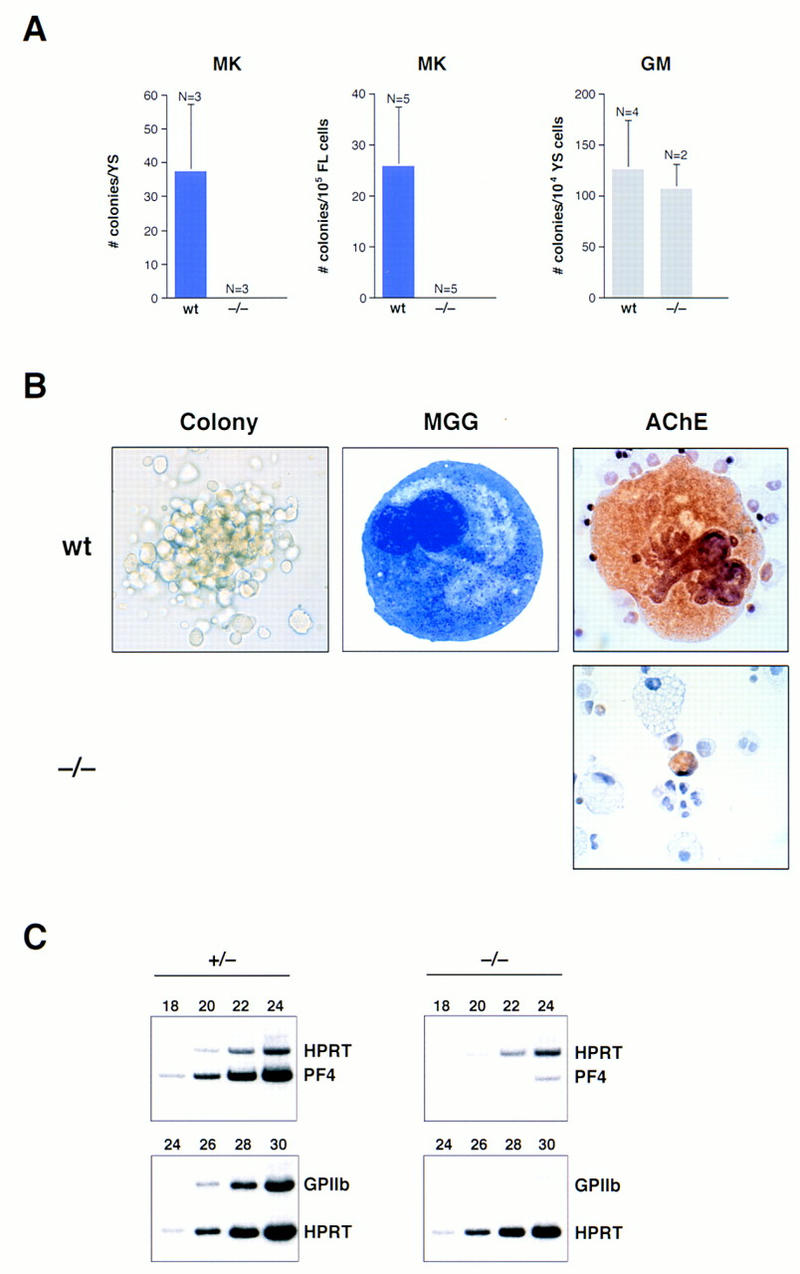
Absolute requirement for FOG in early megakaryopoiesis in vitro. Yolk sac and fetal liver cells from wild-type and homozygous mutant (−/−) embryos were assayed in semisolid medium for megakaryocytic colonies. (A) Number of megakaryocytic (MK) and granulocyte-macrophage (GM) colonies observed after 6 days of cultivation. Numbers (N) at the top of each error bar indicate the number of embryos analyzed. Error bars represent SEM. (YS) yolk sac; (FL) fetal liver. (B) May–Grunwald–Giemsa and AChE staining of cells from wild-type and mutant (−/−) colonies. Wild-type megakaryocytes were readily identified by their large size, multilobed nuclei, granular cytoplasm, and AChE positivity, which appears as orange/brown cytoplasmic staining. Small, AChE-positive cells were very rarely observed in mutant cultures. Original magnification, 630×. (C) RT–PCR analysis of PF4 and GPIIb mRNAs from hematopoietic colonies cultured in the presence of Tpo. RT–PCR amplification of HPRT transcripts served as an internal control for quantitation. Samples were amplified for six cycles in the presence of HPRT primers, before addition of PF4 primers.
To investigate further the nature of the megakaryocytic defect resulting from loss of FOG, we examined the expression of megakaryocyte-specific mRNAs by semiquantitative RT–PCR. Consistent with the morphological and cytochemical data, transcripts for platelet factor 4 (PF4), an early marker of the megakaryocytic lineage (Poncz et al. 1987), were reduced >20-fold in mutant compared to control cultures (Fig. 7C). Similarly, transcripts for glycoprotein IIb (GPIIb), another early marker of the megakaryocytic lineage (Molla et al. 1992), were virtually undetectable in mutant cultures (Fig. 7C). The absence of FOG, therefore, results in an extremely early block in megakaryocyte development.
In contrast, the development of the myeloid lineage appeared normal. As shown in Figure 7A, approximately equal numbers of granulocyte–macrophage (GM) colonies were obtained when wild-type or mutant yolk sac cells were cultivated in the presence of granulocyte–colony-stimulating factor (G-CSF), GM-CSF, IL-3, and IL-11. Furthermore, macrophages and granulocytes recovered from wild-type and mutant GM colonies were morphologically indistinguishable (Fig. 8A–D). Mast cells derived from mutant progenitor cells appeared morphologically normal and stained with toluidine blue (Fig. 8E–H). However, mast and mast-like cells, the latter characterized by multilobed nuclei, mast cell-like granules, and AChE-negative staining, appeared to be abnormally abundant, particularly in cultures grown in Epo-KL or Tpo-KL, but not KL alone (data not shown). As FOG is not expressed in mast cell lines or in primary mast cells differentiated in vitro (Tsang et al. 1997), we speculate that these mast-like cells may represent aberrant bi- or tripotential precursors that cannot proceed normally along the megakaryocytic or erythroid differentiation pathways.
Figure 8.
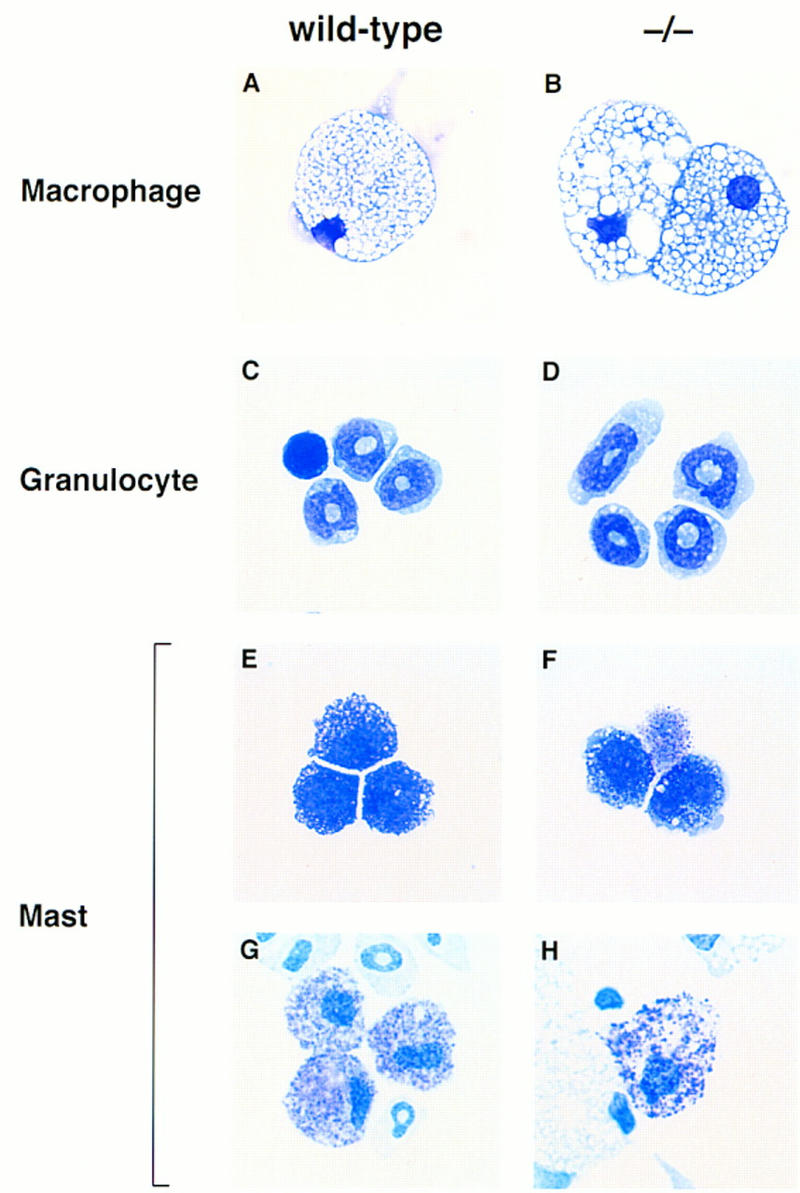
Morphological assessment of myeloid and mast cells lacking FOG. Yolk sac and fetal liver cells from wild-type and mutant (−/−) embryos were assayed in semisolid medium for granulocyte–macrophage or mast colonies. Macrophages (A,B) and granulocytes (C,D) from wild-type and mutant myeloid colonies were morphologically indistinguishable. Cells with mast cell morphology (E,F) and toluidine blue-positive granules (G,H) were also identified in wild-type and mutant cultures. Original magnifications, 630× (macrophages), 1000× (granulocytes and mast cells).
FOG−/− ES cells fail to differentiate into mature erythroid or megakaryocytic cells in vitro
To confirm the specific effects of the FOG null mutation on the erythroid and megakaryocytic lineages, we examined the hematopoietic development of wild-type and FOG−/− ES cells using a two-step in vitro differentiation assay (Keller et al. 1993). In this assay, the temporal appearance of hematopoietic precursors in developing embryoid bodies (EBs) parallels that found in the embryo. Embryoid bodies generated from two independent FOG−/− ES cell clones were disaggregated at select times and replated in the presence of various growth factors (see Materials and Methods). Both wild-type and FOG−/− EBs gave rise to primitive erythroid (EryP) precursors with replating in the presence of Epo. However, mutant EryP colonies were found at a reduced frequency and were typically pale or pink rather than red (data not shown). As shown in Figure 9B, cells within these colonies appeared to be partially arrested at a proerythroblast-like stage, closely resembling the primitive red blood cells of FOG−/− embryos (see Fig. 4A, right). Similarly, mutant EryD cells displayed a prominent defect in maturation, with many of the cells appearing proerythroblast-like or megaloblastic (Fig. 9D). Finally, in contrast to wild-type EBs, mutant EBs failed to generate any megakaryocytic colonies when replated in the presence of Tpo (Fig. 9E). In contrast, macrophage precursor-derived colonies developed from FOG−/− EBs (Fig. 9G), albeit at a slightly reduced frequency. Taken together, our in vivo and in vitro data demonstrate an intrinsic defect in FOG−/− hematopoietic progenitors that impairs selectively development of the erythroid and megakaryocytic lineages.
Figure 9.
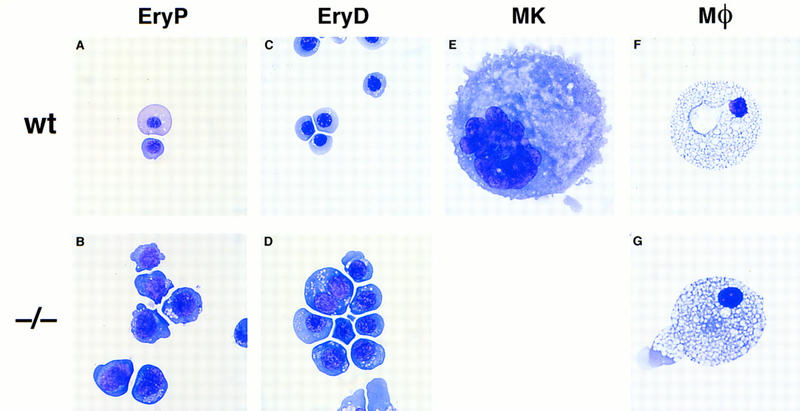
In vitro differentiation of FOG−/− ES cells. (A–G) May–Grunwald–Giemsa staining of cells from primitive erythroid (EryP) colonies (A,B), definitive erythroid (EryD) colonies (C,D), megakaryocyte colonies (E), and macrophage colonies (F,G). Mutant EryP and EryD cells display a prominent defect in maturation, as evidenced by their large size and uncondensed nuclei. No megakaryocytic cells were obtained upon replating mutant embryoid bodies in the presence of thrombopoietin. Wild-type and mutant macrophages were morphologically indistinguishable. Original magnifications, 1000× (erythroid and megakaryocytic cells), 630× (macrophages).
Discussion
Recent evidence suggests that the activities of GATA transcription factors may be modulated through interactions with complex zinc proteins such as FOG and Ush (Cubadda et al. 1997; Haenlin et al. 1997; Tsang et al. 1997). In accordance with this proposal, we have now shown that the absence of FOG results in a marked, but partial, arrest in erythroid maturation, closely resembling the GATA-1− phenotype. In contrast to GATA-1 deficiency, however, loss of FOG leads to the specific ablation of the megakaryocytic lineage, pointing to an absolute requirement for FOG during early megakaryopoiesis. FOG thus appears to act through both GATA-1-dependent as well as GATA-1-independent mechanisms. These findings have important implications for the roles of FOG and other FOG-like proteins in regulating transcription and development.
FOG, an essential cofactor for GATA-1 in erythroid cells
The consequences of FOG deficiency for erythroid development are remarkably similar to those seen in the absence of GATA-1. First, erythroid commitment does not require either FOG or GATA-1, as evidenced by the formation of precursors in their absence. Rather, FOG and GATA-1 are essential for the terminal maturation of erythroid precursors. Precursors lacking GATA-1 arrest in development at the proerythroblast stage (Weiss et al. 1994; Pevny et al. 1995) and undergo apoptosis (Weiss and Orkin 1995). Similarly, erythroid precursors lacking FOG exhibit a prominent, albeit partial, arrest also at the proerythroblast stage. FOG−/− erythroid precursors, however, appear to persist longer than GATA-1− precursors, with many acquiring megaloblastic characteristics before undergoing apoptosis (data not shown). Second, both primitive and definitive erythroid precursors arrest at a similar developmental stage in the absence of either FOG or GATA-1 (Fujiwara et al. 1996), suggesting a similar requirement for the two proteins in both erythroid lineages. This is distinct from other nuclear regulatory proteins, such as core-binding factor (CBF) (Okuda et al. 1996; Wang et al. 1996) and c-myb (Mucenski et al. 1991), whose loss results in normal yolk sac-derived erythropoiesis but severe defects in definitive erythropoiesis. Third, FOG−/− ES cells, like GATA-1− ES cells (Pevny et al. 1991), do not contribute to mature red blood cells of adult chimeras, indicating a cell autonomous defect in erythroid progenitors lacking expression of either protein. Finally, the blockage of erythroid maturation induced by loss of FOG or GATA-1 has similar consequences for the embryo; both FOG−/− and GATA-1− embryos (Fujiwara et al. 1996) die during midembryonic development with severe anemia.
This striking similarity of phenotypes strengthens the notion that FOG and GATA-1 act in concert (Fig. 10), providing a combinatorial signal for erythroid-specific differentiation. One plausible mechanism is that FOG functionally couples DNA-bound GATA-1 to the general transcriptional machinery in erythroid cells. Alternatively, FOG might serve to bridge distant DNA-bound GATA-1 molecules, thereby facilitating and stabilizing long-range interactions (e.g., local promoter-enhancer associations). Although we have been unable to demonstrate specific DNA binding by FOG itself (A. Tsang and S.H. Orkin, unpubl.), we have not excluded the possibilities that FOG recognizes DNA in association with GATA-1 or influences the DNA-binding specificity of GATA-1. By analogy to the B-cell-restricted coactivator Bob1/OBF1/OCA-B, which functions with octamer-binding transcription factors (Gstaiger et al. 1996), the complex of FOG and GATA-1 might recognize only a subset of GATA-containing sites, leading to the differential activation of critical target genes during erythroid maturation.
Figure 10.
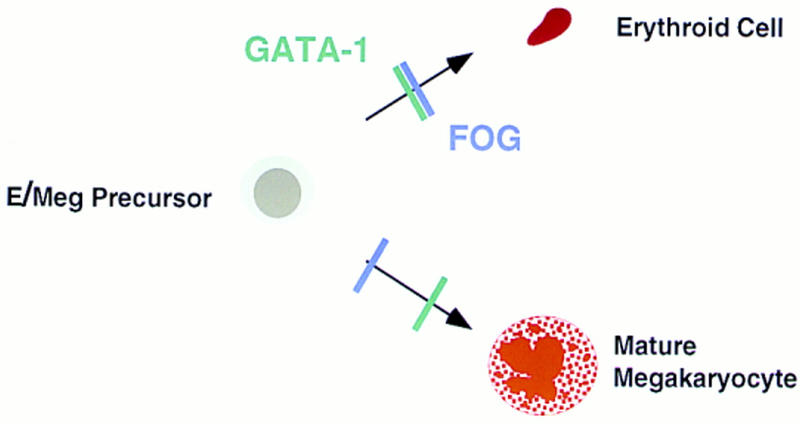
Developmental blocks imposed by loss of FOG or GATA-1. The consequences of FOG or GATA-1 deficiency for erythroid development are remarkably similar, strengthening the notion that the two proteins act in concert to drive erythroid-specific differentiation. In contrast to GATA-1 deficiency, loss of FOG leads to the specific ablation of the megakaryocytic lineage at a very early stage, pointing to a pivotal, GATA-1-independent role for FOG in early megakaryopoiesis.
Taken together with earlier findings, the presented results establish FOG and GATA-1 as components of an essential protein complex in erythroid cells. As GATA-1 interacts with other ubiquitously expressed as well as hematopoietic-restricted factors (Merika and Orkin 1995; Osada et al. 1995), this complex is likely to contain additional proteins. Of particular interest are the leukemia oncoproteins SCL/tal-1, a basic helix–loop–helix transcription factor, and Rbtn2/LMO2, a LIM-domain nuclear protein. Recent work suggests that by interacting with both SCL/tal-1 and GATA-1, Rbtn2/LMO2 assembles a large, DNA-binding complex in erythroid cells (Wadman et al. 1997). This complex presumably contains FOG as well. Interestingly, the phenotype resulting from targeted disruptions of either SCL/tal-1 or Rbtn2/LMO2, namely the complete absence of embryonic red blood cell formation (Warren et al. 1994; Shivdasani et al. 1995a), is clearly distinct from the consequences of GATA-1 or FOG loss. SCL/tal-1 is further distinguished by its absolute requirement in the formation of all adult hematopoietic lineages (Porcher et al. 1996). Thus, in contrast to FOG and GATA-1, SCL/tal-1, and presumably Rbtn2/LMO2, function at the very earliest, definable stage of hematopoietic development. Taken together, these findings predict the existence of various, highly dynamic protein complexes involving factors such as GATA-1, FOG, Rbtn2/LMO2, and SCL/tal-1 that change in abundance and composition as hematopoietic progenitors arise, commit to specific lineages, and differentiate along single pathways (Orkin 1998).
FOG and other GATA transcription factors
Previously, we have shown that FOG is capable of interacting physically with other GATA factors in addition to GATA-1 (Tsang et al. 1997). This finding, together with the observation that FOG RNA is present in multipotential progenitor cell lines and at lower levels in certain T-lymphoid lines (Tsang et al. 1997) raised the intriguing possibility that FOG might act as a cofactor for the other hematopoietic GATA factors. Our present data, however, do not support an in vivo role for FOG in either GATA-2 or GATA-3 function. GATA-2, for example, has been shown by targeted gene disruption to be critical for maintaining the proliferative capacity of immature progenitor cells and for development of the mast cell lineage (Tsai et al. 1994; Tsai and Orkin 1997). In the absence of FOG, neither multipotential progenitors nor mast cells appeared markedly deficient, hence arguing against a role for FOG in GATA-2 function. GATA-3, an essential and specific regulator of early thymocyte development (Ting et al. 1996), is also apparently not dependent on FOG for in vivo function. Preliminary analysis of the lymphopoietic potential of FOG−/− ES cells in the RAG-2-deficient blastocyst complementation assay suggests that FOG, in contradistinction to GATA-3, is not required for the development of the T cell lineage (A. Tsang, R. Monroe, F. Alt, and S.H. Orkin, unpubl.). Early embryonic lethality precludes assessment of the role of FOG in additional tissues, such as intestine and adult liver, where FOG is coexpressed with other members of the GATA factor family (Laverriere et al. 1994). Thus, the functional requirement for FOG in vivo may be more stringent than suggested by its pattern of expression vis-à-vis other GATA proteins.
A GATA-1 independent requirement for FOG in early megakaryopoiesis
Our findings establish a pivotal, GATA-1 independent role for FOG in the development of the megakaryocytic lineage. In its absence, megakaryocytes fail to develop from either hematopoietic progenitors in the embryo or from FOG−/− ES cells differentiated in vitro. Although the presence of rare, AChE-positive cells and low levels of PF4 transcripts (Fig. 7B) suggests that megakaryocytic commitment takes place in the absence of FOG, at least to a small extent, these findings also demonstrate a nonredundant function for FOG in early megakaryocyte development. This profound consequence of FOG loss was unexpected given the proposed mechanism of action of FOG as a GATA-1 cofactor in both erythroid and megakaryocytic cells (Tsang et al. 1997). Although recent studies have demonstrated a critical role for GATA-1 in the megakaryocytic lineage, GATA-1 appears to be necessary at a somewhat later stage of development (Fig. 10). Specifically, GATA-1− megakaryocytes express PF4 and GPIIb, and exhibit deregulated proliferation and severely impaired cytoplasmic maturation (Shivdasani et al. 1997). The early effect of FOG deficiency on megakaryocyte development does not preclude a role for FOG as a GATA-1 cofactor during later stages. Rather, we speculate that FOG exhibits dual functions in megakaryocytes: an early, GATA-1-independent role, perhaps at the very earliest stage of development from the bipotential erythroid/megakaryocytic progenitor, and a later, GATA-1-dependent role during terminal megakaryocyte maturation.
To date, the specific ablation of the megakaryocytic lineage has not been a phenotypic consequence of any transcription factor gene knockout. Indeed, previous studies have identified only one other transcription factor besides GATA-1, the basic leucine zipper protein NF-E2 (Andrews et al. 1993), that is specifically required for the development of megakaryocytes. Like GATA-1, however, NF-E2 is only necessary for advanced stages of megakaryocyte maturation, as endomitosis and expression of lineage-specific markers such as AChE appear to be unaffected in the absence of NF-E2 (Shivdasani et al. 1995b). The only other genes required for the formation of megakaryocytes are those that are essential for the earliest hematopoietic progenitors, for example SCL/tal-1 (Porcher et al. 1996) and CBF (Okuda et al. 1996; Wang et al. 1996), and as such are required for the development of all hematopoietic lineages. Thus, among the known hematopoietic nuclear regulatory factors, FOG is unique in defining the earliest stage in megakaryocyte development. Although the identity of this stage has not been established, we speculate that FOG may be required for the proliferation and survival of an early megakaryocyte-specific progenitor or perhaps for the differentiation of a bi- or tripotent progenitor along the megakaryocytic lineage. The latter is indirectly supported by the abundance of peculiar cell types in mutant cultures characterized by multilobed nuclei, mast cell-like granules, and red cell morphology. In the absence of other known genes required specifically for early stages in megakaryocyte development, it is likely that FOG-regulated target genes will function as novel components of developmental pathways critical for the survival, proliferation, and differentiation of the earliest megakaryocyte progenitors.
Implications of FOG for other GATA cofactors
Recent evidence suggests that a novel family of complex zinc finger proteins, exemplified by FOG and Ush, function to modulate the transcriptional activities of GATA factors in mammalian and Drosophila development, respectively (Cubadda et al. 1997; Haenlin et al. 1997; Tsang et al. 1997). Whereas FOG cooperates with GATA-1 in erythroid and megakaryocytic differentiation, Ush antagonizes the effects of the GATA factor Pannier during bristle patterning. Thus, the activities of GATA proteins may be modulated either positively or negatively through interaction with their respective cofactors. The aim of our present work was to address the validity of this paradigm. We have shown that FOG, like GATA-1, is indeed essential for terminal erythroid maturation, thereby supporting the proposed role of FOG as a GATA-1 cofactor in red blood cells. The unexpected finding of an absolute requirement for FOG during early, rather than late, megakaryocyte maturation, reveals an additional, GATA-1 independent role for FOG as an essential and specific regulator of early megakaryocyte development. Defining how FOG functions in early megakaryopoiesis will likely reveal novel GATA-independent mechanisms by which FOG and related finger proteins regulate transcription and development.
Materials and methods
All recombinant DNA work was done using standard techniques (Sambrook et al. 1989). Details of plasmid constructions are available upon request.
Targeted disruption of the murine FOG gene
Overlapping FOG genomic clones were isolated from a λFixII mouse strain 129 library (Stratagene). The exon/intron structure was determined by restriction enzyme mapping, DNA sequencing, and PCR. To generate the first targeting construct, a 2.9-kb XbaI–KpnI fragment containing FOG 5′ genomic DNA and a 2.8-kb HindIII fragment containing 3′ genomic DNA were cloned into pPNT (Tybulewicz et al. 1991). The second targeting vector was constructed by inserting a 10-kb 5′ BamHI–BstEII fragment and a 1.6-kb 3′ BglII–HindIII fragment into pTKLNCL (a gift from R. Mortensen, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA). The two targeting constructs were linearized with NotI and XhoI, respectively, and electroporated into CJ-7 ES cells. Transfectants were selected in G418 (280 μg/ml) and ganciclovir (2 μm) and expanded for Southern blot analysis. The frequency of homologous recombination with each targeting vector was 1 in 132 and 1 in 124, respectively. Two independently generated, targeted ES cell clones with normal karyotypes were injected into C57BL/6 blastocysts. Male chimeras were mated to C57BL/6 females, and heterozygous offspring were intercrossed to generate homozygous mutants. Genotyping of pups and embryos was done by Southern blot analysis. No differences in phenotype were observed between mice derived from the two independent ES cell clones, nor between mice of different genetic backgrounds (mixed 129/Sv-C57BL/6 or inbred 129/Sv).
Generation of FOG−/− ES cells
Heterozygous FOG+/− ES cell clones were passaged on gelatin-treated plates as described (Weiss et al. 1994) and grown in high concentrations (1.2–3.0 mg/ml) of G418 (Mortensen et al. 1992). Homozygous mutant ES cell clones were identified by Southern blot analysis.
Histology and cytology
E10.5 yolk sacs and E11.5 embryos were fixed in Bouin’s solution and embedded in paraffin. Sections (5 μm) were cut and stained with hematoxylin and eosin for histological examination. Cytocentrifuge preparations were stained with May–Grunwald–Giemsa or toluidine blue for morphological assessment, or for AChE activity according to standard protocol.
In vitro hematopoietic progenitor colony assay
Yolk sacs and fetal livers were dissected from E10.5 and E11.5 embryos under sterile conditions. Cells were disaggregated (Wong et al. 1986) and then plated in methylcellulose media containing 30% FCS and supplemented with one or more of the following recombinant growth factors: Epo (2 U/ml), rat KL (50 ng/ml), human Tpo (0.5 μg/ml), IL-3 (10 ng/ml), IL-6 (2 ng/ml), human IL-11 (5 ng/ml), human G-CSF (0.5 ng/ml), and GM-CSF (5 ng/ml).
In vitro ES cell hematopoietic differentiation
In vitro differentiation of ES cells was performed as described (Keller et al. 1993; Weiss et al. 1994). Primitive erythroid, EryD, and megakaryocyte (MK) colonies were generated by cultivation in Epo, Epo-KL, and Tpo, respectively. Macrophage (Mφ) colonies were obtained following growth in G-CSF, GM-CSF, IL-3, and IL-11.
Hemoglobin assay
Hemoglobin analysis was performed as described (Pevny et al. 1991).
Semiquantitative RT–PCR and RNase protection assay
Total RNA was isolated from cultured hematopoietic colonies or peripheral blood of viable E11.5 embryos by standard procedures (Chomczynski and Sacchi 1987). First strand cDNA synthesis and PCR (94°C for 1 min, 55°C for 1 min, 72°C for 1.5 min) in the presence of tracer [α-32P]dCTP were performed essentially as described (Weiss et al. 1994). Primer sets were PF4 and GPIIb (Shivdasani et al. 1995b), GATA-1 and HPRT (Weiss et al. 1994), and FOG 5′-CACCCTGTGCAGGAACCAGT-3′ and 5′-GGGTTTCTCTTCCGTCGCCG-3′ (product 246 bp). RNase protection assays were performed according to standard protocol using previously reported RNA probes for βH1 globin (Perkins et al. 1995) and α-, β-, ζ-, and ε-globins (Baron and Maniatis 1986).
Acknowledgments
We thank R. Mortensen for the targeting vector pTKLNCL, and C. Browne, K. Cunniff, and S. Goff for technical assistance. A.P.T. was supported by National Institutes of Health (NIH) training grant T32-GM07753-18. A portion of this work was supported by a grant from the NIH to S.H.O., who is an Investigator of the Howard Hughes Medical Institute.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 USC section 1734 solely to indicate this fact.
References
- Andrews NC, Erdjument-Bromage H, Davidson MB, Tempst P, Orkin SH. Erythroid transcription factor NF-E2 is a haematopoietic-specific basic-leucine zipper protein. Nature. 1993;362:722–728. doi: 10.1038/362722a0. [DOI] [PubMed] [Google Scholar]
- Baron MH, Maniatis T. Rapid reprogramming of globin gene expression in transient heterokaryons. Cell. 1986;46:591–602. doi: 10.1016/0092-8674(86)90885-8. [DOI] [PubMed] [Google Scholar]
- Blobel GA, Simon MC, Orkin SH. Rescue of GATA-1-deficient embryonic stem cells by heterologous GATA-binding proteins. Mol Cell Biol. 1995;15:626–633. doi: 10.1128/mcb.15.2.626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- Cubadda Y, Heitzler P, Ray RP, Bourouis M, Ramain P, Gelbart W, Simpson P, Haenlin M. u-shaped encodes a zinc finger protein that regulates the proneural genes achaete and scute during the formation of bristles in Drosophila. Genes & Dev. 1997;11:3083–3095. doi: 10.1101/gad.11.22.3083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans T, Felsenfeld G. The erythroid-specific transcription factor eryf1: A new finger protein. Cell. 1989;58:877–885. doi: 10.1016/0092-8674(89)90940-9. [DOI] [PubMed] [Google Scholar]
- Fujiwara Y, Browne CP, Cunniff K, Goff SC, Orkin SH. Arrested development of embryonic red cell precursors in mouse embryos lacking transcription factor GATA-1. Proc Natl Acad Sci. 1996;93:12355–12358. doi: 10.1073/pnas.93.22.12355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gstaiger M, Georgiev O, van Leeuwen H, van der Vliet P, Schaffner W. The B cell coactivator Bob1 shows DNA sequence-dependent complex formation with Oct-1/Oct-2 factors, leading to differential promoter activation. EMBO J. 1996;15:2781–2790. [PMC free article] [PubMed] [Google Scholar]
- Haenlin M, Cubadda Y, Blondeau F, Heitzler P, Lutz Y, Simpson P, Ramain P. Transcriptional activity of Pannier is regulated negatively by heterodimerization of the GATA DNA-binding domain with a cofactor encoded by the u-shaped gene of Drosophila. Genes & Dev. 1997;11:3096–3108. doi: 10.1101/gad.11.22.3096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller G, Kennedy M, Papayannopoulou T, Wiles MV. Hematopoietic commitment during embryonic stem cell differentiation in culture. Mol Cell Biol. 1993;13:472–486. doi: 10.1128/mcb.13.1.473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kudla B, Caddick MX, Langdon T, Martinez-Rossi NM, Bennett CF, Sibley S, Davies RW, Arst HN., Jr The regulatory gene areA mediating nitrogen metabolite repression in Aspergillus nidulans. Mutations affecting specificity of gene activation alter a loop residue of a putative zinc finger. EMBO J. 1990;9:1355–1364. doi: 10.1002/j.1460-2075.1990.tb08250.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kulessa H, Frampton J, Graf T. GATA-1 reprograms avian myelomonocytic cells into eosinophils, thromboblasts and erythroblasts. Genes & Dev. 1995;9:1250–1262. doi: 10.1101/gad.9.10.1250. [DOI] [PubMed] [Google Scholar]
- Kuo CT, Morrisey EE, Anandappa R, Sigrist K, Lu MM, Parmacek MS, Soudais C, Leiden JM. The GATA-4 transcription factor is required for ventral morphogenesis and heart tube formation. Genes & Dev. 1997;11:1048–1060. doi: 10.1101/gad.11.8.1048. [DOI] [PubMed] [Google Scholar]
- Laverriere AC, MacNeill C, Mueller C, Poelmann RE, Burch JB, Evans T. GATA-4/5/6, a subfamily of three transcription factors transcribed in developing heart and gut. J Biol Chem. 1994;269:23177–23184. [PubMed] [Google Scholar]
- Martin DIK, Orkin SH. Transcriptional activation and DNA-binding by the erythroid factor GF-1/NF-E1/Eryf 1. Genes & Dev. 1990;4:1886–1898. doi: 10.1101/gad.4.11.1886. [DOI] [PubMed] [Google Scholar]
- McDonald TP, Jackson CW. Thrombopoietin derived from human embryonic kidney cells stimulates an increase in DNA content of murine megakaryocytes in vivo. Exper Hematol. 1990;18:758–763. [PubMed] [Google Scholar]
- Merika M, Orkin SH. Functional synergy and physical interactions of the erythroid transcription factor GATA-1 and Krüppel family proteins, Sp1 and EKLF. Mol Cell Biol. 1995;15:2437–2447. doi: 10.1128/mcb.15.5.2437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Molkentin JD, Lin Q, Duncan SA, Olson EN. Requirement of the transcription factor GATA-4 for heart tube formation and ventral morphogenesis. Genes & Dev. 1997;11:1061–1072. doi: 10.1101/gad.11.8.1061. [DOI] [PubMed] [Google Scholar]
- Molla A, Andrieux A, Chapel A, Schweitzer A, Berthier R, Marguerie G. Lack of transcription and expression of the aIIb integrin in human early haematopoietic stem cells. Br J Haematol. 1992;82:635–639. doi: 10.1111/j.1365-2141.1992.tb06937.x. [DOI] [PubMed] [Google Scholar]
- Mortensen RM, Conner DA, Chao S, Geisterfer-Lowrance AAT, Seidman JG. Production of homozygous mutant ES cells with a single targeting construct. Mol Cell Biol. 1992;12:2391–2395. doi: 10.1128/mcb.12.5.2391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mucenski ML, McLain K, Kier AB, Swerdlow SH, Schreiner CM, Miller TA, Pietryga DW, Scott WJ, Jr, Potter SS. A functional c-myb gene is required for normal fetal hepatic hematopoiesis. Cell. 1991;65:677–689. doi: 10.1016/0092-8674(91)90099-k. [DOI] [PubMed] [Google Scholar]
- Okuda T, van Deursen J, Hiebert SW, Grosveld G, Downing JR. AML1, the target of multiple chromosomal translocations in human leukemia, is essential for normal fetal liver hematopoiesis. Cell. 1996;84:321–330. doi: 10.1016/s0092-8674(00)80986-1. [DOI] [PubMed] [Google Scholar]
- Orkin SH. GATA-binding transcription factors in hematopoietic cells. Blood. 1992;80:575–581. [PubMed] [Google Scholar]
- ————— . Transcription factors regulating early hematopoietic development and lineage commitment. In: Monroe JG, Rothenberg EV, editors. Molecular biology of B-cell and T-cell development. Totowa, NJ: Humana Press; 1998. . (In Press.) [Google Scholar]
- Osada H, Grutz G, Axelson H, Forster A, Rabbitts TH. Association of erythroid transcription factors: Complexes involving the LIM protein RBTN2 and the zinc-finger protein GATA1. Proc Natl Acad Sci. 1995;92:9585–9589. doi: 10.1073/pnas.92.21.9585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkins AC, Sharpe AH, Orkin SH. Lethal β-thalassemia in mice lacking the erythroid CACCC-transcription factor EKLF. Nature. 1995;375:318–322. doi: 10.1038/375318a0. [DOI] [PubMed] [Google Scholar]
- Pevny L, Simon MC, Robertson E, Klein WH, Tsai SF, D’Agati V, Orkin SH, Costantini F. Erythroid differentiation in chimaeric mice blocked by a targeted mutation in the gene for transcription factor GATA-1. Nature. 1991;349:257–260. doi: 10.1038/349257a0. [DOI] [PubMed] [Google Scholar]
- Pevny L, Lin CS, D’Agati V, Simon MC, Orkin SH, Costantini F. Development of hematopoietic cells lacking transcription factor GATA-1. Development. 1995;121:163–172. doi: 10.1242/dev.121.1.163. [DOI] [PubMed] [Google Scholar]
- Poncz M, Surrey S, LaRocco P, Weiss MJ, Rappaport EF, Conway TM, Schwartz E. Cloning and characterization of platelet factor 4 cDNA derived from a human erythroleukemia cell line. Blood. 1987;69:219–223. [PubMed] [Google Scholar]
- Porcher C, Swat W, Rockwell K, Fujiwara Y, Alt FW, Orkin SH. The T cell leukemia oncoprotein SCL/tal-1 is essential for development of all hematopoietic lineages. Cell. 1996;86:47–57. doi: 10.1016/s0092-8674(00)80076-8. [DOI] [PubMed] [Google Scholar]
- Ramain P, Heitzler P, Haenlin M, Simpson P. Pannier, a negative regulator of achaete and scute in Drosophila, encodes, a zinc finger protein with homology to the vertebrate transcription factor GATA-1. Development. 1993;119:1277–1291. doi: 10.1242/dev.119.4.1277. [DOI] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: A laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Shivdasani RA. Transcription factors in megakaryocyte differentiation and gene expression. In: Kuter DJ, Hunt P, Sheridan W, Zucker-Franklin D, editors. Thrombopoiesis and thrombopoietins. Totawa, NJ: Humana Press; 1997. pp. 189–202. [Google Scholar]
- Shivdasani RA, Mayer EL, Orkin SH. Absence of blood formation in mice lacking the T-cell leukaemia oncoprotein tal-1/SCL. Nature. 1995a;373:432–434. doi: 10.1038/373432a0. [DOI] [PubMed] [Google Scholar]
- Shivdasani RA, Rosenblatt MF, Zucker-Franklin D, Jackson CW, Hunt P, Saris CJ, Orkin SH. Transcription factor NF-E2 is required for platelet formation independent of the actions of thrombopoietin/MGDF in megakaryocyte development. Cell. 1995b;81:695–704. doi: 10.1016/0092-8674(95)90531-6. [DOI] [PubMed] [Google Scholar]
- Shivdasani RA, Fujiwara Y, McDevitt MA, Orkin SH. A lineage-selective knockout establishes the critical role of transcription factor GATA-1 in megakaryocyte growth and platelet development. EMBO J. 1997;16:3965–3973. doi: 10.1093/emboj/16.13.3965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spieth J, Shim YH, Lea K, Conrad R, Blumenthal T. elt-1, an embryonically expressed Caenorhabditis elegans gene homologous to the GATA transcription factor family. Mol Cell Biol. 1991;9:4651–4659. doi: 10.1128/mcb.11.9.4651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ting CN, Olson MC, Barton KP, Leiden JM. Transcription factor GATA-3 is required for development of the T-cell lineage. Nature. 1996;384:474–478. doi: 10.1038/384474a0. [DOI] [PubMed] [Google Scholar]
- Tsai FY, Orkin SH. Transcription factor GATA-2 is required for proliferation/survival of early hematopoietic cells and mast cell formation, but not for erythroid and myeloid terminal differentiation. Blood. 1997;89:3636–3643. [PubMed] [Google Scholar]
- Tsai SF, Martin DI, Zon LI, D’Andrea AD, Wong GG, Orkin SH. Cloning of cDNA for the major DNA-binding protein of the erythroid lineage through expression in mammalian cells. Nature. 1989;339:446–451. doi: 10.1038/339446a0. [DOI] [PubMed] [Google Scholar]
- Tsai FY, Keller G, Kuo FC, Weiss M, Chen J, Rosenblatt M, Alt FW, Orkin SH. An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature. 1994;371:221–226. doi: 10.1038/371221a0. [DOI] [PubMed] [Google Scholar]
- Tsang AP, Visvader JE, Turner CA, Fujiwara Y, Yu C, Weiss MJ, Crossley M, Orkin SH. FOG, a multitype zinc finger protein, acts as a cofactor for transcription factor GATA-1 in erythroid and megakaryocytic differentiation. Cell. 1997;90:109–119. doi: 10.1016/s0092-8674(00)80318-9. [DOI] [PubMed] [Google Scholar]
- Tybulewicz VLJ, Crawford CE, Jackson PK, Bronson RT, Mulligan RC. Neonatal lethality and lymphopenia in mice with a homozygous disruption of the c-abl proto-oncogene. Cell. 1991;65:1153–1163. doi: 10.1016/0092-8674(91)90011-m. [DOI] [PubMed] [Google Scholar]
- Visvader JE, Elefanty AG, Strasser A, Adams JM. GATA-1 but not SCL induces megakaryocytic differentiation in an early myeloid line. EMBO J. 1992;11:4557–4564. doi: 10.1002/j.1460-2075.1992.tb05557.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Visvader JE, Crossley M, Hill J, Orkin SH, Adams JM. The C-terminal zinc finger of GATA-1 or GATA-2 is sufficient to induce megakaryocytic differentiation of an early myeloid cell line. Mol Cell Biol. 1995;15:634–641. doi: 10.1128/mcb.15.2.634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wadman IA, Osada H, Grutz GG, Agulnick AD, Westphal H, Forster A, Rabbitts TH. The LIM-only protein Lmo2 is a bridging molecule assembling an erythroid, DNA-binding complex which includes the TAL1, E47, GATA-1 and Ldb1/NLI proteins. EMBO J. 1997;16:3145–3157. doi: 10.1093/emboj/16.11.3145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Q, Stacy T, Miller JD, Lewis AF, Gu T-L, Huang X, Bushweller JH, Bories J-C, Alt FW, Ryan G, Liu PP, Wynshaw-Boris A, Binder M, Marin-Padilla M, Sharpe AH, Speck NA. The CBFβ subunit is essential for CBFα2(AML1) function in vivo. Cell. 1996;87:697–708. doi: 10.1016/s0092-8674(00)81389-6. [DOI] [PubMed] [Google Scholar]
- Warren AJ, Colledge WH, Carlton MBL, Evans MJ, Smith AJH, Rabbitts TH. The oncogenic cysteine-rich LIM domain protein Rbtn2 is essential for erythroid development. Cell. 1994;78:45–57. doi: 10.1016/0092-8674(94)90571-1. [DOI] [PubMed] [Google Scholar]
- Weiss MJ, Orkin SH. Transcription factor GATA-1 permits survival and maturation of erythroid precursors by preventing apoptosis. Proc Natl Acad Sci. 1995;92:9623–9627. doi: 10.1073/pnas.92.21.9623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiss MJ, Keller G, Orkin SH. Novel insights into erythroid development revealed through in vitro differentiation of GATA-1 embryonic stem cells. Genes & Dev. 1994;8:1184–1197. doi: 10.1101/gad.8.10.1184. [DOI] [PubMed] [Google Scholar]
- Weiss MJ, Yu C, Orkin SH. Erythroid-cell-specific properties of transcription factor GATA-1 revealed by phenotypic rescue of a gene-targeted cell line. Mol Cell Biol. 1997;17:1642–1651. doi: 10.1128/mcb.17.3.1642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitelaw E, Tsai SF, Hogben P, Orkin SH. Regulated expression of globin chains and the erythroid transcription factor GATA-1 during erythropoiesis in the developing mouse. Mol Cell Biol. 1990;10:6596–6606. doi: 10.1128/mcb.10.12.6596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong PMC, Chung SW, Chui DHK, Eaves CJ. Properties of the earliest clonogenic hemopoietic procursors to appear in the developing murine yolk sac. Proc Natl Acad Sci. 1986;83:3851–3854. doi: 10.1073/pnas.83.11.3851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng W, Flavell RA. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell. 1997;89:587–596. doi: 10.1016/s0092-8674(00)80240-8. [DOI] [PubMed] [Google Scholar]
- Zhu J, Hill RJ, Heid PJ, Fukuyama M, Sugimoto A, Priess JR, Rothman JH. end-1 encodes an apparent GATA factor that specifies the endoderm precursor in Caenorhabditis elegans embryos. Genes & Dev. 1997;11:2883–2896. doi: 10.1101/gad.11.21.2883. [DOI] [PMC free article] [PubMed] [Google Scholar]