

Abstract
Objective
Impaired antisaccade performance is a consistent cognitive finding in schizophrenia. Antisaccades require both response inhibition and volitional motor programming, functions that are essential to flexible responding. We investigated whether abnormal timing of hemodynamic responses (HDRs) to antisaccades might contribute to perseveration of ocular motor responses in schizophrenia. We focused on the frontal eye field (FEF), which has been implicated in the persistent effects of antisaccades on subsequent responses in healthy individuals.
Method
Eighteen chronic, medicated schizophrenia outpatients and 15 healthy controls performed antisaccades and prosaccades during functional MRI. Finite impulse response models provided unbiased estimates of event-related HDRs. We compared groups on the peak amplitude, time-to-peak, and full-width half-max of the HDRs.
Results
In patients, HDRs in bilateral FEF were delayed and prolonged but ultimately of similar amplitude to that of controls. These abnormalities were present for antisaccades, but not prosaccades, and were not seen in a control region. More prolonged HDRs predicted slower responses in trials that followed an antisaccade. This suggests that persistent FEF activity following an antisaccade contributes to inter-trial effects on latency.
Conclusions
Delayed and prolonged HDRs for antisaccades in schizophrenia suggest that the functions necessary for successful antisaccade performance take longer to implement and are more persistent. If abnormally persistent neural responses on cognitively demanding tasks are a more general feature of schizophrenia, they may contribute to response perseveration, a classic behavioral abnormality. These findings also underscore the importance of evaluating the temporal dynamics of neural activity to understand cognitive dysfunction in schizophrenia.
Keywords: schizophrenia, antisaccade, frontal eye field, functional MRI, perseveration, inhibition
1. Introduction
Impaired antisaccade performance is consistently observed in schizophrenia. Antisaccades require inhibition of the prepotent response of looking towards a suddenly appearing visual stimulus (i.e., a prosaccade) and the substitution of a gaze in the opposite direction. Thus, antisaccades require both response inhibition and volitional motor programming, which are essential to adaptive, flexible responding. Here, we investigated the neural basis of antisaccade deficits in schizophrenia in relation to perseveration.
In schizophrenia, the antisaccade error rate is reliably increased (Gooding and Basso 2008) and correct antisaccades have longer latencies (Manoach et al 2002; Radant et al 2007; Reuter et al 2007) and more persistent effects on responses in subsequent trials (Barton et al 2005; Barton et al 2006a; Reuter et al 2007). Specifically, while both healthy and schizophrenia participants demonstrate a ‘prior antisaccade effect’ – saccades have longer latencies when preceded by a correct antisaccade than a prosaccade (Barton et al 2006b; Fecteau et al 2004; Lee et al 2011) – in schizophrenia this effect lasts for two trials and only patients are more likely to commit an error in the same direction as the preceding antisaccade (Barton et al 2005). These findings suggest that antisaccades have more persistent effects on the ocular motor system in schizophrenia, resulting in perseveration of responses.
We have hypothesized that these prior antisaccade effects are mediated by the frontal eye field (FEF, Barton et al 2006b; Manoach et al 2007). In monkey studies, FEF neurons show reduced firing in preparation for antisaccades compared to prosaccades, and lower activity predicts longer antisaccade latencies and fewer errors (Everling and Munoz 2000). This reflects a speed-accuracy trade-off: the lower the activity the longer it takes to reach the threshold for triggering a correct antisaccade and the less likely it is that an erroneous prosaccades will escape. In fMRI studies, there is more FEF activation for antisaccades than prosaccades (e.g., Manoach et al 2007; Ford et al 2005b), which has been interpreted to reflect heightened inhibitory input (DeSouza et al 2003). These findings suggest that FEF activity is important both for response inhibition and generating the volitional antisaccade. Evidence from magnetoencephalography (MEG, Lee et al 2011) and fMRI (Manoach et al 2007) studies of healthy individuals supports the hypothesis that preparatory activity in the FEF also mediates prior antisaccade effects on latency. With MEG, FEF activation is greater following antisaccades than prosaccades, and this is sustained throughout the preparatory period of the subsequent trial. With fMRI, FEF activation is reduced during trials preceded by an antisaccade versus a prosaccade. (For a discussion of differences in the direction of signal changes across techniques please see, Lee et al 2011). These findings suggest that the effects of a prior antisaccade persist into the current trial suppressing FEF activity and prolonging latency.
Prior neuroimaging studies report both decreased amplitude of FEF activation during antisaccades in schizophrenia (Camchong et al 2008; Nakashima et al 1994) and no difference (McDowell et al 2002; Raemaekers et al 2002; Tu et al 2006). Here, we studied both the amplitude and temporal dynamics of the FEF hemodynamic response (HDR) to understand these discrepancies and to illuminate the neural bases of slower and more perseverative correct antisaccades in schizophrenia. We expected that FEF activation to antisaccades would be abnormally prolonged in schizophrenia and that this would correlate with the magnitude of prior antisaccade effects. We also expected that longer latencies for correct antisaccades, which suggest slower volitional motor programming, would be accompanied by a later time-to-peak of the HDR in the FEF.
2. Methods
2.1 Participants
Twenty schizophrenia outpatients were recruited from an urban mental health center. They had been maintained on stable doses of atypical antipsychotic medications for at least six weeks, with the exception of one participant taking fluphenazine. Diagnoses were confirmed with Structured Clinical Interviews for DSM-IV (First et al 1997). Clinical status was characterized with the Positive and Negative Syndrome Scale (Kay et al 1987) and the Brief Psychiatric Rating Scale (Overall and Gorham 1962). Sixteen healthy control participants, screened to exclude a personal history of mental illness (SCID-Non-patient edition, First et al 2002) and a family history of schizophrenia spectrum disorder, were recruited from the community. All participants were screened to exclude substance abuse or dependence within the past six months, a history of significant head injury, and neurological illness. The data from two patients and one control could not be used due to eye tracker malfunction. The remaining 18 patients and 15 controls did not differ in age, gender, handedness (White and Ashton 1976), parental socioeconomic status (Hollingshead 1965), or an estimate of pre-morbid verbal IQ (Table 1, Blair and Spreen 1989). The study was approved by the Partners Human Research Committee and participants gave written informed consent.
Table 1.
Means, standard deviations and group comparisons of demographic data and rating scale scores. The Phi value is the result of a Fisher’s Exact Test. The z value is the result of a nonparametric Mann-Whitney U comparison.
| Subject characteristics | Healthy controls (n=15) | Schizophrenia patients (n=18) | t | p |
|---|---|---|---|---|
| Age | 37 ± 10 | 42 ± 11 | −1.50 | 0.14 |
| Sex | 11M/4F | 13M/5F | Phi = 0.01 | 0.99 |
| Laterality Score (Handedness) | 92 ± 11 | 89 ± 13 | 0.59 | 0.56 |
| Parental SES* | 2.4 ± 0.3 | 2.5 ± 0.2 | z = −0.42 | 0.66 |
| Estimated Verbal IQ | 112 ± 8 | 106 ± 13 | 1.66 | 0.11 |
| Age of Onset | 25 ± 2 | |||
| Length of Illness (years) | 17 ± 2 | |||
| Level of Severity | ||||
| BPRS | 18 ± 2 | Minimal | ||
| PANSS positive | 14 ± 1 | Mild | ||
| PANSS negative | 17 ± 2 | Mild | ||
a lower score denotes higher status
2.2 Saccadic Paradigm
Participants practiced in a mock scanner. They were instructed to respond as quickly and accurately as possible by making a saccade to the stimulus on prosaccade trials and in the opposite direction on antisaccade trials. In addition to a base rate of pay, they received 5 cents for each correct response.
The task consisted of a pseudorandom sequence of prosaccade and antisaccade trials that were balanced for right and left movements and 2, 4, and 6s intervals of fixation (Figure 1). The schedule of events was determined using a technique that optimizes the statistical efficiency of event-related designs (Dale 1999). Participants performed six task runs (5m 22s each) that generated a total of 211 prosaccade trials, 211 antisaccade trials, and 80 fixation intervals. The ISCAN fMRI Remote Eye Tracking Laboratory (ISCAN, Burlington, MA) recorded eye position during scanning using a 60Hz video camera.
Figure 1.
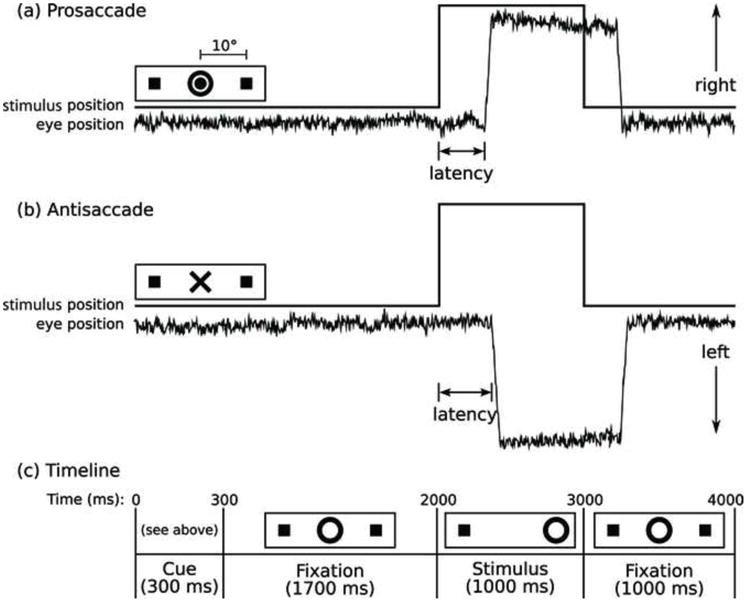
Saccadic paradigm with idealized eye position traces. Saccadic trials lasted 4000 ms and began with an instructional cue at the center of the screen. For half of the participants, orange concentric rings were the cue for a prosaccade trial (A) and a blue X was the cue for an antisaccade trial (B). These cues were reversed for the rest of the participants. The cue was flanked horizontally by two small green squares of 0.2° width that marked the potential locations of stimulus appearance, 10° left and right of center. These squares remained visible throughout the experiment. C: At 300 ms, the instructional cue was replaced by a green fixation ring at the center of the screen, of 0.4° diameter and luminance of 20 cd/m2. After 1700 ms, the ring shifted to one of the two stimulus locations, right or left with equal probability. This was the imperative stimulus to which the participant responded by either making a saccade to it (prosaccade) or to the square on the opposite side (antisaccade). The green ring remained in the peripheral location for 1000 ms and then returned to the center and participants were instructed to follow the ring to the center for 1000 ms of fixation before the start of the next trial. Fixation intervals were simply a continuation of the fixation display that constituted the final second of the previous saccadic trial.
2.3 Analysis of Eye Movements
Eye movements were scored in MATLAB (Mathworks, Natick, MA) using a partially automated program. As in our prior studies (e.g., Manoach et al 2007), saccades were identified as horizontal eye movements with velocities exceeding 47°/s. The onset of a saccade was defined as the point at which the velocity of the eye first exceeded 31°/s. Accuracy was based on whether the initial saccade was in the correct direction. Error rate and the latency of correct saccades with latencies over 130ms (to exclude anticipatory saccades, Fischer and Breitmeyer 1987) were analyzed using randomized block ANOVAs with factors for Group, Task, and subjects nested within group. To examine prior antisaccade effects on the subsequent trial (t+1) we included a factor for the Prior Task. To test whether this effect was more persistent in schizophrenia, lasting for two trials (t+2) rather than one, we included a factor for the Penultimate Task. We also calculated prior antisaccade effects for each participant, defined as the difference in latency between trials preceded by an antisaccade vs. a prosaccade without regard to the identity of the current trial (antisaccade/XX latency – prosaccade/XX) or in the case of the t+2 effect, without regard to the identities of the current or intervening trials (antisaccade/XX/XX latency – prosaccade/XX/XX).
2.4 Image Acquisition
Images were acquired with a 3.0T Siemens Trio whole body high-speed imaging device equipped for echo planar imaging (Erlangen, Germany). Two high-resolution structural images were acquired in the sagittal plane using a high resolution 3D magnetization prepared rapid gradient echo (MPRAGE) sequence (repetition time (TR), 2530ms; echo spacing, 7.25ms; echo time (TE), 3ms; flip angle 7°) with an in-plane resolution of 1mm and 1.3mm slice thickness. Functional images were collected using a gradient echo T2* weighted sequence (TR/TE/Flip = 2000ms/30ms/90°). Twenty contiguous horizontal slices parallel to the intercommissural plane (voxel size: 3.13 × 3.13 × 5mm) were acquired interleaved. The functional sequences included prospective acquisition correction (PACE) for head motion (Thesen et al 2000).
2.5 Analysis of fMRI Data
Analyses used FreeSurfer (Fischl et al 1999) and FreeSurfer Functional Analysis Stream (FS-FAST, Burock and Dale 2000) software. In addition to PACE, functional scans were retrospectively corrected for motion using AFNI (Cox and Jesmanowicz 1999), intensity normalized, and smoothed using a 3D 8mm full-width-half-max Gaussian kernel. Functional images were aligned to the averaged 3D structural image for each participant, which was used to construct inflated (2D) models of individual cortical surfaces. Finite impulse response (FIR) estimates of the HDRs were calculated for each of the four trial types (correct prosaccades, error prosaccades, correct antisaccades, error antisaccades) for each participant. This involved using a linear model to provide unbiased estimates of the average signal intensity at each time point without making a priori assumptions about the shape of the HDR. HDR estimates were computed at 12 time points with an interval of 2s (corresponding to the TR) ranging from 4s prior to the start of a trial to 18s after the start. Temporal correlations in the noise were accounted for by prewhitening using a global estimate of the residual error autocorrelation function truncated at 30s.
To register data across participants, anatomical and functional scans were spatially normalized using a surface-based spherical coordinate system. Registered group data were smoothed with a 2D 4.6mm FWHM Gaussian kernel. fMRI results were displayed on a template brain. Localization of activation was based on an automated parcellation algorithm (Fischl et al 2004). To facilitate comparison with other studies, approximate Talairach coordinates were derived (see supplement).
Group comparisons of activation
To restrict group comparisons to regions activated by the task we created a mask of all positively activated vertices (p<0.001) on the cortical surface in the contrast of all correct saccades versus fixation at 4s, which is the time of peak activation in bilateral FEF in the averaged data of all participants. First, as is standard, we examined group differences in the amplitude of the HDR at 4 s in the contrast of correct antisaccades vs. prosaccades. Second, to determine whether the groups differed in the timing of the HDR, and whether differences were greater for antisaccades, we conducted a three-way ANOVA of Group, Task, and Time (0, 2, 4, 6, 8, 10s) at each vertex. For regions showing a significant Group by Task by Time interaction, we calculated the mean percent signal change at each time point for antisaccades and prosaccades and used the ideal sinc interpolation kernel to construct the HDR functions for each task in each participant. From these HDR functions, we derived three parameters – time-to-peak, peak amplitude (this differs from the standard amplitude analysis since the time of the peak was allowed to vary), and full-width half-max – using the mspeaks function in MATLAB’s Bioinformatics Toolbox (The Mathworks, Inc., Natick, MA). HDR parameters were compared between groups using t-tests.
Relations of HDR parameters with performance
Regressions of performance on HDR parameters included an interaction term of the HDR parameter by group to test whether the slope of the relation differed by group.
Correction for multiple comparisons
We used a clustering method based on 10,000 Monte Carlo simulations of synthesized white Gaussian noise using the smoothing, resampling, and averaging parameters of the functional analyses in the region of interest mask to determine the likelihood that a cluster of a certain size would be found by chance for a given threshold (p≤.05) and to provide cluster-wise probability values (CWP).
Control analyses
Methods for comparing motion between groups and assessing the effects of antipsychotic medications and age on activation are described in the supplement.
3. Results
3.1 Performance
Participants made more antisaccade than prosaccade errors (F(1,31)=23.58, p< 0.0001) and patients made more errors than controls (F(1,31)=8.19, p=0.007). The group by task interaction (F(1,31)=4.45, p=0.04) reflected that patients made significantly more antisaccade (21±17 vs. 9±6%; t(31)=3.55, p=0.001) but not prosaccade (6±7 vs. 3±2%; t(31)=0.84, p=0.41) errors than controls.
Latency data from one patient and two controls could not be scored reliably, and were excluded from analyses. Antisaccades had longer latencies than prosaccades (F(1,28)=108.10, p< 0.0001) and patients showed a trend toward longer latencies than controls (F(1,28)=3.18, p=0.09; antisaccades: 332±70 vs. 293±41ms, t(28)=2.01, p=0.05; prosaccades: 268±99 vs. 240±68ms, t(28)=1.41, p=.17) that did not differ significantly by task (F(1,28)=1.08, p=.31).
A prior antisaccade vs. prosaccade slowed the latency of the current trial (F(1,28)=11.51, p=0.002). As in our prior study (Barton et al 2005), group membership did not affect the magnitude of the prior antisaccade effect (F(1,28)=0.94, p=0.34). Contrary to our expectations, we did not detect significant prior antisaccade effects two trials later (t+2) in either group.
3.2 fMRI data
Amplitude at 4 s
Relative to controls, patients showed significantly reduced activation in the left FEF in the contrast of antisaccades vs. prosaccades at 4s (−26, 2, 46; cluster size=722mm2; CWP=0.0001). There were no other significant group differences.
HDR Timing
Significant Group by Task by Time interactions were observed in the left FEF (Figure 2; −23, −1, 46; cluster size=1183mm2; CWP=0.0001) and the right supplementary eye field (SEF; 9, 2, 48; cluster size=853mm2; CWP=0.001).
Figure 2.
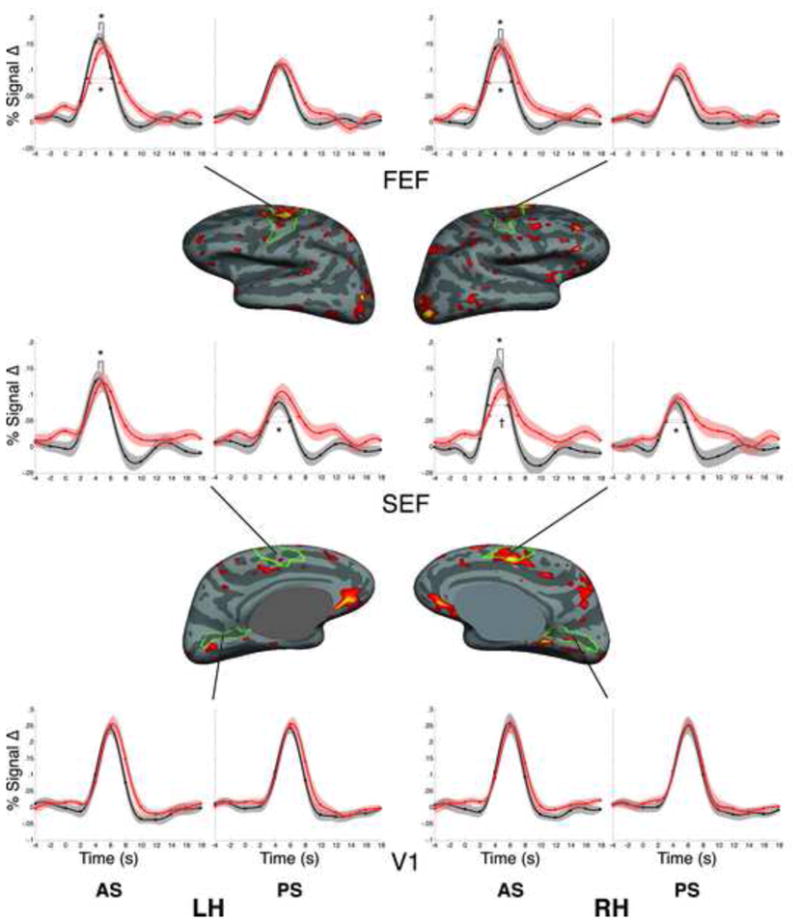
Statistical map of the Group by Time by Task interaction on the inflated medial and lateral cortical surfaces. Clusters in left FEF and right SEF survived correction for multiple comparisons. HDR time course graphs (with standard errors) are displayed for each condition (prosaccades and antisaccades) relative to the fixation baseline for bilateral FEF and SEF, as well as the control region V1. Red lines represent patient data, black lines represent healthy control data. Time in seconds is on the x-axis and percent signal change relative to the fixation baseline is on the y-axis. Significant group differences in ‘time-to-peak’ and full-width half-max are denoted by asterisks (*). Trends are denoted by the symbol †.
To understand these interactions, we calculated the HDR parameters for each trial type in the FEF, SEF, and in a comparison region, primary visual cortex (V1), which is activated by the task but is not involved in ocular motor control. As we did not expect lateralized differences, we examined the HDR parameters in both hemispheres. Within our region of interest mask, we used anatomical criteria to define these regions. We restricted the FEF to vertices in and around the precentral sulcus and gyrus, excluding the medial surface (Paus 1996), the SEF to the medial surface, dorsal to the cingulate sulcus, posterior to the VCA line and anterior to the precentral sulcus (Grosbras et al 1999), and V1 to the calcarine sulcus. Three patients were missing data for prosaccades in the SEF because no peak was detected.
The groups did not differ in peak amplitude in any region for antisaccades or prosaccades. However, patients showed a significantly later time-to-peak in bilateral FEF and SEF for antisaccades, but not for prosaccades (Table 2, Figure 2). In addition, in bilateral FEF, the HDRs for patients were significantly prolonged for antisaccades, but not prosaccades, as indexed by the full-width-half-max. Patients also showed prolonged HDRs bilaterally in the SEF for prosaccades, but this finding was limited by the fact that no peak was detected in the SEF for prosaccades in three patients. There were no group differences in HDR parameters in V1.
Table 2.
Means, standard deviations and group comparisons of peak amplitude, time-to-peak, and full-width half-max (FWHM) of HDRs to antisaccades and prosaccades by region and hemisphere.
| Peak Amplitude | Time-to-peak | FWHM | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ROI | Task | Hemi | HC | SZ | t | p | HC | SZ | t | p | HC | SZ | t | p |
| FEF | AS | LH | .18 ± .05 | .16 ± 05 | 1.14 | .26 | 4.56 ± .64 | 5.21 ± .83 | −2.46 | .02 | 3.67 ± .97 | 4.49 ± 1.3 | −2.01 | .05 |
| RH | .17 ± .06 | .16 ± .06 | 0.25 | .81 | 4.61± .76 | 5.18 ± .84 | −2.03 | .05 | 3.59 ± .64 | 4.31 ± 1.22 | −2.06 | .05 | ||
| PS | LH | .12 ± .03 | .13 ± .03 | −0.60 | .55 | 4.55 ± .79 | 4.83 ± 1.62 | −.60 | .55 | 3.70 ± .90 | 4.19 ± 1.23 | −1.29 | .21 | |
| RH | .11 ± .03 | .12 ± .04 | −0.65 | .52 | 4.86 ± 1.60 | 4.94 ± .82 | −.19 | .86 | 3.55 ± .89 | 4.22 ± 1.36 | −1.63 | .11 | ||
| SEF | AS | LH | .15 ± .07 | .14 ± .07 | 0.34 | .73 | 4.52 ± .75 | 5.24 ± .91 | −2.45 | .02 | 3.33 ± .96 | 4.05 ± 1.89 | −1.33 | .19 |
| RH | .17 ± .09 | .14 ± .06 | 1.25 | .22 | 4.48 ± .67 | 5.26 ± 1.16 | −2.29 | .03 | 3.16 ± .73 | 4.00 ± 1.66 | −1.83 | .08 | ||
| PS* | LH | .11 ± .05 | .12 ± .05 | −0.93 | .36 | 5.14 ± 1.66 | 5.22 ± 1.03 | −.16 | .87 | 3.27 ± .93 | 4.14 ± 1.29 | −2.14 | .04 | |
| RH | .11 ± .05 | .11 ± .04 | 0.05 | .96 | 5.01 ± 1.74 | 5.30 ± 1.33 | −.52 | .61 | 3.07 ± .89 | 3.99 ± .78 | −3.07 | .005 | ||
| V1 | AS | LH | .27 ± .06 | .27 ± .10 | −0.26 | .80 | 5.96 ± .78 | 6.33 ± .65 | −1.49 | .15 | 3.50 ± .71 | 3.66 ± .67 | −.66 | .51 |
| RH | .28 ± .10 | .27 ± .10 | 0.43 | .67 | 5.84 ± .76 | 6.23 ± .58 | −1.65 | .11 | 3.59 ± .64 | 3.93 ± .90 | −1.20 | .24 | ||
| PS | LH | .26 ± .06 | .28 ± .08 | −0.42 | .68 | 5.93 ± .72 | 6.26 ± .63 | −1.39 | .17 | 3.65 ± .68 | 3.80 ± .82 | −.57 | .57 | |
| RH | .27 ± .10 | .27 ± .07 | 0.08 | .94 | 5.84 ± .77 | 6.23 ± .60 | −1.61 | .12 | 3.77 ± .59 | 3.90 ± .96 | −.48 | .63 | ||
In three patients no peak was detected in the SEF for prosaccades, one in both hemispheres, one in the right hemisphere, and one in the left hemisphere.
Relations of FEF HDR parameters with performance
Time-to-peak did not correlate with antisaccade latency, and this did not differ by group. In the left FEF we observed a significant correlation of the prior antisaccade effect (t+1) with the full-width half-max (Figure 3, t(27)=2.05, p=.05) that did not differ by group (t(27)= −0.07, p=.94). A similar, non-significant effect was seen in the right FEF. Finally, we examined whether FWHM predicted the magnitude of the prior antisaccade effect two trails later (t+2) in schizophrenia. We observed a trend in the right FEF (t(16)=1.65, p=.10) and a weaker relation in left FEF (t(16)=1.20, p=.22). There were no relations in controls (p’s>.8).
Figure 3.
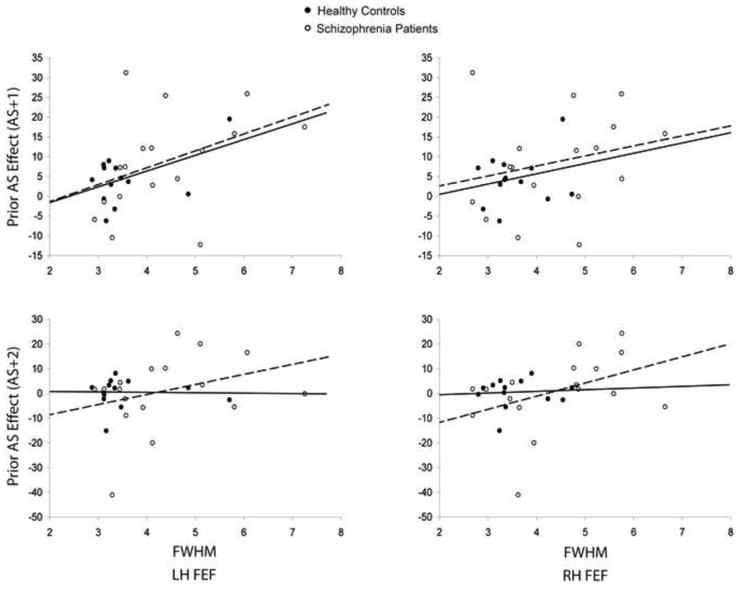
Scatter plots of the relations of the full-width half-max (FWHM) in left and right FEF with the prior antisaccade effect on latency by group with regression lines. Healthy controls are represented by filled black circles and solid lines, and schizophrenia patients are represented by open circles and dashed lines.
Control analyses
Please see supplement for details. The groups did not differ in mean motion. Neither age nor antipsychotic medication dosage was significantly related to any FEF HDR parameter.
4. Discussion
Schizophrenia patients showed abnormally delayed and persistent HDRs in the FEF during antisaccades. This suggests that the processes necessary for successful antisaccade performance are implemented more slowly and are more persistent. That the HDR was delayed, but ultimately of similar amplitude in schizophrenia, underscores the importance of evaluating the temporal dynamics of neural activity to understanding the basis of cognitive dysfunction. If slower, more prolonged neural responses to cognitively demanding tasks are a more general feature of schizophrenia, it may be difficult to rapidly adjust behavior in response to the demands of the moment. Such dynamic adjustments are fundamental to adaptive, flexible behavior and impairments may contribute to perseveration.
As in prior studies, we observed similar significant prior antisaccade effects on saccadic latency for patients and controls – an antisaccade slowed responses in the subsequent trial (Barton et al 2005; Manoach et al 2002). This effect correlated with more prolonged FEF HDRs, which along with the findings of our paired MEG and fMRI studies (Lee et al 2011; Manoach et al 2007), suggests that persistent activity of the FEF following an antisaccade contributes to inter-trial effects on latency. In the present study, we failed to replicate our behavioral finding of a more prolonged prior antisaccade effect on latency in schizophrenia (i.e., that an antisaccade slowed latency two trials later, Barton et al 2005), and this ‘t + 2’ effect only weakly correlated with the more prolonged HDRs for antisaccades (i.e., trend in right FEF, n.s. in left). This failure to replicate the behavioral finding may reflect the inferior temporal resolution of the scanner-based eye tracking system of the present study (60 vs. 1000Hz), which may have been inadequate to detect a small effect on latency (i.e., 20ms in the prior study). A higher resolution eye tracker may be necessary to evaluate the hypothesis that the more prolonged HDR in the FEF during antisaccades in schizophrenia is a neural correlate of the behavioral perseveration that we previously observed (Barton et al 2005; Barton et al 2006a).
Patients also showed a later peak of activation in the FEF for correct antisaccades. This might reflect less efficient implementation of inhibition and/or slower activation of the antisaccade task goal (e.g., Reuter et al 2007), both of which have been proposed to account for impaired antisaccade performance in schizophrenia. The present findings cannot discriminate between these possibilities.
In addition to the delayed peak of the HDR, correct antisaccades were performed more slowly in patients. This is consistent with prior reports of delayed HDRs in the context of slower manual reaction times (Ford et al 2005a; Velanova et al 2007; Yarkoni et al 2009). Prior fMRI studies of schizophrenia have reported both delayed activation in several regions during a visual oddball task (Ford et al 2005a) and normal HDR peak times in primary sensorimotor and supplementary motor regions during a flashing checkerboard (Barch et al 2003). Our findings of normal HDRs in V1 and delayed HDRs in ocular motor regions are consistent with both of these reports. The shape of the HDR has been shown to vary across brain regions, stimulus types, individuals, and groups (Duann et al 2002; Ford et al 2005a; Miezin et al 2000; Velanova et al 2007). Thus, whether HDR timing abnormalities are observed in schizophrenia may depend on both the region and the type of cognitive demand. Greater demand may lead to delayed and prolonged neural responses in higher order regions and, consequently, to slower, more perseverative responses.
The finding of delayed HDRs in schizophrenia resonates with numerous reports of delayed cognitive event-related potentials. The relative dearth of prior reports of delayed HDRs may reflect that most standard fMRI analysis techniques assume a shape to the HDR and only test for group differences in amplitude. Had we limited our investigation to amplitude at the time peak of the HDR in the combined group as is standard, we might have erroneously concluded that patients showed significantly reduced FEF activation. Instead, this reduction reflected a delayed peak in patients, and, at later time points (i.e., 8 and 10s) patients actually showed significantly greater activation than controls, which reflected more prolonged activation rather than increased amplitude. This suggests that a failure to consider the timing of the HDR may lead to erroneous conclusions and contribute to inconsistent findings concerning fMRI activation differences in schizophrenia.
A limitation to the present study is that all of the patients were chronically ill and taking antipsychotic medications. Although controlling for dosage did not alter the findings, this does not exclude a confounding effect of medications. Even if the abnormalities observed were due to medication, progression, or epiphenomena of long-term illness, they remain relevant since the vast majority of patients with schizophrenia are chronic and medicated.
In summary, we propose that delayed and prolonged HDRs in the FEF during antisaccades in schizophrenia contribute to slower and more perseverative ocular motor responses. If delayed and prolonged neural responses to cognitively demanding tasks are a more general feature of schizophrenia, they may contribute to cognitive slowing and to responses that are unduly influenced by the recent past at the expense of more immediate demands.
Supplementary Material
Acknowledgments
Role of Funding Source: Support for this study was provided by National Institute for Mental Health R01 MH67720, F32 MH082514, F32 MH088081; National Alliance for Research on Schizophrenia and Depression; Mental Illness and Neuroscience Discovery (MIND) Institute; National Center for Research Resources P41RR14075; Canada Research Chair and CIHR grant MOP-81270. These agencies had no further role in study design; in the collection, analysis and interpretation of data; in the writing of the report; and/or in the decision to submit the paper for publication.
Footnotes
Previous presentation: Society for Neuroscience, San Diego, November 13–17, 2010
Contributors: Drs. Dyckman, Lee, and Agam contributed to data collection, analysis, and manuscript preparation. Dr. Vangel provided statistical consultation. Dr. Goff provided patient characterization, assisted with recruitment, and consulted regarding the findings. Dr. Barton participated in task design and the conceptualization of the findings. Dr. Manoach oversaw and participated in all aspects of the study. All authors contributed to and have approved the final manuscript.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Barch DM, Mathews JR, Buckner RL, Maccotta L, Csernansky JG, Snyder AZ. Hemodynamic responses in visual, motor, and somatosensory cortices in schizophrenia. Neuroimage. 2003;20:1884–1893. doi: 10.1016/s1053-8119(03)00449-x. [DOI] [PubMed] [Google Scholar]
- Barton JJ, Cherkasova MV, Lindgren KA, Goff DC, Manoach DS. What is perseverated in schizophrenia? Evidence of abnormal response plasticity in the saccadic system. J Abnorm Psychol. 2005;114:75–84. doi: 10.1037/0021-843X.114.1.75. [DOI] [PubMed] [Google Scholar]
- Barton JJ, Goff DC, Manoach DS. The inter-trial effects of stimulus and saccadic direction on prosaccades and antisaccades, in controls and schizophrenia patients. Exp Brain Res. 2006a;174:487–498. doi: 10.1007/s00221-006-0492-9. [DOI] [PubMed] [Google Scholar]
- Barton JJ, Greenzang C, Hefter R, Edelman J, Manoach DS. Switching, plasticity, and prediction in a saccadic task-switch paradigm. Exp Brain Res. 2006b;168:76–87. doi: 10.1007/s00221-005-0091-1. [DOI] [PubMed] [Google Scholar]
- Blair JR, Spreen O. Predicting premorbid IQ: a revision of the National Adult Reading Test. Clin Neuropsychologist. 1989;3:129–136. [Google Scholar]
- Burock MA, Dale AM. Estimation and detection of event-related fMRI signals with temporally correlated noise: a statistically efficient and unbiased approach. Hum Brain Mapp. 2000;11:249–260. doi: 10.1002/1097-0193(200012)11:4<249::AID-HBM20>3.0.CO;2-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Camchong J, Dyckman KA, Austin BP, Clementz BA, McDowell JE. Common neural circuitry supporting volitional saccades and its disruption in schizophrenia patients and relatives. Biol Psychiatry. 2008;64:1042–1050. doi: 10.1016/j.biopsych.2008.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox RW, Jesmanowicz A. Real-time 3D image registration for functional MRI. Magn Reson Med. 1999;42:1014–1018. doi: 10.1002/(sici)1522-2594(199912)42:6<1014::aid-mrm4>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- Dale AM. Optimal experimental design for event-related fMRI. Hum Brain Mapp. 1999;8:109–140. doi: 10.1002/(SICI)1097-0193(1999)8:2/3<109::AID-HBM7>3.0.CO;2-W. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeSouza JF, Menon RS, Everling S. Preparatory set associated with pro-saccades and anti-saccades in humans investigated with event-related FMRI. J Neurophysiol. 2003;89:1016–1023. doi: 10.1152/jn.00562.2002. [DOI] [PubMed] [Google Scholar]
- Duann JR, Jung TP, Kuo WJ, Yeh TC, Makeig S, Hsieh JC, Sejnowski TJ. Single-trial variability in event-related BOLD signals. Neuroimage. 2002;15:823–835. doi: 10.1006/nimg.2001.1049. [DOI] [PubMed] [Google Scholar]
- Everling S, Munoz DP. Neuronal correlates for preparatory set associated with pro-saccades and anti-saccades in the primate frontal eye field. J Neurosci. 2000;20:387–400. doi: 10.1523/JNEUROSCI.20-01-00387.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fecteau JH, Au C, Armstrong IT, Munoz DP. Sensory biases produce alternation advantage found in sequential saccadic eye movement tasks. Exp Brain Res. 2004;159:84–91. doi: 10.1007/s00221-004-1935-9. [DOI] [PubMed] [Google Scholar]
- First MB, Spitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for DSM-IV Axis I Disorders, Research Version, Patient Edition with Psychotic Screen (SCID-I/P W/PSY SCREEN) New York: Biometrics Research, New York State Psychiatric Institute; 1997. [Google Scholar]
- First MB, Spitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for DSM-IV-TR Axis I Disorders, Research Version. New York: Biometrics Research, New York State Psychiatric Institute; 2002. Nonpatient Edition. [Google Scholar]
- Fischer B, Breitmeyer B. Mechanisms of visual attention revealed by saccadic eye movements. Neuropsychologia. 1987;25:73–83. doi: 10.1016/0028-3932(87)90044-3. [DOI] [PubMed] [Google Scholar]
- Fischl B, Sereno MI, Dale AM. Cortical surface-based analysis. II: Inflation, flattening, and a surface-based coordinate system. Neuroimage. 1999;9:195–207. doi: 10.1006/nimg.1998.0396. [DOI] [PubMed] [Google Scholar]
- Fischl B, van der Kouwe A, Destrieux C, Halgren E, Segonne F, Salat DH, et al. Automatically parcellating the human cerebral cortex. Cereb Cortex. 2004;14:11–22. doi: 10.1093/cercor/bhg087. [DOI] [PubMed] [Google Scholar]
- Ford JM, Johnson MB, Whitfield SL, Faustman WO, Mathalon DH. Delayed hemodynamic responses in schizophrenia. Neuroimage. 2005a;26:922–931. doi: 10.1016/j.neuroimage.2005.03.001. [DOI] [PubMed] [Google Scholar]
- Ford KA, Goltz HC, Brown MR, Everling S. Neural processes associated with antisaccade task performance investigated with event-related FMRI. J Neurophysiol. 2005b;94:429–440. doi: 10.1152/jn.00471.2004. [DOI] [PubMed] [Google Scholar]
- Gooding DC, Basso MA. The tell-tale tasks: a review of saccadic research in psychiatric patient populations. Brain Cogn. 2008;68:371–390. doi: 10.1016/j.bandc.2008.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grosbras MH, Lobel E, Van de Moortele PF, LeBihan D, Berthoz A. An anatomical landmark for the supplementary eye fields in human revealed with functional magnetic resonance imaging. Cereb Cortex. 1999;9:705–711. doi: 10.1093/cercor/9.7.705. [DOI] [PubMed] [Google Scholar]
- Hollingshead AB. Two factor index of social position. New Haven, Conn: Yale University Press; 1965. [Google Scholar]
- Kay SR, Fiszbein A, Opler LA. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull. 1987;13:261–276. doi: 10.1093/schbul/13.2.261. [DOI] [PubMed] [Google Scholar]
- Lee AKC, Hämäläinen MS, Dyckman KA, Barton JJS, Manoach DS. Saccadic preparation in frontal eye field is modulated by distinct trial history effects as revealed by magnetoencephalography. Cerebral Cortex. 2011;21:245–253. doi: 10.1093/cercor/bhq057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manoach DS, Lindgren KA, Cherkasova MV, Goff DC, Halpern EF, Intriligator J, Barton JJS. Schizophrenic subjects show deficient inhibition but intact task-switching on saccadic tasks. Biol Psychiatry. 2002;51:816–826. doi: 10.1016/s0006-3223(01)01356-7. [DOI] [PubMed] [Google Scholar]
- Manoach DS, Thakkar KN, Cain MS, Polli FE, Edelman JA, Fischl B, Barton JJ. Neural activity is modulated by trial history: a functional magnetic resonance imaging study of the effects of a previous antisaccade. J Neurosci. 2007;27:1791–1798. doi: 10.1523/JNEUROSCI.3662-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell JE, Brown GG, Paulus M, Martinez A, Stewart SE, Dubowitz DJ, Braff DL. Neural correlates of refixation saccades and antisaccades in normal and schizophrenia subjects. Biol Psychiatry. 2002;51:216–223. doi: 10.1016/s0006-3223(01)01204-5. [DOI] [PubMed] [Google Scholar]
- Miezin FM, Maccotta L, Ollinger JM, Petersen SE, Buckner RL. Characterizing the hemodynamic response: effects of presentation rate, sampling procedure, and the possibility of ordering brain activity based on relative timing. Neuroimage. 2000;11:735–759. doi: 10.1006/nimg.2000.0568. [DOI] [PubMed] [Google Scholar]
- Nakashima Y, Momose T, Sano I, Katayama S, Nakajima T, Niwa S, Matsushita M. Cortical control of saccade in normal and schizophrenic subjects: a PET study using a task-evoked rCBF paradigm. Schizophr Res. 1994;12:259–264. doi: 10.1016/0920-9964(94)90036-1. [DOI] [PubMed] [Google Scholar]
- Overall JE, Gorham DR. The brief psychiatric rating scale. Psychol Reports. 1962;10:799–812. [Google Scholar]
- Paus T. Location and function of the human frontal eye-field: a selective review. Neuropsychologia. 1996;34:475–483. doi: 10.1016/0028-3932(95)00134-4. [DOI] [PubMed] [Google Scholar]
- Radant AD, Dobie DJ, Calkins ME, Olincy A, Braff DL, Cadenhead KS, et al. Successful multi-site measurement of antisaccade performance deficits in schizophrenia. Schizophr Res. 2007;89:320–329. doi: 10.1016/j.schres.2006.08.010. [DOI] [PubMed] [Google Scholar]
- Raemaekers M, Jansma JM, Cahn W, Van der Geest JN, van der Linden JA, Kahn RS, Ramsey NF. Neuronal substrate of the saccadic inhibition deficit in schizophrenia investigated with 3-dimensional event-related functional magnetic resonance imaging. Arch Gen Psychiatry. 2002;59:313–320. doi: 10.1001/archpsyc.59.4.313. [DOI] [PubMed] [Google Scholar]
- Reuter B, Jager M, Bottlender R, Kathmann N. Impaired action control in schizophrenia: the role of volitional saccade initiation. Neuropsychologia. 2007;45:1840–1848. doi: 10.1016/j.neuropsychologia.2006.12.006. [DOI] [PubMed] [Google Scholar]
- Thesen S, Heid O, Mueller E, Schad LR. Prospective acquisition correction for head motion with image-based tracking for real-time fMRI. Magn Reson Med. 2000;44:457–465. doi: 10.1002/1522-2594(200009)44:3<457::aid-mrm17>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- Tu PC, Yang TH, Kuo WJ, Hsieh JC, Su TP. Neural correlates of antisaccade deficits in schizophrenia, an fMRI study. J Psychiatr Res. 2006;40:606–612. doi: 10.1016/j.jpsychires.2006.05.012. [DOI] [PubMed] [Google Scholar]
- Velanova K, Lustig C, Jacoby LL, Buckner RL. Evidence for frontally mediated controlled processing differences in older adults. Cereb Cortex. 2007;17:1033–1046. doi: 10.1093/cercor/bhl013. [DOI] [PubMed] [Google Scholar]
- White K, Ashton R. Handedness assessment inventory. Neuropsychologia. 1976;14:261–264. doi: 10.1016/0028-3932(76)90058-0. [DOI] [PubMed] [Google Scholar]
- Yarkoni T, Barch DM, Gray JR, Conturo TE, Braver TS. BOLD correlates of trial-by-trial reaction time variability in gray and white matter: a multi-study fMRI analysis. PLoS One. 2009;4:e4257. doi: 10.1371/journal.pone.0004257. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.