

Abstract
Since the early pioneering work of Ballinger and Reckard demonstrating that transplantation of islets of Langerhans into diabetic rodents could normalize their blood glucose levels, islet transplantation has been proposed to be a potential treatment for type 1 diabetes 1,2. More recently, advances in human islet transplantation have further strengthened this view 1,3. However, two major limitations prevent islet transplantation from being a widespread clinical reality: (a) the requirement for large numbers of islets per patient, which severely reduces the number of potential recipients, and (b) the need for heavy immunosuppression, which significantly affects the pediatric population of patients due to their vulnerability to long-term immunosuppression. Strategies that can overcome these limitations have the potential to enhance the therapeutic utility of islet transplantation.
Islet transplantation under the mouse kidney capsule is a widely accepted model to investigate various strategies to improve islet transplantation. This experiment requires the isolation of high quality islets and implantation of islets to the diabetic recipients. Both procedures require surgical steps that can be better demonstrated by video than by text. Here, we document the detailed steps for these procedures by both video and written protocol. We also briefly discuss different transplantation models: syngeneic, allogeneic, syngeneic autoimmune, and allogeneic autoimmune.
Keywords: Medicine, Issue 50, islet isolation, islet transplantation, diabetes, murine, pancreas
Protocol
1. Preparation and Calibration of Liberase Reagent
This protocol utilizes Liberase TL (Roche, Cat# 05 401 020 001) enzyme for pancreatic digestion.
Purchase 100-200mg at a time and make a large batch. Re-suspend lyophilized powder in sterile HPLC grade water so that the concentration is approximately 26 Wünsch units per ml. This should be approximately 5 mg/ml. See the package insert for details on the Wünsch units contained in the specific lot. Place vials on ice for 30 minutes with gentile swirling every few minutes. Check to ensure powder is completely dissolved at the end of 30 minutes.
Pool all dissolved Liberase together and mix with gentle swirling (do NOT vortex or shake). Aliquot 24.3 Wünsch units to pre-chilled Eppendorph tubes, quick freeze on liquid nitrogen (treat it as any enzymes) and store at -80°C. This should be approximately 0.935ml per Eppendorph tube. Each aliquot is for single use (enough for 11 mice) and should not be stored or re-frozen once thawed. In general, the stock is stable for at least 6 months.
- For each new batch, calibrate the optimal incubation time. Use the frozen stock, not the freshly dissolved stock, for calibration, to mimic the real experimental conditions as much as possible.
- Calibrate the water bath that you intend to use to exactly 37°C using a mercury thermometer. Use the same incubator in future experiments.
- Prior to use, thaw a tube of Liberase on ice and add it (0.935ml) to 21.6ml serum free RPMI, making the final working concentration 1.08 Wünsch units per ml. Serum, BAS and EDTA inhibit Liberase. Zinc and calcium are cofactors. Store on ice and use within 1.5 hours. This is enough for approximately 11 mice (2ml/mouse).
- See step 2.3 for pancreas perfusion. Perfuse as many mice as possible in 1 hr. Then use one or two mice for each incubation time and test 10, 12, 14, 16, 18 minute incubation times. If possible, include 30 second intervals between times as the timing of digestion is very important.
- Complete the islet isolation protocol and analyze islet yield and quality from each incubation time point. Ultimately the yield should not change too much between peak times, but the quality of islets at those intervals may vary. For subsequent experiments, use the conditions that result in the greatest number of islets that are healthy 48 hours after isolation. The age of the mice appears to affect the optimal incubation time. So, use mice within a similar age range. Ideally, use 8-12 week old mice; however, even 8-10 month old mice can yield good islets. For calibration purposes, use younger mice.
2. Surgical Procedure
- Prepare all equipment and media before euthanizing the islet donor mice.
- Autoclave or flame sterilize instruments before use. All reagents and instruments (touching the samples) should be sterile. The isolation and hand picking of islets can be performed outside of a laminar flow hood without greatly increasing the incidence of contamination. However, sterilization of tools that come into contact with pancreas or islets is critical for avoiding contamination. The following tools are needed:
- 1 pair of dissecting scissors for abdominal incision.
- 2 pairs of forceps.
- 1 hemostatic forceps to clamp off the bile duct.
- 0.419 mm diameter wire mesh.
- 0.1 mm diameter nylon mesh.
- Bend several 27 gauge needles so that a 70 degree angle is introduced approximately halfway down the length of the needle. The beveled edge of the needle should face the inside of the elbow.
- Fill a spray bottle with 70% ethanol.
- Set up a dissecting microscope with an adequate light source on a lab bench or in a hood.
- Pre-chill centrifuge to 4°C. Centrifuge must have a swing bucket rotor, be capable of 900xG, be refrigerated, and have a break that can be disengaged. On a Sorvall RT6000D with the Sorvall H1000B rotor, a 900xG speed is 2400RPM. Slower spins are done at 800RPM.
- Place the following reagents on ice.
- RPMI (1 liter with 10% serum)
- RPMI (200ml without serum)
- 50ml conical tubes (1 for each pancreas)
- 5ml syringe.
- Equilibrate Histopaque1077 to room temperature.
- Thaw a single use aliquot of calibrated Liberase on ice and dilute in 22.5ml serum free RPMI. Serum, BAS and EDTA inhibit Liberase. Zinc and calcium are cofactors. Store on ice and use within 1.5 hours. This is enough for approximately 11 mice (2ml/mouse).
- Warm 37°C water bath.
- Have on hand as many mice as you can cannulate in 1 hour (approximately 10 - 18 mice).
- Prepare mouse for cannulation. First euthanize the mouse by cervical dislocation or CO2 asphyxiation. While mouse is expiring, fill the 5ml syringe with 2ml Liberase working stock (1.08 Wünsch units /ml).
- Place mouse in supine position on a paper towel at the dissecting scope and spray the abdomen with 70% ethanol before opening its abdomen with a V-incision from the pubic region to both front legs. Fold skin over chest to reveal the abdominal cavity.
- Position the mouse with the head pointing towards you. Locate the duodenal entry of the common bile duct as highlighted in FIG.1. This can be done without looking through dissecting scope objective.
- Clamp the duodenal opening with hemostat as demonstrated in FIG.2. Clamp position is critical for blocking the flow of Liberase into the intestines. If clamp is too high or too low, Liberase will not perfuse the pancreas. Position hemostat, so that when compressed, it runs exactly along the pancreatic/intestine boarder.
- Before cannulating the common bile duct, it may be necessary to reposition the liver by pressing it up against the diaphragm, so that the entire length of the common bile duct is exposed (FIG.3). Once the length of the bile duct exposed, use the bent 27-gague needle attached to the Liberase filled syringe for cannulation. There are two techniques for cannulating the bile duct.
- In the first technique, or 'free hand' method, the hemostat clamped on the duodenum is pulled away from the head of the mouse towards the tail so that the common bile duct becomes taut. There is a confluence in the bile duct close to the liver where bile draining from the gall bladder and enzymes from the liver come together before entering the intestines. The inside of the V formed by this confluence is exposed by pulling the bile duct tight and is an ideal place to initiate cannulation of the duct (FIG.4). If positioned correctly, the needle will slide right through the V, and directly cannulate the lower portion of the duct. Insert the needle several millimeters into the duct before dispensing the Liberase. Optimal needle placement is important to prevent backflow into the liver and gall bladder. In addition, it is important that the needle not be inserted so far that it obstructs the splenic duct. The splenic duct is a difficult to see branch of the common bile duct and it drains the splenic tail of the pancreas, an islet rich area. If the needle slides past the opening for the splenic duct, there will be less than complete perfusion of the splenic tail and potentially reduced digestion of this area. Optimal islet yield occurs when your needle depth is far enough that no Liberase escapes to the liver/gall bladder but not so far that you've passed the splenic duct. You can test for successful cannulation by dispensing a small amount of Liberase. If you see the Liberase filling the duct then dispense the rest of the 2mls. If the area around the duct begins to fill, stop dispensing, reposition the needle, and try again.
- In the second technique, or 'forceps assist' method, the hemostat clamped on the duodenum is pulled away from the head of the mouse towards the tail so that the common bile duct becomes taut. Then, using the bent 27-gague needle attached to the 5ml syringe, puncture the fascia below the common bile duct close to the liver. Use the needle to clear the fascia from the duct (FIG.5). With the needle still in place, set the hemostat down and pick up a pair of forceps. Use one arm of the open forceps to lift up the bile duct where the needle has cleared the fascia. The ridges in the forceps create a tract that you can use to guide the needle into the duct. While pulling the duct and forceps towards you, slide the needle into the duct using the forceps as a backstop. Test for cannulation by dispensing a small amount of Liberase. If the duct begins to fill (FIG.5 Arrow), dispense the rest of the 2mls. If the area around the duct begins to fill, stop dispensing, reposition the needle and try again.
As the 2mls of Liberase fills the pancreas, the area near the duodenum begins to expand first, followed by the region on top of the stomach, and finally the splenic tail (FIG.6). Look for even expansion of the pancreas (FIG.6). If one area begins to expand and you don't see the white tissue evenly expanding and spreading out, it is likely that the common bile duct has not been cannulated, yet the needle is inside the pancreatic capsule. Filling the capsule with Liberase will not result in a high islet yield as the surface area exposed to the enzyme will be low. If this occurs, stop perfusion and reposition the needle.
Once the pancreas has been perfused, it can be removed from the mouse by pulling it free from the points of contact with the intestines, stomach, and spleen. Start by removing the hemostat from the duodenum. Then, use the forceps to lift the duodenum and separate the pancreas from the intestines with a second pair of forceps (FIG.7A-B). This is done by holding the second pair of forceps steady while pulling the intestines out of the abdomen. Next, pull the pancreas free from the top of the stomach and the spleen (FIG.7C-D). Finally, lift the pancreas out of the abdomen and cut it free from the remaining fascia connections (FIG.7E-F).
Place the pancreas in a 50ml conical tube and leave it on ice while working on the other mice. Start a 1 hour timer. For maximal islet yield, place only 1 pancreas in each tube. It is not recommended to leave the perfused pancreata on ice for more than 1 hour, as the Liberase will begin to degrade the tissue. At the end of the hour, promptly move to step 3.0 for all pancreata that were perfused.
Repeat steps 2.3 - 2.6 for the remaining mice or until the 1hour timer started in Step 2.6 has expired.
3. Purifying Islets from the Perfused Pancreata
Before the islets can be purified from the pancreas, the tissue must be digested at 37°C. Group the 50ml tubes bearing the perfused islets in an open bottom rack that fits into the 37°C water bath. Ensure that all caps are well secured and submerge tubes in the 37°C water bath for the precise amount of time pre-determined for the current batch of Liberase (See step 1.4). At the end of the incubation, move the tubes to ice and add RPMI1640 with 10% serum to 20mls per tube. Serum containing media will stop the Liberase digestion.
Disassociate the tissue by shaking the tubes vigorously 40 times in 10 seconds. This step is critical for optimal recovery of islets. Even with optimal Liberase perfusion and tightly calibrated digestion time, islet yield will be low if the tissue is not broken up. The aggressive handling of the samples in this step does not appear to harm the mouse islets and is necessary to free them from exocrine cell clusters.
- To separate the islets from the digested pancreas, complete the following steps.
- Pool digested pancreata so that there are approximately 2.5 pancreata per tube. In doing so, ten tubes will be condensed to 4 tubes with 50mls of media each.
- Centrifuge tubes for 2 minutes at 800RPM and 4°C.
- Pour off the supernatant and resuspend the pellets in 15-25mls media with 10% FBS by vortexing gently (adjust vortex intensity to approximately 50% of maximum) for several seconds.
- Pour the resuspended slurry through the 0.419mm wire mesh and funnel into a fresh 50ml conical tube. This separates out the non-digested tissue, fat and lymph. Rinse the initial tube with an additional 10mls of media containing 10% FBS and pour through the wire mesh. Repeat this process for the remaining tubes.
- Centrifuge the tubes for 2 minutes at 800RPM and 4°C.
- Pour off the supernatant and carefully invert the tubes on a paper towel. Watch to make sure that the islets don't slide off. Wipe the inside of the tubes with a paper towel to remove residual media, being careful not to disturb the cell pellet. Return tubes to an upright position.
- Resuspend the pellets in 5mls of room temperature histopaque1077 by vortexing gently for several seconds. Ensure that the suspension is homogeneous before adding an additional 5mls of histopaque around the inside of the tube. This washes the islets off the wall of the tube.
- Overlay histopaque with 10mls of serum free RPMI, being careful to maintain a sharp interface between the histopaque and media. Add the media by pipetting slowly down the side of the tube at a rate of 1ml per 10 seconds. The media should be on the top, and histopaque on the bottom. Serum affects the density of media and should not be used during this step.
- Spin samples in a Centrifuge equilibrated to 20°C at 2400RPM (900xG) for 20 minutes with very slow acceleration and no braking. This step separates the islets from the exocrine cells. The islets will migrate to the interface between the serum free media and Histopaque while the exocrine cells will form a pellet at the bottom of the tube.
- Collect the islets from the histopaque/media interface with a disposable 10ml serological pipette. Islets will stick to glass, therefore glass pipettes should not be used unless they have been siliconized. Place islets into a fresh 50ml conical tube. Several tubes can be pooled at this point. Fill the tubes to 50ml with serum containing RPMI.
- Centrifuge tubes for 2 minutes at 800RPM and 4°C.
- Pour off the supernatant and resuspend the islets in 50mls of serum containing RPMI by vortexing gently.
- Centrifuge the tubes for 90 seconds at 800RPM and 4°C.
- Pour off the supernatant and repeat wash 1 more time.
- Pour off the supernatant and take tubes to the tissue culture hood. Add pen/strep to RPMI containing 10% FBS to a final concentration of 1% to make the islet culture media. Resuspend the islets in 5mls of this culture media.
- Invert a 0.1mm nylon cell strainer and place it on a 15ml conical tube. Slowly pass the resuspended islet slurry through the strainer. The islets will be retained by the strainer while the exocrine cells pass through into the 15ml conical tube. This step dramatically increases the purity of islets with respect to exocrine cell contamination at the end of the protocol.
- Rinse the 50ml conical tube with an additional 6mls of media and pipette it through the strainer.
- Place 3mls of culture media as a large drop in a 10-cm petri dish. Invert the cell strainer right side up, so that the surface used to capture the islets can be dipped into the media drop. Touching the filter to the media will transfer most of the islets into a non-tissue culture treated petri dish. Using a non-treated petri dish is critical to avoid islet adhesion and dispersal to the surface of the plate. Free floating islets are much more easily picked for separation into experimental groups.
- Pipette an additional 5-7mls of culture media through the strainer into the petri dish to wash any residual islets off of the strainer into the dish.
- Check the islet yield and quality under a 4x objective on an inverted microscope. Swirling the dish in a tight oval will collect the islets in the center of the dish. If the islets look good, they can either be picked immediately for experimental use or separated evenly among 4-6 10cm petri plates (per 10 mice). Culturing too many islets in the same dish causes the islets to become necrotic and degrade. For experiments, 250-300 islets per 6cm dish with 2-3mls of media does not appear to stress the islets.
- There will be some usable islets in the 15ml conical tube that collected the 0.1mm nylon cell strainer flow through. These islets can be recovered following three to four rounds of gravity sedimentation. After approximately 4 minutes of sedimentation, aspirate all but approximately 1ml of media from the tube and refill it to the top with culture media. Cap the tube and invert to mix. Repeating this process several times removes many of the single cell exocrine cells that are slow to sediment, and enriches the heavier aggregates of endocrine cells (islets) that quickly sink. After the final aspiration, resuspend in 7mls of culture media and plate in a fresh petri dish.
4. Working with Isolated Islets
The process of isolation is fairly stressful on the islets; histopaque is toxic, shaking and centrifugation steps induce sheer stress, and the removal from host blood supply as well as in vitro culture can induce hypoxia. We and others have shown that isolation induces beta-cell death 4-5. The effect from these stresses is seen by the sloughing of cells from the periphery of the islet and the darkening and expulsion of the central core of the islet (central necrosis). While the sloughing of cells from the periphery can be tolerated, the central necrosis of the islet disqualifies it for use in experiments. Typically, the larger the islet, the greater the chance that central necrosis will be a problem. Efforts aimed at reducing the islet's post isolation stress response will increase the yield of usable islets.
Therefore, we use a previously reported technique of incubating the islets in a 22-27°C tissue culture incubator for the first 48-72 hours following isolation 6-8. This reduced temperature dampens the islets stress response and smoothes their transition to in vitro culture. If a 22-27°C tissue culture incubator is not available, it is possible to achieve the desired temperature range in a standard tissue culture incubator by turning off the heat control and placing approximately 20lbs of freezer bricks equilibrated to -80°C. This results in approximately 16-24hours of reduced temperature in the incubator. Replace the -80°C freezer bricks as needed. We have not observed an additional benefit from of incubation at this lower temperature longer than 72hrs.
- In addition to the low temperature incubation, we recommend changing the media 24-48 hours after isolation. Secreted factors from stressed and dead islets accumulate in the media following isolation and can promote additional islet stress. To change the media, complete the following steps.
- Transfer the media and islets from the petri dish to a 15ml conical tube using a plastic pipette.
- Wash the plate with an additional 3mls of culture media to collect residual islets.
- Add this additional 3mls to the 15ml conical tube and allow islets to gravity sediment for 5min.
- Aspirate all but 1ml of media from the 15ml conical tube, then resuspend the islets in 4mls of islet culture media.
- Transfer the islets and media to a fresh petri plate. Add an additional 3mls of culture media to the 15ml conical tube to collect any residual islets and add this to the petri plate.
- Change the islet media every three to five days thereafter.
- When picking islets for an experiment, it is not recommended to mix islets from different isolations. The molecular baseline of the islets is influenced by many things, including the amount of time they have been in culture and the isolation stress that varies from prep to prep. To reduce variability, islet experiments must be performed on islets that were isolated at the same time. However, if this is not possible, we strongly suggest pooling all islets into a single group prior to picking them for experiments. To pick islets for experiments, complete the following steps.
- If islets are incubating in more than one dish, pool all islets into a single dish by following steps 4.3.1 through 4.3.5 listed above. If multiple dishes of islets are involved, a 50ml conical tube should be used instead of the 15ml tube. This can reduce variability induced by subtle differences in culture conditions between plates during the post-isolation recovery period.
- During gravity sediment of the islets, set up an inverted microscope equipped with a 4x or 10x objective. Collect a p200 micropipette and a box of sterile p200 tips.
- Transfer the sedimented islets to a single plate and move the plate to the microscope. Swirl the plate to collect the islets into the center. Remove the lid to the plate and use the p200 pipette to pick healthy islets. Healthy islets have smooth boarders and no dark center regions (Figure 8). Try to maintain an even distribution of islet sizes throughout experimental dishes as islet size can alter the response to treatment.
- The number of islets per experimental group can vary based on the needs of the experiment. We estimate that one islet has approximately 1000-2000 cells and will yield 0.3-1μg of protein and 25ng of total RNA. For in vitro assays, a typical experimental group can be between 100 and 250 islets plated in 35mm or 6cm dishes with 1-3ml of media. For transplantation, each recipient mouse will require between 150 and 400 islets depending on the experimental design (See step 5.2.2 and Discussion for more details).
- To minimize the variations introduced by the hand picking process, we use the orifice of the p200-pipette tip as a guide to estimate islet size. Most islets have a diameter approximately half of the orifice diameter, and we count each of them as one standard islet. Any islets bigger or smaller than that, we assign a different islet count accordingly. We collect islets in batches of 20-50 and transfer them to the designated experimental dishes. If the p200-pipette is set at the volume of 180 microliters, one can collect multiple islets (typically more than 50 islets) into one tip. Thus, even though the islets are picked one at a time, one can collect many islets within each p200-tip by controlling the pipetman release mechanism. This facilitates the picking of hundreds or even more than a thousand of islets needed for experiments. We also use this batch-wise operation to help even out the distribution of islets with similar quality and size.
5. Subcapsular Kidney Islet Transplantation
- Induce diabetes in transplant recipient mice.
- Place mice on a 4 to 6 hour fast. Optimal transplant recipients should be between 6 to 10 weeks old and weigh 20 to 25 grams at time of fast. To fast the mice, remove the food from the feeding rack and always place the mice in a fresh cage. This ensures a true fast by separating mice from any residual bits of food that may have previously fallen into their cage. Plan for 80% to 90% of mice to become diabetic.
- Three hours into the fast, prepare the Sodium Citrate Buffer. Weigh out 0.735g of enzyme grade Sodium Citrate (Na Citrate, Mr 294.10 g/mol) and dissolve it in 25mls of sterile deionized water. Adjust the pH to 4.5 using HCl. This buffer should be made fresh for each round of experiments.
- Place 0.05g Streptozotocin (STZ, Mr 265.2 g/mol) into a 1.5ml Eppendorph tube. This amount is enough to induce diabetes in approximately 8 mice. Prepare an adequate number of 1.5ml tubes for the number of mice to be injected. STZ is light sensitive, so cover each tube with aluminum foil.
- After 4 hours of fast, resuspend one tube of STZ with 1ml of freshly made Na Citrate Buffer. Once resuspended, the STZ-Na Citrate solution loses activity within 15 to 20 minutes, so this step should only be done immediately before injecting the mice.
- Inject each mouse intraperitoneally with an appropriate volume of the STZ-Na Citrate solution to achieve a final dose of 190mg/kg mouse. This dose may vary by the strain and age of mouse; therefore it may be necessary to perform a dose optimization if the incidence of diabetes is too low or if too many mice are dying in the 3 to 5 days following injection. Common doses used range from 150mg/kg mouse to 250mg/kg mouse.
- Supply food and water once all mice have been injected.
- Test mice for non-fasting hyperglycemia (>350mg/kg) 2 to 4 days after injection. Mice that demonstrate 3 consecutive days of non-fasting hyperglycemia may be used as transplant recipients.
- Prepare islets for transplantation
- Isolate islets as described above in steps 2.0 through 4.0. If necessary, islets can be isolated on the day of transplantation or before.
- Pick islets into groups for transplantation by placing them into sterile 1.5ml Eppendorph tubes. Keep tubes on ice. The number of islets needed to restore normoglycemia may vary given the conditions of the experiment (islet donor strain, weight and strain of recipient, and any additional treatments given to the recipient) and the person who is picking the islets. The size of the recipient mouse is one major variable that influences minimal islet number, as larger mice will require more islets. We consistently find that 150 to 250 islets only marginally restores normoglycemia while 300 islets or more is curative in an 18 to 20gram C57BL/6 recipient mouse. In these experiments, the islet donor mice were either C57BL/6 or Balb/c.
- Transplant islets to subcapsular kidney pouch
- Assemble the following materials (all surgical instruments must be sterilized): Table 1. Equipment for Transplantation
25μl Hamilton Syringe 6" of PE 50 flexible tubing cut so one side is beveled Gel loading and p200 pipette tips Sterile Eppendorph tubes (1 per recipient) Isoflurane and Isoflurane vaporizer Eppendorph tube rack McPherson-Vannas Scissors (1) Bent arm forceps (2) Hemostats (1) Wound clip with clips 27 gauge needle Flame pulled glass capillary tube probes* Cotton tipped applicators 50ml conical tube of sterile PBS Sutures Electric Shears 70% Ethanol Spray bottle Ball point pen Betadine clear plastic sterile drape - Weigh and tag all diabetic recipient mice. Assign each mouse to an experimental group prior to transplantation so that each group has similarly matched body weights.
- Set up a surgical field within an open front hood equipped with the gas outlet from the Isoflurane vaporizer.
- Anesthetize a recipient mouse with isoflurane and then shave its flank with the electric shears. Shave the mouse outside of the area where the transplants will be performed.
- Return mouse to anesthesia and position it lying perpendicular to and directly over the shaft of a glass rod so that the shaved flank faces up. Spray flank with 70% Ethanol. The mouse is to be prepped with a surgical scrub of alternating betadine and alcohol repeated three times. Drape the mouse with clear sterile drape allowing access to the shaved flank.
- Prepare the islets for transplantation while the mouse is being anesthetized. Do this by completing the following steps.
- Place a sterile gel loading tip into the opening of a 1.5ml Eppendorph tube so that there is a gentle bend which makes a U shape in the narrow end of the tip. The opening of the tip should be facing directly up out of the Eppendorph tube. Do not introduce a kink at any point in the gel loading tip as this will result in shearing of the islets and a reduction in the number of islets that are transplanted.
- Gently remove from ice a tube of islets that was prepared in step 5.2.2. All of the islets should be settled at the bottom of the tube. Using a p200 pipette, gently collect the islets in a minimal volume from the bottom of the tube. Transfer these islets through the large opening of the gel loading tip prepared in step 5.3.6.1. The islets should settle into the narrow length or narrow restriction of the gel loading tip.
- After the islets have sedimented, remove as much of the media from the gel loading tip as possible. This can be accomplished using a fresh gel loading tip attached to a p200 pipette by aspirating the media through the large opening of the islet-bearing gel loading tip.
- Transfer the sedimented islets into the PE 50 flexible tubing (~ 15cm in length). This can be done by first attaching the non-beveled end of the tube to the narrow opening of the islet-bearing gel loading tip before attaching the tip to a p200 pipette. The islets can then be pushed through 60% of the length of the tubing.
- Transfer the islet bearing PE 50 tubing to a 25μl Hamilton syringe. Make sure the syringe plunger has been withdrawn before transferring the tube. Connect the non-beveled edge of the PE 50 tubing to the needle of the syringe.
- Depress the plunger of the syringe until the islets are approximately 0.5cm away from the beveled opening.
- Set the syringe aside so that the islets may settle into a single mass near the beveled opening of the tube while the kidney is being prepared to receive the islets.
- Using your fingers, localize the kidney through the skin of the mouse under anesthesia. The shaft of the ball point pen should be directly underneath the area where the kidney is located and positioned perpendicular to the spinal cord.
- Once the kidney is localized, use the McPherson-Vannas scissors to make a 2cm incision through the dermis directly above it in a direction perpendicular to the spinal cord. This incision will expose the peritoneal wall.
- With the McPherson-Vannas scissors make a 1cm incision through the peritoneal wall in the same direction as the cut through the dermal layer. It is important that the opening created by this incision be smaller than the kidney being exposed.
- Using the shaft of the glass rod as a backstop, force the kidney through the peritoneal opening by pressing down on the adjacent surface. If the opening is slightly smaller than the kidney, the kidney will squeeze through the opening and rest stably on the surface of the mouse without any additional manipulation or contact. This positioning is optimal for the subsequent transplantation steps. If the opening is too big, the kidney will fall back into the peritoneal cavity, making it difficult to complete subsequent steps.
- Wet the surface of the exposed kidney with ice cold sterile PBS using a soaked cotton tipped applicator. Repeat this step every few minutes or as needed to prevent the kidney capsule from drying out.
- Using the 27 gauge needle, make a 0.2cm incision through the kidney capsule across the anterior surface of the kidney moving from the left lateral side towards the right lateral side.
- Using the flame pulled glass capillary tube probe, create a pouch between the kidney capsule and kidney parenchyma. Do so by inserting the probe through the incision made in step 5.3.12 and move it in a posterior direction along the dorsal-lateral surface of the kidney. An ideal probe will have a bend in it that matches the natural arc of this surface of the kidney. This will allow the probe to reach the most posterior end of the kidney without tearing open a large anterior opening. It is important to make this pouch as long and narrow as possible. Short wide pouches are not ideal as they allow islets to escape from the capsule. Long and narrow pouches allow the islets to be deposited toward the posterior end of the kidney with minimal loss from the capsular pouch.
- Retrieve the 25μl Hamilton syringe prepared in step 5.3.6 and move the islets to the very edge of the beveled opening of the flexible tube.
- Insert the beveled edge of the flexible tubing into the subcapsular pouch prepared in step 5.3.13. The beveled edge should be facing up, so that the long edge is in contact with the kidney parenchyma. Move the tube to the most posterior end of the capsule.
- Slowly transfer the islets from the PE 50 tube into the subcapsular pouch by depressing the plunger of the syringe. As the islets fill the pouch slowly back the flexible tube out making more room for the islets to be deposited. Ensure that all islets are transferred from the tube into the pouch. Do not fill the pouch with air or excess liquid.
- After removing the PE 50 tube from the pouch, use the flame pulled glass capillary tube probe to gently pack the islets into the proximal end of the pouch. Do this by sliding the glass probe along the outer surface of the kidney moving in an anterior to posterior direction. Use caution not to pack the islets too tightly as the posterior end of the subcapsular pouch may rupture and release the islets. This step minimizes the loss of islets from the anterior opening of the pouch when the kidney is placed back inside the peritoneal cavity.
- Using a bent arm forceps lift the peritoneal lining adjacent to the opening made for the kidney and then use a PBS soaked cotton tipped applicator to gently push the kidney back into the peritoneal cavity.
- Close the mouse by applying sutures to the peritoneal wall and wound clips to the dermal layer.
- Return the mouse to its cage and repeat steps 5.3.4 through 5.3.19 for the remaining mice. Mice should be kept warm with a heating pad beneath the cage or a heating lamp with the eyes protected until the mouse fully recovers from anesthesia. Animals should be provided with acetominophine in the water (6 mg/mL).
- Check non-fasting blood glucose levels 24hrs later. All mice should be normoglycemic (blood glucose <200mg/dl) at post-operation day (POD) 1.
6. Representative Results
Two main procedures are described in this protocol: (1) islet isolation and (2) islet transplantation under the kidney capsule. In the islet isolation procedure, islet yields typically range between 100-150 islets per mouse depending on the age and strain. The purity of these islets with respect to their separation from the exocrine cells of the pancreas can vary widely between each preparation. However the use of room temperature Histopaque1077 in step 3.3.7 and a 0.1mm nylon filter in step 3.3.17 typically allow for greater than 99% purity of islets. This purity is achieved before any additional hand picking of islets. Representative images of the islets following isolation are shown in Figure 8D. As seen in these images, the islets typically have a rough peripheral border immediately following isolation. However, with time in culture, healthy islets will acquire and maintain a smooth border (Figure 8D). In some islets, a central necrotic region will develop, appearing as a dark area in the core. This necrotic center is often expelled from the islet as captured by time-lapse imaging (Supplemental Video 1). The remaining islets appear hollow in the center (data not shown) and presumably not functional. Thus, we exclude islets with dark center during the hand picking process. Importantly, we found that a low temperature incubation following the isolation procedure (Step 4.2) can dramatically reduce the number of islets that will develop central necrosis in the culture over time.
In the islet transplantation procedure, two steps are critical: chemical induction of diabetes and delivery of islets to the kidney sub-capsule area. For diabetes induction, the goal is to achieve hyperglycemic in greater than 90% of STZ-injected mice. These mice will survive without transplantation for several weeks. The optimal dose of STZ to achieve this goal varies, depending on mouse strains and age, and should be determined by experimentally. We found that a difference as small as 20mg/kg body weight can make a big difference. Thus, a titration with narrow intervals should be carried out. For islet transplantation, a key consideration is the models, which can have four immunological contexts: syngeneic, allogeneic, syngeneic autoimmune, and allogeneic autoimmune. In the syngeneic model, the donor strain is the same as the recipient strain. Thus, the islets would not invoke the adaptive-immune response from the recipients. This allows the researchers to investigate the issues surrounding "primary non-function," a term referring to islet dysfunction and loss due to reasons other than specific immune rejection by the recipients. Primary non-function is thought to be the main reason for islet failure in the early transplantation stage and for the requirement of a large number of islets (≥2 pancreata/patient) to achieve euglycemia. Thus, it is a critical issue for islet transplantation. In this model, a marginal islet mass that does not completely restore normoglycemia should be used and the mice should be examined for blood glucose at the early stage following transplantation, typically between POD 1 to POD 7. In our experience, a marginal mass of islets is typically between 150 and 250 average sized islets per 20 gram recipient using the C57BL/6 strain of mice. With the syngeneic model, one can use genetically or pharmacologically modified islets to test whether a particular modulation affects the therapeutic efficacy of the islets at the early stage after transplantation.
In the allogeneic and autoimmune models, the islet grafts are exposed to a specific immune attack by the host adaptive-immunity, which involves T-lymphocytes, B-lymphocytes, and other immune cells. Adaptive-immunity is a delayed immune response; to assay this response, it is necessary to transplant a saturating number of islets that effectively restores normoglycemia at the early stage after transplantation to minimize the impact of primary non-function. One can then monitor the length of time before the islets are rejected as indicated by the loss of normoglycemia to determine the effects of any modification of islets on graft rejection. In our experience, greater than 300 islets are required per 20 gram mouse and 15 to 21 days are the rejection time for an allogeneic transplant using Balb/c (H-2d) as donors and C57BL/6 (H-2b) as recipients.
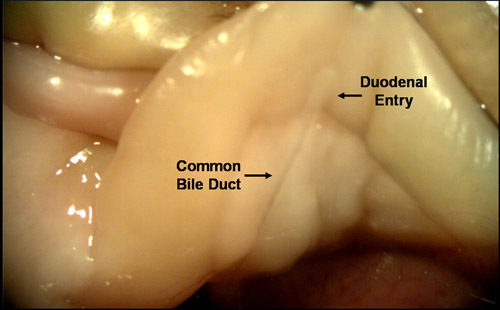 Figure 1. Locating the duodenal opening of the common bile duct. Endogenous digestive enzymes draining from the liver, pancreas and the gall bladder pass through the common bile duct before entering the intestinal tract at the duodenum. The direction of flow through this duct can be reversed if pressure is applied to the system by adding external fluid. This will result in the pancreas becoming full and distended.
Figure 1. Locating the duodenal opening of the common bile duct. Endogenous digestive enzymes draining from the liver, pancreas and the gall bladder pass through the common bile duct before entering the intestinal tract at the duodenum. The direction of flow through this duct can be reversed if pressure is applied to the system by adding external fluid. This will result in the pancreas becoming full and distended.
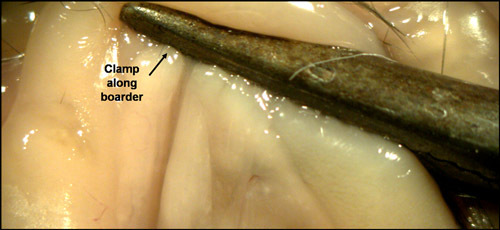 Figure 2. Clamping the duodenal opening of the common bile duct. In order to fill the pancreas with Liberase, it is necessary to block the duodenal opening of the common bile duct. If this is not achieved properly, Liberase will fill the intestines instead of the pancreas.
Figure 2. Clamping the duodenal opening of the common bile duct. In order to fill the pancreas with Liberase, it is necessary to block the duodenal opening of the common bile duct. If this is not achieved properly, Liberase will fill the intestines instead of the pancreas.
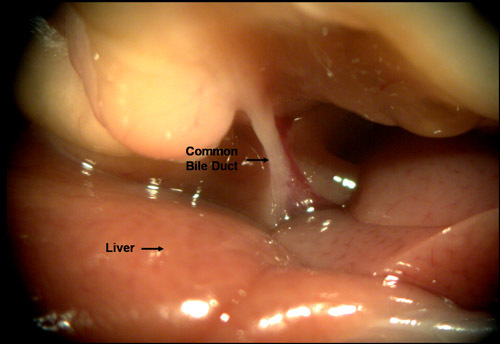 Figure 3. Repositioning the liver against the diaphragm allows visualization of the proximal region of the common bile duct. Cannulation of the common bile duct can be achieved most successfully, if it is attempted near its proximal end (at the V-shaped confluence described in 2.3.1).
Figure 3. Repositioning the liver against the diaphragm allows visualization of the proximal region of the common bile duct. Cannulation of the common bile duct can be achieved most successfully, if it is attempted near its proximal end (at the V-shaped confluence described in 2.3.1).
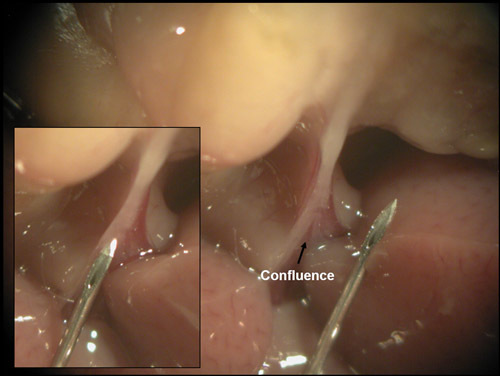 Figure 4. Cannulating the common bile duct by the 'free hand' method. A confluence of the common bile duct becomes visible when the hemostat that is clamped over the duodenum is pulled towards the posterior end of the mouse. Duct cannulation can be achieved by placing the 27 gauge needle at this confluence and driving through it.
Figure 4. Cannulating the common bile duct by the 'free hand' method. A confluence of the common bile duct becomes visible when the hemostat that is clamped over the duodenum is pulled towards the posterior end of the mouse. Duct cannulation can be achieved by placing the 27 gauge needle at this confluence and driving through it.
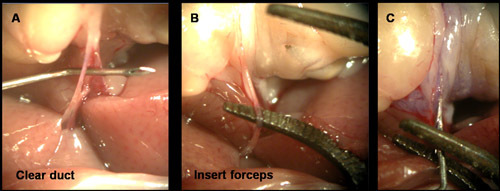 Figure 5. Cannulating the common bile duct by the 'forceps assist' method. As mentioned in the text, sometimes the fascial tissue surrounding the duct makes it difficult to cannulate. (A) In these cases, it is useful to clear the facial layer with the 27 gauge needle. (B) The duct can then be clearly visualized and supported by a pair of bent arm forceps. (C) Using the forceps as a backstop, the 27 gauge needle can be inserted into the duct for delivery of Liberase.
Figure 5. Cannulating the common bile duct by the 'forceps assist' method. As mentioned in the text, sometimes the fascial tissue surrounding the duct makes it difficult to cannulate. (A) In these cases, it is useful to clear the facial layer with the 27 gauge needle. (B) The duct can then be clearly visualized and supported by a pair of bent arm forceps. (C) Using the forceps as a backstop, the 27 gauge needle can be inserted into the duct for delivery of Liberase.
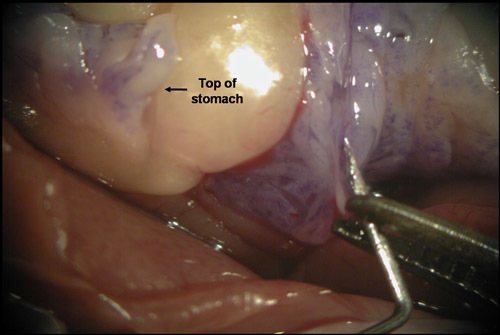 Figure 6. Even expansion of the pancreas indicates optimal needle placement. Given the translucent nature of the duct and surrounding fascial tissues, in some instances it may appear that the duct has been cannulated when in fact it has not. If this occurs and the needle penetrates into the pancreatic capsule, the duodenal region of the pancreas will begin to expand upon delivery of the Liberase. However, the entire pancreas will not be perfused and this will result in low islet yield. To avoid this problem, watch for even expansion of the pancreas by specifically looking for signs of enzyme entering the region of the pancreas attached to the anterior portion of the stomach.
Figure 6. Even expansion of the pancreas indicates optimal needle placement. Given the translucent nature of the duct and surrounding fascial tissues, in some instances it may appear that the duct has been cannulated when in fact it has not. If this occurs and the needle penetrates into the pancreatic capsule, the duodenal region of the pancreas will begin to expand upon delivery of the Liberase. However, the entire pancreas will not be perfused and this will result in low islet yield. To avoid this problem, watch for even expansion of the pancreas by specifically looking for signs of enzyme entering the region of the pancreas attached to the anterior portion of the stomach.
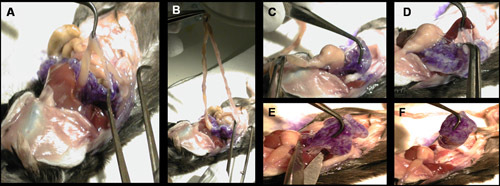 Figure 7. Removing the perfused pancreas. To remove the pancreas from the mouse, several points of contact with the viscera must be broken. (A-B) Separate the pancreas from the intestines. (C) Separate the pancreas from the stomach. (D) Separate the pancreas from the intestines. (E-F) Cut any remaining contacts with the peritoneal cavity.
Figure 7. Removing the perfused pancreas. To remove the pancreas from the mouse, several points of contact with the viscera must be broken. (A-B) Separate the pancreas from the intestines. (C) Separate the pancreas from the stomach. (D) Separate the pancreas from the intestines. (E-F) Cut any remaining contacts with the peritoneal cavity.
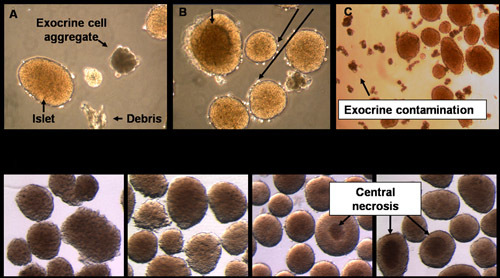 Figure 8. Representative images of islets. The relative purity and quality of the islets can be estimated microscopically upon completion of the isolation procedure. (A) While the goal is to purify islets from the pancreas, exocrine cells and debris are occasionally present. (B) The stress of islet isolation can induce cell death that manifests as dark central regions of the islets and sloughing of cells from the islet periphery. (C) When picking islets for an experiment, avoid picking the contaminating exocrine cells. (D) Islets change in appearance with time in culture. As islets recover from isolation, they acquire a smooth peripheral surface. Occasionally in larger islets a dark central region is visible. These cells stain positive for cell death.
Figure 8. Representative images of islets. The relative purity and quality of the islets can be estimated microscopically upon completion of the isolation procedure. (A) While the goal is to purify islets from the pancreas, exocrine cells and debris are occasionally present. (B) The stress of islet isolation can induce cell death that manifests as dark central regions of the islets and sloughing of cells from the islet periphery. (C) When picking islets for an experiment, avoid picking the contaminating exocrine cells. (D) Islets change in appearance with time in culture. As islets recover from isolation, they acquire a smooth peripheral surface. Occasionally in larger islets a dark central region is visible. These cells stain positive for cell death.
Supplemental Video 1. Time lapse microscopy of post-isolation islets. In this 16hr time lapse video the islets can be seen developing necrotic centers that are ultimately ejected. Please click here for the video.
Discussion
In the sections above, we described detailed steps for isolating and transplanting the mouse pancreatic islets. Here, we briefly highlight the steps that are critical for the success of the procedures.
1. Islet isolation
The key steps are identifying an optimal enzymatic digestion time (1.4), positioning the hemostat clamp over the duodenal opening of the common bile duct (2.2.3), cannulating the common bile duct (2.3), vigorous shaking of the pancreas after the 37°C digest (3.2), and removal of exocrine cell contamination (3.3.7 and 3.3.16). These steps have a dramatic influence on the quality, yield and purity of islets. Perhaps the most technically challenging step is the cannulation of the common bile duct. In many mice, the common bile duct can be difficult to visualize due to surrounding fascia. Therefore, it is common that attempts to cannulate the common bile duct result in penetration of the fascia tissue only. In these instances, the Liberase begins to fill the surrounding connective tissue and does not perfuse the pancreas. If this is observed, stop the flow of Liberase and reposition the needle so that it penetrates into the lumen of the bile duct. With practice, it is possible to quickly identify the optimal position for needle placement and to complete the cannulation without damaging the friable duct.
2. Islet transplantation
The critical steps are the STZ induction of diabetes (5.1) and the efficient delivery of islets to the subcapsular pouch (5.3.14-5.3.16). STZ is a naturally occurring glucose analogue that is selectively toxic to the insulin secreting cells β-cells of islets 9. However, this selective toxicity occurs in a relatively narrow concentration range. This is evidenced by the fact that high doses of STZ can result in rapid (2-3 days) lethality in the absence of hyperglycemia. Therefore, care should be taken to identify the optimal dose of STZ for the strain and age of mice being used before any large scale experiments are initiated. The physical delivery of the islets to the subcapsular pouch is also a key step. To achieve consistent results, it is imperative to minimize the loss of islets during their delivery. This can occur if there are defects or tears in the kidney capsule covering the pouch or if the injected volume is too large. Damage to the capsule can be avoided by gentle and careful manipulation, and by using smooth glass probe to prepare the subcapsular pouch. Additionally, the injection volume can be minimized with careful removal of the supernatant in step 5.3.6.3. If the supernatant is removed down to the level of the sedimented islets, there is typically more than enough room in the subcapsular pouch to place 400 to 500 islets. However, if the injection volume is too large, the islets are more likely to leak out of the pouch during the transplant process, compromising the reproducibility of the experiment.
Within this protocol, it is possible to modify some of the steps and still achieve satisfactory outcomes. These include the following: (a) the use of a different enzyme to digest the pancreas, (b) a different method for delivering the enzyme to the pancreas, (c) the use of a continuous Ficoll-sodium-diatrizoate (FSD) gradient instead of the single density Histopaque1077 step, and (d) islet transplantation to locations other than the kidney subcapsule.
In this protocol, Liberase is used to weaken the cell-cell contacts within the pancreas. Liberase is essentially a mixture of highly purified collagenases that have proven useful in releasing islets from the pancreas. However, less purified collagenases are capable of accomplishing a similar result. Therefore, it is possible to use different commercially available enzyme preparations for digesting the pancreas, as long as the digestion condition is optimized to achieve high islet quality, yield and purity.
It is also possible to digest the pancreas without ductal perfusion of enzyme. Delivery of Liberase through the common bile duct allows for maximal exposure of pancreatic surface area to the enzyme. This results in more even digestion of the pancreas and greater release of intact islets. However, other steps to increase the surface area of the pancreas to the enzyme can be used. For example, mincing the pancreas before incubating it in Liberase or directly injecting Liberase through the pancreatic capsule can be used to isolate islets. However, no other method of Liberase delivery is as effective as perfusion through the common bile duct. Therefore, mincing or direct injection should be reserved as a last resort, if the common bile duct is damaged and not perfusable.
An alternative method for separating the islets from the exocrine cells involves the use of a FSD gradient instead of Histopaque1077 10. The efficient separation of islets from exocrine cells in this protocol is in part due to a density differential between the two cell types. Histopaque1077 has a density of 1.0771 g/ml at 25°C. At this temperature, Histopaque is denser than the islets but less dense than the exocrine cells. Therefore, under the force of centrifugation, Histopaque1077 pellets the exocrine cells while lifting the islets to the surface. In principle, any other substance with a density that that can separate islets from exocrine cells can be used in this step and the FSD gradient is one such substance.
With respect to the location for islets transplantation, the kidney capsule used in this protocol is one of the few possible sites. While the kidney subcapsule is the most commonly reported site of transplantation in mice, other transplantation sites have been found to be effective and they include the hepatic portal vein, subretinal space, testis, epididymal fat pad, spleen, and even pancreas 11-14. Each of these locations has their own advantages and challenges. Therefore, the selection of a site for islet transplantation should be determined by the questions addressed and specific circumstances of the experiments.
It is believed that strategies that can overcome the limitations on islet transplantation will dramatically enhance its therapeutic potential. Therefore, the use of a murine model as detailed by this protocol offers an attractive approach for identifying such strategies.
Disclosures
No conflicts of interest declared.
Acknowledgments
This work is supported by RO1 DK064938 (to T.H.).
References
- Shapiro AM. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl J Med. 2000;343:230–238. doi: 10.1056/NEJM200007273430401. [DOI] [PubMed] [Google Scholar]
- Weir GC, Bonner-Weir S. Islet transplantation as a treatment for diabetes. J Am Optom Assoc. 1998;69:727–732. [PubMed] [Google Scholar]
- Gangemi A. Islet transplantation for brittle type 1 diabetes: the UIC protocol. Am J Transplant. 2008;8:10–1111. doi: 10.1111/j.1600-6143.2008.02234.x. [DOI] [PubMed] [Google Scholar]
- Zmuda E. Deficiency of ATF3, an adaptive-response gene, protects islets and ameliorates inflammation in a syngenic transplantation model. Diabetologia. 2010. Forthcoming. [DOI] [PMC free article] [PubMed]
- Bottino R. Response of Human Islets to Isolation Stress and the Effect of Antioxidant Treatment. Diabetes. 2004;53:2559–2568. doi: 10.2337/diabetes.53.10.2559. [DOI] [PubMed] [Google Scholar]
- Markmann JF. The effect of islet cell culture in vitro at 24 degrees C on graft survival and MHC antigen expression. Transplantation. 1990;49:272–277. doi: 10.1097/00007890-199002000-00009. [DOI] [PubMed] [Google Scholar]
- Terasaka R, Lacy PE, Hauptfeld V, Bucy RP, Davie JM. The effect of cyclosporin-A, low-temperature culture, and anti-Ia antibodies on prevention of rejection of rat islet allografts. Diabetes. 1986;35:83–88. doi: 10.2337/diab.35.1.83. [DOI] [PubMed] [Google Scholar]
- Ricordi C, Lacy PE, Sterbenz K, Davie JM. Low-temperature culture of human islets or in vivo treatment with L3T4 antibody produces a marked prolongation of islet human-to-mouse xenograft survival. Proc Natl Acad Sci U S A. 1987;84:8080–8084. doi: 10.1073/pnas.84.22.8080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnedl WJ, Ferber S, Johnson JH, Newgard CB. STZ transport and cytotoxicity. Specific enhancement in GLUT2-expressing cells. Diabetes. 1994;43:1326–1333. doi: 10.2337/diab.43.11.1326. [DOI] [PubMed] [Google Scholar]
- Eckhard M, Brandhorst D, Brandhorst H, Brendel MD, Bretzel RG. Optimization in osmolality and range of density of a continuous ficoll-sodium-diatrizoate gradient for isopycnic purification of isolated human islets. Transplantation Proceedings. 2004;36:2849–2854. doi: 10.1016/j.transproceed.2004.09.078. [DOI] [PubMed] [Google Scholar]
- Gores PF, Rabe F, Sutherland DE. Prolonged survival of intraportal versus subrenal capsular transplanted islet allografts. Transplantation. 1987;43:747–749. [PubMed] [Google Scholar]
- Nasr IW. Testicular Immune Privilege Promotes Transplantation Tolerance by Altering the Balance between Memory and Regulatory T Cells. J Immunol. 2005;174:6161–6168. doi: 10.4049/jimmunol.174.10.6161. [DOI] [PubMed] [Google Scholar]
- Merani S, Toso C, Emamaullee J, Shapiro AMJ. Optimal implantation site for pancreatic islet transplantation. British Journal of Surgery. 2008;95:1449–1461. doi: 10.1002/bjs.6391. [DOI] [PubMed] [Google Scholar]
- Inoue M, Maeno T, Hatchell DL. Survival of allografted pancreatic islets in the subretinal space in rats. Ophthalmic Res. 2003;35:48–53. doi: 10.1159/000068197. [DOI] [PubMed] [Google Scholar]