

Abstract
The shewanellae are ubiquitous in aquatic and sedimentary systems that are chemically stratified on a permanent or seasonal basis. In addition to their ability to utilize a diverse array of terminal electron acceptors, the microorganisms have evolved both common and unique responding mechanisms to cope with various stresses. This paper focuses on the response and adaptive mechanism of the shewanellae, largely based on transcriptional data.
1. Introduction
Stress is an inevitable part of the life of all organisms. This is especially true about microorganisms, which reside and thrive in almost all environments on earth, including some considered extremely harsh [1]. Common environmental factors that affect the activities of microorganisms include temperature, pH, water availability, nutrient limitation, presence of various chemicals, osmolarity, pressure, and radiation [2]. Consequently, for every microorganism the ability to adapt rapidly to changes in environments is essential for its survival and prosperity. Regulation that modulates the microbial adaptation to environmental disturbances is rather complex. The most important and efficient control occurs at the level of transcription. Many single stress-induced regulatory circuits have been identified, which enable cells to cope with specific stresses. However, given that microbial cells live in a dynamic environment where multiple factors fluctuate constantly at the same time, stress responses are generally carried out by a regulatory network composed of a series of individual circuits which are highly connected [3].
Most of our understanding of microbial stress response mechanisms has come from the study of model microorganisms, particularly Escherichia coli and Bacillus subtilis. Extensive physiological and genetic analyses of the stress response systems in these two bacteria have helped us to elucidate the complexity of the process, function of critical proteins, and regulation [4]. While model organisms will continue to provide insights into the fundamental properties of the stress response systems, efforts should be extended to other microorganisms, especially those that are of scientific, environmental, and economic importance.
As one of representatives, the family of Shewanellaceae (order Alteromonadales, class γ-proteobacteria) is emerging in recent years. The genus Shewanella consists of rod-shaped, Gram-negative, aerobic or facultatively anaerobic, polarly flagellated, readily cultivated γ-proteobacteria [5–8]. While many Shewanella isolates remain uncharacterized, 52 species have been recognized to date [9]. Shewanellae are renowned for its ability to use a diverse range of electron acceptors for anaerobic respiration, including fumarate, nitrate, nitrite, thiosulfate, elemental sulfur, trimethylamine N-oxide (TMAO), dimethyl sulfoxide (DMSO), Fe(III), Mn(III) and (IV), Cr(VI), U(VI), As(V), V(V), and others [10, 11]. As a result of this property, Shewanellae have drawn much attention in the fields of bioremediation, biogeochemical circulation of minerals, and bioelectricity [12, 13]. In addition, Shewanellae have now served as the model for ecological and evolutionary studies at the whole genome level because of its diverse habitats and the availability of up to 26 genome sequences [14, 15].
A number of Shewanella strains are currently under physiological investigation [11]. However, stress responses have focused nearly exclusively on Shewanella oneidensis, the first genome of the shewanellae to be sequenced [16]. The availability of the genome sequence allowed development of high-throughput technologies such as microarrays and proteomics tools, with which an array of assays has been carried out to decipher the ability of S. oneidensis to respond to and survive external stresses. While impacts of most of common environmental factors have been examined, oxidative stress imposed by H2O2 is surprisingly untouched. In this paper, we consider all insights into the stress response mechanisms revealed thus far in S. oneidensis and broaden our discussion to other sequenced species if necessary.
2. Stress Responses to Temperature Fluctuation
Variation in growth temperature is a common stress encountered in nature. Stress response to sudden fluctuation in growth temperature, has become a model system for studying the impact of environmental stresses on biological systems. The hallmark of this adaptive cellular response is the induction of a limited set of proteins, called Heat shock proteins (Hsps) or Cold shock proteins (Csps). In general, Hsps play important roles in protein folding, degradation, assembly of protein complexes, and transport of proteins across membranes whereas Csps function as RNA chaperons to regulate ribosomal translation, rate of mRNA degradation and termination of transcription [17–19].
Using whole-genome DNA microarrays, temporal gene expression profiles of S. oneidensis MR-1 in response to temperature variations have been investigated [20, 21]. Expression profiles indicate that temperature fluctuation has a pleiotropic effect on the bacterial transcriptomes. Both heat and cold shock responses appear to share a couple of common features, including that approximately 15% of the total genes are significantly affected (P < 0.05) over a 25-min period, that the global changes in mRNAs are rapid and transient, and that a similar set of proteins are induced to manage energy production and protein damage. For instance, most of genes encoding enzymes in the Entner-Doudoroff pathway and the pentose cycle are highly induced upon a temperature alteration.
In the case of heat shock response, two lines of evidence suggest that S. oneidensis copes with the situation with mechanism similar to that employed by E. coli. First, the majority of the genes that showed homology to known Hsps in E. coli such as DnaK, DnaJ, GroEL, GroES, GrpE, HtpG, and Lon/La proteases were highly induced. Second, the identified σ 32 consensus sequences (CTTGAAA-13/15bp-CCCCAT) of both bacteria for heat shock gene promoters are virtually the same (Figure 1), indicating that the induction of most Hsps owns to a rapid and transient increase in the intracellular concentration of an alternative σ factor, σ 32 encoded by rpoH. Nevertheless, novel findings are not scarce. After numerous attempts, we failed to remove rpoH from the genome, implicating that σ 32 is essential in S. oneidensis (unpublished result). Additionally, some hypothetical proteins (i.e., SO2017) are under the control of σ 32, suggesting that S. oneidensis recruits new proteins to overcome increased temperature (Table 1).
Figure 1.
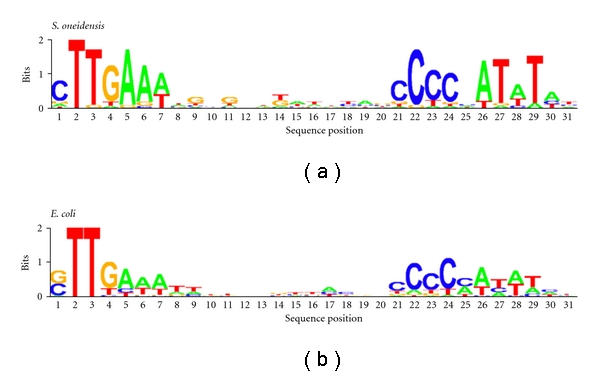
Comparison of consensus σ 32-recognition sequences of E. coli and S. oneidensis. The E. coli and S. oneidensis sequences used were from the published reports by Nonaka et al. [22] and Gao et al. [20], respectively. The sequences were initially aligned by clustalx and the sequence logo was prepared using public software at http://www.bioinf.ebc.ee/EP/EP/SEQLOGO/.
Table 1.
Genes predicted to be under the direct control of σ 32 in S. oneidensis.
| Locus | Gene | Product | Start | End | Sequence | Weight |
|---|---|---|---|---|---|---|
| SO2016 | htpG | heat shock protein HtpG | −84 | −55 | CTTGAAAAGTGGATTTGCAGCCCCATTTTA | 20.3 |
| SO4162 | hslV | ATP-dependent protease HslV | −83 | −54 | CTTGAATTCTGGCTATCCATCCCCATATTT | 20.1 |
| SO1126 | dnaK | chaperone protein DnaK | −78 | −48 | CTTGAAAAAAAATGCGTCCGGCCCCATATCT | 18.6 |
| SO0406 | trxA | thioredoxin 1 | −80 | −51 | CTTGAAAAGCTATTTTTCAGCCCCAATATA | 18.4 |
| SO1524 | grpE | heat shock protein GrpE | −74 | −45 | CTTGAAACGTCAAAATTGATCCCCATAATA | 18.2 |
| SO2593 | conserved hypothetical protein | −262 | −232 | CTTGAAATGGGGAGTTTAACTCCCCATTTTT | 17.9 | |
| SO3577 | clpB | clpB protein | −77 | −48 | CTTGAATTTGGTTAAATAGCCCCCATCTTT | 16.8 |
| SO0452 | trxC | thioredoxin 2 | −60 | −31 | CTTTAAATTCGCCGCAGCGCCCCCATATCT | 15.7 |
| SO2017 | conserved hypothetical protein | −106 | −76 | CTTGAGTTGAGACGCAAGTGCCCCGATTTAC | 14.4 | |
| SO1796 | lon | ATP-dependent protease La | −68 | −39 | ATTGAAAGGGCATAAACCGCCCCAATATAC | 14 |
| SO2277 | ibpA | 16 kDa heat shock protein A | −167 | −138 | CTTGAAATCCGTTTTCCTATCCTTATATCT | 13.5 |
| SO0703 | groES | chaperonin GroES | −123 | −93 | CTTGGATCTGGCGGGGGTGAACCCCATATCA | 13.3 |
| SO4492 | conserved hypothetical protein | −76 | −48 | GTTGAAAAGAATTGATTTGCCCCAAGATA | 12.8 | |
| SO1794 | clpP | ATP-dependent Clp protease, proteolytic subunit | −83 | −55 | CTTGACTTGATTAGCAGTTCGCCATTTAT | 12.8 |
| SO1163 | conserved hypothetical protein | −60 | −31 | CTTGAATCGGGTATAATCGCCACCATATAG | 12.7 | |
| SO3863 | modA | molybdenum ABC transporter, periplasmic molybdenum-binding protein | −206 | −177 | CTTGAGTAAATGTTATTGTCCCCGATCAAT | 12.3 |
| SO1196 | rrmJ | ribosomal RNA large subunit methyltransferase J | −65 | −36 | GTTGAAAAACCGCTATTCTACCCTTATATA | 12.2 |
| SO2723 | HIT family protein | −47 | −17 | ATTGAATTGCTAGTATACTATCCCAATTAAC | 11.8 | |
| SO1213 | hydrolase, TatD family | −240 | −211 | GTTTAAAGGCGGTGATTCACCGCCTTTTTT | 11.8 | |
| SO2705 | topA | DNA topoisomerase I | −77 | −49 | CTTGAAACTCTCAGTGCAACCCTCTATAT | 11.1 |
| SO3501 | conserved hypothetical protein | −297 | −268 | CATGAATTTGGCAACGGCACCGCCATTTTC | 11 | |
| SO2728 | htpX | peptidase HtpX | −101 | −71 | GTAGAAAAACTCTTATCTTTACCCCTTGAAT | 10.6 |
| SO1473 | smpB | SsrA-binding protein | −69 | −39 | GTTGAAATAGCTCAAATAAACCCTTATATCC | 10.3 |
| SO0698 | fsxA | fxsA protein | −64 | −34 | CTTGAATTAAGACCGGATTGCCCCCATTTAG | 10.3 |
| SO3402 | hypothetical protein | −396 | −367 | ATTGAAAAGGGCCTTTATGGCCCTTTTTCG | 10.2 | |
| SO1937 | fur | ferric uptake regulation protein | −164 | −135 | CTTGAATTGCCGCAATTTATTGCAATTTCA | 10.2 |
| SO2706 | astB | succinylarginine dihydrolase | −40 | −11 | TTTGAATAAATAATAACCTTCCCTATCACA | 9.7 |
| SO0868 | hypothetical protein | −93 | −63 | GTTTAAATGGGGAGAAAACAACTCCATTTTA | 9.4 | |
| SO3961 | rpoN | RNA polymerase sigma-54 factor | −83 | −53 | CTTGAATTTGGCAGCGCAAAGCGCCATCAGT | 9.4 |
| SO0930 | tkt | Transketolase | −161 | −133 | CTTGAATAGTTCATCCTTAAGCCATTTTT | 9.3 |
| SO3528 | hypothetical protein | −195 | −167 | AATGAAAAGAGGCTTTTAGCCTCTTTTTT | 9.3 | |
| SO1580 | TonB-dependent heme receptor | −57 | −28 | CTTTGATGCCTATAATGCCGCCCTATTTTT | 9.3 | |
| SO2314 | ISSo1, transposase OrfA | −227 | −197 | GTTAAAATGACAAGCATGGAGCGCAATATCT | 9.2 | |
| SO1903 | hypothetical protein | −71 | −42 | TTTGGGATTATTTAATTCCCCCCCATTTAT | 9.2 | |
| SO1097 | conserved hypothetical protein | −63 | −33 | CATGAAATCTGCGATAATCAGCGCCTTATTT | 9.2 | |
| SO0595 | hypothetical protein | −327 | −298 | CTTGATTAGAGCCACGTCGCTCCAATTTTT | 9.2 | |
| SO4719 | conserved hypothetical protein | −44 | −16 | CTAGGCATTTGAGTTGGAACCCTATTTTT | 9.1 | |
| SO4287 | motA | chemotaxis motA protein | −127 | −99 | CTTGAATTTAGTAGATTTTCCTTATAATG | 9.1 |
| SO3113 | tgt | queuine tRNA-ribosyltransferase | −96 | −67 | GTTGAACCTTTTAGATCTGTCCCTATCTCT | 9 |
Genome screening with σ 32 weight matrix is performed using RSAT at http://rsat.ulb.ac.be/rsat/RSAT_home.cgi [23]. Genes with a weight score over 9 are shown.
Unlike E. coli, most Shewanella strains are psychrotolerant. In terms of the canonical Csps S. oneidensis possesses three (of which two (SO1648 and SO2787) are cold inducible) whereas E. coli has nine (of which four are cold inducible) [19]. Both SO1648 and SO2787 are important in growth at low temperatures evidenced in the mutational analysis [21]. The S. oneidensis genome carries two more genes encoding Csd(cold shock domain)-containing proteins (SO0733, 203 aa; SO1732, 224 aa) whose C-terminal lacks sequence similarity to any known proteins. Intriguingly, such a structure has been found only in eukaryotes, with the exception of Mycobacterium [24]. Neither SO0733 nor SO1732 is found to be induced upon a decrease in temperature or influences growth at low temperature, indicating that these Csd-containing proteins may not be involved in cold stress response.
S. piezotolerans WP3 is another Shewanella that has been studied in respect of response to low temperatures. Strikingly, none of its Csps are cold inducible, suggesting that these proteins may not play an indispensible role in the process [25]. Instead, the organism utilizes other strategies to overcome temperature downshifts. These include increased production of EPA (eicosapentaenoic acid) and BCFA (branched-chain fatty acid) [26], induced expression of RNA helicase DeaD which may facilitate transcription, morphological changes in cell membrane, and elevated assembly of lateral flagella (The organism possesses both polar and lateral flagella.) [25]. In addition, a novel filamentous phage (SW1) is found to be significantly induced at low temperature but the significance of this event in the cold adaptation of S. piezotolerans WP3 is unknown [27].
3. Stress Responses to Acidic and Alkaline pH
Microorganisms live in a volatile environment where extracellular pH changes frequently. To minimize the acid- or alkaline-induced damage, various adaptive strategies have evolved [28, 29]. Studies on E. coli have revealed that bacterial cells activate outward H+ pumps such as K+/proton antiporters in response to acute cytoplasmic acidification and sodium proton antiporters, which bring in 2 H+ for each Na+ extruded, to adapt to alkaline pH in the presence of Na+. To survive upon prolonged acid stress exposure, cells rely on the arginine and glutamate decarboxylase/antiporter systems, which are thought to counteract external acidification through the consumption of intracellular protons and the generation of alkaline amines. Additional acid tolerance responses include regulation of proton permeability by induction of membrane proteins and lipid modification enzyme. In the case of alkaline stress, amino acid metabolic enzymes such as tryptophan deaminase (TnaA) and o-acetylserine sulfhydrylase A (CysK) are induced to reverse alkalinization by metabolizing amino acids to produce acidic products.
The response of S. oneidensis to acid and alkaline stresses intersects with other stresses evidenced by elevated expression of RpoS, a central regulator of stationary-phase gene expression [30]. It is reasonable to speculate that S. oneidensis cells upon altered pH mimic those at the stationary phase. In respect of response to acidic pH, the mechanism of S. oneidensis is fundamentally different from that of E. coli. The most important and effective player of E. coli in mediating acid resistance is the glutamate-dependent (Gad) system, which is missing in all sequenced Shewanellae [31]. Additionally, none of genes encoding H+ ex-pumps are found to be induced. Instead, proteins showing substantial induction are rather diverse, including those functioning in cell envelope structure (e.g., csg genes), glycogen biosynthesis (glg operon), fatty acid metabolism (fadBA), glutamate synthesis (gltBD), phosphate transport (so1724 and pstB-1), and regulation (e.g., rpoS and phoU). This observation indicates that the molecular effects of acute acidic pH are profound and multifarious. Upon alkaline pH, as in E. coli Na+/H+ antiporter systems (NhaA) are particularly important in maintaining a pH-homeostatic mechanism, thus enabling S. oneidensis to survive and adapt to external alkaline conditions.
4. Stress Responses to Osmolarity
The bacterial response to hypertonic stress includes a range of mechanisms. The most important one is regulation of aquaporins in the outer membrane for water intake by the stationary-phase sigma factor, RpoS [32]. It is common that upon the stress condition K+ uptake is activated and K+ ions are maintained at high levels. Additionally, cells accumulate neutral, polar, small molecules, such as glycine betaine (GB), proline, trehalose, or ectoine [33]. These compatible solutes serve as osmoprotectants and are synthesized and/or imported into the cell. Many Shewanella species are marine microorganisms and therefore are naturally tolerant to relatively high levels of salt. Although some like S. oneidensis, are obtained from freshwater environments, they are able to grow in the presence of up to 0.6 M NaCl [34].
The primary response of S. oneidensis to hyperosmotic conditions is similar to E. coli. Genes encoding K+ uptake proteins, Na+ efflux system components, and glutamate synthesis are found to be highly induced. Nonetheless, some novel mechanisms are observed. Genes encoding proteins involved in accumulation of compatible osmolytes are either missing in the genome or transcriptionally unaffected when encountered stress. Interestingly, genes encoding TCA cycle are particularly active, probably producing much needed ATP for ion transport. This may also explain that S. oneidensis shows reduced motility and chemotaxis responding capability under the stress given that the assembly of flagella is extremely energy consuming [34].
5. Stress Responses to Radiation
Radiation is potentially lethal and mutagenic to all organisms. Although DNA is the major chromophore in general, effects of radiation are in fact pleiotropic [35, 36]. S. oneidensis, one of the most radiation-sensitive organisms known so far, is approximately 1 order of magnitude more susceptible to all wavelengths of solar UV, UV, and ionizing radiation than E. coli [35, 37–40]. This is strikingly because the organism similar to E. coli possesses the complete set of genes for photo-reactivation, and nucleotide excision repair, and SOS response, primary mechanisms that protect cells from DNA damages and radiation-induced oxidative stress [16, 41, 42]. All of these S. oneidensis genes appear to be functional and crucial in the cellular response to radiation, supported by significant upregulation in transcriptional analyses. It is interesting to note that Shewanella strains vary significantly in their susceptibility to radiation although compared to E. coli they are still much less resistant. The general trend is that the more radiation exposure is in the habitat where the organisms are isolated the less sensitive they are [37]. For instance, S. oneidensis MR-1 from lake sediment and S. putrefaciens 200 from a crude oil pipeline are more sensitive to radiation than S. algae from the surface of a red alga and S. oneidensis MR-4 from the surface of the Black Sea [33].
It has been suggested that the hypersensitivity to radiation may be in part due to the activation of prophage [38–40]. Radiation has been used as a standard approach to induce prophage in a variety of bacteria [43, 44]. In S. oneidensis, upon radiation the majority of LambdaSo, MuSo1, and MuSo2 genes are induced and phage particles have been found in the cultures, indicating that a great number of cells are lysed by lytic phages. It has also been implicated that a large number of iron-containing proteins may be partially accountable for the susceptibility. Compared to E. coli which hosts only five to seven cytochrome c proteins, S. oneidensis contains 41 such proteins, some of which are electron transport proteins and essential in respiration [45, 46]. Damages on these proteins by reactive oxygen species (ROS) generated in cells upon radiation would likely cause two detrimental results [47]. First, damaged proteins per se may be dysfunctional, directly reducing ability to survive or thrive. Second, damaged proteins release irons into cultures, which further induce ROS production [48]. This second wave of ROS may be more fatal because it comes at the onset of recovery of seriously damaged cells. Furthermore, the finding that the intracellular Mn/Fe concentration ratios correlate well with resistance to radiation may explain the hypersensitivity of S. oneidensis, which has the lowest ratio among bacteria tested so far [35, 49].
6. Stress Responses to Heave Metals
Many of metal elements are required for microbial growth mostly as cofactors in metabolic pathways. However, they exert deleterious effects under conditions of elevated concentration [50]. Shewanellae have attracted much attention because of their ability to reduce metal ions including chromium, cobalt, iron, manganese, technetium, uranium, and vanadium, some of which are not needed and highly toxic for most organisms [10, 51, 52]. At the low level these metal ions are taken as electron acceptors by cells and mildly induced some stress-associated genes [53]. However, at the high concentration some of them elicited a distinctively different pattern [54–60]. The cellular resistance mechanisms displayed by microorganisms are diverse and include biosorption, diminished intracellular accumulation through either direct obstruction of the ion uptake system or active chromate efflux, precipitation, and reduction of metals to less toxic form. Multiple regulatory circuits are found to work together to cope with the stress response of S. oneidensis to heavy metal compounds. The major ones include those modulating oxidative stress protection, detoxification, protein stress protection, iron acquisition, and DNA repair [50].
The molecular response of S. oneidensis to heavy metal shock elicits a distinctively different transcriptional profile compared with metal reduction [53–60]. This observation is consistent with that metal reduction and toxicity resistance mechanisms are to be unlinked cellular processes [61]. Responses of S. oneidensis to acute stresses imposed by a variety of heavy metals share a common strategy: survive first and then exert both general and specific stress responses. As a result, S. oneidensis up-regulates its resistance-nodulation-cell division (RND) protein family genes that facilitate cation export and thus confer heavy metal resistance. Once the first line of defense is initiated, cells employ both general and specific stress responses that are inseparable from each other to recover from the crisis. Alternative sigma factors including RpoS, RpoH, RpoE, along with stress-response-related genes are induced, leading to induction of a variety of detoxification, resistance, and transport functions. Such coordinated expression of stress response and detoxification mechanisms in S. oneidensis may offer an advantage to thrive in anoxic metal-reducing conditions in aquatic sediment and submerged soil systems where substantial amounts of heavy metals can be generated.
Two specific responding mechanisms are particularly worth noting. The first is that genes/proteins involved in iron transport are transcriptionally active and implicated to play an important role in the process. Although induction of siderophore biosynthetic and iron transport genes may not be a direct consequence of intracellular iron limitation, several lines of evidence suggest that it is more likely to be indirect by interfering with the Fur (ferric uptake regulator) protein, which eventually results in derepression of the iron regulon. Several reports have demonstrated that Co2+, Mn2+, or other divalent cations interact with the Fur-binding sites [62, 63]. Moreover, iron-chelating siderophores from other microorganisms have been shown to be able to bind other metals, such as thorium, uranium, vanadium, and plutonium [64, 65]. By increasing siderophore production, cells can reduce toxicity of heavy metals by sequestration. The other is that sulfur transport and assimilation is promoted. While the underlying mechanism is currently unknown, an explanation is offered. In S. oneidensis, reactive oxygen species (ROS) produced in cells by heavy metal stresses can damage iron-containing proteins. As cysteine residues in these proteins are essential to their functions, an extra mount of cysteine is needed for protection. To this end, cells elevate transportation of inorganic sulfate which is reduced and incorporated into bioorganic compounds via assimilatory sulfate reduction, which is the major route of cysteine biosynthesis in most microorganisms [66].
7. Concluding Remarks
As a potential strategy for the reductive immobilization or detoxification of environmental contaminants, in situ bioremediation has received much interest and attention in last 20 years and are becoming more prevalent today. As its intrinsic feature, the application puts its work force, mostly bacteria, “in situ” facing the unpredictability of individual microbial processes and constant fluctuations in environments. Thanks to the availability of the S. oneidensis genome sequence, stress responses of the microorganism have been extensively investigated, generating a handful of insights into mechanisms adopted to cope with detrimental conditions. Nonetheless, adaptive mechanisms of Shewanella to environmental stresses are still a large playing field for three reasons. First, a number of common stressful agents, especially reactive oxygen species, are not visited. Second, the complex components and regulation in the bacterial stress responses discussed in this paper are mostly based on transcriptional profiling and thus experimental validation is urgently warranted. Last, but definitely not the least, the genus is composed of members which are not only isolated from extremely diverse habitats but also lack unifying phenotypic features, prompting exploration to be extended to other ecological groups of the Shewanellae.
Acknowledgments
This study was financially supported by Major Program of Science and Technology Department of Zhejiang (2009C12061), by the Fundamental Research Funds for the Central Universities, and by Major State Basic Research Development Program (973 Program: 2010CB833803).
References
- 1.Gerday C, Glansdorff N. Physiology and Biochemistry of Extremophiles. Washington, DC, USA: American Society for Microbiology; 2007. [Google Scholar]
- 2.Storz G, Hengge-Aronis R. Bacterial Stress Responses. Washington, DC, USA: American Society for Microbiology; 2000. [Google Scholar]
- 3.Ron E. Bacterial stress response. In: Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E, editors. The Prokaryotes. Singapore: Springer; 2006. [Google Scholar]
- 4.Fields S, Johnston M. Whither model organism research? Science. 2005;307(5717):1885–1886. doi: 10.1126/science.1108872. [DOI] [PubMed] [Google Scholar]
- 5.MacDonell MT, Colwell RR. Phylogeny of the Vibrionaceae, and recommendation for 2 new genera, Listonella and Shewanella . Systematic and Applied Microbiology. 1985;6(2):171–182. [Google Scholar]
- 6.Gauthier G, Gauthier M, Christen R. Phylogenetic analysis of the genera Alteromonas, Shewanella, and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera, Alteromonas (emended) and Pseudoalteromonas gen. nov., and proposal of twelve new species combinations. International Journal of Systematic Bacteriology. 1995;45(4):755–761. doi: 10.1099/00207713-45-4-755. [DOI] [PubMed] [Google Scholar]
- 7.Ivanova EP, Flavier S, Christen R. Phylogenetic relationships among marine Alteromonas-like proteobacteria: emended description of the family Alteromonadaceae and proposal of Pseudoalteromonadaceae fam. nov., Colwelliaceae fam. nov., Shewanellaceae fam. nov., Moritellaceae fam. nov., Ferrimonadaceae fam. nov., Idiomarinaceae fam. nov. and Psychromonadaceae fam. nov. International Journal of Systematic and Evolutionary Microbiology. 2004;54(5):1773–1788. doi: 10.1099/ijs.0.02997-0. [DOI] [PubMed] [Google Scholar]
- 8.Gao H, Obraztova A, Stewart N, et al. Shewanella loihica sp. nov., isolated from iron-rich microbial mats in the Pacific Ocean. International Journal of Systematic and Evolutionary Microbiology. 2006;56(8):1911–1916. doi: 10.1099/ijs.0.64354-0. [DOI] [PubMed] [Google Scholar]
- 9.Verma P, Pandey PK, Gupta AK, et al. Shewanella indica sp. nov., isolated from the sediment of Arabian Sea. doi: 10.1099/ijs.0.026310-0. International Journal of Systematic and Evolutionary Microbiology. In press. [DOI] [PubMed] [Google Scholar]
- 10.Nealson KH, Saffarini D. Iron and manganese in anaerobic respiration: environmental significance, physiology, and regulation. Annual Review of Microbiology. 1994;48(1):311–343. doi: 10.1146/annurev.mi.48.100194.001523. [DOI] [PubMed] [Google Scholar]
- 11.Fredrickson JK, Romine MF, Beliaev AS, et al. Towards environmental systems biology of Shewanella . Nature Reviews Microbiology. 2008;6(8):592–603. doi: 10.1038/nrmicro1947. [DOI] [PubMed] [Google Scholar]
- 12.Fredrickson JK, Zachara JM. Electron transfer at the microbe-mineral interface: a grand challenge in biogeochemistry. Geobiology. 2008;6(3):245–253. doi: 10.1111/j.1472-4669.2008.00146.x. [DOI] [PubMed] [Google Scholar]
- 13.Lovley DR. The microbe electric: conversion of organic matter to electricity. Current Opinion in Biotechnology. 2008;19(6):564–571. doi: 10.1016/j.copbio.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 14.Konstantinidis KT, Serres MH, Romine MF, et al. Comparative systems biology across an evolutionary gradient within the Shewanella genus. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(37):15909–15914. doi: 10.1073/pnas.0902000106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Simpson PJL, Richardson DJ, Codd R. The periplasmic nitrate reductase in Shewanella: the resolution, distribution and functional implications of two NAP isoforms, NapEDABC and NapDAGHB. Microbiology. 2010;156(2):302–312. doi: 10.1099/mic.0.034421-0. [DOI] [PubMed] [Google Scholar]
- 16.Heidelberg JF, Paulsen IT, Nelson KE, et al. Genome sequence of the dissimilatory metal ion-reducing bacterium Shewanella oneidensis . Nature Biotechnology. 2002;20(11):1118–1123. doi: 10.1038/nbt749. [DOI] [PubMed] [Google Scholar]
- 17.Lindquist S, Craig EA. The heat-shock proteins. Annual Review of Genetics. 1988;22:631–677. doi: 10.1146/annurev.ge.22.120188.003215. [DOI] [PubMed] [Google Scholar]
- 18.Yura T, Nakahigashi K. Regulation of the heat-shock response. Current Opinion in Microbiology. 1999;2(2):153–158. doi: 10.1016/S1369-5274(99)80027-7. [DOI] [PubMed] [Google Scholar]
- 19.Weber MH, Marahiel MA. Bacterial cold shock responses. Science Progress. 2003;86(1-2):9–75. doi: 10.3184/003685003783238707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gao H, Wang Y, Liu X, et al. Global transcriptome analysis of the heat shock response of Shewanella oneidensis . Journal of Bacteriology. 2004;186(22):7796–7803. doi: 10.1128/JB.186.22.7796-7803.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Gao H, Yang ZK, Wu L, Thompson DK, Zhou J. Global transcriptome analysis of the cold shock response of Shewanella oneidensis MR-1 and mutational analysis of its classical cold shock proteins. Journal of Bacteriology. 2006;188(12):4560–4569. doi: 10.1128/JB.01908-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Nonaka G, Blankschien M, Herman C, Gross CA, Rhodius VA. Regulon and promoter analysis of the E. coli heat-shock factor, sigma32, reveals a multifaceted cellular response to heat stress. Genes and Development. 2006;20(13):1776–1789. doi: 10.1101/gad.1428206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Turatsinze JV, Thomas-Chollier M, Defrance M, van Helden J. Using RSAT to scan genome sequences for transcription factor binding sites and cis-regulatory modules. Nature Protocols. 2008;3(10):1578–1588. doi: 10.1038/nprot.2008.97. [DOI] [PubMed] [Google Scholar]
- 24.Phadtare S, Inouye M. Genome-wide transcriptional analysis of the cold shock response in wild-type and cold-sensitive, quadruple-csp-deletion strains of Escherichia coli . Journal of Bacteriology. 2004;186(20):7007–7014. doi: 10.1128/JB.186.20.7007-7014.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li S, Xiao X, Sun P, Wang F. Screening of genes regulated by cold shock in Shewanella piezotolerans WP3 and time course expression of cold-regulated genes. Archives of Microbiology. 2008;189(6):549–556. doi: 10.1007/s00203-007-0347-1. [DOI] [PubMed] [Google Scholar]
- 26.Wang F, Xiao X, Ou HY, Gai Y. Role and regulation of fatty acid biosynthesis in the response of Shewanella piezotolerans WP3 to different temperatures and pressures. Journal of Bacteriology. 2009;191(8):2574–2584. doi: 10.1128/JB.00498-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wang F, Li Q, Xiao X. A novel filamentous phage from the deep-sea bacterium Shewanella piezotolerans WP3 is induced at low temperature. Journal of Bacteriology. 2007;189(19):7151–7153. doi: 10.1128/JB.00569-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Foster JW. Microbial responses to acid stress. In: Storz G, Hengge-Aronis R, editors. Bacterial Stress Responses. Washington, DC, USA: American Society for Microbiology; 2000. [Google Scholar]
- 29.Saito H, Kobayashi H. Bacterial responses to alkaline stress. Science Progress. 2003;86(4):271–282. doi: 10.3184/003685003783238635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Leaphart AB, Thompson DK, Huang K, et al. Transcriptome profiling of Shewanella oneidensis gene expression following exposure to acidic and alkaline pH. Journal of Bacteriology. 2006;188(4):1633–1642. doi: 10.1128/JB.188.4.1633-1642.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Foster JW. Escherichia coli acid resistance: tales of an amateur acidophile. Nature Reviews Microbiology. 2004;2(11):898–907. doi: 10.1038/nrmicro1021. [DOI] [PubMed] [Google Scholar]
- 32.Csonka LN. Physiological and genetic responses of bacteria to osmotic stress. Microbiological Reviews. 1989;53(1):121–147. doi: 10.1128/mr.53.1.121-147.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jebbar M, Sohn-Bosser L, Bremer E, Bernard T, Blanco C. Ectoine-induced proteins in Sinorhizobium meliloti include an ectoine ABC-type transporter involved in osmoprotection and ectoine catabolism. Journal of Bacteriology. 2005;187(4):1293–1304. doi: 10.1128/JB.187.4.1293-1304.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liu Y, Gao W, Wang Y, et al. Transcriptome analysis of Shewanella oneidensis MR-1 in response to elevated salt conditions. Journal of Bacteriology. 2005;187(7):2501–2507. doi: 10.1128/JB.187.7.2501-2507.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ghosal D, Omelchenko MV, Gaidamakova EK, et al. How radiation kills cells: survival of Deinococcus radiodurans and Shewanella oneidensis under oxidative stress. FEMS Microbiology Reviews. 2005;29(2):361–375. doi: 10.1016/j.femsre.2004.12.007. [DOI] [PubMed] [Google Scholar]
- 36.Daly MJ, Gaidamakova EK, Matrosova VY, et al. Protein oxidation implicated as the primary determinant of bacterial radioresistance. PLoS Biology. 2007;5(4, article e92) doi: 10.1371/journal.pbio.0050092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Qiu X, Sundin GW, Chai B, Tiedje JM. Survival of Shewanella oneidensis MR-1 after UV radiation exposure. Applied and Environmental Microbiology. 2004;70(11):6435–6443. doi: 10.1128/AEM.70.11.6435-6443.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Qiu X, Sundin GW, Wu L, Zhou J, Tiedje JM. Comparative analysis of differentially expressed genes in Shewanella oneidensis MR-1 following exposure to UVC, UVB, and UVA radiation. Journal of Bacteriology. 2005;187(10):3556–3564. doi: 10.1128/JB.187.10.3556-3564.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Qiu X, Tiedje JM, Sundin GW. Genome-wide examination of the natural solar radiation response in Shewanella oneidensis MR-1. Photochemistry and Photobiology. 2005;81(6):1559–1568. doi: 10.1562/2005-04-15-RA-490. [DOI] [PubMed] [Google Scholar]
- 40.Qiu X, Daly MJ, Vasilenko A, et al. Transcriptome analysis applied to survival of Shewanella oneidensis MR-1 exposed to ionizing radiation. Journal of Bacteriology. 2006;188(3):1199–1204. doi: 10.1128/JB.188.3.1199-1204.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Friedberg EC, Walker GC, Siede W. DNA Repair and Mutagenesis. Washington, DC, USA: American Society for Microbiology; 1995. [Google Scholar]
- 42.Janion C. Inducible SOS response system of DNA repair and mutagenesis in Escherichia coli . International Journal of Biological Sciences. 2008;4(6):338–344. doi: 10.7150/ijbs.4.338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lamont I, Brumby AM, Egan JB. UV induction of coliphage 186: prophage induction as an SOS function. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(14):5492–5496. doi: 10.1073/pnas.86.14.5492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liu Y, Zhang Q, Fang C, Zhu S, Tang Y, Huang S. Effect of glutathione on UV induction of prophage lambda. Archives of Microbiology. 2005;183(6):444–449. doi: 10.1007/s00203-005-0019-y. [DOI] [PubMed] [Google Scholar]
- 45.Gao H, Barua S, Liang Y, et al. Impacts of Shewanella oneidensis c-type cytochromes on aerobic and anaerobic respiration. Microbial Biotechnology. 2010;3(4):455–466. doi: 10.1111/j.1751-7915.2010.00181.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Blattner FR, Plunkett G, III, Bloch CA, et al. The complete genome sequence of Escherichia coli K-12. Science. 1997;277(5331):1453–1462. doi: 10.1126/science.277.5331.1453. [DOI] [PubMed] [Google Scholar]
- 47.Py B, Barras F. Building Feg-S proteins: bacterial strategies. Nature Reviews Microbiology. 2010;8(6):436–446. doi: 10.1038/nrmicro2356. [DOI] [PubMed] [Google Scholar]
- 48.Imlay JA. Cellular defenses against superoxide and hydrogen peroxide. Annual Review of Biochemistry. 2008;77:755–776. doi: 10.1146/annurev.biochem.77.061606.161055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Daly MJ, Gaidamakova EK, Matrosova VY, et al. Accumulation of Mn(II) in Deinicoccus radiodurans facilitates gamma-radiation resistance. Science. 2004;306(5698):1025–1028. doi: 10.1126/science.1103185. [DOI] [PubMed] [Google Scholar]
- 50.Nies DH. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiology Reviews. 2003;27(2-3):313–339. doi: 10.1016/S0168-6445(03)00048-2. [DOI] [PubMed] [Google Scholar]
- 51.Liu C, Gorby YA, Zachara JM, Fredrickson JK, Brown CF. Reduction kinetics of Fe(III), Co(III), U(VI), Cr(VI), and Tc(VII) in cultures of dissimilatory metal-reducing bacteria. Biotechnology and Bioengineering. 2002;80(6):637–649. doi: 10.1002/bit.10430. [DOI] [PubMed] [Google Scholar]
- 52.Carpentier W, Sandra K, De Smet I, Brigé A, De Smet L. Microbial reduction and precipitation of vanadium by Shewanella oneidensis . Applied and Environmental Microbiology. 2003;69(6):3636–3639. doi: 10.1128/AEM.69.6.3636-3639.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Beliaev AS, Klingeman DM, Klappenbach JA, et al. Global transcriptome analysis of Shewanella oneidensis MR-1 exposed to different terminal electron acceptors. Journal of Bacteriology. 2005;187(20):7138–7145. doi: 10.1128/JB.187.20.7138-7145.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Saltikov CW, Cifuentes A, Venkateswaran K, Newman DK. The ars detoxification system is advantageous but not required for As(V) respiration by the genetically tractable Shewanella species strain ANA-3. Applied and Environmental Microbiology. 2003;69(5):2800–2809. doi: 10.1128/AEM.69.5.2800-2809.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Bencheikh-Latmani R, Williams SM, Haucke L, et al. Global transcriptional profiling of Shewanella oneidensis MR-1 during Cr(VI) and U(VI) reduction. Applied and Environmental Microbiology. 2005;71(11):7453–7460. doi: 10.1128/AEM.71.11.7453-7460.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Brown SD, Martin M, Deshpande S, et al. Cellular response of Shewanella oneidensis to strontium stress. Applied and Environmental Microbiology. 2006;72(1):890–900. doi: 10.1128/AEM.72.1.890-900.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Brown SD, Thompson MR, VerBerkmoes NC, et al. Molecular dynamics of the Shewanella oneidensis response to chromate stress. Molecular and Cellular Proteomics. 2006;5(6):1054–1071. doi: 10.1074/mcp.M500394-MCP200. [DOI] [PubMed] [Google Scholar]
- 58.Chourey K, Thompson MR, Morrell-Falvey J, et al. Global molecular and morphological effects of 24-hour chromium(VI) exposure on Shewanella oneidensis MR-1. Applied and Environmental Microbiology. 2006;72(9):6331–6344. doi: 10.1128/AEM.00813-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Toes A-CM, Daleke MH, Kuenen JG, Muyzer G. Expression of copA and cusA in Shewanella during copper stress. Microbiology. 2008;154(9):2709–2718. doi: 10.1099/mic.0.2008/016857-0. [DOI] [PubMed] [Google Scholar]
- 60.Mugerfeld I, Law BA, Wickham GS, Thompson DK. A putative azoreductase gene is involved in the Shewanella oneidensis response to heavy metal stress. Applied Microbiology and Biotechnology. 2009;82(6):1131–1141. doi: 10.1007/s00253-009-1911-1. [DOI] [PubMed] [Google Scholar]
- 61.Cervantes C, Campos-Garcia J, Devars S, et al. Interactions of chromium with microorganisms and plants. FEMS Microbiology Reviews. 2001;25(3):335–347. doi: 10.1111/j.1574-6976.2001.tb00581.x. [DOI] [PubMed] [Google Scholar]
- 62.Funahashi T, Fujiwara C, Okada M, et al. Characterization of Vibrio parahaemolyticus manganese-resistant mutants in reference to the function of the ferric uptake regulatory protein. Microbiology and Immunology. 2000;44(12):963–970. doi: 10.1111/j.1348-0421.2000.tb02591.x. [DOI] [PubMed] [Google Scholar]
- 63.Hantke K. Iron and metal regulation in bacteria. Current Opinion in Microbiology. 2001;4(2):172–177. doi: 10.1016/s1369-5274(00)00184-3. [DOI] [PubMed] [Google Scholar]
- 64.Baysse C, De Vos D, Naudet Y, et al. Vanadium interferes with siderophore-mediated iron uptake in Pseudomonas aeruginosa . Microbiology. 2000;146(10):2425–2434. doi: 10.1099/00221287-146-10-2425. [DOI] [PubMed] [Google Scholar]
- 65.John SG, Ruggiero CE, Hersman LE, Tung CS, Neu MP. Siderophore mediated plutonium accumulation by Microbacterium flavescens (JG-9) Environmental Science and Technology. 2001;35(14):2942–2948. doi: 10.1021/es010590g. [DOI] [PubMed] [Google Scholar]
- 66.Kertesz MA. Riding the sulfur cycle—metabolism of sulfonates and sulfate esters in gram-negative bacteria. FEMS Microbiology Reviews. 2000;24(2):135–175. doi: 10.1016/S0168-6445(99)00033-9. [DOI] [PubMed] [Google Scholar]