

Abstract
Background
Ethanol intake levels characteristic of adult males and females emerge post-pubertally. The present set of experiments examined the consequences of pre-pubertal and adult gonadectomies to explore whether the presence of gonadal hormones at puberty exerts organizational influences and/or plays an activational role in age- and sex-typical patterns of ethanol consumption.
Methods
Male and female Sprague-Dawley rats were gonadectomized (GX), received sham gonadectomy (SH) or were left non-manipulated (NM) at one of two ages, either pre-pubertally on postnatal day (P) 23 (early) or post-pubertally in adulthood on P70 (late). Early surgery animals were tested for ethanol consumption either during adolescence (P28-39) or in adulthood at the same age that late surgery animals were tested (P75-86). Voluntary ethanol consumption was indexed using a 2 hr limited-access paradigm, with access to two bottles: one containing water, and the other a sweetened ethanol solution.
Results
Age of GX did not impact patterns of ethanol consumption. Removal of testicular hormones in males, regardless of age of removal, elevated consumption levels in adulthood to female-typical levels. Ovariectomy did not have notable effects on ethanol drinking in females. Ethanol intake and preference of early SH males was significantly greater than that of both late SH and NM males. Removal of the gonads prior to puberty did not influence ethanol drinking or preference during adolescence in either males or females.
Conclusions
These results suggest that testicular hormones play an activational role in lowering ethanol intake and preference of adult male rats. Pubertal hormones, in contrast, were found to exert little influence on ethanol drinking or preference during adolescence, although the effect of surgical manipulation itself during development was found to exert a long-lasting facilitatory effect on ethanol consumption in adulthood.
Keywords: adolescence, ethanol consumption, puberty, gonadal hormones, sex differences
Introduction
Research examining alcohol effects in humans has predominantly focused on males. Yet, epidemiological and clinical data have shown notable sex differences in alcohol use, sensitivity to certain alcohol effects and propensity for abuse and dependence. For instance, although women average fewer drinks per occasion than men (Chan, 2007; Dawson, 1993), they have shorter intervals between the onset of drinking and the emergence of problem drinking than men (Greenfield, 2002). Women also differ from men in their sensitivity to a number of acute and chronic consequences of ethanol, including ethanol-induced cognitive and behavioral impairments, and subjective intoxication (e.g., Fillmore and Weafer, 2004; Mills and Bisgrove, 1983; Mumenthaler et al., 1993; NIAAA, 2004; Wang et al., 2003). For example, when BECs were equated across sex, women were found to be less sensitive to alcohol-induced impairment of inhibitory control than men (Fillmore and Weafer, 2004), but more sensitive to alcohol-related cognitive disruptions in a divided attention task (Mills and Bisgrove, 1983) and impairments in short- and long- term memory (Mumenthaler et al., 1999). In addition to these acute effects, women are also more sensitive than men to long-term consequences of drinking, with women more at risk for ethanol-induced liver disease, heart damage and brain damage (Mann et al., 2005; Urbano-Márquez et al., 1995).
In studies using laboratory animals, modest sex differences have been observed in adult rats and mice in tests of ethanol-induced social suppression (Varlinskaya et al., 2010), locomotor activity (Middaugh et al., 1992), and hang-over related decreases in social behavior (Varlinskaya and Spear, 2004). Sex differences in ethanol intake are also well-established in laboratory rodents during adulthood, although in contrast to the typical sex difference seen in human consumption (Chan, 2007; Dawson, 1993; SAMSHA, 2008), adult female rats and mice drink more ethanol on a g/kg basis than adult males in a number of paradigms. This sex difference in ethanol intake has been observed in intake tests ranging from 24 hr continuous access (Doremus et al., 2005; Lancaster et al., 1996; Yoneyama et al., 2008) and limited-access 2-bottle choice situations (Chester et al., 2006; Le et al., 2001; Vetter-O’Hagen et al., 2009) to operant self-administration tests (Blanchard and Glick, 1995; Blanchard et al., 1993). Such intake differences between the sexes are evident in outbred rats (e.g., Doremus et al., 2005; Lancaster et al., 1996) as well as in rats and mice selected for high or low alcohol intake (e.g., Chester et al., 2006; Le et al., 2001).
These adult-typical sex differences in ethanol intake are not reliably observed in adolescent rodents. For instance, recent research in our laboratory has found adolescent sex differences opposite those seen in adulthood, with adolescent male rats consuming more g/kg ethanol than adolescent females in a limited-access paradigm (Anderson et al., 2009; Vetter-O’Hagen et al., 2009). Sex differences in adolescents are less prevalent in continuous ethanol access paradigms, with one report finding no differences between the sexes (Lancaster et al., 2006), while another reported greater intake among adolescent female relative to male rats (Truxell et al., 2007).
Similar to the developmental pattern observed among rodents, sex differences in alcohol consumption reported in human adults likewise appear to emerge only gradually during development. Several recent studies have reported that young adolescent males and females between the ages of 12 and 17 show similar frequencies and quantities of alcohol use that begin to diverge thereafter (McPherson et al., 2004; Patton et al., 2004; Substance Abuse and Mental Health Services Administration [SAMHSA], 2008). Even among adults aged 18-25, the most recent SAMHSA report showed that frequency of current use did not differ by sex, although men reported more heavy and binge drinking than their female counterparts (SAMHSA, 2008). Taken together, it appears that in both humans and laboratory animals, species-typical sex differences in alcohol sensitivity and drinking patterns tend to emerge gradually after puberty, suggesting a possible influence of rising gonadal hormone levels on ethanol intake (see Witt, 2007).
Indeed, ethanol intake not only differs by sex, but also by age in both humans (Johnston et al., 2004) and laboratory rodents, with adolescent humans drinking approximately 2 times more alcohol per occasion than adults (SAMSHA, 2007) and adolescent rats exhibiting 2-3 fold higher ethanol consumption than their adult counterparts (Brunell and Spear, 2005; Doremus et al., 2005). This relatively high intake typical of adolescent rats decreases dramatically across the adolescent to adult transition For example, in a study examining the pattern of elevated intake observed during adolescence, males showed a clearly elevated plateau during early and mid adolescence (P28-39) followed by a sharp decline at approximately P40 to reach intakes modestly elevated over adult-typical consumption levels, with drinking gradually declining thereafter to reach those characteristic of adulthood by approximately P70 (Vetter et al., 2007). Unfortunately, this study did not include females, and what data are available are mixed as to whether adolescent female intake declines into adulthood, with some studies reporting that adolescent males and females drink more than adults of both sexes (Doremus et al., 2005), whereas others found no intake differences between adolescent and adult females (Vetter-O’Hagen et al., 2009).
Traditionally, organizational sculpting of brain structure by gonadal hormone exposure was thought to be restricted to a critical period during the prenatal and early postnatal period (Phoenix et al., 1959). Recent evidence, however, suggests that another organizational period for steroid-dependent organization of brain structure and behavior may occur during adolescence, with increases in sex hormones at puberty acting to induce further organizational influences on the brain to give rise to sexually dimorphic reproductive behaviors, along with other sex-typical behaviors (e.g., agonistic, aggressive and anxiety-related behaviors -- see Primus & Kellogg, 1989, 1990; Romeo et al., 2003; Schulz et al., 2004; Schulz and Sisk, 2006). It has yet to be determined whether increases in gonadal hormones occurring during adolescence play an organizational role in other post-pubertally emerging adult sex-typical behaviors, such as sex differences in ethanol intake and sensitivity.
The present set of experiments examined the consequences of pre-pubertal as well as adult gonadectomies to assess whether the presence of gonadal hormones exerts organizational influences at puberty and/or plays an activational role during adolescence or in adulthood on age- and sex-typical patterns of ethanol consumption indexed via voluntary limited-access intake tests.
Materials and Methods
Subjects
A total of 160 male and female Sprague-Dawley rats bred in our colony were used in these experiments. On postnatal day (P) 1, the day after birth, litters were culled to 8-10 pups, with 6 animals of one sex and 4 of the other kept whenever possible. Offspring were weaned and pair-housed with a same-sex littermate in a temperature-controlled vivarium on a 14:10 light/dark cycle (lights on at 0700) and given ad libitum access to food (Purina Rat Chow, Lowell, MA) and water until the onset of experimental procedures. At all times, animals were treated in accordance with guidelines for animal care established by the National Institutes of Health (Institute of Laboratory Animal Resources, Commission on Life Science, 1996), using protocols approved by Binghamton University Institutional Animal Care and Use Committee. Throughout these experiments, no more than one animal per litter was placed into any given experimental condition.
Experimental Design
In order to examine the influence of pubertal hormones on ethanol consumption, both male and female animals were gonadectomized (GX) or received sham gonadectomy (SH) at one of two ages, either pre-pubertally on P23 (early) or post-pubertally in adulthood on P70 (late). Early surgery animals were either tested shortly after surgery (i.e., during adolescence) or in adulthood at the same age that late surgery animals were tested. Six experimental groups from each sex were used (n=10/sex/group), for a total of 12 experimental groups: early GX, early SH, late GX, and late SH animals tested in adulthood; early GX and early SH animals tested in adolescence. An additional control group of non-manipulated (NM) males and females were also tested at each age (n=10/sex/age) in order to examine the possible influence of the surgical manipulation process per se on ethanol intake.
Surgery
Two days prior to surgery, animals were rehoused with a littermate of the same age and sex and assigned to the same surgery condition. Animals were anesthetized using isoflurane (3-3.5% initially) and maintained at surgical levels of anesthesia throughout the surgery via nose cone supplementation (3% repeated as necessary). For castration in males, each testis was removed, a suture made in each tunic and in the inguinal ring (to prevent possible herniation) and the incision closed with Vetbond tissue adhesive (3M, St. Paul, MN). For ovariectomies, an incision was made on the dorsal side of the animal, caudal to the last rib and through the skin perpendicular to the midline. On each side of this incision, an opening was made in the muscle wall via blunt dissection, with the oviduct on each side sutured proximal to the ovary, the ovary excised, and the muscle wall sutured. For sham gonadectomies, the same procedures were followed, except that reproductive tissue was not manipulated, nor the gonads removed. Immediately following surgery and again the following day, animals received a subcutaneous injection of the anti-inflammatory agent Ketofen (5 mg/kg). Following surgery, animals were returned to their home cages, with a wire-mesh divider used to separate each housing pair for a recovery period of approximately 72 hrs. On the fourth day, the divider was removed.
Ethanol Intake Testing
The 12 days of ethanol intake testing began on the fifth day after surgery for the early GX and SH animals tested during adolescence (i.e., P28-39) and for the late GX and SH animals tested during adulthood (P75-86). Early GX and SH animals assigned to be tested during adulthood were left non-manipulated after the post-surgical period until ethanol intake testing in adulthood (P75-86). On the day before ethanol intake testing began, animals were weighed to establish a pre-test baseline body weight prior to the initiation of water deprivation. Animals were then water deprived for 24 hrs prior to the first 2 hr limited-access ethanol intake session. Fifteen min prior to each session, animals in each housing pair were separated from each other in their home cage by a mesh divider (the same divider used post-surgically) in order to assess intakes of each animal individually. During the 2 hr limited-access sessions, each animal was given access to two bottles: one containing water, and the other ethanol sweetened with 0.1% saccharin at concentrations of 6% ethanol (experimental days 1-4) and 10% ethanol (experimental days 5-12). The position of the two bottles was alternated daily to avoid development of side preferences. Fifteen min after the end of each day’s 2 hr access session, the divider was removed from the home cage. Approximately 30 min after each daily access session, each pair of animals received a daily supplemental water allocation, calculated as described below. This procedure has been successfully used to motivate animals to drink fluids within the restricted 2 hr period in previous studies in our laboratory (Vetter-O’Hagen et al., 2009).
A preliminary study examining the functional effects of gonadectomy on body weight gain of pre- and post-pubertally gonadectomized male and female rats relative to their sham gonadectomized and non-manipulated counterparts revealed that body weights of these ad libitum-fed and watered animals varied not only as a function of age and sex, but also across surgery condition. Among early surgery (P23) animals, significant weight differences did not emerge in either sex until after adolescence, with GX males weighing less than their SH or NM counterparts and early GX females weighing more than their SH or NM counterparts in adulthood. Similar weight differences in adulthood were observed in late GX animals, with GX males weighing less than controls and a trend for GX females to weigh more than controls (Vetter-O’Hagen and Spear, 2009). Using body weight data from that study, all animals in the current study were provided with sufficient supplementary water to maintain weight trajectories that were approximately 85% that of non-deprived animals of the corresponding age, sex and surgery condition. To accomplish this goal, body weights of each housing pair of animals were monitored daily and the amount of supplemental water provided to each pair was adjusted daily by 1-3 ml in order to maintain each animal along the pre-determined weight trajectory appropriate for each age, sex and surgical condition. All supplemental water was fully consumed overnight before the start of the ethanol intake session. In previous experiments using the same procedure, pair-housed animals given supplementary water titrated in this manner each gained weight daily and maintained appropriate weight trajectories for their age and sex throughout the experimental period (Vetter-O’Hagen et al., 2009). Since provision of supplemental water was used to maintain animals on the proper weight trajectory for each condition, differences in supplemental amounts water were expected. During adolescence, as expected given the lack of group differences in body weight at that time, analyses of variance comparing mean supplemental water (mean ± SEMs per cage) provided across days in the GX (32.8 ± 0.3), SH (32.5 ± 0.2) and NM (32.6 ± 0.2) groups did not differ significantly by sex or surgery condition. In adults, consistent with the differences in weight trajectories across groups, the analysis of variance on supplemental water (mean mls ± SEMs per cage) provided across days revealed greater intake in GX (46.3 ± 0.4 ml) than SH (44.5 ± 0.3 ml) and NM (43.6 ± 0.7 ml) females (F(2, 22) = 13.49, p< .01), whereas in males, GX (52.6 ± 0.4) animals received less supplemental water than SH (54.1 ± 0.4) and NM (54.4 ± 0.4) animals (F(2, 22) = 12.65, p< .01).
Blood samples from the tail were collected immediately following the 2 hr access session half-way through ethanol intake testing (i.e. on day 6) for analysis of blood ethanol concentrations (BECs). Trunk blood was collected after the last intake session (day 12) for analysis of BEC and gonadal hormone levels (testosterone or estradiol for males and females, respectively). Whole blood for BEC analyses and plasma for hormone assays were stored at −80° C until the time of assay. Gonadal hormone assays were used to confirm removal of gonadal tissue via comparison of gonadectomized groups with sham and non-manipulated groups.
Blood Analyses
For analysis of BECs, an aliquot of blood was thawed and BECs were determined by means of head-space gas chromatography, using a Hewlett Packard (HP) 5890 series II Gas Chromatograph, a HP 7694E Headspace Sampler, and HP Chemstation software that compares the peak area under the curve in each sample with those of standard curves derived from reference standard solutions. At the time of assay, whole blood samples were placed in airtight vials, heated in a water bath (55 °C) for 7 min, and a 1.0 ml sample from the gas head space extracted with an airtight syringe and injected onto the column (see Silveri and Spear, 2000).
For analysis of testosterone and estradiol, plasma samples were thawed, and testosterone and estradiol levels assessed via radioimmunoassay using a 125I RIA double antibody kit from MP Biomedicals (Solon, OH), with specificities of 100% for each hormone assayed. Testosterone assay sensitivity was 0.03 ng/ml, with inter- and intra-assay coefficients of variation of 10.1% and 6.6%, respectively. Estradiol assay sensitivity was 7.2 pg/ml, with inter-assay coefficients of 9% and intra-assay coefficients of 7.3%. Samples and standards for each hormone assay were run in duplicate, using a Packard Cobra II Autogamma Counter, with disintegrations per min averaged against a standard curve.
Data Analysis
In order to address whether gonadectomy impacted ethanol consumption, preference, BECs, hormone levels and total fluid intake in adulthood, analyses of the data first focused on adult test gonadectomy effects using separate 2 sex × 2 surgery condition (GX vs. SH) × 2 surgery age (early vs. late) × 6 block (2-day blocks) repeated measures analyses of variance (ANOVAs). Due to high variability in intake measures across days, all intake and preference data were combined into 2-day blocks before analysis. The second set of analyses of the adult test data focused on surgical manipulation effects on intake and preference by comparing the adult test data from SH manipulated animals to that of NM animals using 2 sex × 3 surgical manipulation (early SH vs. late SH vs. NM) × 6 block repeated measures ANOVAs.
In the analysis of ethanol intake, preference and total fluid data of adolescent-tested animals, 2 sex × 3 surgery condition (GX vs. SH vs. NM) × 6 block repeated measures ANOVAs were used, with similar 2-way ANOVAs minus the repeated measure of block used in the analysis of pre-test body weight, BEC and hormone data.
Prior to these analyses, Levene’s tests were used to test for homogeneity of variance (HV) in each data set, with testosterone data being the only data set that violated HV assumptions due to floor effects in both adolescent and adult GX males. Consequently, in adults, testosterone data from GX and SH males were analyzed using the nonparametric Kruskal-Wallis H test. Comparisons of plasma testosterone levels between SH and NM adult males did not violate the HV assumption, thus ANOVAs were used for those comparisons. Among adolescents, GX, SH and NM testosterone data was analyzed using the Kruskal-Wallis H test, followed by Mann-Whitney U tests to determine significant differences between the surgery conditions. Significant ANOVA results involving interactions with sex as a factor were explored further using separate ANOVAs for each sex, with Fisher’s LSD tests used to determine the locus of significant main effects and interactions. Block effects unrelated to other variables are de-emphasized in the results.
Results
Impact of Age of Gonadectomy on Adult Assessments
Ethanol intake
The analysis of mean daily g/kg ethanol intake in adulthood revealed significant main effects of sex (F(1, 72) = 12.35, p< .01), surgery condition (F(1, 72) = 9.9, p< .01), and block (F(5, 360) = 25.56, p< .01), along with a significant interaction among these three variables (F(5, 360) = 2.57, p< .05). In general, females drank more than males, with intakes generally lowest among SH males and ethanol intake of GX males increasing across test blocks to reach levels similar to that of females by the end of testing (Fig. 1 top panel). When the data were analyzed separately by sex to explore sex-dependent gonadectomy effects in more detail, GX males were found to consume more ethanol than SH males, regardless of age of surgery [main effect of surgery condition (F(1, 36) = 10.72, p< .05)] (see Fig. 1 top, left panel). The only effect involving surgery age was a block × surgery age interaction (F(5, 180) = 3.26, p< .01), with ethanol intake greater in early than late surgery males during the first block. Among females, no significant effects involving surgery condition or surgical age were evident, although there was a trend (p= .07) for late SH females to drink less than early SH females and both GX groups (see Fig. 1 top, right panel).
Fig. 1.
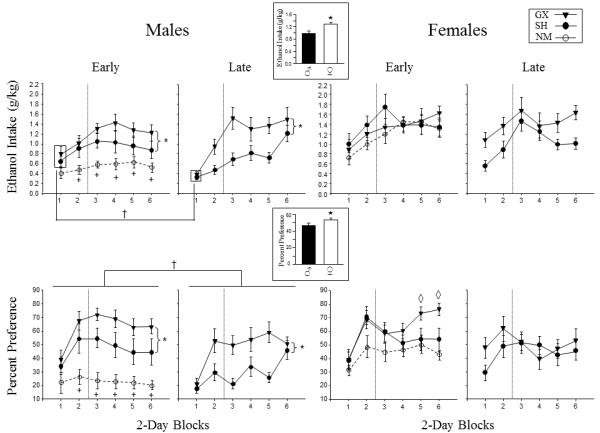
Mean ethanol intake (g/kg) and percent preference of early and late gonadectomized (GX) adult males and females is shown compared to their sham (SH) counterparts across the six 2-day blocks of measurement. The vertical dashed line represents the increase in ethanol concentration (from 6% to 10%) provided to animals during the 2 hr limited-access session and the bars represent standard error. The * symbol indicates the significant difference (p< .05) between GX and SH males collapsed across block. The † symbol denotes significant differences (p< .05) between the early and late surgery groups during the blocks indicated. The ◊ denotes the significant difference (p< .05) between GX and SH females during the blocks indicated. Sex differences (p< .05; indicated by the ★ symbol) are shown in inserts, with the data collapsed across all other variables. Although the non-manipulated (NM) groups were analyzed separately with the early and late SH groups, for the sake of comparison, mean ethanol intake and percent preference is shown in the early surgery panels for males and females, with the + symbol indicating significant differences (p< .05) between early SH and NM males.
Preference data
Preference scores were calculated via the formula: [((ethanol solution intake)/(ethanol solution intake + water intake)) *100], with values >50% reflecting a preference for the ethanol solution and scores <50% reflecting a water preference. In the analysis of percent preference scores following early and late GX or SH surgery, main effects of surgery condition (F(1, 72) = 10.12, p< .01) and of surgical age (F(1, 72) = 13.86, p< .01) were tempered by interactions of these variables with block and with sex [(F(5, 360) = 2.77, p< .05) and (F(5, 360) = 2.93, p< .05), respectively]. Gonadectomy markedly increased ethanol preference of males to reach levels seen in GX and SH females, an effect that emerged with the increase in preference scores seen in all groups following the first test block (Fig. 1 bottom panel). The separate ANOVA of the male data also revealed evidence for significantly greater preference scores among GX than SH males beginning during the 2nd block (block × surgery condition interaction (F(5, 180) = 2.32, p< .05), with this ANOVA also revealing significantly greater preferences among early than late surgery males [main effect of surgical age (F(1, 36) = 10.38, p< .01) – see Fig. 1 bottom, left panel]. In the ANOVA of the female preference data (Fig. 1 bottom, right panel), preference scores of early GX females increased to reach levels significantly elevated over all other groups by the last 2 test blocks, with no significant differences among females in the late surgery groups [block × surgery condition × surgery age interaction (F(5, 180) = 2.81, p< .05)].
Pre-test body weight, BEC, hormone, and total fluid data
The analysis of pre-test body weight data revealed main effects of sex (F(1, 72) = 547.93, p< .01) and surgery age (F(1, 72) = 23.99, p< .01) as well as a sex × surgery condition × surgery age interaction (F(1, 72) = 15.14, p< .01). In addition to the typical sex difference in body weights, sex-dependent effects of gonadectomy on body weights were seen in early surgery animals, with GX males weighing significantly less than early SH males by the time they had reached adulthood, whereas the converse was seen in females, with early GX females weighing significantly more than SH females (Table 1). Trends for similar patterns of weight differences were seen among the late surgery animals, although these differences did not reach statistical significance (males: p= .06; females p= .07) at the time body weights were assessed (4 days post-surgery). In the analysis of BECs, no significant sex, surgery condition or age of surgery effects emerged in early or late SH or GX animals on either day that samples were taken (see Table 1). As expected, gonadal hormone levels in adulthood were significantly reduced after both early and late gonadectomy in males and females (Table 1), with testosterone levels in GX males significantly lower than those of SH males (H(1, 40) = 30.88, p< .01), as were estradiol levels in GX relative to SH females [main effect of surgery condition: (F(1, 36) =12.15, p< .01)]. The ANOVA of total fluid intake revealed significant main effects of sex (F(1, 72) = 8.48, p< .01), with females generally drinking more relative to their body weight than males, surgery age (F(1, 72) = 10.38, p< .01) and block (F(5, 360) = 79.74, p< .01), and were tempered by several significant interactions involving these factors as well as surgery condition. The ANOVA of total fluid intake in males revealed, significant effects of surgery age (F(1,36) = 6.81, p< .05), block (F(5, 180) = 36.34, p< .01) and their interaction (F(5, 180) = 12.92, p< .01), with late surgery males drinking more than early surgery males in blocks 3-6 regardless of surgery condition (late surgery mean intake ± SEM across blocks: 36.70 ± 2.32 ml/kg; early surgery: 30.36 ± 1.75 ml/kg). Among females, this effect of surgery age (F(1, 36) = 4.82, p< .05) was tempered by a surgery condition × surgery age interaction (F(1, 36) = 6.12, p< .05), with late GX females (means ± SEMs) drinking significantly more ml/kg total fluid () than late SH females () and than early GX () and early SH females ().
Table 1.
Adult Pre-test Body Weights, Blood Ethanol Concentrations (BECs), and Hormone Levels ± SEMs
| Hormonal Status | Body Weight (g) | BECs (mg/dl) | Hormone Levels | |||||
|---|---|---|---|---|---|---|---|---|
| Males | Females | Males | Females | |||||
| Early Surgery | Males | Females | Day 6 | Day 12 | Day 6 | Day 12 | Testosterone (ng/ml) | Estradiol (pg/ml) |
| GX | 380.0 ± 9.9 * | 331.3 ± 9.7 * | 32.1 ± 7.2 | 24.1 ± 9.3 | 19.2 ± 8.2 | 19.8 ± 4.9 | 0.0 ± 0.0 * | 62.5 ± 3.4 * |
| SH | 438.9 ± 11.6# | 268.2 ± 4.7# | 25.1 ± 5.3 | 10.8 ± 5.6 | 25.3 ± 9.8 | 20.2 ± 4.6 | 1.6 ± 0.8 | 82.1 ± 7.4 |
|
| ||||||||
| Late Surgery | ||||||||
|
| ||||||||
| GX | 387.5 ± 4.3 | 269.2 ± 4.8 | 32.4 ± 9.0 | 17.8 ± 5.6 | 27.1 ± 7.2 | 19.4 ± 5.8 | 0.0 ± 0.0 * | 62.1 ± 3.0 * |
| SH | 407.9 ± 5.8 | 250.0 ± 5.0 | 8.4 ± 2.5 | 32.4 ± 26.3 # | 27.0 ± 8.1 | 21.0 ± 5.4 # | 1.3 ± 0.6 | 74.8 ± 4.8 |
|
| ||||||||
| NM | 402.3 ± 8.4 | 258.2 ± 5.9 | 5.6 ± 3.8 | 1.2 ± 0.8 | 14.1 ± 7.8 | 14.1 ± 10.8 | 1.0 ± 0.4 | 74.6 ± 2.4 |
GX animals differed significantly from their same-sex SH counterparts (p< .01)
SH animals differed significantly from NM animals regardless of sex (p< .05)
Surgical Manipulation Effects
Ethanol intake and preference data
The separate 2 sex × 3 surgical manipulation (early SH vs. late SH vs. NM) × 6 block ANOVAs comparing ethanol intake and percent preference of early and late SH animals with that of NM (see Fig. 1) revealed main effects of sex, with females consuming more ethanol and showing greater preference scores than males [intake: (F(1,54) = 25.89, p< .01); preference: (F(1, 54) = 12.81, p< .01), respectively]. Significant effects of surgical manipulation [(F(2, 54) = 4.01, p< .05); (F(2, 54) = 7.00, p< .01)] and block [(F(5, 270) = 13.84, p< .01); (F(5, 270) = 8.03, p< .01)] were also evident for both measures. In the analysis of the male data, the early SH group showed elevated ethanol intake and preference relative to that of NM and, to a lesser extent, late SH animals on most days of testing [effects of surgical manipulation [(F(2, 27) = 3.97, p< .05); (F(2,27) = 6.74, p< .01)], and surgical manipulation × block [(F(10, 135) = 2.39, p< .01); (F(10, 135) = 1.88, p≤ .05)]. Among females, no significant differences emerged among the early and late SH and NM groups.
Pre-test body weight, BEC, hormone, and total fluid data
In the analysis of pre-test body weight data, a main effect of sex (F(1, 54) = 696, p< .01) was evident, with males weighing more than females. A main effect of surgery age (F(2, 54) = 7.15, p< .01) also emerged, with early SH animals weighing significantly more than late SH and NM animals regardless of sex; this finding, however, appears to be driven more by the males given that this effect persisted only in the analysis of males when the data were analyzed separately by sex. In the ANOVA focused on comparing day 6 BEC levels of early and late SH animals with NM animals, there were no significant effects. On day 12, BECs in the late SH group were significantly greater than NM animals [main effect of age of surgery (F(2, 54) = 4.10, p< .05)], regardless of sex (Table 1). Gonadal hormone levels in NM males and females did not differ significantly from their same-sex SH counterparts (Table 1). The analysis of total fluid intake revealed a significant main effect of sex (F(1, 54) = 4.72, p< .05) and a block × surgery age interaction (F(10, 270) = 2.51, p< .01), with females drinking more total fluid than males and total fluid intake significantly lower in early SH than NM animals only in block 4.
Impact of Early Gonadectomy and Surgical Manipulation during Adolescence
The analysis of ethanol intake and percent preference data during adolescence revealed only significant main effects of block, with no significant effects of sex or surgery condition (Table 3). Likewise, no significant effects of surgery condition emerged in the analyses of body weights and BECs on day 6, with only a main effect of sex (F(1, 54) = 31.79, p< .01) emerging in the analysis of pre-test body weight (Table 2). In the analysis of BECs on day 12, however, GX animals displayed higher BECs than NM, but not SH animals, regardless of sex [main effect of surgery condition (F(2, 54) = 4.49, p< .05). Testosterone levels among males differed significantly by group (H(2, 30) = 10.05, p< .05), with lower plasma testosterone levels in GX males compared to their SH and NM counterparts. Among females, a trend for lower estradiol levels in GX compared to SH and NM was revealed (F(2, 27) = 2.75, p=.08), although, this effect did not reach statistical significance (Table 2). Total fluid intake also did not differ significantly by sex or surgery condition.
Table 3.
Pre-pubertal Gonadectomy and Surgical Manipulation Effects on Ethanol Intake and Preference During Adolescence (means ± SEMs) Collapsed Across Blocks
| GX |
SH |
NM |
||||
|---|---|---|---|---|---|---|
| Males | Females | Males | Females | Males | Females | |
| Ethanol Intake (g/kg) | 2.2 ± 0.2 | 2.3 ± 0.3 | 1.8 ± 0.2 | 2.1 ± 0.1 | 1.9 ± 0.1 | 1.7 ± 0.1 |
| Percent Preference | 40.4 ± 2.7 | 44.4 ± 4.4 | 35.1 ± 4.7 | 40.5 ± 2.7 | 36.3 ± 3.0 | 38.6 ± 4.1 |
Table 2.
Adolescent Pre-test Body Weights, Blood Ethanol Concentrations (BECs), and Hormone Levels ± SEMs
| Hormonal Status | Body Weight | BECs (mg/dl) | Hormone Levels | |||||
|---|---|---|---|---|---|---|---|---|
| Males | Females | Males | Females | |||||
| Males | Females◊ | Day 6 | Day 12 | Day 6 | Day 12 | Testosterone (ng/ml) | Estradiol (pg/ml) | |
| GX | 87.4 ± 1.4 | 82.9 ± 2.5 | 35.1 ± 9.4 | 35.9 ± 8.7 ** | 26.5 ± 6.9 | 34.5 ± 9.6 ** | 0.00 ± 0.00* | 52.4 ± 2.6 |
| SH | 91.4 ± 1.7 | 80.4 ± 2.2 | 28.4 ± 9.4 | 34.9 ± 7.0 | 20.1 ± 4.5 | 12.3 ± 2.5 | 0.13 ± 0.06 | 65.4 ± 5.3 |
| NM | 91.4 ± 2.3 | 80.8 ± 2.0 | 21.9 ± 10.3 | 9.6 ± 7.4 | 15.7 ± 6.5 | 16.9 ± 6.5 | 0.14 ± 0.05 | 57.2 ± 3.6 |
GX males differed significantly from their SH counterparts (p< .01)
GX animals differed significantly from their NM counterparts (p< .05)
Females differed significantly from males
Discussion
Overall, age at removal of the gonads did not impact patterns of ethanol consumption in the current study. Removal of testicular hormones in males, regardless of age of removal, resulted in female-typical levels of consumption and preference in adulthood, whereas ovariectomy had little effect on ethanol drinking of adult females. Age of the surgical manipulation process itself exerted a notable influence on adult ethanol consumption among males, with intake and preference of early SH males significantly greater than that of both late SH and NM males; similar trends did not reach significance in females.
In contrast, the presence or absence of gonadal hormones and the surgical manipulation process itself exerted little influence on ethanol drinking or preference in either females or males tested during adolescence. This lack of impact of GX among adolescent females could be in part a function of estradiol levels, which were not significantly attenuated by ovariectomy, perhaps not surprisingly given that estradiol levels are still rising during adolescence (Ojeda & Skinner, 2006) and hence removal of the ovaries may have had less of an impact on hormone levels at this age. This does not seem to be the case in adolescent males, with testosterone levels below detectable levels in GX males.
No significant effects of sex were observed in ethanol intake or preference during adolescence, although typical sex differences in ethanol intake were observed in adulthood, with adult females generally consuming more ethanol (g/kg) than males. This finding of greater ethanol intake among adult females than males is consistent with previous adult ethanol intake data in both rats and mice (i.e. Chester et al., 2006; Doremus et al., 2005; Vetter-O’Hagen et al., 2009; Yoneyama et al., 2008). The lack of sex differences in amount of g/kg ethanol consumed during adolescence, however, contrasts with previous studies from our laboratory that found adolescent male rats to drink more ethanol (g/kg) in a limited-access setting than adolescent females (Anderson et al., 2009; Vetter-O’Hagen et al., 2009). Intake differences across sex during adolescence are not always evident in rats, however (e.g., see Lancaster et al., 2006), and in mice, females of both ages tend to drink more than their male counterparts (Moore et al., 2010; Tambour et al., 2008). While it is possible that the stress of surgical manipulation in the GX and SH animals could have disrupted expression of sex differences in ethanol consumption during adolescence in the present study, there was also no sign of consumption differences between NM male and female adolescents, weakening this suggestion.
Sex-typical differences in ethanol intake and preference during adulthood were disrupted in GX males, with GX in males increasing ethanol intake to levels similar to that of females across the measurement period. In general, similar patterns were observed for preference scores, with GX markedly increasing the ethanol preference of males to reach levels comparable to those seen in GX and SH females. Thus, without testicular hormones, adult male rats tended to display more female-typical levels of ethanol consumption and preference in adulthood. These findings suggest a possible suppressant role of testicular hormones on drinking and preference during adulthood.
This effect of increased ethanol intake and preference in GX males compared to their SH counterparts was evident regardless of the developmental period in which testicular hormones were removed, with ethanol intake and preference elevated in adulthood in both early and late GX males. These results are more consistent with an activational rather than organizational role of testosterone and/or testicular hormones. Similar to these findings, recent work from another laboratory found that preweanling GX in Long Evans rats increased ethanol consumption in adult males relative to their SH counterparts (Sherrill et al., 2011). However, the Sherrill et al. (2011) study did not include animals similarly gonadectomized in adulthood, leading the authors to conclude that the effects of pre-pubertal gonadectomy on adult-typical sex differences in ethanol consumption were likely due to organizational effects exerted by gonadal hormones during puberty. The results of the present study, which included an adult gonadectomized group and tested in adulthood, do not support this conclusion. Additional support for an activational rather than organizational effect of gonadal hormones on ethanol consumption comes from a recent follow-up study in our laboratory focusing on gonadectomy in adult males which replicated the enhancement of ethanol intake by GX seen in the present study, and found that replacement of testosterone in castrated adult males was sufficient to return drinking behavior to levels comparable to that of SH males – data providing further evidence that testosterone acts in an activational manner to suppress ethanol consumption under these test circumstances (Vetter-O’Hagen et al., under revision). Nevertheless, a number of other studies that have examined ethanol consumption or preference in castrated males have reported no difference between gonadectomized and intact males (Almeida et al., 1998; Cailhol and Mormede, 2001; Hilakivi-Clarke, 1996). Daily dihydrotestosterone treatment was found, however, to decrease ethanol consumption in a continuous access, 2-bottle choice paradigm in both castrated and intact males, findings consistent with a possible suppressive effect of testosterone on ethanol drinking in males (Almeida et al., 1998). Another recent study concluded that alteration of ethanol drinking by testicular hormones may be influenced by environmental factors, with castration decreasing drinking in a continuous access paradigm in individually housed males with high baseline levels of drinking, but having no impact on males in a crowded housing situation (Begg and Weisinger, 2008). It is possible that methodological differences such as housing condition, drinking paradigm or deprivation status could explain the varying effects of castration found across studies.
Effects of gonadectomy in adult females were minimal, with the analyses focusing on females revealing no significant impact of ovariectomy on ethanol consumption. Likewise, ethanol preference of adult females was generally unaffected by GX, although greater preference in early GX relative to SH females was seen during the last test blocks, an effect that may be spurious as it was not evident in the analysis of intake nor was it seen in the late GX group. The lack of notable effects of ovariectomy on ethanol intake is in agreement with several studies which have reported that removal of the ovaries did not affect ethanol consumption in adult female rats (Almeida et al., 1998; Cailhol and Mormede, 2001) and mice (Hilakivi-Clarke, 1996). Other studies, however, have reported modest decreases in ethanol intake after gonadectomy in females (Becker et al., 1985), particularly when baseline levels of ethanol intake were equated across surgical groups and intake was examined within each subject in terms of change from baseline (Ford et al., 2002a). Although potentially subtle ovariectomy effects might have emerged if the design of the current experiments had included a pre-surgery baseline intake test, pre-surgery data would have been difficult to relate to adult intake data for the early surgery group. It is also possible that stage of estrous cycle in SH females could have influenced ethanol intake in the intact adult females, perhaps masking cycle-specific effects that may have been disrupted by ovariectomy. However, variability in the intake and preference data among adult SH and NM females was not greater than any other group, as would be anticipated if phase of estrous cycle had exerted notable influences on overall ethanol consumption and/or preference. Other studies have found that total ethanol intake was unaffected by stage of estrous cycle when females were allowed to cycle freely (Ford et al., 2002b; Roberts et al., 1998), although the microstructure of ethanol drinking was reported to vary across estrous phase (Ford et al., 2002b).
Another potential explanation for the lack of an effect of ovariectomy in females could be due to high levels of circulating plasma estradiol in GX females, which may have masked potential effects of ovarian removal on ethanol intake and preference. While estradiol levels of GX females were significantly lower than SH females these declines were relatively modest (~20-25%) in the GX groups. Typically, ovariectomy has been found to decrease plasma estradiol levels partially, but not completely (e.g. ~58% - Kim et al., 2004; ~46% - Moien-Afshari et al., 2003). A number of extra-ovarian sources contribute to circulating estradiol levels, including, estradiol secretion from the adrenals (Shaikh and Shaikh, 1975) and production by tissues in the lung, liver, kidney, intestine and uterus as well as in adipose tissue deposits (Takahashi, 1961). These nonovarian sources of estradiol likely contribute to the maintenance of significant estradiol levels among females in the current study, although it is possible that residual estradiol in GX females may also reflect in part incomplete gonadal removal, given that we did not confirm complete gonadal tissue removal through dissection at autopsy. Interestingly, most prior work examining the role of gonadal hormones in ethanol-related behavior do not report hormone levels after gonadectomy or use any other technique for verifying complete removal of gonadal tissue (Almeida et al., 1998; Begg and Weisinger, 2008; Cailhol and Mormede, 2001; Hilakivi-Clarke, 1996; Sherrill et al., 2011), with only a few reporting verification of tissue removal, but not circulating hormone levels (Gililland and Finn, 2007; Becker et al., 1985).
It is not simply the case that ovariectomy was without consequence in the females in this study. Although devoid of effects on ethanol intake, ovariectomy did have other effects, with early GX females tested in adulthood showing significantly greater increases in body weight than control females (and a trend for a similar effect in late GX females) prior to the initiation of water deprivation or ethanol intake testing, and late GX females showing significant elevations in total fluid intake relative to all other groups.
The process of early surgical manipulation itself was found to exert an impact on the later ethanol intake of males in adulthood, with the drinking of early SH males elevated relative to that of late SH and NM control males, an effect that was less robust among females (given that the effect did not reach significance in females when the data were analyzed separately by sex). It is possible that this consequence of the surgical process itself may be associated with the stress of this manipulation among juvenile males. Indeed, other stressors early in life have been shown to influence adult ethanol drinking. For instance, exposure to 10 days of footstock stress during adolescence was found to elevate later adult drinking behavior relative to non-stressed controls in both male and female high acohol preferring (HAP2 line) mice, an effect that was not evident when mice were exposed to the same stressor as adults (Chester et al., 2008). Likewise, maternal separation stress in preweanling rodents has repeatedly been shown to increase ethanol consumption in adulthood (Cruz et al., 2008; Huot et al., 2001; Rockman et al., 1987). Animals in the early SH group in the current study were surgically manipulated at P23 – i.e., during the juvenile period between conventional weaning (P21) and adolescence (P28-42, see Spear, 2000). While few studies to our knowledge have focused specifically on the influence of stress during this period on later ethanol consumption, our results combined with the results of others suggest that exposure to stressors during development can alter drinking in adulthood, an effect that may be especially robust in males. This effect appears to take some time to be expressed, given that early SH animals did not differ from NM controls in their preference or consumption of ethanol during the adolescent period per se.
It is possible, although unlikely, that procedural variables may have contributed to the increased ethanol intake effect observed in GX compared to SH adult males in this experiment. For example, since a sweetened ethanol solution (0.1% saccharin) was used in this study, the changes observed in ethanol solution drinking and preference after gonadectomy in males could potentially reflect changes in sweet taste preference. Yet, changes in sweet preference have not been previously seen following castration in male rats (Zucker, 1969) or hamsters (Zucker et al., 1972), although reductions in saccharin preference have sometimes (Zucker, 1969), but not always (Kenney and Redick, 1980), been reported following ovariectomy in female rats. Given these prior data, if the presence of the sweetener influenced post-gonadectomy intakes of the sweetened ethanol it would have been expected that gonadectomy would have disrupted ethanol consumption among females rather than males in the present study.
Group differences in water deprivation level could have also played a role in the ethanol intake effects observed among males. For example, because level of deprivation was adjusted to produce equivalent decreases in percentage of normal body weight trajectories for each age/sex/surgical condition group, some groups (e.g., adult GX males and adult SH females), received less supplemental water than other groups (e.g., adult SH males and adult GX females), in order to maintain weights appropriate for their sex and surgical condition. Since supplemental water was entirely consumed overnight, groups that received a smaller water ration could have had a longer duration of deprivation. This could have potentially led to differential levels of thirst motivation, thereby affecting ethanol consumption. Were this the case, it would be expected that the groups showing the greatest ethanol consumption would also have consumed the most total fluid (i.e., water + ethanol solution) during the limited access intake session. This does not seem to be the case. For example, GX males showed greater EtOH consumption but not total fluid consumption, whereas late GX females had significantly elevated total fluid consumption but did not consume more ethanol than the other groups of females. Level of water deprivation also does not appear to contribute to the early SH effect observed in males for two reasons, the first being that supplemental water provided to early and late SH males did not differ, and the second being that the pattern of total fluid intake in early SH males was actually somewhat lower than that of NM, although this effect only reached significance during the 4th intake block, an effect opposite of what would be expected if these animals were more water deprived (and hence more motivated to drink fluid) than NM or late SH males. Further evidence that deprivation level is not a contributing factor to the elevated ethanol consumption of GX males in the present study are recent findings by Sherrill et al. (2011) showing similar effects without the use of food or water deprivation (Sherrill et al., 2011). Thus, collectively, the changes in ethanol intake after removal of the testes in males observed in this study do not appear to be related to group differences in level of water deprivation.
Blood ethanol concentrations taken at the end of the 2 hr limited-access session do not appear to accurately reflect group differences in ethanol consumption in both the adolescent and adult tested groups. Perhaps most notable was the lack of a significant effect among the adult tested animals, with GX males not displaying significantly elevated BECs relative to SH males, even though robust differences in ethanol drinking were evident between these two groups. Additionally, among the adolescent tested animals, some group differences were revealed in the analysis of BECs on day 12 that were not reflected in the ethanol intake data, with GX animals displaying greater BECs than NM animals regardless of sex, but with no significant differences in ethanol intake or preference across these groups. The most likely explanation for this apparent dissociation between ethanol drinking and BECs is that the time-point chosen for blood sampling was too long after the initiation of drinking. Previous reports of the temporal pattern of ethanol drinking during limited-access sessions have found that in ethanol preferring (P) rats, animals drink most of the total fluid consumed during a session in the first 12 min of the access period (Bell et al., 2006) and in C57BL/6J mice, similar temporal patterns of licking were observed, with number of sipper tube contacts greatest during the first 20-30 min of the limited-access session (Ford et al., 2005; Rhodes et al., 2006). Thus, in the current experiment, it is likely that BECs had peaked and begun to decline by the time blood samples were taken at the end of the 2 hr access period, thereby missing the time when group differences in BECs may have been most pronounced. For example, among the adolescent test groups, GX animals could have consumed more ethanol later in the intake session, compared to their NM counterparts, leading to a later peak in blood ethanol levels and higher levels at the time of blood sampling. Another explanation for the lack of significantly greater BECs among adult GX males could be metabolic changes induced by castration. In rats, some studies have shown that levels of alcohol dehydrogenase are elevated in gonadectomized males (Mezey and Potter, 1985; Mezey et al., 1980) -- these castration-induced increases in liver enzymatic activity have sometimes (Cicero et al., 1980), but not always (Cicero et al., 1990) been found to be sufficient to result in lower BECs. In C57BL/6J and DBA/2J mice, however, gonadectomy has been found to have little impact on BECs or ethanol clearance rates after a large dose of ethanol in males and females (Gililland and Finn, 2007).
Taken together, the results of the current study provide evidence for an activational role of testosterone or other testicular products in the sex differences observed in ethanol drinking in adult rats. Since the age of removal of the gonads did not impact ethanol consumption, it can be concluded that age-related and sex-related patterns of ethanol consumption under these limited-access conditions do not appear to be related to organizational influences of pubertal hormones. Furthermore, pre-pubertal removal of the gonads did not impact levels of ethanol intake during adolescence, suggesting that non-gonadal factors contribute to elevations in ethanol consumption seen during this developmental period.
Acknowledgements
The authors would like to thank Julia Shultz for performing the gonadectomy and sham surgeries and Judy Sharp for analyzing the hormonal and blood ethanol concentration assays.
Funding acknowledgements: This research was supported by NIAAA grant R01-AA017355
References
- Anderson RI, Vetter-O’Hagen CS, Spear LP. Effects of housing/test conditions on limited-access ethanol consumption in adolescent and adult male and female rats. Alcohol Clin Exp Res. 2009;33:220A. [Google Scholar]
- Almeida OF, Shoaib M, Deicke J, Fischer D, Darwish MH, Patchev VK. Gender differences in ethanol preference and ingestion in rats. The role of the gonadal steroid environment. J Clin Invest. 1998;101:2677–2685. doi: 10.1172/JCI1198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker HC, Anton RF, De Trana C, Randall CL. Sensitivity to ethanol in female mice: effects of ovariectomy and strain. Life Sci. 1985;37:1293–1300. doi: 10.1016/0024-3205(85)90244-9. [DOI] [PubMed] [Google Scholar]
- Begg DP, Weisinger RS. The role of adrenal or testicular hormones in voluntary ethanol and NaCl intake of crowded and individually housed rats. Physiol Behav. 2008;93:408–413. doi: 10.1016/j.physbeh.2007.09.007. [DOI] [PubMed] [Google Scholar]
- Bell RL, Rodd ZA, Lumeng L, Murphy JM, McBride WJ. The alcohol-preferring P rat and animal models of excessive alcohol drinking. Addict Biol. 2006;11:270–288. doi: 10.1111/j.1369-1600.2005.00029.x. [DOI] [PubMed] [Google Scholar]
- Blanchard BA, Glick SD. Sex differences in mesolimbic dopamine responses to ethanol and relationship to ethanol intake in rats. Recent Dev Alcohol. 1995;12:231–241. doi: 10.1007/0-306-47138-8_15. [DOI] [PubMed] [Google Scholar]
- Blanchard BA, Steindorf S, Wang S, Glick SD. Sex differences in ethanol-induced dopamine release in nucleus accumbens and in ethanol consumption in rats. Alcohol Clin Exp Res. 1993;17:968–973. doi: 10.1111/j.1530-0277.1993.tb05650.x. [DOI] [PubMed] [Google Scholar]
- Brunell SC, Spear LP. Effect of stress on the voluntary intake of a sweetened ethanol solution in pair-housed adolescent and adult rats. Alcohol Clin Exp Res. 2005;29:1641–1653. doi: 10.1097/01.alc.0000179382.64752.13. [DOI] [PubMed] [Google Scholar]
- Cailhol S, Mormede P. Sex and strain differences in ethanol drinking: effects of gonadectomy. Alcohol Clin Exp Res. 2001;25:594–599. [PubMed] [Google Scholar]
- Chan KK, Neighbors C, Gilson M, Larimer ME, Marlatt G Alan. Epidemiological trends in drinking by age and gender: providing normative feedback to adults. Addict Behav. 2007;32:967–976. doi: 10.1016/j.addbeh.2006.07.003. [DOI] [PubMed] [Google Scholar]
- Chester JA, de Paula Barrenha G, DeMaria A, Finegan A. Different effects of stress on alcohol drinking behaviour in male and female mice selectively bred for high alcohol preference. Alcohol Alcohol. 2006;41:44–53. doi: 10.1093/alcalc/agh242. [DOI] [PubMed] [Google Scholar]
- Chester JA, Barrenha GD, Hughes ML, Keuneke KJ. Age- and sex-dependent effects of footshock stress on subsequent alcohol drinking and acoustic startle behavior in mice selectively bred for high-alcohol preference. Alcohol Clin Exp Res. 2008;32:1782–1794. doi: 10.1111/j.1530-0277.2008.00763.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cicero TJ, Bernard JD, Newman K. Effects of castration and chronic morphine administration on liver alcohol dehydrogenase activity and the metabolism of ethanol in the male sprague-dawley rat. J Pharmacol Exp Ther. 1980;215:317–324. [PubMed] [Google Scholar]
- Cicero TJ, Greenwald J, Nock B, O’Connor L. Castration-induced changes in the response of the hypothalamic-pituitary axis to alcohol in the male rat. J Pharmacol Exp Ther. 1990;22:456–461. [PubMed] [Google Scholar]
- Cruz FC, Quadros IM, da Planeta CS, Miczek KA. Maternal separation stress in male mice: long-term increases in alcohol intake. Psychopharmacology (Berl) 2008;201:459–468. doi: 10.1007/s00213-008-1307-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dawson DA. Patterns of alcohol consumption: beverage effects on gender differences. Addiction. 1993;88:133–138. doi: 10.1111/j.1360-0443.1993.tb02771.x. [DOI] [PubMed] [Google Scholar]
- Doremus TL, Brunell SC, Rajendran P, Spear LP. Factors influencing elevated ethanol consumption in adolescent relative to adult rats. Alcohol Clin Exp Res. 2005;29:1796–1808. doi: 10.1097/01.alc.0000183007.65998.aa. [DOI] [PubMed] [Google Scholar]
- Fillmore MT, Weafer J. Alcohol impairment of behavior in men and women. Addiction. 2004;99:1237–1246. doi: 10.1111/j.1360-0443.2004.00805.x. [DOI] [PubMed] [Google Scholar]
- Ford MM, Eldridge JC, Samson HH. Ethanol consumption in the female Long-Evans rat: a modulatory role of estradiol. Alcohol. 2002a;26:103–113. doi: 10.1016/s0741-8329(01)00203-8. [DOI] [PubMed] [Google Scholar]
- Ford MM, Eldridge JC, Samson HH. Microanalysis of ethanol self-administration: estrous cycle phase-related changes in consumption patterns. Alcohol Clin Exp Res. 2002b;26:635–643. [PubMed] [Google Scholar]
- Ford MM, Nickel JD, Finn DA. Treatments with and withdrawal from finasteride alters ethanol intake patterns in male C57BL/6J mice: potential role of endogenous neurosteroids? Alcohol. 2005;37:23–33. doi: 10.1016/j.alcohol.2005.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gililland KR, Finn DA. The impact of gonadectomy and adrenalectomy on acute withdrawal severity in male and female C57BL/6J and DBA/2J mice following a single high dose of ethanol. Alcohol Clin Exp Res. 2007;31:1846–1857. doi: 10.1111/j.1530-0277.2007.00509.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenfield SF. Women and alcohol use disorders. Harv Rev Psychiatry. 2002;10:76–85. doi: 10.1080/10673220216212. [DOI] [PubMed] [Google Scholar]
- Hilakivi-Clarke L. Role of estradiol in alcohol intake and alcohol-related behaviors. J Stud Alcohol. 1996;57:162–170. doi: 10.15288/jsa.1996.57.162. [DOI] [PubMed] [Google Scholar]
- Huot RL, Thrivikraman KV, Meaney MJ, Plotsky PM. Development of adult ethanol preference and anxiety as a consequence of neonatal maternal separation in Long Evans rats and reversal with antidepressant treatment. Psychopharmacology (Berl) 2001;158:366–373. doi: 10.1007/s002130100701. [DOI] [PubMed] [Google Scholar]
- Institute of Laboratory Animal Research, National Research Council . Guide for the Care and Use of Laboratory Animals. National Academies Press; Washington, DC: 2010. [Google Scholar]
- Johnston LD, O’Malley PM, Bachman JG, Schulenburg JE. Monitoring the future national results on adolescent drug use: Overview of key findings, 2003. National Institute on Drug Abuse. 2004:1–59. [Google Scholar]
- Kenney NJ, Redick JH. Effects of ovariectomy and subsequent estradiol replacement on intake of sweet solutions. Physiol Behav. 1980;24:807–809. doi: 10.1016/0031-9384(80)90418-7. [DOI] [PubMed] [Google Scholar]
- Kim SW, Kim NN, Jeong S, Munarriz R, Goldstein I, Traish AM. Modulation of rat vaginal blood flow and estrogen receptor by estradiol. J Urol. 2004;172:1538–1543. doi: 10.1097/01.ju.0000137744.12814.2e. [DOI] [PubMed] [Google Scholar]
- Lancaster FE, Brown TD, Coker KL, Elliott JA, Wren SB. Sex differences in alcohol preference and drinking patterns emerge during the early postpubertal period. Alcohol Clin Exp Res. 1996;20:1043–1049. doi: 10.1111/j.1530-0277.1996.tb01945.x. [DOI] [PubMed] [Google Scholar]
- Le AD, Israel Y, Juzytsch W, Quan B, Harding S. Genetic selection for high and low alcohol consumption in a limited-access paradigm. Alcohol Clin Exp Res. 2001;25:1613–1620. doi: 10.1111/j.1530-0277.2001.tb02168.x. [DOI] [PubMed] [Google Scholar]
- National Institute on Alcohol Abuse and Alcoholism Alcohol- An important women’s health issue. Alcohol Alert. 2004;62 [Google Scholar]
- Mann K, Ackermann K, Croissant B, Mundle G, Nakovics H, Diehl A. Neuroimaging of gender differences in alcohol dependence: are women more vulnerable? Alcohol Clin Exp Res. 2005;29:896–901. doi: 10.1097/01.alc.0000164376.69978.6b. [DOI] [PubMed] [Google Scholar]
- Moien-Afshari F, Kenyon E, Choy JC, Battistini B, McManus BM, Laher I. Long-term effects of ovariectomy and estrogen replacement treatment on endothelial function in mature rats. Maturitas. 2003;45:213–223. doi: 10.1016/s0378-5122(03)00149-x. [DOI] [PubMed] [Google Scholar]
- McPherson M, Casswell S, Pledger M. Gender convergence in alcohol consumption and related problems: issues and outcomes from comparisons of New Zealand survey data. Addiction. 2004;99:738–748. doi: 10.1111/j.1360-0443.2004.00758.x. [DOI] [PubMed] [Google Scholar]
- Mezey E, Potter JJ. Effect of castration on the turnover of rat liver alcohol dehydrogenase. Biochem Pharmacol. 1985;34:369–372. doi: 10.1016/0006-2952(85)90045-0. [DOI] [PubMed] [Google Scholar]
- Mezey E, Potter JJ, Harmon SM, Tsitouras PD. Effects of castration and testosterone administration on rat liver alcohol dehydrogenase activity. Biochem Pharmacol. 1980;29:3175–3180. doi: 10.1016/0006-2952(80)90582-1. [DOI] [PubMed] [Google Scholar]
- Middaugh LD, Frackelton WF, Boggan WO, Onofrio A, Shepherd CL. Gender differences in the effects of ethanol on C57BL/6 mice. Alcohol. 1992;9:257–260. doi: 10.1016/0741-8329(92)90062-f. [DOI] [PubMed] [Google Scholar]
- Mills KC, Bisgrove EZ. Body sway and divided attention performance under the influence of alcohol: dose-response differences between males and females. Alcohol Clin Exp Res. 1983;7:393–397. doi: 10.1111/j.1530-0277.1983.tb05492.x. [DOI] [PubMed] [Google Scholar]
- Moore EM, Mariani JN, Linsenbardt DN, Melon LC, Boehm SL. Adolescent C57BL/6J (but not DBA/2J) mice consume greater amounts of limited-access ethanol compared to adults and display continued elevated ethanol intake into adulthood. Alcohol Clin Exp Res. 2010;34:734–742. doi: 10.1111/j.1530-0277.2009.01143.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mumenthaler MS, Taylor JL, O’Hara R, Yesavage JA. Gender differences in moderate drinking effects. Alcohol Res Health. 1999;23:55–64. [PMC free article] [PubMed] [Google Scholar]
- Ojeda SR, Skinner MK. Puberty in the Rat. In: Plant TM, editor. Knobil & Neill’s Physiology of Reproduction. Vol. 2, Reproductive Processes and Their Control. Elsevier Academic Press; San Diego: 2006. pp. 2061–2126. [Google Scholar]
- Patton GC, McMorris BJ, Toumbourou JW, Hemphill SA, Donath S, Catalano RF. Puberty and the onset of substance use and abuse. Pediatrics. 2004;114:e300–306. doi: 10.1542/peds.2003-0626-F. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phoenix CH, Goy RW, Gerall AA, Young WC. Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology. 1959;65:369–382. doi: 10.1210/endo-65-3-369. [DOI] [PubMed] [Google Scholar]
- Primus RJ, Kellogg CK. Pubertal-related changes influence the development of environment-related social interaction in the male rat. Dev Psychobiol. 1989;22:633–643. doi: 10.1002/dev.420220608. [DOI] [PubMed] [Google Scholar]
- Primus RJ, Kellogg CK. Gonadal hormones during puberty organize environment-related social interaction in the male rat. Horm Behav. 1990;24:311–323. doi: 10.1016/0018-506x(90)90012-m. [DOI] [PubMed] [Google Scholar]
- Roberts AJ, Smith AD, Weiss F, Rivier C, Koob GF. Estrous cycle effects on operant responding for ethanol in female rats. Alcohol Clin Exp Res. 1998;22:1564–1569. [PubMed] [Google Scholar]
- Rockman GE, Hall A, Markert L, Glavin GB. Early weaning effects on voluntary ethanol consumption and stress responsivity in rats. Physiol Behav. 1987;40:673–676. doi: 10.1016/0031-9384(87)90116-8. [DOI] [PubMed] [Google Scholar]
- Rhodes JS, Ford MM, Yu CH, Brown LL, Finn DA, Garland T, Crabbe JC. Mouse inbred strain differences in ethanol drinking to intoxication. Genes Brain Behav. 2006;6:1–18. doi: 10.1111/j.1601-183X.2006.00210.x. [DOI] [PubMed] [Google Scholar]
- Romeo RD, Schulz KM, Nelson AL, Menard TA, Sisk CL. Testosterone, puberty, and the pattern of male aggression in Syrian hamsters. Dev Psychobiol. 2003;43:102–108. doi: 10.1002/dev.10125. [DOI] [PubMed] [Google Scholar]
- Schulz KM, Richardson HN, Zehr JL, Osetek AJ, Menard TA, Sisk CL. Gonadal hormones masculinize and defeminize reproductive behaviors during puberty in the male Syrian hamster. Horm Behav. 2004;45:242–249. doi: 10.1016/j.yhbeh.2003.12.007. [DOI] [PubMed] [Google Scholar]
- Schulz KM, Sisk CL. Pubertal hormones, the adolescent brain, and the maturation of social behaviors: Lessons from the Syrian hamster. Mol Cell Endocrinol. 2006;254-255:120–126. doi: 10.1016/j.mce.2006.04.025. [DOI] [PubMed] [Google Scholar]
- Shaikh AA, Shaikh SA. Adrenal and ovarian steroid secretion in the rat estrous cycle temporally related to gonadotropins and steroid levels found in peripheral plasma. Endocrinology. 1975;96:37–44. doi: 10.1210/endo-96-1-37. [DOI] [PubMed] [Google Scholar]
- Sherrill LK, Koss WA, Foreman ES, Gulley JM. Effects of pre-pubertal gonadectomy and binge-like ethanol exposure during adolescence on drinking behavior in adult male and female rats. Behav Brain Res. 2011;216:569–575. doi: 10.1016/j.bbr.2010.08.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silveri MM, Spear LP. Ontogeny of ethanol elimination and ethanol-induced hypothermia. Alcohol. 2000;20:45–53. doi: 10.1016/s0741-8329(99)00055-5. [DOI] [PubMed] [Google Scholar]
- Substance Abuse and Mental Health Services Administration . National Survey on Drug Use and Health. U.S. Dept of Health and Human Services, Substance Abuse and Mental Health Services Administration, Office of Applied Studies; 2007. [Google Scholar]
- Substance Abuse and Mental Health Services Administration . National Survey on Drug Use and Health. U.S. Dept of Health and Human Services, Substance Abuse and Mental Health Services Administration, Office of Applied Studies; 2008. [PubMed] [Google Scholar]
- Spear LP. The adolescent brain and age-related behavioral manifestations. Neurosci Biobehav Rev. 2000;24:417–463. doi: 10.1016/s0149-7634(00)00014-2. [DOI] [PubMed] [Google Scholar]
- Takahashi K. On the extra-ovarian origin of estrogen. Tohoku J Exper Med. 1961;75:207–222. doi: 10.1620/tjem.75.207. [DOI] [PubMed] [Google Scholar]
- Tambour S, Brown LL, Crabbe JC. Gender and age at drinking onset affect voluntary alcohol consumption but neither the alcohol deprivation effect nor the response to stress in mice. Alcohol Clin Exp Res. 2008;32:2100–2106. doi: 10.1111/j.1530-0277.2008.00798.x. [DOI] [PubMed] [Google Scholar]
- Truxell EM, Molina JC, Spear NE. Ethanol intake in the juvenile, adolescent, and adult rat: effects of age and prior exposure to ethanol. Alcohol Clin Exp Res. 2007;31:755–765. doi: 10.1111/j.1530-0277.2007.00358.x. [DOI] [PubMed] [Google Scholar]
- Urbano-Marquez A, Estruch R, Fernandez-Sola J, Nicolas JM, Pare JC, Rubin E. The greater risk of alcoholic cardiomyopathy and myopathy in women compared with men. JAMA. 1995;274:149–154. doi: 10.1001/jama.1995.03530020067034. [DOI] [PubMed] [Google Scholar]
- Varlinskaya EI, Doremus-Fitzwater TL, Spear LP. Repeated restraint stress alters sensitivity to the social consequences of ethanol in adolescent and adult rats. Pharmacol Biochem Behav. 2010;96:228–235. doi: 10.1016/j.pbb.2010.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varlinskaya EI, Spear LP. Acute ethanol withdrawal (hangover) and social behavior in adolescent and adult male and female Sprague-Dawley rats. Alcohol Clin Exp Res. 2004;28:40–50. doi: 10.1097/01.ALC.0000108655.51087.DF. [DOI] [PubMed] [Google Scholar]
- Vetter-O’Hagen CS, Spear LP. The effects of gonadectomy on age- and sex-typical patterns of ethanol consumption in Sprague-Dawley rats. Alcohol Clin Exp Res. 2009;33:97A. doi: 10.1111/j.1530-0277.2011.01555.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetter-O’Hagen C, Varlinskaya E, Spear L. Sex differences in ethanol intake and sensitivity to aversive effects during adolescence and adulthood. Alcohol Alcohol. 2009;44:547–554. doi: 10.1093/alcalc/agp048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vetter-O’Hagen CS, Weiss KE, Spear LP. Evidence for the suppressant effects of testosterone on sex-typical ethanol intake in adult male sprague-dawley rats. (under revision) [DOI] [PMC free article] [PubMed]
- Vetter CS, Doremus-Fitzwater TL, Spear LP. Time course of elevated ethanol intake in adolescent relative to adult rats under continuous, voluntary-access conditions. Alcohol Clin Exp Res. 2007;31:1159–1168. doi: 10.1111/j.1530-0277.2007.00417.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang GJ, Volkow ND, Fowler JS, Franceschi D, Wong CT, Pappas NR, Netusil N, Zhu W, Felder C, Ma Y. Alcohol intoxication induces greater reductions in brain metabolism in male than in female subjects. Alcohol Clin Exp Res. 2003;27:909–917. doi: 10.1097/01.ALC.0000071740.56375.BA. [DOI] [PubMed] [Google Scholar]
- Witt ED. Puberty, hormones, and sex differences in alcohol abuse and dependence. Neurotoxicol Teratol. 2007;29:81–95. doi: 10.1016/j.ntt.2006.10.013. [DOI] [PubMed] [Google Scholar]
- Yoneyama N, Crabbe JC, Ford MM, Murillo A, Finn DA. Voluntary ethanol consumption in 22 inbred mouse strains. Alcohol. 2008;42:149–160. doi: 10.1016/j.alcohol.2007.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zucker I. Hormonal determinants of sex differences in saccharin preference, food intake and body weight. Physiol Behav. 1969;4:595–602. [Google Scholar]
- Zucker I, Wade GN, Ziegler R. Sexual and hormonal influences on eating, taste preferences and body weight of hamsters. Physiol Behav. 1972;8:101–111. doi: 10.1016/0031-9384(72)90135-7. [DOI] [PubMed] [Google Scholar]