

Abstract
Novel chemical entities (NCEs) may be investigated for emetic liability in a range of unpleasant experiments involving retching, vomiting or conditioned taste aversion/food avoidance in sentient animals. We have used a range of compounds with known emetic /aversive properties to examine the possibility of using the social amoeba, Dictyostelium discoideum, for research into identifying and understanding emetic liability, and hence reduce adverse animal experimentation in this area. Twenty eight emetic or taste aversive compounds were employed to investigate the acute (10 min) effect of compounds on Dictyostelium cell behaviour (shape, speed and direction of movement) in a shallow chemotaxic gradient (Dunn chamber). Compound concentrations were chosen based on those previously reported to be emetic or aversive in in vivo studies and results were recorded and quantified by automated image analysis. Dictyostelium cell motility was rapidly and strongly inhibited by four structurally distinct tastants (three bitter tasting compounds - denatonium benzoate, quinine hydrochloride, phenylthiourea, and the pungent constituent of chilli peppers - capsaicin). In addition, stomach irritants (copper chloride and copper sulphate), and a phosphodiesterase IV inhibitor also rapidly blocked movement. A concentration-dependant relationship was established for five of these compounds, showing potency of inhibition as capsaicin (IC50 = 11.9±4.0 µM) > quinine hydrochloride (IC50 = 44.3±6.8 µM) > denatonium benzoate (IC50 = 129±4 µM) > phenylthiourea (IC50 = 366±5 µM) > copper sulphate (IC50 = 1433±3 µM). In contrast, 21 compounds within the cytotoxic and receptor agonist/antagonist classes did not affect cell behaviour. Further analysis of bitter and pungent compounds showed that the effect on cell behaviour was reversible and not cytotoxic, suggesting an uncharacterised molecular mechanism of action for these compounds. These results therefore demonstrate that Dictyostelium has potential as a non-sentient model in the analysis of the molecular effects of tastants, although it has limited utility in identification of emetic agents in general.
Introduction
Emetic research employs a range of animal models, either to identify the emetic liability of a novel chemical entity (NCE) or to characterise mechanisms giving rise to emesis [1]. Common models can be divided into those that have the ability to vomit (e.g. ferret, house musk shrew, dog and cat), and those that lack the emetic reflex (e.g. rats and mice) [2]. In rats, pica, the ingestion of a non-nutritive substance such as kaolin, and conditioned taste aversion/food avoidance (CTA/CFA) are used as an emetic-like readout [3]. Considerable variability in the sensitivity to emetic compounds exists between animal models, due to the multiple pathways available for induction of the reflex, and differences in receptor pharmacology and distribution, and metabolic pathway regulation [4], [5]. This variability therefore makes it difficult to establish a single animal model for emetic research, and encourages a multi-model approach and increasing animal usage [5].
The very nature of emetic research has the potential to cause considerable distress in the subjects, and some emetic compounds (e.g. cisplatin) induce intense retching and vomiting and a protracted emetic response that can last for several days [6]. To reduce the number of animals needed for these adverse tests, Holmes et al. [5] suggested a tiered approach to identify potential emetic liability of NCEs early in compound optimisation. In this approach, a series of individual assays would be performed in order to reduce the final number of compounds tested on sentient models. The first tier in this approach would involve the in silico analysis of novel compounds under investigation, whereby structures of known emetic efficacy are compared with novel compounds under investigation employing previously recorded data from in vivo studies. Secondly, a simple non-sentient model would be used to screen for compounds showing strong effects associated with other emetic compounds. Thirdly, tissue culture experiments would be employed using mammalian cell lines to predict emetic liability, and then finally animal models would be incorporated. This approach could substantially reduce the number of animal experiments by excluding many compounds with potential emetic liability at an earlier stage. This tiered approach requires development of a simple non-sentient model system capable of identifying emetic liability of compounds in a high-throughput type screen.
Dictyostelium is a simple model system, widely used in the analysis of cell signalling, development, and cell behaviour during movement [7]–[10]. The genome of the model has been sequenced [11], identifying a wide range of homologues related to human disease proteins and associated intracellular signalling pathways. Many of these proteins and related pathways are absent in other simple model systems such as Saccharomyces cerevisiae and Schizosaccharomyces pombe [11], [12], suggesting Dictyostelium may have specific advantages over other commonly used non-sentient models.
Dictyostelium is increasingly being used in biomedical research [12]–[14], in for example, the analysis of mitochondrial disease [15], in Alzheimer's disease signaling [16], and in understanding pathways of microbial infection [17]. In many of these studies, and in other more pharmacologically-oriented projects, Dictyostelium has been used to analyse drug-induced changes in behaviour during movement at a cellular level [18]. For example, in the analysis of bipolar disorder drugs valproic acid and lithium [19], [20], for potential chemotherapy research [13], [21], [22], and for the vasodilator nitric oxide [23].
In this paper, we explore the utility of using Dictyostelium as a simple non-sentient model in the tiered approach to reducing animals in testing for emetic liability, as proposed by Holmes et al. [5]. This was investigated by monitoring cell behaviour (speed, shape and direction of movement) following exposure to a range of compounds known to induce emesis, pica or CTA/CFA. The broad categories of compounds investigated (summarised in Table 1) include: tastants (bitter and pungent [“hot”] compounds), cytotoxic anti-cancer agents, selective receptor agonists and antagonists and metal salts. Our results indicate that tastants (both bitter and hot compounds) cause a rapid, pronounced and concentration-dependent effect on cell behaviour, although a range of cytotoxic and receptor agonist/antagonists compounds giving rise to emetic or taste aversive responses had no effect. These results suggest that Dictyostelium may provide a new model for the analysis of bitter and hot compound perception and signalling, although it shows little functionality as a generalised predictor of emetic function for novel chemical entities.
Table 1. Emetic or taste aversive compounds assessed for effects on Dictyostelium behaviour during chemotaxis.
| Generic Target | Target Receptor/ Mechanism of Action | Common Name | Concentration | Effect on Dictyostelium | Species | Dose Range | Reference |
| Receptor Agonist/ Tastant | T2R Receptor ligand | Denatonium Benzoate | 0.05–10 mM | Y | R*, H* | 0.01–10 mM | [36], [39], [63], [72] |
| Phenylthiourea | 0.05–5 mM | Y | H* | 2–5 mM | [63] | ||
| Quinine HCl | 0.05–1 mM | Y | R* | 0.0082–250 mM | [36], [73] | ||
| TRPV1 Receptor Agonist | Capsaicin | 0.01–0.3 mM | Y | S | 0.04–0.4 mg/kg | [28], [74] | |
| Resiniferatoxin | 1 µM, 10 µM | N | S | 0.1–1000 µg/kg | [27], [29], [75], [76] | ||
| Cytotoxic | Cytotoxic/DNA Damage?? | 5-Fluorouracil | 250 µM | N | F,R,S,H | 35–100 mg/kg | [77]–[79] |
| Actinomycin D | 10 µM, 700 µM | N | D,R | 0.13–0.25 mg/kg | [79], [80] | ||
| Cisplatin | 50 µM, 300 µM | N | F,D,R,S, H | 3–20 mg/kg | [6], [29], [80]–[87] | ||
| Cycloheximide | 5 mM | N | F,D | 20 mg/kg | [81] | ||
| Methotrexate | 50 µM, 250 µM | N | S,H | 80 mg/kg | [77], [78] | ||
| Streptozotocin | 1 µM | N | H | 14–27 mg/kg | [88]–[90] | ||
| Vincristine | 1 µM | N | R | 0.1–1 mg/kg | [79] | ||
| ReceptorAntagonist | Extracellular Enzyme Inhibitor | Digoxin | 1 µM | N | C,H | 0.2–0.6 mg/kg | [91], [92] |
| PDEIV Inhibitor | Rolipram | 10 µM, 700 µM | Y | F,D,R,S | 0.5–10 mg/kg | [60], [93]–[95] | |
| SSRI/Transmitter Uptake Inhibitor | Fluoxetine | 6.5 µM | N | S | 60 mg/kg | [96] | |
| Receptor Agonist | 5-HT Receptor Agonist | 5-hydroxytryptamine | 1 µM, 100 µM | N | S | 4–10 mg/kg | [96]–[98] |
| Dopamine Receptor Agonist | Apomorphine HCl | 10 µM, 1 mM | N | F,D,R,H | 0.1–10 mg/kg | [80], [81], [83], [85], [99]–[101] | |
| Ligand Gated Ion Channel Activator | Veratridine HCl | 30 µM, 500 µM | N | D, C | 0.02–0.25 mg/kg | [102]–[104] | |
| Neurokinin Receptor Agonist | Substance P | 1 µM | N | D | 0.03–0.2 mg/kg | [105] | |
| Nicotinic Receptor Agonist | Nicotine | 6 µM, 100 µM | N | F,D,R,S,H | 1.5–20 mg/kg | [29], [75], [87], [97], [106]–[109] | |
| Opioid Receptor Agonist | Loperamide HCl | 1 µM, 100 µM | N | F | 0.5 mg/kg | [85], [110], [111] | |
| Other | CNS depressant | Lithium Chloride | 10 mM | N | R,S,H | 50–200 mg/kg | [109], [112], [113] |
| Enteroendocrine Cell Stimulant | Metformin | 500 µM, 10 mM | N | H* | 1–30 µM | [114], [115] | |
| Free Radical Generator | Pyrogallol | 500 µM, 10 mM | N | S | 128 mg/kg | [27], [116] | |
| Gastric mucosal irritant | Copper Sulphate | 0.16– 5 mM | Y | F,D,R,S, H | 5–120 mg/kg | [29], [85], [97], [109], [117], [118] | |
| Copper Chloride | 1.6 mM | Y | Based on concentratio of copper sulphate: [29], [85], [97], [109], [117], [118] | ||||
| Zinc Sulphate | 1.6 mM | N | H | 1% | [119] | ||
| Prostaglandin | PGF2α | 1 µM, 100 µM | N | F,S | 1–13.5 mg/kg | [117], [120] |
A range of emetic or taste aversive compounds within the categories of tastants, cytotoxic agents, generalised receptor agonists/antagonists, and other compounds were selected for acute exposure to chemotaxing Dictyostelium cells. These compounds have a range of target receptors and/or mechanisms of action, as indicated. Compound concentrations employed in these Dictyostelium experiments (shown here) were derived from the experimental dose range for each compound used in emetic-related experiments in other species: F = Ferret; D = Dog; R = Rat; S = Shrew; C = Cat; H = Human. For in vivo experiments in the rat, the table refers to the dose at which pica was observed, for all other species it refers to the emetic dose. An effect on chemotaxis (defined as a significant change in cell velocity following acute treatment (see Figure 3 and 4)) is represented by Y (Yes), with no effect denoted N (No). * Caused conditioned taste aversion responses in the rat or data derived from in vitro studies.
Results
To investigate the utility of employing Dictyostelium as a model for the study of tastants, cytotoxic agents, receptor agonists/antagonists and other emetic or aversive compounds, we first defined a standard assay. In this assay, Dictyostelium cell behaviour was monitored by time lapse photography every 6 seconds over a 15 min period (under control conditions) ( Figure 1 ) within a chemotactic gradient (moving towards cAMP). Computer-generated outlines of individual cells enabled the quantification of cell velocity, aspect and angle of movement ( Figure 2 and Movie S1). These three measurements encapsulate the complete basic behaviour of moving cells. In addition, an X, Y coordinate plot is provided illustrating the path length and direction of movement of individual cells throughout the recorded period. Under these conditions, cells exhibited stable behaviour that did not significantly change over the 15 min period monitored ( Figure 3 ).
Figure 1. Time-dependent image series of Dictyostelium cells during chemotaxis. A–O:

Cells moving towards a gradient of cAMP (5 µM) over a 15 min period, with images shown for each min. Two cells are indicated (arrows) over the test period. Bar = 10 µm.
Figure 2. Analysis of Dictyostelium cell behaviour.
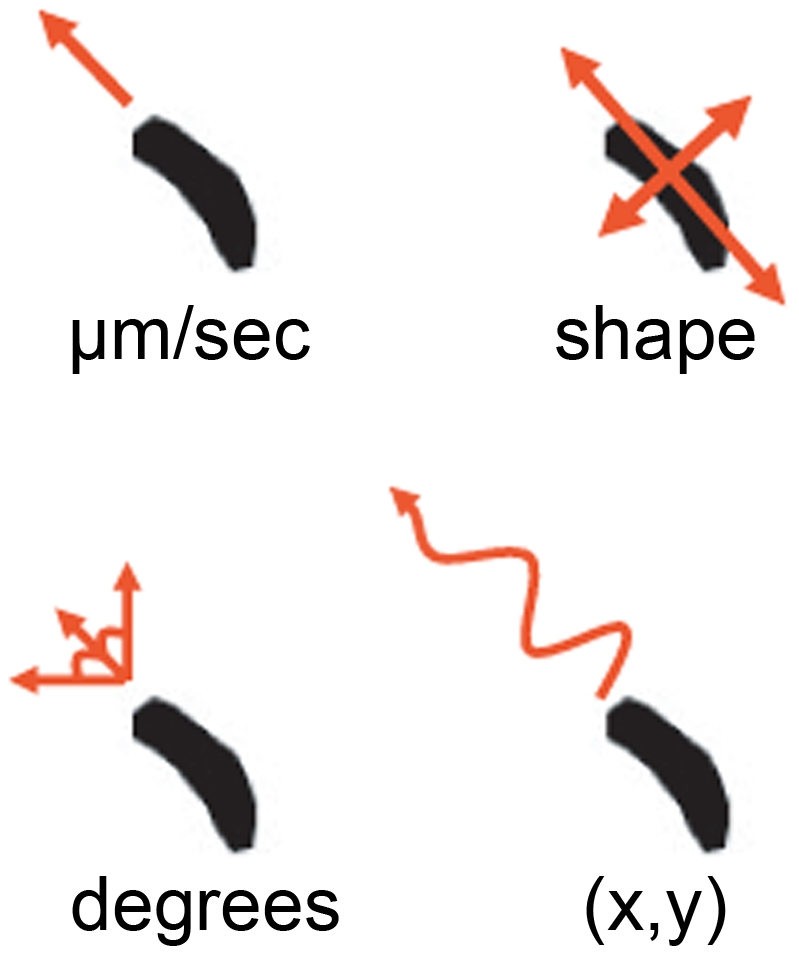
Cells moving under a chemotactic gradient were analysed using ImageProPlus software to determine cell velocity ( µm/min); cell aspect (shape -measured as a ratio between the diameters of cells across each axis, where a value of 1 represents a circle); cell angle (degrees-where cell migration was measured in comparison to the y-axis); and cell tracking (where the co-ordinates of individual cells were illustrated following normalisation to (0,0) at 5 min) in order to illustrate changes in migration before and after compound addition.
Figure 3. Analysis of Dictyostelium cell behaviour over a 15 min period under control conditions.

Mean data representing 85 cell chemotaxis over a 15 min period for A: cell velocity; B: cell aspect; C: cell angular movement; D: cell tracking (where the co-ordinates of individual cells were illustrated following normalisation to (0,0) at 5 min, represented by single lines for 0–5 min (blue) or 5–15 min (dashed red) and cell direction has been adjusted so that cells are moving up the page). Data from A–C is presented as mean of triplicate experiments analysing approximately 30 cells in each.
This standard assay enabled the analysis of compounds with known emetic or aversive responses in a range of species, on Dictyostelium cell behaviour ( Table 1 ). For each compound and concentration, at least triplicate experiments were recorded (monitoring approximately 30 cells each), establishing the behaviour of cells for five min prior to compound addition. Following drug addition, images were then recorded for a further ten min to monitor acute drug effects. The concentrations of compounds used in these tests are based upon concentrations used in vivo (e.g. copper sulphate), plasma concentrations (e.g. cisplatin) or concentrations shown to be active in vitro in mammalian tissues relevant to the emetic reflex (e.g. RTX on neurones, denatonium on intestinal epithelial cells) as shown in Table 1 . A compound was determined to have an effect on cell behaviour if the average cell velocity or aspect changed significantly (P<0.05) between the first five min period (prior to addition of the drug) and the final five min of the assay. Where a substance was without apparent effect at in vitro concentrations, experiments were then repeated at 10 – 200 fold higher concentration ( Table 1 ) to reduce the risk of obtaining a false-negative result.
Of the 28 compounds screened, seven evoked a significant acute effect on Dictyostelium cell behaviour ( Tables 1 and 2 , Figures 4 and 5 ). These were: denatonium benzoate, phenylthiourea, quinine hydrocholoride; copper chloride and sulphate salts; capsaicin; and rolipram. The effect of all of these compounds was a concomitant loss of velocity, cell shape and angular movement. Strength of effect also varied, where for example, addition of 5 mM copper sulphate caused Dictyostelium cells to slowly stop moving and lose shape over a ten min period ( Figure 4 and Movie S2), although still generating a significant decrease in velocity (P = 0.014) and change in aspect (P = 0.047). In contrast, the addition of denatonium benzoate (5 mM) caused an immediate loss in cell velocity and aspect ( Figure 5 ). This variation in time of onset for drug effects is also seen in the X,Y coordinate plots for these compounds ( Figure 3D , 4D and 5D ). Interestingly, a number of compounds that have also been shown to have tastant activity related to those tested here did not inhibit cell behaviour in this assay (e.g. the hot compound resiniferatoxin and the bitter compound cycloheximide).
Table 2. Statistical significance of the concentration-dependent acute reduction in Dictyostelium cell velocity.
| Compound | Concentration (mM) | P-value (Velocity) |
| Capsaicin | 0.01 | NS |
| 0.05 | 0.025 | |
| 0.10 | 0.041 | |
| 0.20 | 0.001 | |
| 0.30 | 0.006 | |
| Copper Chloride | 1.60 | 0.045 |
| Copper Sulphate | 0.16 | NS |
| 0.80 | NS | |
| 1.20 | NS | |
| 1.60 | 0.032 | |
| 2.40 | 0.028 | |
| 5.00 | 0.005 | |
| Denatonium Benzoate | 0.05 | NS |
| 0.50 | 0.015 | |
| 1.00 | 0.037 | |
| 5.00 | 0.021 | |
| 10.0 | 0.06 | |
| Phenylthiourea | 0.05 | NS |
| 0.20 | NS | |
| 0.50 | NS | |
| 1.00 | 0.024 | |
| 2.00 | 0.011 | |
| 5.00 | 0.040 | |
| Quinine Hydrocholoride | 0.05 | NS |
| 0.10 | 0.049 | |
| 0.20 | 0.002 | |
| 0.50 | 0.029 | |
| 1.00 | 0.035 | |
| Rolipram | 0.01 | NS |
| 0.70 | 0.007 |
Concentration range of compounds showing a significant acute effect on Dictyostelium cell velocity between the first 5 and final 5 min of the assay. T-tests performed were 2-tailed paired student t-tests, with ∼30 cells measured in each replicate. NS = not significant.
Figure 4. Analysis of Dictyostelium cell behaviour with addition of 5 mM copper sulphate after 5 Min.

Mean data representing 124 cell chemotaxis over a 15 min period following addition of copper sulphate (5 mM) at 5 min (arrow) for A: cell velocity; B: cell aspect; C: cell angular movement; D: cell tracking (where the co-ordinates of individual cells were illustrated following normalisation to (0,0) at 5 min, represented by single lines for 0–5 min (blue) or 5–15 min (dashed red) and cell direction has been adjusted so that cells are moving up the page). Data from A–C is presented as a mean of triplicate experiments analysing approximately 40 cells in each.
Figure 5. Analysis of Dictyostelium cell behaviour with addition of 5 mM denatonium benzoate after 5 Min.

Mean data representing 89 cells chemotaxis over a 15 min period following addition of denatonium benzoate (5 mM) at 5 min (arrow) for A: cell velocity; B: cell aspect; C: cell angular movement; D: cell tracking (where the co-ordinates of individual cells were illustrated following normalisation to (0,0) at 5 min, represented by single lines for 0–5 min (blue) or 5–15 min (dashed red) and cell direction has been adjusted so that cells are moving up the page). Data from A–C is presented as a mean of triplicate experiments analysing approximately 30 cells in each.
We then investigated the concentration-dependence of this effect for denatonium benzoate, phenylthiourea, quinine hydrochloride, copper sulphate and capsaicin. A relationship between compound concentration and change in velocity was found in all cases, as indicated by the secondary plots ( Figure 6 ). IC50 values calculated from these experiments suggest the ranking of potency to be; capsaicin (IC50 = 11.9±4.0 µM, R2 = 0.78) > quinine hydrochloride (IC50 = 44.3±6.8 µM, R2 = 0.61) > denatonium benzoate (IC50 = 129±4 µM, R2 = 0.65) > phenylthiourea (IC50 = 366±5 µM, R2 = 0.50) > copper sulphate (IC50 = 1433±3 µM, R2 = 0.54).
Figure 6. Concentration-dependent reduction of cell velocity for emetic and taste aversive compounds.

Reduction in cell velocity compared to untreated cells was plotted against log10 concentrations, where compound concentration was chosen from those showing a non-significant effect on cell behaviour; up to a maximum of a 200-fold increase from this value. All data is presented as a mean ± S.E.M of triplicate experiments, comparing mean cell velocity during the first 5 min and final 5 min with increasing concentration.
Since the block in cell behaviour during these experiments may occur through a variety of mechanisms including cell toxicity or death, we continued the analysis of tastants on Dictyostelium by monitoring the reversibility of behaviour effects, a potential role of cell death in this effect, and the results of long-term exposure (during development). To assess the reversibility of tastant action, we analysed the recovery of cells following compound exposure. In these experiments, cell movement was recorded for 4.5 min in the absence of a chemotactic (cAMP) gradient, prior to the addition of each tastant for 4.5 min (using concentrations at eight-fold higher that the IC50 value, representing approximate concentrations at which the concentration-dependence curve begins to plateau ( Figure 6 )). This length of exposure was chosen to show a significant reduction in cell velocity. The buffer containing the tastant was then replaced with fresh buffer (lacking tastant) and cells were observed for a further 26 min ( Figure 7 ). Under control conditions (in the absence of tastant), cell handling gave rise to a small non-significant drop in velocity, which then returned towards the initial velocity by the end of the test period. In the presence of all tastants, cells show an initial significant reduction in velocity upon exposure compared to untreated cells, which is consistent with earlier experiments ( Figure 6 ). However, following the removal of the 3 mM phenylthiourea, 1 mM denatonium benzoate, or 350 µM quinine hydrochloride, cells then increased in velocity, returning towards the rate of movement of untreated cells under control conditions, and showing no significant difference for the last time periods measured ( Figure 7 ). In contrast, cells exposed to 100 µM capsaicin did not recover velocity in the time period measured here (not shown). However, reducing the concentration of capsaicin to 50 µM enabled cells to recover velocity ( Figure 7 ).
Figure 7. Analysis of Dictyostelium cell behaviour recovery post-tastant exposure.

Mean data representing cell velocity during random cell movement over a 35 min period, with average cell velocity measured for 4.5 min under control conditions (at −4.5–0 min), prior to the addition of tastants at 0 min (D), followed by removal of tastants at 4.5 min (W), and the recording of recovery up to 31 min. Tastant concentrations used are: 1 mM denatonium benzoate, 3 mM phenylthiourea, 350 µM quinine hydrocholide, and 50 µM capsaicin. Data is presented as a mean ± S.E.M for each 4.5 min period, with triplicate experiments analysing approximately 30 cells in each. Grey bars indicate the equivalent time periods used in the analysis of cell velocity comparing control and each condition. N/S = non significant, * = P<0.05.
To further analyse cell viability following tastant exposure, we then measured cell survival following 10 and 30 min treatment. These experiments also initially employed tastant concentrations at eight-fold IC50 values ( Figure 6 ) and cell viability was determined using trypan blue staining ( Table 3 ). In the presence of phenylthiourea, denatonium benzoate and quinine hydrochloride, the percentage of surviving cells were above 95% after 10 min of exposure and above 90% after 30 min of exposure, indicating cell death is not the cause of the block in cell behaviour following treatment with these compounds. However, 100 µM capsaicin reduced cell survival to 51% and 57% after 10 and 30 min respectively. We therefore reduced capsaicin concentration to 50 µM, to show a 98% cells survived after both 10 and 30 min of exposure, also indicating that cell death was not the cause of capsaicin-induced block in cell movement at or below this concentration.
Table 3. Analysis of tastants on Dictyostelium cell viability.
| Compound/Exposure | Mean Cell Count (alive) | S.E.M | Mean Cell Count (dead) | S.E.M | Cell Total | Cell Viability (% cells Surviving) | P-value |
| Time (Min) | |||||||
| Control | |||||||
| 10 | 135 | 2.2 | 1 | 0.6 | 136 | 99 | N/A |
| 30 | 135 | 4.3 | 0 | 0 | 135 | 100 | N/A |
| 1 mM DB | |||||||
| 10 | 181 | 16.5 | 7 | 0 | 188 | 96 | 0.006 |
| 30 | 183 | 5.9 | 16 | 3.2 | 199 | 92 | 0.021 |
| 350 µM QHCl | |||||||
| 10 | 139 | 12.1 | 1 | 1 | 140 | 99 | NS |
| 30 | 146 | 2.59 | 3 | 1.2 | 150 | 97 | NS |
| 3 mM PTU | |||||||
| 10 | 128 | 8.7 | 0 | 0 | 128 | 100 | NS |
| 30 | 119 | 7.1 | 4 | 0.9 | 123 | 97 | 0.021 |
| 100 µM Capsaicin | |||||||
| 10 | 87 | 1.5 | 83 | 7.7 | 170 | 51 | 0.002 |
| 30 | 74 | 4.1 | 57 | 1.8 | 131 | 57 | 0.0004 |
| 50 µM Capsaicin | |||||||
| 10 | 174 | 1.3 | 3 | 1.5 | 177 | 98 | NS |
| 30 | 150 | 8.9 | 3 | 0.8 | 153 | 98 | 0.021 |
Chemotactically competent Dictyostelium cells were exposed to compounds at indicated concentrations and cell viability assessed after 10 and 30 min using trypan blue. All experiments were performed in triplicate. N/A = Not applicable NS = Not significant.
Finally, we examined the chronic effect of tastants on Dictyostelium development by exposing cells to each compound for 24 hours during starvation on a nitrocellulose filter ( Figure 8 ). Under control conditions, cells were able to chemotax together to form a mound and ultimately develop into a multicellular fruiting body composed of a spore head held above the substratum by dead, vacuolated stalk cells [13]. Repeating these experiments in the presence of tastants (at eight-fold IC50 values, Figure 6 ) did not inhibit fruiting body formation ( Figure 8 ). Furthermore, development in the presence of phenylthiourea (3 mM), quinine hydrochloride (350 µM) and capsaicin (100 µM) did not alter the general structure of the fruiting body (spore head and stalk), however in the presence of denatonium benzoate (1 mM), development was slowed, with a reduced number of immature fruiting bodies present after 24 hours. This effect was overcome after prolonged incubation (48 hours; data not shown).
Figure 8. Analysis of tastants on Dictyostelium development.

Cells were allowed to develop over 24 hours in the presence of control conditions, 3 mM phenylthiourea, 1 mM denatonium benzoate, 350 µM quinine hydrochloride and 100 µM capsaicin. All images are representative of triplicate experiments indicating cell survival after 24-hours exposure to each compound. Scale bar represents 1 mm.
Discussion
Research into the mechanisms by which diverse compounds induce emesis, and the related phenomena of pica and CTA/CFA, traditionally employs a range of unpleasant experiments on several animal species (primarily ferret, dog and rat), with considerable heterogeneity in dose and response for many compounds between different models [5]. It would thus be of significant advantage to provide early indicators of potential emetic liability during drug development prior to in vivo animal studies. We therefore tested a broad spectrum of emetic and aversive compounds from each major class of emesis-inducing chemical group, for their acute effects on Dictyostelium behaviour during chemotaxis. It was found that a range of structurally discrete tastants (capsaicin – the pungent compound in chilli peppers; and denatonium benzoate, phenylthiourea, and quinine hydrochloride – all bitter tasting compounds), a stomach irritant (copper containing compounds), and a phosphodiesterase IV (PDE4) inhibitor (rolipram) ( Table 1 and 2 ) all caused rapid disruption of cellular behaviour, including a simultaneous decrease in cell velocity and loss of cell shape leading to rounding. Following a block in cell movement, the angle of movement also approached zero degrees. Interestingly, no treatments were found to cause changes in cell direction independent of speed or cell shape. It is worth noting that a range of emetic/aversive compounds that have been previously shown to cause chronic effects on Dictyostelium development, including lithium and cytotoxic compounds such as cisplatin [20], [22], [24], [25], did not acutely affect cell velocity, shape or angular direction in this study. The differences in compound effects may be due to the short exposure time (ten min) used here, in comparison to previous reports that employ treatment times of 1–24 hours.
Capsaicin, a vanilloid, is the active compound found in chillies responsible for causing a burning taste sensation and it has been demonstrated to be an agonist for the transient receptor potential vanilloid-1 (TRPV1) receptor [26]. The TRPV1 receptor has been implicated in the induction of emesis via the acute release of endogenous substance P [27], [28]. Identification of the mechanism of action of Capsaicin in Dictyostelium may provide a novel model for molecular research in this area, and provide further insight into a mechanism of action (through either receptor activation or inhibition, or via intracellular cell signalling pathway regulation). Initial examination of the Dictyostelium proteome failed to find any proteins with significant homology to known human, mouse or worm (C. elegans) TRPV1 proteins (based upon BLAST analysis with TRPV receptors - Table S1), and this therefore raises the possibility of an alternative mechanism of action of pungent tasting compounds in regulating Dictyostelium cell behaviour. Surprisingly, the ultra potent analogue of capsaicin, resiniferatoxin, did not significantly affect Dictyostelium cell behaviour at the concentrations used here, although it is capable of inducing emesis in animal models [28], [29]. It is unclear if structural or physicochemical differences between the two compounds give rise to altered efficacy in regulating the Dictyostelium target.
Bitter taste has been thought to be perceived in humans since a pre-Neanderthal age, providing a mechanism for identifying potentially toxic substances [30]. It has been shown that bitter taste can initiate the sensation of nausea [31]. Response to bitter tasting compounds has also been shown across a wide spectrum of model systems from mammals to frogs, fish, Drosophila, and C. elegans [32]–[38]. Denatonium benzoate, phenylthiourea and quinine hydrochloride are all bitter compounds, and all caused significant and dramatic changes in cell velocity and aspect. All three compounds are proposed T2R receptor ligands involved in this bitter taste detection, and this mechanism functions in emesis in humans at high concentrations [39]–[42]. The common effect observed here for three structurally-independent bitter compounds, at concentrations used in emetic based research ( Table 1 ), suggests a T2R receptor-like mechanism of action in Dictyostelium. However, BLAST analysis of the Dictyostelium proteome using 25 different human T2R receptors (with these proteins sharing a 30–70% homology [[43]–[45]); 24 mouse [43], six insect (Drosophila; NCBI 36094, 117484, 38935, 117498, 117349, 117492) [34], three worm (C. elegans; NCBI 178326, 177117, 188314) [32], [37] and three candidate fish (zebrafish; NCBI 664690, 553134, 798975) [35], [46] bitter receptors, again did not identify any Dictyostelium proteins showing significant homology within its genome (See Table S2). Furthermore, other related receptors such as the mammalian TRPM5 receptor (NCBI 29850), also associated with bitter taste detection [47], [48], did not have recognisable Dictyostelium homologues (based on BLAST analysis – see Table S3). Although the lack of recognised bitter receptors in Dictyostelium (as defined by protein sequence homology to other species) suggests a potential novel mechanism for the detection of bitter compounds, it must be noted that receptors for other taste-related compounds have been found in other models that lack homology to established tastant receptors (e.g. sweet receptors in Drosophila [49]). Identifying the mechanism (or molecular target) of bitter taste perception in Dictyostelium may thus provide a novel mechanism of action for bitter tasting compounds, leading to the subsequent analysis of this mechanism in humans.
Copper compounds are essential in the diet but at higher concentrations can causes gastrointestinal upsets, which include nausea, vomiting, cramps and diarrhoea [50], [51]. This has lead to the formation of regulatory guidelines for copper levels in drinking water [50], [51]. Analysis of the molecular mechanism of this effect in mammalian systems suggests the gastro-duodenal luminal concentration of copper sulphate is the key to emesis induction, thus implicating a gut mucosal-triggered reaction, and this is supported by neurophysiological studies [52]–[56]. However, the molecular emetic mechanism of copper in the gut remains unclear. Several studies have analysed a role for copper in Dictyostelium [57], and these have suggested that cells are highly resistant to copper through high cellular export [57]. The molecular mechanism of copper may be due to an inhibition of ATP-dependent ion currents, controlled by P2X receptors [58], [59].
Phosphodiesterase IV (PDE4) inhibitors are proposed for use as anti-inflammatory agents (e.g. in asthma) but may cause nausea and vomiting as side-effects [60]. Phosphodiesterases are responsible for degrading cAMP and cGMP in Dictyostelium [61], and there is one potential PDE4 homologue in Dictyostelium that shares a 33–35% identity with the 4 human isoforms A–D. Inhibition of phosphodiesterases would be expected to elevate extracellular cAMP levels, leading to saturation of cAMP receptors in chemotaxis, and thus the observed inhibition of cell behaviour shown in our assay. Rolipram did not exert an effect on Dictyostelium cell behaviour at low concentrations (10 µM), but blocked cell movement at higher concentrations (700 µM). The high concentration required for this effect may thus reflect a non-specific action of Rolipram on Dictyostelium phosphodiesterases [62].
The acute block in Dictyostelium cell behaviour caused by tastants may occur through a range of mechanisms including interaction with unknown receptor(s) or other molecular target(s), or through basic cytotoxicological mechanisms. To investigate these potential toxicological mechanisms, we carried out a range of short- and long-term exposure experiments on Dictyostelium. Using a concentration derived from the IC50 value for each compound (eight fold higher), we initially showed that all cells treated with bitter compounds recovered from acute (4.5 min) exposure ( Figure 7 ). This suggests that inhibition of cell behaviour by these tastants is not through a cytotoxicological mechanism. This was then confirmed by measuring cell death following 10 and 30 min exposure, since these compounds had no significant effect on cell viability ( Table 3 ). In contrast 100 µM capsaicin treatment did not enable cell recovery over 26 min, and caused a large reduction in cell viability following 10 min exposure. This effect however is likely to be maximal, since increased exposure (30 min) to capsaicin did not further reduce cell viability. However, reduction of capsaicin concentration to 4-fold over IC50 values enabled cells to recover velocity and did not reduce cell viability. These combined experiments suggest that tastants examined here do not function through an acute toxicological mechanism to block Dictyostelium cell behaviour.
To extend these toxicological assays for the analysis of longer exposure periods, we also examined the role of tastants on Dictyostelium development ( Figure 8 ). In these experiments, cells were exposed to tastants (again at eight-fold IC50 values) for 24 hours, and the ability to develop into mature fruiting bodies was monitored as previously described [10]–[12]. Long term exposure to all compounds did not block fruiting body formation, clearly indicating that these compounds are not lethal to Dictyostelium at high concentrations for extended exposure. Phenylthiourea, quinine and capsaicin exposure also did not alter fruiting body morphology, whereas denatonium benzoate slowed development, with immature fruiting bodies present after 24 hours, that later developed into mature structures. These combined cytotoxicological and developmental experiments suggest that bitter and hot compounds do not block Dictyostelium behaviour through toxic or irreversible mechanisms.
All pharmacological studies must consider drug concentrations, to differentiate between potential target-specific and non-specific effects. In mammalian experimental systems, bitter tasting compounds have been shown to cause effects on mouse intestinal STC-1 [63], [64] as well transfected HEK-293 cells [65] at similar concentrations to those used in our experiments (up to 10 mM and 1 mM respectively), suggesting that Dictyostelium is as sensitive as other models for detecting bitter taste. Similarly, capsaicin has also been used in human-based taste experiments at concentrations shown here to affect Dictyostelium behaviour [66], [67] (100 µM).
Since the molecular mechanisms of these compounds in chemotactic cell behaviour remains unknown, it is not possible to infer a commonly targeted signalling pathway. However, in mammalian systems, T2R receptor signalling is regulated by a TRPV1-like receptor, TRPM5, thus both ‘bitter’ and ‘hot’ compounds share a common signalling pathway [68]. Although Dictyostelium does not contain proteins with high amino acid sequence similarity to either of these receptors, further investigation will be necessary to determine if Dictyostelium's ability to detect these compounds involves a common signalling pathway.
An important limitation of this work is that primary assay used in this investigation only monitors the acute effects of test substances (within 10 min of exposure), thus any delayed effect would not be observed. For example, cytotoxic agents (e.g. cisplatin) used in anti-cancer treatments cause DNA damage, and this effect may not lead to significant changes in cell behaviour within the ten min response time recorded. These compounds do, however, give rise to a chronic block in development following longer exposure [22].
Conclusions
A broad range of emetic and aversive compounds within the categories of tastants (e.g. bitter and hot compounds), cytotoxic agents, or generalised receptor agonists/antagonists were tested to determine if Dictyostelium cell behaviour could be used to investigate the molecular mechanisms of these compounds. We show that Dictyostelium provides a limited model for emetic or aversive compound identification. However, Dictyostelium may enable an exciting new avenue for research into the molecular mechanisms of bitter and hot compounds, since these compounds have a rapid and strong effect on behaviour, the compounds have an uncharacterised molecular mechanism of action, and we have demonstrated the compounds are unlikely to affect cell behaviour via toxicological means. Further investigation into the molecular mechanism of tastants on Dictyostelium may thus provide novel mechanism(s) of bitter and hot compound action.
Materials and Methods
Chemicals
The following chemicals were obtained from Sigma Aldrich Co. Ltd (Dorset, UK), and are provided with catalogue numbers: 5-fluorouracil (2,4-Dihydroxy-5-fluoropyrimidine; F6627), actinomycin D (2-Amino-(N,N)-1-bis(hexadecahydro-6,13-diisopropyl-2, 5, 9-trimethyl-1,4,7,11,14-pentaoxo-1H-pyrrolo[2], [1]-[1], [4], [7], [10], [13] oxatetraazacyclohexadecin-10-yl)-4,6-dimethyl-3-oxo-3H-phenoxazine-1,9-dicarboxamide; A1410), capsaicin (8-Methyl-N-vanillyl-trans-6-nonenamide; M2028), cisplatin (cis-Dichlorodiammine platinum(II); 479306), copper chloride (203149), copper sulphate (cupric sulphate pentahydrate; C8027), cycloheximide (3-[2-(3,5-Dimethyl-2-oxocyclohexyl)-2-hydroxyethyl] glutarimide; C7698), denatonium benzoate (N,N-Diethyl-N-[(2,6-dimethylphenyl carbamoyl) methyl] benzyl ammonium benzoate; D5765), digoxin (12β-Hydroxydigitoxin; D6003), fluoxetine ((±)-N-Methyl-γ-[4-(trifluoromethyl)phenoxy]benzenepropanamine hydrochloride; F132), lithium chloride (L9650), loperamide hydrochloride (4-(p-Chlorophenyl)-4-hydroxy-N,N-dimethyl-α,α-diphenyl-1-piperidinebutyramide hydrochloride; L4762), metformin (1,1-Dimethylbiguanide hydrochloride; 04635), methotrexate (4-Amino-10-methylfolic acid hydrate), nicotine (3-(1-methylpyrrolidin-2-yl)pyridine; M4010), PGF2α ((5Z,9α,11α,13E,15S)-9,11,15-Trihydroxyprosta-5,13-dienoic acid tris salt; P0424), phenylthiourea (1-Phenyl-2-thiourea; P7629), pyrogallol (1,2,3-Trihydroxybenzene; P0381), quinine hydrochloride ((R)-[(2S,4R,5R)-5- ethenyl-1 -azabicyclo [2.2.2]octan-2-yl]- (6-methoxyquinolin-4-yl) methanol dihydrate hydrochloride; Q1125), resiniferatoxin (4-Hydroxy-3-methoxy- [(2S,3aR,3bS,6aR,9aR,9bR,10R,11aR)- 3a,3b,6,6a,9a, 10,11,11a-octahydro-6a-hydroxy-8, 10-dimethyl-11a-(1-methylethenyl)-7-oxo-2-(phenylmethyl)-7H-2,9b -epoxyazuleno[5,4-e]-1,3-benzodioxol-5-yl] benzeneacetate; R8756), rolipram (4-[3-(Cyclopentyloxy)-4-methoxyphenyl]-2-pyrrolidinone; R6520), streptozocin (N-(Methylnitrosocarbamoyl)-α-D-glucosamine; S0130), veratridine (3-Veratroylveracevine; V5754), vincristine (22-Oxovincaleukoblastine sulfate salt; V8388) and zinc sulphate monohydrate (96495). The following compound (with catalogue number) were obtained from Tocris Bioscience Ltd: (Bristol, UK) 5-hydroxytryptamine (3-(2-Aminoethyl)-1H-indol-5-ol hydrochloride; 3547), apomorphine hydrochloride (R(–)-10,11-Dihydroxyaporphine; 2073) and substance P ((2S)-2-[[(2S)-1-[(2S)-6-amino-2-[[(2S)-1-[(2S)-2-amino-5-(diaminomethylideneamino) pentanoyl]pyrrolidine-2-carbonyl]amino]hexanoyl]pyrrolidine-2-carbonyl]amino]-N-[(2S)-5-amino-1-[[(2S)-1-[[(2S)-1-[[2-[[(2S)-1-[[(2S)-1-amino-4-methylsulfanyl-1-oxobutan-2-yl] amino]-4-methyl-1-oxopentan-2-yl]amino]-2-oxoethyl]amino]-1-oxo-3-phenylpropan-2-yl] amino]-1-oxo-3sphenylpropan-2-yl]amino]-1,5-dioxopentan-2-yl]pentanediamide; 1156). All compounds were dissolved in dimethylsulfoxide (DMSO) (5-fluorouracil, actinomycin D, cycloheximide, digoxin, fluoxetine, rolipram, quinine hydrochloride, capsaicin and resiniferatoxin) or phosphate buffer (16.5 mM KH2PO4, 3.8 mM K2HPO4, pH 6.2) (lithium chloride, cisplatin, streptozotocin, vincristine, metformin, pyrogallol, copper sulphate, copper chloride, zinc sulphate, substance P, nicotine, loperamide hydrochloride, PGF2α, denatonium benzoate, phenylthiourea), apart from 5-hydroxytryptamine, apomorphine hydrochloride (dissolved in 0.9% ascorbic acid); methotrexate and veratridine (dissolved in 0.1% sodium hydroxide).
Cell Behaviour Assay
To prepare Dictyostelium cells (Ax2) for behaviour analysis experiments, cells were grown in shaking suspension in Axenic medium (Formedium Co. Ltd, Norfolk, UK), washed and resuspended in phosphate buffer at 1.7×106 cells/ml. Cells were then pulsed for 5 hours with 30 nΜ cyclic adenosine monophosphate (cAMP) (Sigma Co. Ltd, Dorset, UK) at 6 min intervals whilst shaking at 120 rpm. Cells were then washed in phosphate buffer, resuspended at 1×107 cells/ml, and used in a Dunn chamber (Hawksley, Sussex, UK) assay [69], migrating toward 5 µM cAMP. A stable chemotactic gradient was allowed to form over a 30 min period, prior to recording cell shape and position using an Olympus IX71 microscope at 40x magnification with a QImaging RetigaExi Fast1394 digital camera. Cell images were recorded every 6 seconds over a 15 min period, with the initial 5 min period recorded prior to addition of test compounds (within a 10 µL aliquot diluted in 5 µM cAMP) to the outer well of the Dunn chamber. Subsequent images were recorded over the following ten min period for each compound, and at each concentration, with a minimum of three independent experiments for each drug/concentration and an average cell number of ∼30 cells quantified per experiment. Cell recordings were prepared in the second quadrant of the Dunn chamber, enabling cell angular movement to be recorder at around −50 degrees. Solvent only controls were carried out for all experiments to ensure readouts were based upon compounds listed, with for example, no effect of DMSO shown at 0.6%– the highest concentration used in the experiments described here.
Dictyostelium Recovery Following Tastant Exposure
Dictyostelium cells (Ax2) were pulsed as described above, re-suspended at 1.7×105 cells/mL, and 250 µL aliquots of cells were added to Lab-Tek 8-well chambered coverglass wells (Thermo Fisher, Leicestershire, UK) and allowed to adhere for 45 min. Cell movement was recorded as above at intervals of 18 seconds for a total of 35 min (in the absence of a chemotactic gradient). Cells were allowed to establish a base-line velocity for 4.5 min, prior to the addition of 10 µL of tastant to give indicated final concentrations. Following 4.5 min tastant exposure, cell buffer containing tastants was aspirated from the chamber and replaced with 250 µL phosphate buffer (over a 30 second period), and cells were monitored for a further 25.5 min. All experiments were performed in a minimum of triplicate individual assays, at each compound concentration.
Cell Viability Assay
Dictyostelium cells (Ax2) were pulsed as described, re-suspended at 2.5×105 cells/mL, exposed to tastants (at indicated concentrations) for 7 or 27 min and then stained with 0.4% trypan blue solution (final concentration 0.067%) for 3 min prior to live counting. Dead cells were identified as a distinctive blue colour since live cells did not change colour. Experiments were performed in triplicate.
Development Assay
Dictyostelium development assays were performed in triplicate experiments as previously described [13], [70], [71].
Data Analysis and Statistics
Changes in cell velocity, aspect (the ratio between the major and minor axes of an elliptical shape such as a cell) and angular movement ( Figure 2 ) were monitored for every cell within each of the 600 frames recorded over the 15 min period and analysed by ImagePro software (Media Cybernetics, Buckinghamshire, UK). Compound effects were compared using the mean velocity and aspect of cells between the first 5 min and the final 5 min and significance was determined using a two-tailed paired student t-test (P≤0.05). Angle was also measured throughout in order to observe any changes in the direction of cell movement. The relative co-ordinates of cells (X,Y) were also mapped and represented in a line-tracking plot whereby the co-ordinates of cells at 5 min (when the compound was added) were normalised to (0,0).
In quantifying velocity, concentration-related drug response was calculated by subtracting the mean velocity of cell movement in the final five min from the mean velocity in the first 5 min for each compound at each concentration. These were plotted against drug concentration (log10) to quantify the change in velocity with drug concentration. A non-linear three parameter log 10 concentration response curve was fitted by GraphPad Prism (GraphPad Software Inc v5.02, San Diego, USA) using a least squares fit. Using GraphPad, the subsequent concentration at half maximal compound inhibition (IC50) was calculated well as the R2 value in order to display the accuracy of the curve fit.
Cell velocity was quantified in the Dictyostelium recovery experiments as described above. Significance was determined using unpaired one-tailed student t-tests (based on a unidirectional drop in velocity as observed in Dictyostelium cell behaviour assay) by comparing mean velocity of control cells and equivalent mean velocities at each compound concentration. Tastant dependent effects were determined by comparing 0–4.5 min and 27-31 min periods (grey bar) between control and tastant treated cells to assess initial decrease in cell velocity and recovery of velocity ( Figure 7 ).
Significance was determined in cell viability assays by comparing the percentage cell viability between control conditions and each compound concentration using paired 2-tailed student t-tests.
Supporting Information
Dictyostelium chemotaxis was monitored by time-lapse photography to record cells moving within a Dunn Chamber towards the chemo-attractant, cAMP, across the screen from left to right. Images were taken every 6 seconds over a 15 minute period. Computer generated cell outlines enables average cell velocity, shape and direction of movement to be quantified.
(AVI)
Dictyostelium chemotaxis was monitored by time-lapse photography to record cells moving within a Dunn Chamber towards the chemo-attractant, cAMP, across the screen from bottom right to top left. Images were taken every 6 seconds over a 15 minute period. Computer generated cell outlines enables average cell velocity, shape and direction of movement to be quantified. Cell movement was recorded over a 5 minute period, prior to the addition a range of emetic or aversive compounds (indicated by a black screen flash), and cell behaviour was recorded for a further 10 minutes. In the movie shown here, the stomach irritant copper sulphate (5 mM ) was added, causing Dictyostelium cells to slowly stop moving and lose shape over the ten min period tested.
(AVI)
Homology search results (BLAST analysis) of the Dictyostelium genome for proteins showing amino acid similarity to TRPV receptors from multiple species. Potential homologues are defined by an E-value of less than 1.00E-40 [11], thus Dictyostelium does not contain proteins showing significant sequence similarity to be considered as homologues.
(DOCX)
Homology search results (BLAST analysis) of the Dictyostelium genome for proteins showing amino acid similarity to known bitter receptors from multiple species. Potential homologues are defined by an E-value of less than 1.00E-40 [11], thus Dictyostelium does not contain proteins showing significant sequence similarity to be considered as homologues. N/A = Not applicable.
(DOCX)
Homology search results (BLAST analysis) of the Dictyostelium genome for proteins showing amino acid similarity to TRPM5 receptors from human and mouse. Potential homologues are defined by an E-value of less than 1.00E-40 [11], thus Dictyostelium does not contain proteins showing significant sequence similarity to be considered as homologues. N/A = Not applicable.
(DOCX)
Acknowledgments
Advice on statistics was thankfully received from Fiona Reid, St George's University of London. Thanks to Media Cybernetics UK (David Wiles and James McKechnie) for help with data analysis.
Footnotes
Competing Interests: The authors have declared that no competing interests exist.
Funding: University Federation for Animal Welfare (UFAW) summer studentship to JM and UFAW PhD studentship to SR, a South Western Academic Network (SWan) small equipment grant to RSBW and PLRA, and a Wellcome Trust Project Grant 082640 and National Centre for Replacement Refinement and Reduction (NC3R)s Project grant G0900775 to RSBW. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Stern, RM, Koch KL, Andrews, PLR Nausea-Mechanisms and Management. The functions, identification and measurement of nausea and related behaviour in animals. Oxford University Press. 2011;171-239 [Google Scholar]
- 2.Sanger GJ, Holbrook JD, Andrews PL. The translational value of rodent gastrointestinal functions: a cautionary tale. Trends Pharmacol Sci. 2011;7:402–409. doi: 10.1016/j.tips.2011.03.009. [DOI] [PubMed] [Google Scholar]
- 3.Liu YL, Malik N, Sanger GJ, Friedman MI, Andrews PL. Pica--a model of nausea? Species differences in response to cisplatin. Physiol Behav. 2005;85:271–277. doi: 10.1016/j.physbeh.2005.04.009. [DOI] [PubMed] [Google Scholar]
- 4.Andrews PL, Horn CC. Signals for nausea and emesis: Implications for models of upper gastrointestinal diseases. Auton Neurosci. 2006;125:100–115. doi: 10.1016/j.autneu.2006.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Holmes AM, Rudd JA, Tattersall FD, Aziz Q, Andrews PL. Opportunities for the replacement of animals in the study of nausea and vomiting. Br J Pharmacol. 2009;157:865–880. doi: 10.1111/j.1476-5381.2009.00176.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Percie du Sert N, Rudd JA, Apfel CC, Andrews PL. Cisplatin-induced emesis: systematic review and meta-analysis of the ferret model and the effects of 5-HT(3) receptor antagonists. Cancer Chemother Pharmacol. 2010;3:667–686. doi: 10.1007/s00280-010-1339-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Manahan CL, Iglesias PA, Long Y, Devreotes PN. Chemoattractant signaling in Dictyostelium discoideum. Annu Rev Cell Dev Biol. 2004;20:223–253. doi: 10.1146/annurev.cellbio.20.011303.132633. [DOI] [PubMed] [Google Scholar]
- 8.Bagorda A, Mihaylov VA, Parent CA. Chemotaxis: moving forward and holding on to the past. Thromb Haemost. 2006;95:12–21. [PubMed] [Google Scholar]
- 9.McMains VC, Liao XH, Kimmel AR. Oscillatory signaling and network responses during the development of Dictyostelium discoideum. Ageing Res Rev. 2008;7:234–248. doi: 10.1016/j.arr.2008.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.King JS, Insall RH. Chemotaxis: finding the way forward with Dictyostelium. Trends Cell Biol. 2009;19:523–530. doi: 10.1016/j.tcb.2009.07.004. [DOI] [PubMed] [Google Scholar]
- 11.Eichinger L, Pachebat JA, Glockner G, Rajandream MA, Sucgang R, et al. The genome of the social amoeba Dictyostelium discoideum. Nature. 2005;435:43–57. doi: 10.1038/nature03481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Williams RS, Boeckeler K, Graf R, Muller-Taubenberger A, Li Z, et al. Towards a molecular understanding of human diseases using Dictyostelium discoideum. Trends Mol Med. 2006;12:415–424. doi: 10.1016/j.molmed.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 13.Boeckeler K, Williams RS. Vol. 1. John Wiley & Sons Ltd.; 2007. Dicytostelium as a Biomedical Model. In: Encyclopedia of Life Sciences. Available: http://www.els.net/WileyCDA/ElsArticle/refId-a0006038.html. Accessed 2007 Oct. [Google Scholar]
- 14.Pakes NK, Jayasinghe SN, Williams RSB. Bio-electrospraying and aerodynamically assisted bio-jetting the model eukaryotic Dictyostelium discoideum: assessing stress and developmental competency post treatment. J R Soc Interface. 2011;61:1185–1191. doi: 10.1098/rsif.2010.0696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Francione LM, Annesley SJ, Carilla-Latorre S, Escalante R, Fisher PR. The Dictyostelium model for mitochondrial disease. Semin Cell Dev Biol. 2010;22:120–130. doi: 10.1016/j.semcdb.2010.11.004. [DOI] [PubMed] [Google Scholar]
- 16.McMains VC, Myre M, Kreppel L, Kimmel AR. Dictyostelium possesses highly diverged presenilin/{gamma}-secretase that regulates growth and cell-fate specification and can accurately process human APP: a system for functional studies of the presenilin/{gamma}-secretase complex. Dis Model Mech. 2010;9-10:581–594. doi: 10.1242/dmm.004457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Francione L, Smith PK, Accari SL, Taylor PE, Bokko PB, et al. Legionella pneumophila multiplication is enhanced by chronic AMPK signalling in mitochondrially diseased Dictyostelium cells. Dis Model Mech. 2009;2:479–489. doi: 10.1242/dmm.003319. [DOI] [PubMed] [Google Scholar]
- 18.Jin T, Xu X, Fang J, Isik N, Yan J, et al. How human leukocytes track down and destroy pathogens: lessons learned from the model organism Dictyostelium discoideum. Immunol Res. 2009;43:118–127. doi: 10.1007/s12026-008-8056-7. [DOI] [PubMed] [Google Scholar]
- 19.Xu X, Muller-Taubenberger A, Adley KE, Pawolleck N, Lee VW, et al. Attenuation of phospholipid signaling provides a novel mechanism for the action of valproic acid. Eukaryot Cell. 2007;6:899–906. doi: 10.1128/EC.00104-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.King JS, Teo R, Ryves J, Reddy JV, Peters O, et al. The mood stabiliser lithium suppresses PIP3 signalling in Dictyostelium and human cells. Dis Model Mech. 2009;2:306–312. doi: 10.1242/dmm.001271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Misty R, Martinez R, Ali H, Steimle PA. Naringenin is a novel inhibitor of Dictyostelium cell proliferation and cell migration. Biochem Biophys Res Commun. 2006;345:516–522. doi: 10.1016/j.bbrc.2006.04.047. [DOI] [PubMed] [Google Scholar]
- 22.Li G, Alexander H, Schneider N, Alexander S. Molecular basis for resistance to the anticancer drug cisplatin in Dictyostelium. Microbiology. 2000;146:2219–2227. doi: 10.1099/00221287-146-9-2219. [DOI] [PubMed] [Google Scholar]
- 23.Tao Y, Howlett A, Klein C. Nitric oxide inhibits the initiation of cAMP pulsing in D. discoideum without altering receptor-activated adenylate cyclase. Cell Signal. 1996;8:26–34. doi: 10.1016/0898-6568(95)02011-x. [DOI] [PubMed] [Google Scholar]
- 24.Min J, Traynor D, Stegner AL, Zhang L, Hanigan MH, et al. Sphingosine kinase regulates the sensitivity of Dictyostelium discoideum cells to the anticancer drug cisplatin. Eukaryot Cell. 2005;4:178–189. doi: 10.1128/EC.4.1.178-189.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.King J, Keim M, Teo R, Weening KE, Kapur M, et al. Genetic control of lithium sensitivity and regulation of inositol biosynthetic genes. PLoS One. 2010;5:e11151. doi: 10.1371/journal.pone.0011151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Alexander SP, Mathie A, Peters JA. British Journal Pharmacology. S; 2008. Guide to Receptors and Channels (GRAC).209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Andrews PL, Dovey E, Hockaday J, Hoyle CH, Woods AJ, et al. The development of the emetic reflex in the house musk shrew, Suncus murinus. Brain Res Dev Brain Res. 2000;121:29–34. doi: 10.1016/s0165-3806(00)00022-5. [DOI] [PubMed] [Google Scholar]
- 28.Smith JE, Paton JF, Andrews PL. An arterially perfused decerebrate preparation of Suncus murinus (house musk shrew) for the study of emesis and swallowing. Exp Physiol. 2002;87:563–574. doi: 10.1113/eph8702424. [DOI] [PubMed] [Google Scholar]
- 29.Andrews PL, Okada F, Woods AJ, Hagiwara H, Kakaimoto S, et al. The emetic and anti-emetic effects of the capsaicin analogue resiniferatoxin in Suncus murinus, the house musk shrew. Br J Pharmacol. 2000;130:1247–1254. doi: 10.1038/sj.bjp.0703428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lalueza-Fox C, Gigli E, de la Rasilla M, Fortea J, Rosas A. Bitter taste perception in Neanderthals through the analysis of the TAS2R38 gene. Biol Lett. 2009;5:809–811. doi: 10.1098/rsbl.2009.0532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Peyrot des GC, Beauchamp GK, Stern RM, Koch KL, Breslin PA. Bitter taste induces nausea. Curr Biol. 2011;21:R247–R248. doi: 10.1016/j.cub.2011.02.028. [DOI] [PubMed] [Google Scholar]
- 32.Hilliard MA, Bergamasco C, Arbucci S, Plasterk RH, Bazzicalupo P. Worms taste bitter: ASH neurons, QUI-1, GPA-3 and ODR-3 mediate quinine avoidance in Caenorhabditis elegans. EMBO J. 2004;23:1101–1111. doi: 10.1038/sj.emboj.7600107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Shi P, Zhang J. Contrasting modes of evolution between vertebrate sweet/umami receptor genes and bitter receptor genes. Mol Biol Evol. 2006;23:292–300. doi: 10.1093/molbev/msj028. [DOI] [PubMed] [Google Scholar]
- 34.McBride CS, Arguello JR, O'Meara BC. Five Drosophila genomes reveal nonneutral evolution and the signature of host specialization in the chemoreceptor superfamily. Genetics. 2007;177:1395–1416. doi: 10.1534/genetics.107.078683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Oike H, Nagai T, Furuyama A, Okada S, Aihara Y, et al. Characterization of ligands for fish taste receptors. J Neurosci. 2007;27:5584–5592. doi: 10.1523/JNEUROSCI.0651-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tordoff MG, Alarcon LK, Lawler MP. Preferences of 14 rat strains for 17 taste compounds. Physiol Behav. 2008;95:308–332. doi: 10.1016/j.physbeh.2008.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ezak MJ, Hong E, Chaparro-Garcia A, Ferkey DM. Caenorhabditis elegans TRPV channels function in a modality-specific pathway to regulate response to aberrant sensory signaling. Genetics. 2010;185:233–244. doi: 10.1534/genetics.110.115188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Isono K, Morita H. Molecular and cellular designs of insect taste receptor system. Front Cell Neurosci. 2010;4:20. doi: 10.3389/fncel.2010.00020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sibert JR, Frude N. Bittering agents in the prevention of accidental poisoning: children's reactions to denatonium benzoate (Bitrex). Arch Emerg Med. 1991;8:1–7. doi: 10.1136/emj.8.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tajima T, Watanabe N, Kogawa Y, Takiguchi N, Kato J, et al. Chemotaxis of the nematode Caenorhabditis elegans toward cycloheximide and quinine hydrochloride. J Biosci Bioeng. 2001;91:322–324. doi: 10.1263/jbb.91.322. [DOI] [PubMed] [Google Scholar]
- 41.Clapp TR, Trubey KR, Vandenbeuch A, Stone LM, Margolskee RF, et al. Tonic activity of G-alpha gustducin regulates taste cell responsivity. FEBS Lett. 2008;582:3783–3787. doi: 10.1016/j.febslet.2008.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tepper BJ, White EA, Koelliker Y, Lanzara C, d'Adamo P, et al. Genetic variation in taste sensitivity to 6-n-propylthiouracil and its relationship to taste perception and food selection. Ann N Y Acad Sci. 2009;1170:126–139. doi: 10.1111/j.1749-6632.2009.03916.x. [DOI] [PubMed] [Google Scholar]
- 43.Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJ, et al. A novel family of mammalian taste receptors. Cell. 2000;100:693–702. doi: 10.1016/s0092-8674(00)80705-9. [DOI] [PubMed] [Google Scholar]
- 44.Chandrashekar J, Mueller KL, Hoon MA, Adler E, Feng L, et al. T2Rs function as bitter taste receptors. Cell. 2000;100:703–711. doi: 10.1016/s0092-8674(00)80706-0. [DOI] [PubMed] [Google Scholar]
- 45.Pronin AN, Tang H, Connor J, Keung W. Identification of ligands for two human bitter T2R receptors. Chem Senses. 2004;29:583–593. doi: 10.1093/chemse/bjh064. [DOI] [PubMed] [Google Scholar]
- 46.Ishimaru Y, Okada S, Naito H, Nagai T, Yasuoka A, et al. Two families of candidate taste receptors in fishes. Mech Dev. 2005;122:1310–1321. doi: 10.1016/j.mod.2005.07.005. [DOI] [PubMed] [Google Scholar]
- 47.Liman ER. TRPM5 and taste transduction. Handb Exp Pharmacol. 2007;179:287–298. doi: 10.1007/978-3-540-34891-7_17. [DOI] [PubMed] [Google Scholar]
- 48.Perez CA, Huang L, Rong M, Kozak JA, Preuss AK, et al. A transient receptor potential channel expressed in taste receptor cells. Nat Neurosci. 2002;5:1169–1176. doi: 10.1038/nn952. [DOI] [PubMed] [Google Scholar]
- 49.Gordesky-Gold B, Rivers N, Ahmed OM, Breslin PA. Drosophila melanogaster prefers compounds perceived sweet by humans. Chem Senses. 2008;33:301–309. doi: 10.1093/chemse/bjm088. [DOI] [PubMed] [Google Scholar]
- 50.Araya M, McGoldrick MC, Klevay LM, Strain JJ, Robson P, et al. Determination of an acute no-observed-adverse-effect level (NOAEL) for copper in water. Regul Toxicol Pharmacol. 2001;34:137–145. doi: 10.1006/rtph.2001.1492. [DOI] [PubMed] [Google Scholar]
- 51.Olivares M, Araya M, Pizarro F, Uauy R. Nausea threshold in apparently healthy individuals who drink fluids containing graded concentrations of copper. Regul Toxicol Pharmacol. 2001;33:271–275. doi: 10.1006/rtph.2000.1440. [DOI] [PubMed] [Google Scholar]
- 52.Kayashima N, Iwasaki M, Hayama T. Site of emetic action of oral copper sulfate in dogs. (II) Importance of lower duodenum. Jpn J Pharmacol. 1978;28:797–801. doi: 10.1254/jjp.28.797. [DOI] [PubMed] [Google Scholar]
- 53.Kayashima N, Tanaka M, Iwasaki M, Hayama T. Site of emetic action of oral copper sulfate in dogs. (I) Thresholds of various portions of gastrointestinal tract to locally applied copper sulfate. Jpn J Pharmacol. 1978;28:775–781. doi: 10.1254/jjp.28.775. [DOI] [PubMed] [Google Scholar]
- 54.Lee KY, Park HJ, Chey WY. Studies on mechanism of retching and vomiting in dogs. Effect of peripheral dopamine blocker on myoelectric changes in antrum and upper small intestine. Dig Dis Sci. 1985;30:22–28. doi: 10.1007/BF01318366. [DOI] [PubMed] [Google Scholar]
- 55.Niijima A, Jiang ZY, Daunton NG, Fox RA. Effect of copper sulphate on the rate of afferent discharge in the gastric branch of the vagus nerve in the rat. Neurosci Lett. 1987;80:71–74. doi: 10.1016/0304-3940(87)90497-6. [DOI] [PubMed] [Google Scholar]
- 56.Makale MT, King GL. Surgical and pharmacological dissociation of cardiovascular and emetic responses to intragastric CuSO4. Am J Physiol. 1992;263:R284–R291. doi: 10.1152/ajpregu.1992.263.2.R284. [DOI] [PubMed] [Google Scholar]
- 57.Burlando B, Evangelisti V, Dondero F, Pons G, Camakaris J, et al. Occurrence of Cu-ATPase in Dictyostelium: possible role in resistance to copper. Biochem Biophys Res Commun. 2002;291:476–483. doi: 10.1006/bbrc.2002.6463. [DOI] [PubMed] [Google Scholar]
- 58.Bavan S, Straub VA, Blaxter ML, Ennion SJ. A P2X receptor from the tardigrade species Hypsibius dujardini with fast kinetics and sensitivity to zinc and copper. BMC Evol Biol. 2009;9:17. doi: 10.1186/1471-2148-9-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ludlow MJ, Durai L, Ennion SJ. Functional characterization of intracellular Dictyostelium discoideum P2X receptors. J Biol Chem. 2009;284:35227–35239. doi: 10.1074/jbc.M109.045674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Aoki M, Fukunaga M, Sugimoto T, Hirano Y, Kobayashi M, et al. Studies on mechanisms of low emetogenicity of YM976, a novel phosphodiesterase type 4 inhibitor. J Pharmacol Exp Ther. 2001;298:1142–1149. [PubMed] [Google Scholar]
- 61.Bader S, Kortholt A, van Haastert PJ. Seven Dictyostelium discoideum phosphodiesterases degrade three pools of cAMP and cGMP. Biochem J. 2007;402:153–161. doi: 10.1042/BJ20061153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rascon A, Viloria ME, De-Chiara L, Dubra ME. Characterization of cyclic AMP phosphodiesterases in Leishmania mexicana and purification of a soluble form. Mol Biochem Parasitol. 2000;106:283–292. doi: 10.1016/s0166-6851(99)00224-8. [DOI] [PubMed] [Google Scholar]
- 63.Chen MC, Wu SV, Reeve, Rozengurt E. Bitter stimuli induce Ca2+ signaling and CCK release in enteroendocrine STC-1 cells: role of L-type voltage-sensitive Ca2+ channels. Am J Physiol Cell Physiol. 2006;291:C726–C739. doi: 10.1152/ajpcell.00003.2006. [DOI] [PubMed] [Google Scholar]
- 64.Masuho I, Tateyama M, Saitoh O. Characterization of bitter taste responses of intestinal STC-1 cells. Chem Senses. 2005;30:281–290. doi: 10.1093/chemse/bji022. [DOI] [PubMed] [Google Scholar]
- 65.Talavera K, Yasumatsu K, Yoshida R, Margolskee RF, Voets T, et al. The taste transduction channel TRPM5 is a locus for bitter-sweet taste interactions. FASEB J. 2008;22:1343–1355. doi: 10.1096/fj.07-9591com. [DOI] [PubMed] [Google Scholar]
- 66.Green BG, Hayes JE. Capsaicin as a probe of the relationship between bitter taste and chemesthesis. Physiol Behav. 2003;79:811–821. doi: 10.1016/s0031-9384(03)00213-0. [DOI] [PubMed] [Google Scholar]
- 67.Green BG, varez-Reeves M, George P, Akirav C. Chemesthesis and taste: evidence of independent processing of sensation intensity. Physiol Behav. 2005;86:526–537. doi: 10.1016/j.physbeh.2005.08.038. [DOI] [PubMed] [Google Scholar]
- 68.Devantier HR, Long DJ, Brennan FX, Carlucci SA, Hendrix C, et al. Quantitative assessment of TRPM5-dependent oral aversiveness of pharmaceuticals using a mouse brief-access taste aversion assay. Behav Pharmacol. 2008;19:673–682. doi: 10.1097/FBP.0b013e3283123cd6. [DOI] [PubMed] [Google Scholar]
- 69.Zicha D, Dunn GA, Brown AF. A new direct-viewing chemotaxis chamber. J Cell Sci. 1991;99:769–775. doi: 10.1242/jcs.99.4.769. [DOI] [PubMed] [Google Scholar]
- 70.Williams RSB, Cheng L, Mudge AW, Harwood AJ. A common mechanism of action for three mood-stabilizing drugs. Nature. 2002;417:292–295. doi: 10.1038/417292a. [DOI] [PubMed] [Google Scholar]
- 71.Terbach N, Shah R, Kelemen R, Klein PS, Gordienko D, et al. Identifying an uptake mechanism for the antiepileptic and bipolar disorder treatment valproic acid using the simple biomedical model Dictyostelium. J Cell Sci. 2011;124:2267–2276. doi: 10.1242/jcs.084285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Glendinning JI, Yiin YM, Ackroff K, Sclafani A. Intragastric infusion of denatonium conditions flavor aversions and delays gastric emptying in rodents. Physiol Behav. 2008;93:757–765. doi: 10.1016/j.physbeh.2007.11.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Scott TR, Giza BK. A measure of taste intensity discrimination in the rat through conditioned taste aversions. Physiol Behav. 1987;41:315–320. doi: 10.1016/0031-9384(87)90394-5. [DOI] [PubMed] [Google Scholar]
- 74.Rudd JA, Wai MK. Genital grooming and emesis induced by vanilloids in Suncus murinus, the house musk shrew. Eur J Pharmacol. 2001;422:185–195. doi: 10.1016/s0014-2999(01)01041-x. [DOI] [PubMed] [Google Scholar]
- 75.Andrews PL, Torii Y, Saito H, Matsuki N. The pharmacology of the emetic response to upper gastrointestinal tract stimulation in Suncus murinus. Eur J Pharmacol. 1996;307:305–313. doi: 10.1016/0014-2999(96)00275-0. [DOI] [PubMed] [Google Scholar]
- 76.Cheng FH, Andrews PL, Moreaux B, Ngan MP, Rudd JA, et al. Evaluation of the anti-emetic potential of anti-migraine drugs to prevent resiniferatoxin-induced emesis in Suncus murinus (house musk shrew). Eur J Pharmacol. 2005;508:231–238. doi: 10.1016/j.ejphar.2004.12.022. [DOI] [PubMed] [Google Scholar]
- 77.Matsuki N, Ueno S, Kaji T, Ishihara A, Wang CH, et al. Emesis induced by cancer chemotherapeutic agents in the Suncus murinus: a new experimental model. Jpn J Pharmacol. 1988;48:303–306. doi: 10.1254/jjp.48.303. [DOI] [PubMed] [Google Scholar]
- 78.Pollera CF, Nardi M, Marolla P, Pinnaro P, Terzoli E, et al. Effective control of CMF-related emesis with high-dose dexamethasone: results of a double-blind crossover trial with metoclopramide and placebo. Am J Clin Oncol. 1989;12:524–529. doi: 10.1097/00000421-198912000-00014. [DOI] [PubMed] [Google Scholar]
- 79.Yamamoto K, Nakai M, Nohara K, Yamatodani A. The anti-cancer drug-induced pica in rats is related to their clinical emetogenic potential. Eur J Pharmacol. 2007;554:34–39. doi: 10.1016/j.ejphar.2006.09.058. [DOI] [PubMed] [Google Scholar]
- 80.Eglen RM, Lee CH, Smith WL, Johnson LG, Whiting RL, et al. RS 42358-197, a novel and potent 5-HT3 receptor antagonist, in vitro and in vivo. J Pharmacol Exp Ther. 1993;266:535–543. [PubMed] [Google Scholar]
- 81.Andrews PL, Davis CJ, Bingham S, Davidson HI, Hawthorn J, et al. The abdominal visceral innervation and the emetic reflex: pathways, pharmacology, and plasticity. Can J Physiol Pharmacol. 1990;68:325–345. doi: 10.1139/y90-047. [DOI] [PubMed] [Google Scholar]
- 82.Fukui H, Yamamoto M, Ando T, Sasaki S, Sato S. Increase in serotonin levels in the dog ileum and blood by cisplatin as measured by microdialysis. Neuropharmacology. 1993;32:959–968. doi: 10.1016/0028-3908(93)90060-g. [DOI] [PubMed] [Google Scholar]
- 83.Knox AP, Strominger NL, Battles AH, Carpenter DO. Behavioral studies of emetic sensitivity in the ferret. Brain Res Bull. 1993;31:477–484. doi: 10.1016/0361-9230(93)90112-o. [DOI] [PubMed] [Google Scholar]
- 84.Kris MG, Cubeddu LX, Gralla RJ, Cupissol D, Tyson LB, et al. Are more antiemetic trials with a placebo necessary? Report of patient data from randomized trials of placebo antiemetics with cisplatin. Cancer. 1996;78:2193–2198. doi: 10.1002/(sici)1097-0142(19961115)78:10<2193::aid-cncr22>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 85.Nakayama H, Yamakuni H, Nakayama A, Maeda Y, Imazumi K, et al. Diphenidol has no actual broad antiemetic activity in dogs and ferrets. J Pharmacol Sci. 2004;96:301–306. doi: 10.1254/jphs.fpj04035x. [DOI] [PubMed] [Google Scholar]
- 86.Horn CC, De Jonghe BC, Matyas K, Norgren R. Chemotherapy-induced kaolin intake is increased by lesion of the lateral parabrachial nucleus of the rat. Am J Physiol Regul Integr Comp Physiol. 2009;297:R1375–R1382. doi: 10.1152/ajpregu.00284.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Parker LA Limebeer CL, Rock EM, Litt DL, Kwiatkowska M, et al. The FAAH inhibitor URB-597 interferes with cisplatin- and nicotine-induced vomiting in the Suncus murinus (house musk shrew). Physiol Behav. 2009;97:121–124. doi: 10.1016/j.physbeh.2009.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Carter, SK, Bakowski MT, Hellman K. Chemotherapy of Cancer. John Wiley and Sons. 1981;pp169-174 [Google Scholar]
- 89.Borison HL, McCarthy LE. Neuropharmacology of chemotherapy-induced emesis. Drugs. 1983;25(Suppl 1):8–17. doi: 10.2165/00003495-198300251-00003. [DOI] [PubMed] [Google Scholar]
- 90.Seymour MT. Emesis in anti-cancer therapy - mechanisms and treatment In: Andrews PLR and Sanger GJ. The pharmacokinetics and pharmacodynamics of chemotherapeutic agents. Hall Medical. 1993;9-44 [Google Scholar]
- 91.Gold H, Greiner T, Cattell M, Modell W, Gluck J, et al. Difference in the relation of cardiac to emetic actions in oral and parenteral digitalization. Am J Med. 1952;13:124–144. doi: 10.1016/0002-9343(52)90152-6. [DOI] [PubMed] [Google Scholar]
- 92.Parsons JA, Summers RJ. Cat assay for the emetic action of digitalis and related glycosides (digitoxin, digoxin, lanatoside C, ouabain and calactin). Br J Pharmacol. 1971;42:143–152. doi: 10.1111/j.1476-5381.1971.tb07094.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Heaslip RJ, Evans DY. Emetic, central nervous system, and pulmonary activities of rolipram in the dog. Eur J Pharmacol. 1995;286:281–290. doi: 10.1016/0014-2999(95)00457-2. [DOI] [PubMed] [Google Scholar]
- 94.Hirose R, Manabe H, Nonaka H, Yanagawa K, Akuta K, et al. Correlation between emetic effect of phosphodiesterase 4 inhibitors and their occupation of the high-affinity rolipram binding site in Suncus murinus brain. Eur J Pharmacol. 2007;573:93–99. doi: 10.1016/j.ejphar.2007.06.045. [DOI] [PubMed] [Google Scholar]
- 95.Davis TG, Peterson JJ, Kou JP, Capper-Spudich EA, Ball D, et al. The identification of a novel phosphodiesterase 4 inhibitor, 1-ethyl-5-{5-[(4-methyl-1-piperazinyl)methyl]-1,3,4-oxadiazol-2-yl}-N-(tet rahydro-2H-pyran-4-yl)-1H-pyrazolo[3,4-b]pyridin-4-amine (EPPA-1), with improved therapeutic index using pica feeding in rats as a measure of emetogenicity. J Pharmacol Exp Ther. 2009;330:922–931. doi: 10.1124/jpet.109.152454. [DOI] [PubMed] [Google Scholar]
- 96.Fujiwara-Sawada M, Imanishi T, Yoshida A, Baba J. Possible involvement of peripheral serotonin 5-HT3 receptors in fluvoxamine-induced emesis in Suncus murinus. J Pharm Pharmacol. 2003;55:271–274. doi: 10.1211/002235702496. [DOI] [PubMed] [Google Scholar]
- 97.Torii Y, Saito H, Matsuki N. Selective blockade of cytotoxic drug-induced emesis by 5-HT3 receptor antagonists in Suncus murinus. Jpn J Pharmacol. 1991;55:107–113. doi: 10.1254/jjp.55.107. [DOI] [PubMed] [Google Scholar]
- 98.Javid FA, Naylor RJ. The effect of serotonin and serotonin receptor antagonists on motion sickness in Suncus murinus. Pharmacol Biochem Behav. 2002;73:979–989. doi: 10.1016/s0091-3057(02)00955-3. [DOI] [PubMed] [Google Scholar]
- 99.Andrews PL, Kovacs M, Watson JW. The anti-emetic action of the neurokinin(1) receptor antagonist CP-99,994 does not require the presence of the area postrema in the dog. Neurosci Lett. 2001;314:102–104. doi: 10.1016/s0304-3940(01)02269-8. [DOI] [PubMed] [Google Scholar]
- 100.Horn CC, Richardson EJ, Andrews PL, Friedman MI. Differential effects on gastrointestinal and hepatic vagal afferent fibers in the rat by the anti-cancer agent cisplatin. Auton Neurosci. 2004;115:74–81. doi: 10.1016/j.autneu.2004.08.011. [DOI] [PubMed] [Google Scholar]
- 101.Osinski MA, Uchic ME, Seifert T, Shaughnessy TK, Miller LN, et al. Dopamine D2, but not D4, receptor agonists are emetogenic in ferrets. Pharmacol Biochem Behav. 2005;81:211–219. doi: 10.1016/j.pbb.2005.03.012. [DOI] [PubMed] [Google Scholar]
- 102.Borison HL, Fairbanks VF. Mechanism of veratrum-induced emesis in the cat. J Pharmacol Exp Ther. 1952;105:317–325. [PubMed] [Google Scholar]
- 103.Swiss ED. The emetic properties of veratrum derivatives. J Pharmacol Exp Ther. 1952;104:76–86. [PubMed] [Google Scholar]
- 104.Bobkov IG. On the role of nodular ganglia of the vagus nerve in the emetic response to aconitine and veratrine. Fiziol Zh SSSR Im I M Sechenova. 1964;50:187–192. [PubMed] [Google Scholar]
- 105.Andrews, PL, Rudd JA. Handbook of Experimental Pharmacology In: Holzer, P. The Role of Tachykinins and the Tachykinin NK1 Receptor in Nausea and Emesis. Springer Verlag. 2004;359-440 [Google Scholar]
- 106.Laffan RJ, Borison HL. Emetic action of nicotine and lobeline. J Pharmacol Exp Ther. 1957;121:468–476. [PubMed] [Google Scholar]
- 107.Jorenby DE, Smith SS, Fiore MC, Hurt RD, Offord KP, et al. Varying nicotine patch dose and type of smoking cessation counseling. JAMA. 1995;274:1347–1352. [PubMed] [Google Scholar]
- 108.Saito R, Takano Y, Kamiya HO. Roles of substance P and NK(1) receptor in the brainstem in the development of emesis. J Pharmacol Sci. 2003;91:87–94. doi: 10.1254/jphs.91.87. [DOI] [PubMed] [Google Scholar]
- 109.Yamamoto K, Ngan MP, Takeda N, Yamatodani A, Rudd JA. Differential activity of drugs to induce emesis and pica behavior in Suncus murinus (house musk shrew) and rats. Physiol Behav. 2004;83:151–156. doi: 10.1016/j.physbeh.2004.08.006. [DOI] [PubMed] [Google Scholar]
- 110.Bhandari P, Bingham S, Andrews PL. The neuropharmacology of loperamide-induced emesis in the ferret: the role of the area postrema, vagus, opiate and 5-HT3 receptors. Neuropharmacology. 1992;31:735–742. doi: 10.1016/0028-3908(92)90034-m. [DOI] [PubMed] [Google Scholar]
- 111.Andrews PL, Bhandari P. Resinferatoxin, an ultrapotent capsaicin analogue, has anti-emetic properties in the ferret. Neuropharmacology. 1993;32:799–806. doi: 10.1016/0028-3908(93)90189-a. [DOI] [PubMed] [Google Scholar]
- 112.Karniol IG, Dalton J, Lader MH. Acute and chronic effects of lithium chloride on physiological and psychological measures in normals. Psychopharmacology (Berl) 1978;57:289–294. doi: 10.1007/BF00426753. [DOI] [PubMed] [Google Scholar]
- 113.Seeley RJ, Blake K, Rushing PA, Benoit S, Eng J, et al. The role of CNS glucagon-like peptide-1 (7-36) amide receptors in mediating the visceral illness effects of lithium chloride. J Neurosci. 2000;20:1616–1621. doi: 10.1523/JNEUROSCI.20-04-01616.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Cubeddu LX, Bonisch H, Gothert M, Molderings G, Racke K, et al. Effects of metformin on intestinal 5-hydroxytryptamine (5-HT) release and on 5-HT3 receptors. Naunyn Schmiedebergs Arch Pharmacol. 2000;361:85–91. doi: 10.1007/s002109900152. [DOI] [PubMed] [Google Scholar]
- 115.Hoffmann IS, Roa M, Torrico F, Cubeddu LX. Ondansetron and metformin-induced gastrointestinal side effects. Am J Ther. 2003;10:447–451. doi: 10.1097/00045391-200311000-00012. [DOI] [PubMed] [Google Scholar]
- 116.Torii Y, Saito H, Matsuki N. Induction of emesis in Suncus murinus by pyrogallol, a generator of free radicals. Br J Pharmacol. 1994;111:431–434. doi: 10.1111/j.1476-5381.1994.tb14753.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kan KK, Jones RL, Ngan MP, Rudd JA. Action of prostanoids on the emetic reflex of Suncus murinus (the house musk shrew). Eur J Pharmacol. 2003;477:247–251. doi: 10.1016/j.ejphar.2003.08.020. [DOI] [PubMed] [Google Scholar]
- 118.Araya M, Chen B, Klevay LM, Strain JJ, Johnson L, et al. Confirmation of an acute no-observed-adverse-effect and low-observed-adverse-effect level for copper in bottled drinking water in a multi-site international study. Regul Toxicol Pharmacol. 2003;38:389–399. doi: 10.1016/j.yrtph.2003.08.001. [DOI] [PubMed] [Google Scholar]
- 119.Gaddum, H J. Zinc sulphate 1% vomiting in a few minutes. Oxford University Press; 1961. Pharmacology.266 [Google Scholar]
- 120.Kan KK, Jones RL, Ngan MP, Rudd JA (25-10-2002) Actions of prostanoids to induce emesis and defecation in the ferret. Eur J Pharmacol. 453:299–308. doi: 10.1016/s0014-2999(02)02424-x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Dictyostelium chemotaxis was monitored by time-lapse photography to record cells moving within a Dunn Chamber towards the chemo-attractant, cAMP, across the screen from left to right. Images were taken every 6 seconds over a 15 minute period. Computer generated cell outlines enables average cell velocity, shape and direction of movement to be quantified.
(AVI)
Dictyostelium chemotaxis was monitored by time-lapse photography to record cells moving within a Dunn Chamber towards the chemo-attractant, cAMP, across the screen from bottom right to top left. Images were taken every 6 seconds over a 15 minute period. Computer generated cell outlines enables average cell velocity, shape and direction of movement to be quantified. Cell movement was recorded over a 5 minute period, prior to the addition a range of emetic or aversive compounds (indicated by a black screen flash), and cell behaviour was recorded for a further 10 minutes. In the movie shown here, the stomach irritant copper sulphate (5 mM ) was added, causing Dictyostelium cells to slowly stop moving and lose shape over the ten min period tested.
(AVI)
Homology search results (BLAST analysis) of the Dictyostelium genome for proteins showing amino acid similarity to TRPV receptors from multiple species. Potential homologues are defined by an E-value of less than 1.00E-40 [11], thus Dictyostelium does not contain proteins showing significant sequence similarity to be considered as homologues.
(DOCX)
Homology search results (BLAST analysis) of the Dictyostelium genome for proteins showing amino acid similarity to known bitter receptors from multiple species. Potential homologues are defined by an E-value of less than 1.00E-40 [11], thus Dictyostelium does not contain proteins showing significant sequence similarity to be considered as homologues. N/A = Not applicable.
(DOCX)
Homology search results (BLAST analysis) of the Dictyostelium genome for proteins showing amino acid similarity to TRPM5 receptors from human and mouse. Potential homologues are defined by an E-value of less than 1.00E-40 [11], thus Dictyostelium does not contain proteins showing significant sequence similarity to be considered as homologues. N/A = Not applicable.
(DOCX)