

Abstract
Urban regions are among the most human-altered environments on Earth and they are poised for rapid expansion following population growth and migration. Identifying the biological traits that determine which species are likely to succeed in urbanized habitats is important for predicting global trends in biodiversity. We provide the first evidence for the intuitive yet untested hypothesis that relative brain size is a key factor predisposing animals to successful establishment in cities. We apply phylogenetic mixed modelling in a Bayesian framework to show that passerine species that succeed in colonizing at least one of 12 European cities are more likely to belong to big-brained lineages than species avoiding these urban areas. These data support findings linking relative brain size with the ability to persist in novel and changing environments in vertebrate populations, and have important implications for our understanding of recent trends in biodiversity.
Keywords: brain size, comparative analyses, urbanization, innovation, colonization
1. Introduction
Human-made environments increasingly dominate the planet and urban areas are among the most rapidly developing ones—by the year 2030, the number of people who dwell in cities will increase by 1.75 billion [1,2]. Urbanization dramatically influences biodiversity on both local and global scales [1]. However, while an urban environment is hostile to many organisms, some species thrive in this novel habitat [3]. Understanding the biological prerequisites for successful colonization of urban habitats is key for predicting the long-term trends in biodiversity in response to human-induced environmental change. This is imperative, since future urbanization is likely to endanger a large number of vertebrate species on Earth [4].
Relatively larger brain size is associated with the ability for behavioural innovations in birds and mammals [5–7] and higher long-term abundance in farmland birds [8]. Additionally, large brain size has been shown to improve the success of avian species in novel environments following human-induced introduction [9]. Based on this evidence, we hypothesized that large brain size can predispose bird species for successful establishment in cities. We used a recent statistical approach [10,11] to show that urban bird species are indeed more likely to possess large brains, as well as belonging to large-brained families, than species that avoid urban habitats.
Our analysis focused on the published records for common species of passerine birds in and around 12 representative cities in France and Switzerland that can be categorized into those that flourish in the city and those that avoid urban habitats [12]. We constructed a phylogenetic mixed model using a Bayesian framework [11] to ask (i) whether brain size corrected for body size is positively associated with species success in the urban environment (species-level analysis) and (ii) whether large-brained families contain a higher proportion of successful colonizers of urban areas (family-level analysis).
2. Material and methods
We obtained published data on urban tolerance, brain size and body mass for 82 species of passerine birds belonging to 22 families. Urban tolerance was assessed in the vicinity of 12 representative cities in central Europe [12]. Bird species that were able to breed in the city centres (i.e. excluding those species that only breed at the edges of cities) were considered successful colonizers [12]. All passerines that were breeding in the area of 2592 km2 (an atlas unit used in the original study) around each of the 12 cities but not recorded as breeding in urban centres were considered as urban avoiders [12]. We matched these records with data on avian brain size compiled from the primary literature [13–15]. This resulted in similar numbers of species for successful (n = 38) and unsuccessful (n = 44) colonizers of city centres (see the electronic supplementary material, table S1).
To investigate whether relative brain size was associated with the successful colonization of urban areas across species, we ran a multi-variate generalized mixed effect model (GLMM) in a phylogenetic framework using a Bayesian approach [11] based on a Markov Chain Monte Carlo (MCMC) algorithm (package MCMCglmm v. 2.0.3 for R v. 2.10.1 [16]). This statistical approach allows the use of binomial response variables while controlling for statistical non-independence of the data points owing to shared ancestry. The first model at the species level included a binomial response variable (breeding in the city ‘yes–no’), ln-transformed brain size and body mass as explanatory variables and phylogeny as a random factor (see the electronic supplementary material, table S1). Owing to lack of ad hoc values for the variance in the data, we used uninformative, non-proper priors for the fixed effects (V = 0.1, ν = −1) and an uninformative, proper prior for the random effect (V = 1, ν = 1, α × ν = 0, α × V = 252, i.e. Cathy prior). To test whether large-brained families contribute proportionally more species to city-dwelling populations, we performed a model with binomial response (no. of colonizer spp. − no. of avoider species) and ln-transformed brain size and body mass (mean values for each family; see the electronic supplementary material, table S2) as explanatory variables. The priors for this model were set as uninformative, proper prior for fixed effects (V = 1, ν = 1) and a Cathy prior (see above) for random effects. For both models, we ran one chain with 1 010 000 iterations with a burn-in of 10 000 and a thinning interval of 100 resulting in a sample size of 1000 per chain. These settings resulted in appropriate conversion of the chain. The topology for the phylogenies was adopted from published sources (see the electronic supplementary material, figures S1 and S2) and because information on branch lengths could not be combined between sources, branch lengths were standardized to 1.
3. Results
Larger brain size was significantly positively associated (indicated by the 95% credibility interval excluding 0) with the ability of a species to prosper in the city (figure 1; binomial phylogenetic mixed model [11]: brain size: posterior mean, 533.8 ± 265.3 s.d.; 95% credibility interval, 124.3–1163.5; body size: posterior mean, −419.1 ± 196.8 s.d.; 95% credibility interval, − 881.8 to − 109.5; figure 1; see the electronic supplementary material, table S3a for complete statistics). In addition, families with larger brains contributed more species to city populations than small-brained families (brain size: posterior mean, 7.59 ± 8.80 s.d.; 95% credibility interval, 3.15–33.99; body size: posterior mean, −5.72 ± 6.12 s.d.; 95% credibility interval, −24.06 to −2.41; figure 2; see electronic supplementary material, table S3b for complete statistics). The significantly negative effect of body mass needs some further investigation. However, models with body or brain size separately revealed no significant effects at the species or at the family level. In addition, the pattern still holds at the species level when using the ln-ratio between brain and body size as predictor variable (not shown). We emphasize that the pattern emerges despite the simplicity of our model, which ignores variation owing to ecological factors [1].
Figure 1.

Box-plot displaying the difference (median, 25% and 75% percentiles and sample minimum and maximum) in relative brain size between species breeding in cities (‘yes’, purple colour) and species that avoid cities (‘no’, blue colour). Relative brain size represents residual values obtained from a linear regression between ln-transformed brain and body mass. Unlike the statistical analyses, this figure is not controlled for phylogeny.
Figure 2.
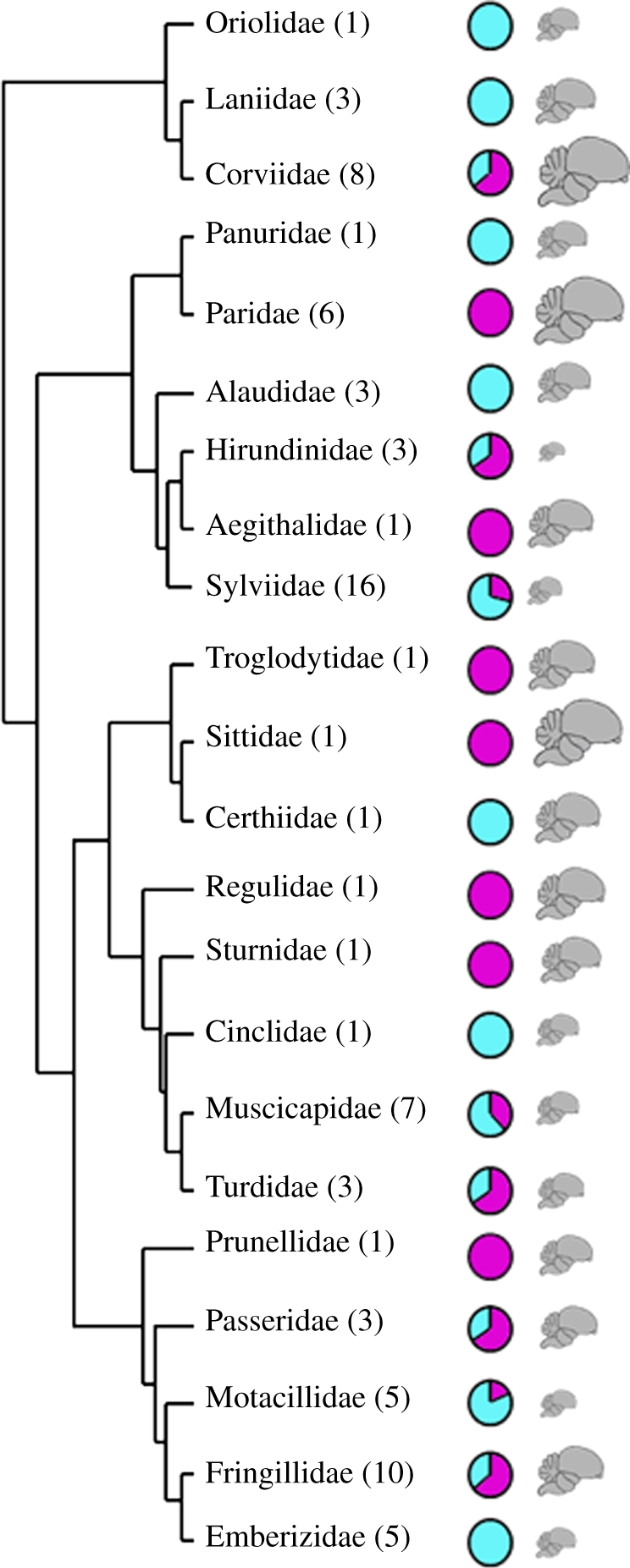
Phylogenetic tree describing the relationship between 22 avian families studied (number of included species per family in parentheses). The pie charts indicate the proportion of urban dwellers (purple colour) versus urban avoiders (blue colour) among species within each family. The schematic of the avian brain is scaled to match the relative mean brain size for each family.
4. Discussion
We showed that species of passerine birds that breed in at least one city centre have relatively larger brains and are more likely to belong to large-brained families than their counterparts that avoid urban habitats. This finding supports the hypothesis that large brain size predisposes avian species for successful establishment in urban environments. Brain size has been repeatedly linked to the ability of animals to adapt to novel or changing environmental conditions [9,17] as well as to innovative behaviour [18], which could prove advantageous in such environmental conditions. Therefore, it is logical to conjecture that brain size can be related to the success in an urban environment, which is both novel and harsh [4].
We used the relative size of the whole brain as our predictor for the proportion of city dwellers within passerine families, although it is possible that the effect is mediated through increased size of particular brain structures. For example, it is possible that were the analyses repeated using telencephalic size, an even stronger result could be found (e.g. [8]). Indeed, innovation rate is most strongly related to the size of isocortex in mammals [6] and nidopallium/mesopallium complex in birds (reviewed in [18]) and it is possible that the same telencephalic structures could play an important role in the relationship between relative brain size and urban breeding reported in this study. The downside of this approach is that currently there are very little data available on brain structure in different bird species, which would reduce our study size. In any case, because the nidopallium/mesopallium is very large and avian taxa with large brains are also characterized by large nidopallium/mesopallium [19], relative brain size, which explains 96 per cent of variance in these telencephalic structures, can be an adequate proxy in comparative analyses [18]. Future studies may gain by adopting a more detailed approach, but it remains to be seen whether the size of nidopallium/mesopallium complex, or any other structure of the avian brain, will prove a better predictor of the successful breeding in urban environments.
Biological factors such as song type [20], environmental tolerance [3] or brain size (these results) can result in only a handful of species succeeding in urban environments, leading to homogenization of fauna in cities [21]. Our study suggests that birds from relatively small-brain size lineages are exposed to greater risks as a result of urbanization. The potential effect of relative brain size on current trends in avian diversity could be even more disastrous if one considers recent findings indicating that small-brained species of British farmland birds were more likely to suffer strong long-term declines compared with large-brained ones [8]. While the urban environment is rapidly developing, the combined effect of urban land and farmland can lead to even faster loss of birds from small-brained lineages from local avifauna.
This study focused on passerines in and around European cities, but certainly other avian taxa flourish in urban environments around the world. Further research is needed to test the generality of these patterns across taxonomic groups and geographical areas. These data also points to a possibility that novel environments, such as cities, may impose selection on brain size in urban populations of birds. Recent work also indicated that an urban environment leads to rapid evolution of morphological traits in birds [22]. We suggest that future research in this field will benefit from focusing on the contemporary evolution of brain size and structure in urban populations of vertebrates.
Acknowledgements
This study was supported by Swedish Research Council fellowships to A.A.M. and N.K., a Swiss National Foundation postdoctoral fellowship to S.I., a Wenner-Gren postdoctoral fellowship and a Juan de la Cierva postdoctoral fellowship from the Ministerio de Ciencia e Innovación to A.G.-V.
References
- 1.Kareiva P., Watts S., McDonald R., Boucher T. 2007. Domesticated nature: shaping landscapes and ecosystems for human welfare. Science 316, 1866–1869 10.1126/science.1140170 (doi:10.1126/science.1140170) [DOI] [PubMed] [Google Scholar]
- 2.UNPD. 2005. World urbanization prospects: the 2005 revision. New York, NY: United Nations Population Division [Google Scholar]
- 3.Bonier F., Martin P. R., Wingfield J. C. 2007. Urban birds have broader environmental tolerance. Biol. Lett. 3, 670–673 10.1098/rsbl.2007.0349 (doi:10.1098/rsbl.2007.0349) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.McDonald R. I., Kareiva P., Formana R. T. T. 2008. The implications of current and future urbanization for global protected areas and biodiversity conservation. Biol. Conserv. 141, 1695–1703 10.1016/j.biocon.2008.04.025 (doi:10.1016/j.biocon.2008.04.025) [DOI] [Google Scholar]
- 5.Lefebvre L., Reader S. M., Sol D. 2004. Brains, innovations and evolution in birds and primates. Brain Behav. Evol. 63, 233–246 10.1159/000076784 (doi:10.1159/000076784) [DOI] [PubMed] [Google Scholar]
- 6.Reader S. M., Laland K. N. 2002. Social intelligence, innovation, and enhanced brain size in primates. Proc. Natl. Acad. Sci. USA 99, 4436–4441 10.1073/pnas.062041299 (doi:10.1073/pnas.062041299) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sol D., Bacher S., Reader S. M., Lefebvre L. 2008. Brain size predicts the success of mammal species introduced into novel environments. Am. Nat. 172, S63–S71 10.1086/588304 (doi:10.1086/588304) [DOI] [PubMed] [Google Scholar]
- 8.Shultz S., Bradbury R. B., Evans K. L., Gregory R. D., Blackburn T. M. 2005. Brain size and resource specialization predict long-term population trends in British birds. Proc. R. Soc. B 272, 2305–2311 10.1098/rspb.2005.3250 (doi:10.1098/rspb.2005.3250) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sol D., Duncan R. P., Blackburn T. M., Cassey P., Lefebvre L. 2005. Big brains, enhanced cognition, and response of birds to novel environments. Proc. Natl Acad. Sci. USA 102, 5460–5465 10.1073/pnas.0408145102 (doi:10.1073/pnas.0408145102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hadfield J. D. 2010. MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J. Stat. Software 33, 1–22 [Google Scholar]
- 11.Hadfield J. D., Nakagawa S. 2010. General quantitative genetic methods for comparative biology: phylogenies, taxonomies and multi-trait models for continuous and categorical characters. J. Evol. Biol. 23, 494–508 10.1111/j.1420-9101.2009.01915.x (doi:10.1111/j.1420-9101.2009.01915.x) [DOI] [PubMed] [Google Scholar]
- 12.Croci S., Butet A., Clergeau P. 2008. Does urbanization filter birds on the basis of their biological traits? Condor 110, 223–240 10.1525/cond.2008.8409 (doi:10.1525/cond.2008.8409) [DOI] [Google Scholar]
- 13.Iwaniuk A. N., Nelson J. E. 2003. Developmental differences are correlated with relative brain size in birds: a comparative analysis. Can. J. Zool. Rev. Can. Zool. 81, 1913–1928 10.1139/z03-190 (doi:10.1139/z03-190) [DOI] [Google Scholar]
- 14.Mlikovsky J. 1990. Brain size in birds: 4. Passeriformes. Acta Soc. Zool. Bohemoslov. 54, 27–37 [Google Scholar]
- 15.Garamszegi L. Z., Moller A. P., Erritzoe J. 2002. Coevolving avian eye size and brain size in relation to prey capture and nocturnality. Proc. R. Soc. Lond. B 269, 961–967 10.1098/rspb.2002.1967 (doi:10.1098/rspb.2002.1967) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.R Development Core Team 2008. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; (ISBN 3-900051-900007-900050) See http://www.R-project.org [Google Scholar]
- 17.Reader S. M., MacDonald K. 2003. Environmental variability and primate behavioural flexibility. In Animal innovation (eds Reader S. M., Laland K. N.). Oxford, UK: Oxford University Press [Google Scholar]
- 18.Lefebvre L., Bolhuis J. J. 2003. Positive and negative correlates of feeding innovations in birds: evidence for limited modularity. In Animal innovation (eds Reader S. M., Laland K. N.). Oxford, UK: Oxford University Press [Google Scholar]
- 19.Rehkamper G., Frahm H. D., Zilles K. 1991. Quantitative development of brain and brain structures in birds (Galliformes and Passeriformes) compared to that in mammals (insectivores and primates). Brain Behav. Evol. 37, 125–143 10.1159/000114353 (doi:10.1159/000114353) [DOI] [PubMed] [Google Scholar]
- 20.Slabbekoorn H., den Boer-Visser A. 2006. Cities change the songs of birds. Curr. Biol. 16, 2326–2331 10.1016/j.cub.2006.10.008 (doi:10.1016/j.cub.2006.10.008) [DOI] [PubMed] [Google Scholar]
- 21.McKinney M. L. 2006. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 127, 247–260 10.1016/j.biocon.2005.09.005 (doi:10.1016/j.biocon.2005.09.005) [DOI] [Google Scholar]
- 22.Badyaev A. V., Young R. L., Oh K. P., Addison C. 2008. Evolution on a local scale: developmental, functional, and genetic bases of divergence in bill form and associated changes in song structure between adjacent habitats. Evolution 62, 1951–1964 10.1111/j.1558-5646.2008.00428.x (doi:10.1111/j.1558-5646.2008.00428.x) [DOI] [PubMed] [Google Scholar]