

Abstract
Background
The chromosomal location of CUL-5 (11q 22-23) is associated with LOH in breast cancer, suggesting that CUL-5 may be a tumor suppressor. The purpose of this research was to determine if there is differential expression of CUL-5 in breast epithelial cells versus breast cancer cell lines, and normal human tissues versus human tumors. The expression of CUL-5 in breast epithelial cells (HMEC, MCF-10A), and breast cancer cells (MCF-7, MDA-MB-231) was examined using RT-PCR, Northern blot analysis, and Western blot analysis. The expression of mRNA for other CUL family members (CUL-1, -2, -3, -4A, and -4B) in these cells was evaluated by RT-PCR. A normal human tissue expression array and a cancer profiling array were used to examine CUL-5 expression in normal human tissues and matched normal tissues versus tumor tissues, respectively.
Results
CUL-5 is expressed at the mRNA and protein levels by breast epithelial cells (HMEC, MCF-10A) and breast cancer cells (MCF-7, MDA-MB-231). These cells also express mRNA for other CUL family members. The normal human tissue expression array revealed that CUL-5 is widely expressed. The cancer profiling array revealed that 82% (41/50) of the breast cancers demonstrated a decrease in CUL-5 expression versus the matched normal tissue. For the 50 cases of matched breast tissue there was a statistically significant ~2.2 fold decreased expression of CUL-5 in tumor tissue versus normal tissue (P < 0.0001).
Conclusions
The data demonstrate no apparent decrease in CUL-5 expression in the breast cancer cell lines (MCF-7, MDA-MB-231) versus the breast epithelial cells (HMEC, MCF-10A). The decrease in CUL-5 expression in breast tumor tissue versus matched normal tissue supports the hypothesis that decreased expression of CUL-5 may play a role in breast tumorigenesis.
Background
The chromosomal region 11q22-23 is associated with LOH in breast cancer [1-8], suggesting that a gene or genes at this chromosomal region may play a role as a tumor suppressor. One study demonstrated that frequency of LOH at 11q22-23 is 38% in primary breast tumors and 84% in local recurrences after surgery [8]. One candidate gene located at this chromosomal region is CUL-5 [7,8]. CUL-5, first described as a gene product involved in mediating vasopressin-induced increases of intracellular-free calcium in renal medullary epithelial cells [9], belongs to an evolutionarily conserved gene family named the Cullins (CULs) [10,11]. To date, seven mammalian CULs have been identified and designated CUL-1, CUL-2, CUL-3, CUL-4A, CUL-4B, CUL-5, and CUL-7 [10-13].
CUL proteins appear to function as scaffolding proteins within E3 ubiquitin ligase complexes [14], which assist in targeting protein substrates for ubiquitin-mediated degradation by the 26S proteasome [15,16]. The mechanism of targeted degradation consists of three enzymatic steps [15,16]. In step one, an E1 enzyme activates the 76-amino acid protein ubiquitin to a high-energy thiol ester intermediate. In step two, E2 enzyme, referred to as an ubiquitin-conjugating enzyme, assists in the transfer of the activated ubiquitin to a lysine residue on the substrate protein that is bound to the E3 ubiquitin ligase complex. Finally, the substrate protein is polyubiquitinated with a chain a ubiquitins, which is the signal for it to be recognized and degraded by the 26S proteasome. The CUL-based E3 ubiquitin ligases consist of a CUL protein, a RING finger protein (e.g. ROC1/Rbx1, ROC2), an E2 ubiquitin-conjugating enzyme (e.g. Cdc34, Ubc5), an adaptor (e.g. SKP-1, Elongin C), and a substrate recognition protein (e.g. F-box proteins, BC-box proteins) [15,16]. CUL proteins function in E3 ubiquitin ligase complexes as scaffolds that bring together the ubiquitin conjugating and substrate recognition components of these E3 complexes.
CUL-5 has been suggested to be a tumor suppressor in breast tissue despite a lack of information concerning the expression and function of this gene during breast tumorigenesis. The main purpose of this study was to examine the expression of CUL-5 in breast epithelial cells versus breast cancer cell lines and in normal tissues versus matched tumor tissues. Here, we report that MCF-7 and MDA-MB-231 breast cancer cell lines do not demonstrate an apparent decrease in the expression of CUL-5 mRNA or protein when compared to HMEC and MCF-10A breast epithelial cells. However, analysis of 50 cases of matched normal versus breast tumor tissue revealed a statistically significant ~2.2 fold decrease in CUL-5 expression in the tumor tissue versus the matched normal tissue.
Results
Expression of CUL-5 mRNA and protein in human breast epithelial cells and breast cancer cell lines
As shown in Figure 1A, RT-PCR demonstrated that CUL-5 mRNA is expressed by primary HMECs, immortalized but non-tumorigenic MCF-10A cells, and both ER-α positive (MCF-7) and ER-α negative (MDA-MB-231) human breast cancer cell lines, as evidenced by amplification of the predicted 674-bp PCR product. The 674-bp CUL-5 PCR product was not evident in the NT controls (Figure 1A) or when reverse transcriptase was eliminated from the reaction mixture (data not shown). The integrity of the RNA samples was confirmed by amplifying a 411-bp PCR product for ribosomal protein S9 (Figure 1B). The identity of the CUL-5 PCR products from these cells were confirmed by restriction endonuclease digestion with EcoRI, which resulted in the generation of two smaller DNA fragments of the predicted sizes (Table 1 and Figure 1C), and by DNA sequencing (data not shown). Expression of mRNAs for other CUL family members in these same cells was demonstrated by RT-PCR amplification of cDNAs for CUL-1, -2, -3, -4A, and -4B (Figure 2). PCR products for these other CUL family members were not amplified when reverse transcriptase was not included in the reaction mixture (data not shown). PCR products for CUL-1, -2, -3, -4A, and -4B were confirmed by restriction endonuclease digestion with BglII, AatII, EcoRI, Sau3AI and HaeIII, respectively (data not shown). The RT-PCR results for CUL-5 mRNA expression were confirmed by Northern blot analysis, which demonstrated one major transcript of ~7.4 kb and two minor transcripts were observed below the 4.4 kb marker (Figure 3). To examine expression of CUL-5 protein, Western blots were performed using affinity-purified rabbit polyclonal antiserum generated in rabbits immunized with a synthetic CUL-5 peptide. Western blot analysis revealed a major immunoreactive band of Mr ~ 82 kDa, and a minor band of Mr ~ 63 kDa (Figure 4). These bands were not seen when antibody was preincubated with immunizing peptide or when preimmune serum was used (data not shown). Blots were stripped and reprobed with anti-actin antibody to ensure the integrity of the protein samples and the Western blotting procedure (Figure 4).
Figure 1.

RT-PCR analysis for CUL-5 in HMEC, MCF-10A, MCF-7, and MDA-MB-231 cells. Semiquantitative RT-PCR was performed using 1 μg of total RNA with 20, 25, and 29 cycles of PCR using primers for (A) CUL-5, or (B) ribosomal protein S9; (C) The CUL-5 PCR products from HMEC, MCF-10A, MCF-7, and MDA-MB-231 cells were confirmed by restriction endonuclease digestion with EcoRI. The results shown are representative of triplicate experiments using independent RNA samples.
Table 1.
Primers used for RT-PCR analysis of CUL family members
| Primer Sequences | Primer Location and GenBank Accession # |
Annealing Temp. and Predicted product size |
Restriction endonuclease and cut site |
|
CUL-1 forward: 5'-CGCTGGCTTTGTGGCTGCTC-3' CUL-1 reverse: 5'-TGTGGCGGCTGGCGTAGAA-3' |
nt 1510 – 1529 nt 2051 – 2069 XM027270 |
58°C 560-bp |
BglII 1891 |
|
CUL-2 forward: 5'-CAGCGAAAGGGATGACAGAGAATG-3' CUL-2 reverse: 5'-TGAGTAAGAGGCCACGCACCA-3' |
nt 1351 – 1374 nt 1688 – 1708 XM005814.3 |
54°C 358-bp |
AatII 1435 |
|
CUL-3 forward: 5'-AGAGCGGAAAGGAGAAGTCGTAGA-3' CUL-3 reverse: 5'-CTCAAAGTCACCCGCAATAGTT-3' |
nt 833 – 856 nt 1424 – 1445 XM051985.1 |
53°C 613-bp |
EcoRI 1160 |
|
CUL-4A forward: 5'-AACTCCACGCTGCCCTCCATCTG-3' CUL-4A reverse: 5'-TGCTGCCCGCCCCTCACC-3' |
nt 346 – 368 nt 840 – 857 XM027482.1 |
58°C 512-bp |
Sau3AI 452 |
|
CUL-4B forward: 5'-GCGGAGCTGCTTTCACCA-3' CUL-4B reverse: 5'-TTCTTCAACCGTTTCTTTCATCTG-3' |
nt 1400 – 1417 nt 1969 – 1992 NM003588.1 |
54°C 593-bp |
HaeIII 1551 |
|
CUL-5 forward: 5'-GAGTGGCTAAGAGAAGTTGGTATG-3' CUL-5 reverse: 5'-TCTTCTCTCATCCTTTCTGTAGTG-3' |
nt 1521 – 1544 nt 2171 – 2194 NM003478.1 |
58°C 674-bp |
EcoRI 1767 |
|
Ribosomal S9 forward: 5'-GCAGGCGCAGACGGGGAAGC-3' Ribosomal S9 reverse: 5'-CGCGAGCGTGGTGGATGGAC-3' |
nt 6 – 25 nt 397 – 416 NM001013.2 |
62°C 411-bp |
DdeI 349 |
Figure 2.

RT-PCR analysis for CUL-1, -2, -3, -4A, -4B, -5, and ribosomal protein S9 in HMEC, MCF-10A, MCF-7, and MDA-MB-231 cells. RT-PCR was performed using 1 μg of total RNA and 25 cycles of PCR. For all of the cells examined, a 560-bp cDNA for CUL-1, a 358-bp cDNA for CUL-2, a 613-bp cDNA for CUL-3, a 512-bp cDNA for CUL-4A, a 593-bp cDNA for CUL-4B, a 674-bp cDNA for CUL-5, and a 411-bp cDNA for ribosomal protein S9 were amplified by RT-PCR. The results shown are representative of duplicate experiments using independent RNA samples.
Figure 3.

Northern blot analysis for CUL-5 in HMEC, MCF-10A, MCF-7, and MDA-MB-231 cells. Approximately 10 μg of total RNA from HMEC, MCF-10A, MCF-7, MDA-MB-231 was electrophoresed and transferred to a Nytran® supercharge membrane. The blots were hybridized with a 32P-labeled 674-bp CUL-5 cDNA or a 905-bp GAPDH cDNA. The results shown are from a 4-day exposure to x-ray film for CUL-5 and a 5-hr exposure to x-ray film for GAPDH. A major CUL-5 transcript of ~7.4 kb was present in all of the cells examined. The results shown are representative of triplicate experiments using independent RNA samples.
Figure 4.

Western blot analysis for CUL-5 in HMEC, MCF-10A, MCF-7, MDA-MB-231, and sol8 mouse myoblast cells. Approximately 50 μg of protein from each cell type, and 25 μg of protein from sol8 mouse myoblasts was fractionated by SDS-PAGE. The primary antibody was affinity purified anti-CUL-5 (378–393) at ~1.14 μg/ml for 1 hr at room temperature. The secondary antibody was an Immunopure® peroxidase goat anti-rabbit IgG (1:40,000, 1 hr at room temperature). The blots were visualized using the Supersignal® west pico chemiluminescent substrate and exposure to x-ray film. Blots were stripped and reprobed with rabbit polyclonal anti-actin (1:1,000, Sigma) to evaluate loading differences and the integrity of the Western blotting procedure. Duplicate experiments yielded similar results.
Expression of CUL-5 in human tissues
A normal human tissue expression array was incubated with a radiolabeled CUL-5 cDNA probe to determine CUL-5 mRNA expression in adult and fetal human tissues. CUL-5 mRNA is expressed in a wide range of human tissues (Table 2). Using a PhosphorImager to quantify the data the highest relative expression of CUL-5 was found in skeletal muscle (volume = 163430, coordinate 7B of Table 2) and the lowest relative expression was found in ovarian tissue (volume = 11139, coordinate 8G of Table 2). CUL-5 mRNA was also detected in fetal human tissue samples and in human cancer cell lines (Table 2). The integrity of the samples on the array was confirmed by reprobing the blot with a radiolabeled ubiquitin cDNA (data not shown). CUL-5 mRNA was not detected in negative controls, including yeast total RNA, yeast tRNA, E. coli rRNA, E. coli DNA, poly r(A) and Cot-1 DNA; these samples were used to obtain an average background correction value. Minimal hybridization did occur with the human genomic DNA samples on the array, which may represent the 674-bp cDNA hybridizing with the CUL-5 gene.
Table 2.
Analysis of CUL-5 expression in normal human tissues and cancer cell lines using a multiple human tissue expression array. The array was hybridized with the 32P-labeled (2 × 107 cpm) 674-bp CUL-5 cDNA for 20 hrs. Analysis of the array was performed using a PhosphorImager and ImageQuant software. The background-adjusted volumes for each sample are reported for the 2-day exposure of the array to a phosphor-screen.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| A | whole brain 77451 | cerebellum left 67297 | substantia nigra 41295 | heart 97252 | esophagus 44513 | colon, transverse 62300 | kidney 137204 | lung 58777 | liver 85484 | leukemia, HL-60 48972 | fetal brain 43774 | yeast total RNA |
| B | cerebral cortex 73105 | cerebellum right 110567 | accumbens nucleus 82337 | aorta 31370 | stomach 63062 | colon, descending 60792 | skeletal muscle 163430 | placenta 80932 | pancreas 79718 | HeLa S3 32213 | fetal heart 55979 | yeast tRNA |
| C | frontal lobe 68310 | corpus callosum 63455 | thalamus 76840 | atrium, left 83350 | duodenum 54191 | rectum 49102 | spleen 53685 | bladder 61953 | adrenal gland 70467 | leukemia, K-562 58024 | fetal kidney 63966 | E. coli rRNA |
| D | parietal lobe 63575 | amygdala 72591 | pituitary gland 65881 | atrium, right 71626 | jejunum 75472 | thymus 50578 | uterus 46700 | thyroid gland 62460 | leukemia, MOLT-4 40827 | fetal liver 47640 | E. coli DNA | |
| E | occipital lobe 78745 | caudate nucleus 84064 | spinal cord 28621 | ventricle, left 76376 | ileum 66144 | peripheral blood leukocyte 29858 | prostate 57159 | salivary gland 73592 | Burkitt's lymphoma, Raji 31147 | fetal spleen 62971 | poly r(A) | |
| F | temporal lobe 55023 | hippocampus 52220 | ventricle, right 62321 | ileocecum 45452 | lymph node 48852 | testis 48162 | mammary gland 28970 | Burkitt's lymphoma, Daudi 30413 | fetal thymus 53776 | human C0t-1 DNA | ||
| G | paracentral gyrus of cerebral cortex 48569 | medulla oblongata 60577 | inter-ventricular septum 89675 | appendix 56027 | bone morrow 37152 | ovary 11139 | colorectal adenocarcinoma, SW480 54931 | fetal lung 64933 | human DNA 100 ng 6117 | |||
| H | pons 55401 | putamen 70937 | apex of the heart 91122 | colon, ascending 40405 | trachea 38285 | lung, carcinoma, A549 27590 | human DNA 500 ng 9541 |
Expression of CUL-5 in matched normal tissues versus tumor tissues
A cancer profiling array was used to examine CUL-5 expression in matched normal tissues versus tumor tissues. With the 50 cases of breast tissue examined, 41 cases (82%) demonstrated a decrease in CUL-5 expression in the breast tumor tissue versus the matched normal tissue (see Table 3, Figures 5, 6 and 7). In three breast tumor cases with matched metastatic samples, CUL-5 expression was similar in the tumor tissues versus the matched metastatic tissues (Figure 7). With the 50 cases of breast tissue there was a statistically significant (P < 0.0001) ~2.2 fold average decrease in CUL-5 expression in the tumor tissue versus the matched normal tissue (Figure 8). The largest difference was seen in sample #31 on the array with a ~7.4 fold decrease in CUL-5 expression in the tumor tissue versus the normal tissue (Table 3 and Figure 7). On the cancer profiling array three other tissues demonstrated statistically significant decreases in CUL-5 expression in tumor tissue versus the matched normal tissue (Figure 8). These tissues included uterine (~1.5 fold decrease, P = 0.0021, n = 42), colon (~1.4 fold decrease, P = 0.0012, n = 35), and kidney (~1.5 fold decrease, P = 0.0034, n = 20). This trend was also observed in ovarian matched normal and tumor tissues, but was not significant (P = 0.0565, n = 14). Conversely, prostate tissue exhibited a ~1.5 fold increase in CUL-5 mRNA in tumor tissue versus normal tissue (P = 0.0335, n = 4). The integrity of the samples on the cancer profiling array was confirmed by reprobing the array with a radiolabeled ubiquitin cDNA (data not shown). As with the normal human tissue expression array, the CUL-5 674-bp cDNA probe did not hybridize with yeast total RNA, yeast tRNA, E. coli rRNA, E. coli DNA, poly r(A) and Cot-1 DNA.
Table 3.
Clinical information and fold-change in CUL-5 expression for the 50 cases of matched normal tissue and breast tumor tissue. The fold-increase (↑) or fold-decrease (↓) in CUL-5 expression in the tumor tissue compared to the matched normal tissue is shown for each individual matched pair.
| Sample Number | Array Location Normal/Tumor | Patient Information | Fold Change in CUL-5 |
| #1 | 1A/2A | 39-yr-old Caucasian, noninfiltrating intraductal carcinoma | ↓1.89 |
| #2 | 1B/2B | 78-yr-old Caucasian, lobular carcinoma | ↓1.79 |
| #3 | 1C/2C | 49-yr-old Caucasian, infiltrating intraductal carcinoma | ↓1.66 |
| #4 | 1D/2D | 52-yr-old Caucasian, infiltrating intraductal carcinoma | ↓1.90 |
| #5 | 1E/2E | 66-yr-old Caucasian, infiltrating intraductal carcinoma | ↑1.01 |
| #6 | 1F/2F | 65-yr-old Caucasian, lobular carcinoma | ↓3.48 |
| #7 | 1G/2G | 44-yr-old Caucasian, infiltrating ductal carcinoma | ↓2.07 |
| #8 | 1H/2H | 33-yr-old Caucasian, infiltrating ductal carcinoma | ↓3.00 |
| #9 | 1I/2I | 40-yr-old Caucasian, infiltrating ductal carcinoma | ↓2.26 |
| #10 | 1J/2J | 49-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.26 |
| #11 | 1K/2K | 41-yr-old Caucasian, lobular carcinoma | ↓2.34 |
| #12 | 1L/2L | 50-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.66 |
| #13 | 1M/2M | 61-yr-old Caucasian, lobular carcinoma | ↓4.87 |
| #14 | 1N/2N | 64-yr-old Caucasian, infiltrating ductal carcinoma | ↓4.85 |
| #15 | 1O/2O | 40-yr-old Caucasian, infiltrating ductal carcinoma | ↓4.66 |
| #16 | 1P/2P | 50-yr old Caucasian, infiltrating ductal carcinoma | ↓2.10 |
| #17 | 1Q/2Q | 44-yr-old Caucasian, infiltrating ductal carcinoma | ↑1.34 |
| #18 | 1R/2R | 52-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.41 |
| #19 | 1S/2S | 47-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.95 |
| #20 | 1T/2T | 59-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.67 |
| #21 | 1U/2U | 50-yr-old Caucasian, infiltrating ductal carcinoma | ↑1.24 |
| #22 | 1V/2V | 68-yr-old Caucasian, infiltrating ductal carcinoma | ↓2.45 |
| #23 | 1W/2W | 63-yr-old Caucasian, tubular carcinoma | ↓1.51 |
| #24 | 1X/2X | 44-yr-old Caucasian, fibrosarcoma | ↓2.52 |
| #25 | 1Y/2Y | 49-yr-old Caucasian, infiltrating lobular carcinoma | ↓1.84 |
| #26 | 1Z/2Z | 64-yr-old Caucasian, infiltrating ductal carcinoma | ↓4.08 |
| #27 | 1AA/2AA | 50-yr-old Caucasian, infiltrating ductal carcinoma | ↓2.63 |
| #28 | 1BB/2BB | 39-yr-old Caucasian, mixed lobular-ductal carcinoma | ↓3.15 |
| #29 | 1CC/2CC | 66-yr-old Caucasian, infiltrating lobular carcinoma | ↑1.07 |
| #30 | 1DD/2DD | 65-yr-old Caucasian, infiltrating lobular carcinoma | ↓4.68 |
| #31 | 1EE/2EE | 61-yr-old Caucasian, infiltrating lobular carcinoma | ↓7.41 |
| #32 | 1FF/2FF | 38-yr-old Caucasian, infiltrating lobular carcinoma | ↓5.19 |
| #33 | 3A/4A | 58-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.88 |
| #34 | 3B/4B | 64-yr-old Caucasian, infiltrating ductal carcinoma | ↓2.72 |
| #35 | 3C/4C | 47-yr-old Caucasian, tubular adenocarcinoma | ↓2.61 |
| #36 | 3D/4D | 40-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.87 |
| #37 | 3E/4E | 60-yr-old Caucasian, lobular carcinoma | ↓6.37 |
| #38 | 3F/4F | 62-yr-old Caucasian, infiltrating ductal carcinoma | ↓5.01 |
| #39 | 3G/4H | 71-yr-old Caucasian, infiltrating ductal carcinoma | ↓4.51 |
| #40 | 3I/4J | 52-yr-old Caucasian, infiltrating ductal carcinoma | ↓4.16 |
| #41 | 3K/4L | 57-yr-old Caucasian, infiltrating ductal carcinoma | ↓4.42 |
| #42 | 3M/4M | 43-yr-old Caucasian, infiltrating ductal carcinoma | ↓5.08 |
| #43 | 3N/4N | 40-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.30 |
| #44 | 3O/4O | 48-yr-old Caucasian, infiltrating ductal carcinoma | ↓2.65 |
| #45 | 3P/4P | 45-yr-old Caucasian, infiltrating ductal carcinoma | ↑2.28 |
| #46 | 3Q/4Q | 47-yr-old Caucasian, medullary carcinoma | ↑1.52 |
| #47 | 3R/4R | 60-yr-old Caucasian, infiltrating ductal carcinoma | ↑2.69 |
| #48 | 3S/4S | 71-yr-old Caucasian, infiltrating ductal carcinoma | ↑1.06 |
| #49 | 3T/4T | 53-yr-old Caucasian, mucinous adenocarcinoma | ↑1.41 |
| #50 | 3U/4U | 42-yr-old Caucasian, infiltrating ductal carcinoma | ↓1.44 |
Figure 5.

Location of samples on the cancer profiling array. Information concerning the patient samples on this array is available at http://bioinfo.clontech.com/dparray (N = normal tissue; T = tumor tissue; a box indicates a matched normal/tumor pair with an associated metastatic sample).
Figure 6.
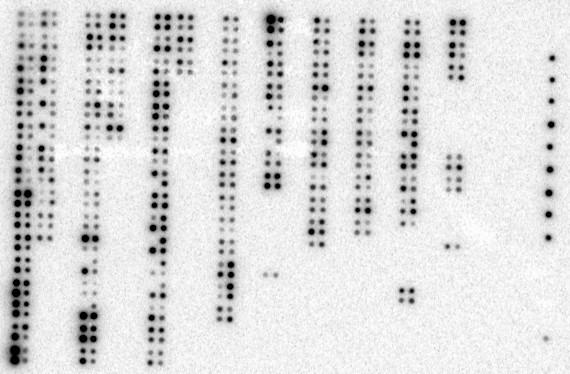
Expression of CUL-5 in normal tissues versus matched tumor tissues. The cancer profiling array was hybribized with the 32P-labeled (2 × 107 cpm) 674-bp CUL-5 cDNA for 20 hrs. The results shown represent the 2-day exposure to the phosphor screen. The location of individual samples on the array is shown in Figure 5.
Figure 7.
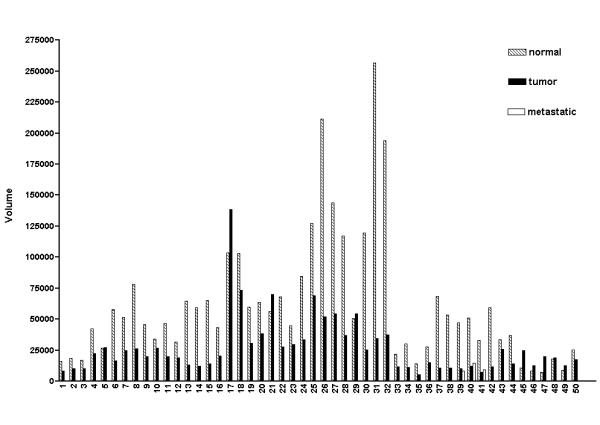
Expression of CUL-5 in the 50 cases of matched normal breast tissue and breast tumor tissue. The data in this figure was derived from quantification of the data in Figure 6 using a PhosphorImager and ImageQuant software, and the background adjusted volumes are shown for each sample. 41 of the 50 cases of breast tissue (82%) exhibited a decrease in CUL-5 expression in the tumor tissue versus the matched normal tissue. Three of the 50 cases had a matched metastatic sample (cases 39, 40, and 41), and the level of CUL-5 expression in the metastatic samples was similar to that seen in the associated tumor tissues.
Figure 8.
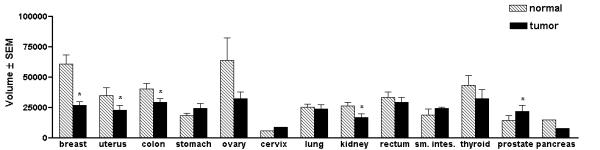
Expression of CUL-5 in matched normal tissue versus tumor tissue. The data in this figure was derived by quantification of the data in Figure 6 using a PhosphorImager and ImageQuant software. The mean volumes ± the SEM are shown. An asterisk indicates a statistically significant difference between the matched normal versus tumor tissue as determined by two-way paired t-tests (P ≤ 0.05). A statistically significant decrease in CUL-5 expression was found in breast tumor tissue (~2.2 fold decrease, P < 0.0001, n = 50), uterine tumor tissue (~1.5 fold decrease, P = 0.0021, n = 42), colon tumor tissue (~1.4 fold decrease, P = 0.0012, n = 35), and kidney tumor tissue (~1.5 fold decrease, P = 0.0034, n = 20), compared to matched normal tissues. A significant increase in CUL-5 expression was found in prostate tumor tissue compared to matched normal tissue (~1.5 fold increase, P = 0.0335, n = 4).
Discussion
The CUL-5 gene is of interest in relation to breast tumorigenesis as a putative tumor suppressor, since it is located on a region of chromosome 11 that is associated with LOH in breast cancer [1-8]. However, there is a lack of information concerning the expression of CUL-5 in breast epithelial cells compared with breast cancer cells and in normal tissues compared with tumor tissues. To address this issue, we used RT-PCR, Northern blot analysis and Western blot analysis to analyze CUL-5 mRNA and protein expression. With these techniques, we demonstrated a major CUL-5 transcript of ~7.4 kb and a major immunoreactive protein of Mr ~ 82 kDa in primary HMECs, immortalized but non-tumorigenic MCF-10A breast epithelial cells, ER-α positive MCF-7 breast cancer cells, and ER-α negative MDA-MB-231 breast cancer cells, which is consistent with previous reports [7,17-19]. While CUL-5 expression was unchanged in breast cancer cells versus breast epithelial cells, the cells used for these studies came from unsynchronized subconfluent proliferating cultures, and cell cycle dependent expression of some CUL family members has been described [19,20]. Investigation of cell cycle dependent expression of CUL-5 is crucial to gain insight into both the mechanism(s) regulating the expression and potential cellular function(s) of CUL-5.
The multiple human tissue expression array demonstrated CUL-5 mRNA expression in many fetal and adult tissues. Ubiquitous expression of CULs is expected, given that CUL-1, -3 and -4A are all critical for proper development; mice nullizygous for these CUL proteins fail to develop past E5.5, E7.5, and E7.5, respectively [21-23]. The finding of CUL-5 expression in various fetal human tissues suggests that CUL-5 may also play a role in embryogenesis and development. CUL-5 was expressed in adult tissues associated with proliferation (e.g. gastrointestinal tract), and in adult tissues not associated with a high level of proliferation (e.g. brain). Ubiquitous tissue expression of CUL-5 mRNA is supported by RT-PCR and Northern blot studies in various human, rat and rabbit tissues [7,17,18,24]. The high level of CUL-5 mRNA expression found in skeletal muscle is consistent with similar findings by others [7,18], and suggests that CUL-5 may play an important role in this tissue. CUL-5 protein expression was originally reported to be restricted to medullary collecting tubule cells and endothelial cells as determined by immunohistochemistry [17]. However, our work indicates that several cultured cell types, including breast epithelial cells, breast cancer cell lines of epithelial origin, and mouse sol8 myoblast cells all express CUL-5 protein. CUL-5 has also recently been show to be ubiquitously expressed in the rat brain by immunocytochemistry [25]. The broad nature of CUL-5 expression suggests that CUL-5 may have a fundamental role in cellular differentiation and proliferation.
We evaluated 50 cases of breast tumor tissue and matched normal tissue, and found a significant ~2.2 fold decreased expression of CUL-5 in tumor tissues compared to normal tissues. Patient samples on this cancer profiling array support the hypothesis that decreased expression of CUL-5 may be associated with breast tumorigenesis. This decrease is remarkable, since a 2 fold reduction in CUL-4A expression can result in embryonic death in mice [23], and less than 2 fold increased expression of CUL-4A prevents granulocytic differentiation and promotes proliferation [26]. The cause of the decreased CUL-5 expression in breast tumor tissue remains unknown, but may reflect both genetic and epigenetic events, and warrants further investigation.
Another potential tumor suppressor gene in breast cancer that is also located on chromosome 11 q22-23 is the ATM gene, which is normally important for DNA damage cell cycle checkpoint control [27]. Similar to our results for CUL-5 expression in MCF-7 and MDA-MB-231 cells, these cells do not express decreased levels of ATM mRNA [28]. In contrast to our results obtained for CUL-5 expression in breast tissue, one study reported that of 11 cases of matched normal breast and cancerous tissue, seven cases had higher expression of ATM mRNA associated with the cancer, three cases expressed similar levels in the normal tissue versus the cancer, and only one case had decreased expression in the cancer [28]. The role of the ATM gene in breast cancer is controversial since mutations in the ATM gene have not been clearly linked to development of breast cancer [29,30].
In addition to differential CUL-5 expression in breast tumor tissue, a statistically significant decrease in CUL-5 expression was found in uterine tumor tissue, colon tumor tissue, and kidney tumor tissue compared to matched normal tissues. A similar trend was seen with ovarian tumor tissue compared to matched normal tissue, although the difference was not statistically significant. An increase in CUL-5 expression was seen in prostate tumor tissue compared to matched normal tissue. Differential expression of CUL-5 in other human tumors suggests that dysregulation of CUL-5 expression may occur during tumorigenesis in these tissues as well. A recent study demonstrated a ~1.5 fold increase in CUL-5 expression in lung adenocarcinoma versus unmatched control lung tissue [31]. Of the 21 matched lung samples on the cancer profiling array used for this study there were seven matched cases with the diagnosis of adenocarcinoma (Figure 5, 27B/28B, 27I/28I, 27J/28J, 27L/28L, 27N/28N, 27O/28O, 27P/28P). The mean volume ± SEM for CUL-5 expression for the normal tissue and matched tumor tissue for these seven cases is 19230 ± 3352 and 22520 ± 6115, respectively. Therefore, The data for lung adenocarcinoma demonstrate a similar trend for higher expression of CUL-5 in the tumor tissue versus normal tissue, although the ~1.2 fold difference was not statistically significant (P = 0.4574, n = 7).
The functional significance of altered CUL-5 expression in cancer is currently not known. CUL-5 was originally described as a gene product involved in vasopressin-mediated increases of intracellular-free calcium [9], and was subsequently shown to attenuate cAMP production in cultured cells [32]. Recently, it was reported that overexpression of CUL-5 in CHO and cos 1 cells attenuates proliferation with an associated decrease in MAPK phosphorylation and an increase in p53 [33]. It is not clear if these reported effects of CUL-5 on cellular signaling and proliferation are E3 ubiquitin ligase dependent or independent. CUL-5 belongs to the CUL protein family, and CULs have diverse roles in development, cell cycle regulation, and transcriptional regulation through their actions as components of E3 ubiquitin ligase complexes [15,16,34]. Loss of function mutations in CUL-1 cause generalized tissue hyperplasia in C. elegans [10]. CUL-1 is a component of an E3 ubiquitin ligase complex called the SCF [35], in which S stands for the adaptor protein SKP-1, C stands for the yeast CUL CDC53, and F stands for a substrate recognition F-box protein. CUL-1 containing E3 ubiquitin ligase complexes are implicated in ubiquitinating a wide range of target proteins such as cyclins E and D, E2f-1, p21, p27, β-catenin and IκB [21,36-49]. CUL-2, -3, -4A and -5 do not associate with the adaptor protein SKP-1, but all of these CULs appear to function as components of E3 ubiquitin ligases, suggesting that they use other adaptor protein(s) besides SKP-1 [50-53]. In support of this idea, CUL-2 forms an E3 ubiquitin ligase complex with the Ring finger protein ROC1/Rbx1, the adaptor Elongin BC, and a BC-box motif containing substrate recognition protein such as the VHL tumor suppressor or mMED8 [34,54-56]. The VHL/Elongin BC/CUL-2/ROC1 E3 ubiquitin ligase complex regulates transcriptional activity of hypoxia-inducible genes by targeting the α subunit of hypoxia-inducible transcription factors for ubiquitin-mediated degradation during non-hypoxic conditions [57-63]. In C. elegans, CUL-2 is critical during G1-S phase transitions, since CUL-2 mutant germ cells exhibit G1-phase arrest with accumulation of CKI-1, a cyclin dependent kinase inhibitor [64]. The CUL-2 mutant C. elegan embryos also exhibited mitotic chromosomes that were unable to condense [64]. Expression of CUL-3 is increased in cultured colon cancer cells versus normal colon cells, and CUL-3 containing E3 ubiquitin ligase complexes are implicated in ubiquitinating cyclins D and E [22,49,65]. The CUL-4A gene is located on an amplicon in breast cancer and was found to be amplified in 16% of the cases of primary breast cancer examined and the mRNA was overexpressed in 47% of breast cancers examined [66]. Research indicates that CUL-4A is involved in cell cycle control, degradation of the DNA repair protein DDB2, embryonic development and hematopoiesis [23,26,67-70]. The recently described CUL-7 which has been shown to play a role in vascular morphogenesis, forms an SCF E3 ubiquitin ligase by interacting with the adaptor SKP-1, the RING finger protein ROC1, and the F-box protein fbx29 [12,13]. Recent studies demonstrate that CUL-5 is capable of forming E3 ubiquitin ligase complexes with elongin BC, as is the case with CUL-2 [51]. The CUL-5/Elongin BC complex was shown to interact with a number of BC-box motif containing proteins, such as MUF1, VHL, RNA polymerase II elongation factor Elongin A, and suppressor of cytokine signaling proteins (e.g. SOCS1 and WSB-1) [51]. It is of interest that CUL-5 was recently shown to be involved in MDM2-independent and adenovirus-dependent ubiquitin-mediated degradation of p53 [71,72].
Despite growing evidence of functional roles of CUL family members, CUL-5 remains largely uncharacterized, including its possible involvement in breast tumorigenesis. Future plans include examination of CUL-5 protein expression in breast cancer biopsy specimens by immunohistochemistry. Also, to elucidate the cellular role(s) of CUL-5 in breast tumorigenesis, experiments are planned to modulate CUL-5 expression and to identify CUL-5 interacting proteins from breast epithelial cells and breast cancer cells.
Conclusions
Breast epithelial cells (HMEC, MCF-10A) and breast cancer cells (MCF-7, MDA-MB-231) all express CUL-5 mRNA and protein without an apparent decrease in CUL-5 expression in the breast cancer cells versus the breast epithelial cells. CUL-5 mRNA is ubiquitously expressed in human tissues, which is indicative of an important fundamental cellular role for CUL-5. Examination of 50 individual cases of matched breast tumor tissue and normal tissue revealed a 2.2 fold decrease in the expression of CUL-5 in the tumor tissue versus the matched normal tissue, thus supporting a potential role for dysregulation of CUL-5 expression in breast tumorigenesis.
Methods
Cell culture
Primary human mammary epithelial cells (HMEC) were obtained from Clonetics (Walkersville, MD) and maintained using medium and instructions provided by the manufacturer. The MCF-10A breast epithelial cells, which are immortalized, but nontumorigenic [73,74], were obtained from the ATCC (Manassas, VA; CRL-10317) and maintained in Ham's F12:Dulbecco's modified Eagle's medium (1:1, Sigma, St. Louis, MO) supplemented with 20 ng/ml epidermal growth factor (Gibco Invitrogen Corp., Carlsbad, CA), 100 ng/ml cholera toxin (Sigma), 0.01 mg/ml insulin (Sigma) 500 ng/ml hydrocortisone (Sigma) and 5% horse serum (Gibco Invitrogen Corp.). The ER-α positive MCF-7 breast cancer cell line [75] and the ER-α negative MDA-MB-231 breast cancer cell line [76] were obtained from the ATCC (HTB-22 and HTB-26, respectively) and maintained in Dulbecco's modified Eagle's medium containing 10% iron-supplemented calf serum (Hyclone, Logan, UT). All cells were maintained at 37°C in a humidified environment with 5% CO2/95% air.
RT-PCR analysis
Total RNA was isolated from subconfluent proliferating cultures using TRI REAGENT® according to the instructions provided by the manufacturer (MRC, Cincinnati, OH). RT-PCR was performed using ~1 μg of total RNA for each sample and the One-Step Superscript™ kit from Invitrogen. The reaction volumes were 50 μl and contained SUPERSCRIPT™ II reverse transcriptase/Platinum® Taq DNA polymerase (1 μl), dNTPs (0.2 mM of each), magnesium sulfate (1.2 mM) and gene specific sense/antisense primers (0.2 μM) at concentrations that were recommended by the manufacturer. The primers used for RT-PCR of CUL-5 and other CUL family members are shown in Table 1, and were designed using LASERGENE software (DNASTAR, Inc., Madison, WI). The RT-PCR conditions consisted of 30 min at 50°C, 2 min at 94°C, followed by a denaturation step of 30 sec at 94°C, an annealing step of 1 min (see Table 1 for annealing temperatures), an extension step of 1 min at 72°C, and a final extension step of 10 min at 72°C. Semiquantitative RT-PCR was performed for CUL-5, using 20, 25, and 29 cycles, utilizing a technique similar to that previously described [77]. The RT-PCR analyses for CUL-5 expression were performed in triplicate with independent RNA samples. The CUL-5 RT-PCR products from HMEC, MCF-10A, MCF-7, and MDA-MB-231 were cloned using the TOPO TA Cloning® kit (Invitrogen) and confirmed by DNA sequencing using an ABI Model 3700 sequencer. The CUL-5 PCR products from HMEC, MCF-10A, MCF-7, and MDA-MB-231 were also confirmed by restriction endonuclease digestion with EcoRI (see Table I). The CUL-1, -2, -3, -4A and -4B PCR analyses were performed in duplicate using independent RNA samples and 25 cycles of PCR. The CUL-1, -2, -3, -4A, and -4B PCR products were validated by restriction endonuclease digestion with BglII, AatII, EcoRI, Sau3AI, and HaeIII, respectively (see Table I). The NT negative control samples received molecular biology grade water instead of RNA, and the minus reverse transcriptase samples received 2 units of Platinum® Taq DNA polymerase without the SUPERSCRIPT™ II reverse transcriptase. Integrity of the RNA samples was confirmed by amplifying a 411-bp PCR product for ribosomal protein S9.
Northern blot analysis
Approximately 10 μg of total RNA from HMEC, MCF-10A, MCF-7, MDA-MB-231 cells, or 5 μg of RNA ladder (0.24–9.5 Kb, Invitrogen) was electrophoresed for 3.5 – 4 hrs at 70 volts using a 1% agarose gel containing 0.66 M formaldehyde and 1X MOPS. After electrophoresis, gels were washed 3 × 5 min with DEPC-treated water and the RNA was transferred overnight by capillary transfer to a Nytran® supercharge membrane (Schleicher & Schuell, Keene, NH) using 20X SSC (Ambion, Austin, TX) and the Turboblotter™ rapid downward transfer system (Schleicher & Schuell). Following the transfer, RNA was immobilized to the membrane by UV irradiation using a model 1800 UV-Stratalinker (Statagene, La Jolla, CA) and by baking for 2 hrs at 70°C. Membranes were stained with blot stain blue (Sigma), according to the procedure provided by the manufacturer, to visualize the RNA ladder. Membranes were then prehybridized with ULTRAhyb™ hybridization buffer (Ambion) for 0.5 – 1 hr at 42°C. The CUL-5 cDNA probe used for Northern blot was prepared by RT-PCR using RNA from HMEC and the primer pairs described in Table 1. The CUL-5 PCR product was purified using the QIAquick® PCR purification kit (QIAGEN, Valencia, CA) and the DNA concentration determined by measuring the absorbance at 260 nm. The purified DNA (~25 ng) was radiolabeled using deoxycitidine 5'-triphosphate [α-32P] (3000 Ci/mmol, Perkin Elmer Life Sciences, Boston, MA) and the DECAprime™ II DNA labeling kit (Ambion), according to the instructions provided by the manufacturer. The radiolabeled probe was separated from the unincorporated nucleotides using NucTrap® probe purification columns (Stratagene). The radiolabeled DNA was denatured prior to hybridization by adding 0.1 volumes of 1 N NaOH and incubating for 10 min at 37°C. After prehybidization, the membranes were hybridized for 18 hrs at 42°C with ~107cpm of denatured radiolabeled probe in ULTRAhyb (Ambion). Following hybridization, the membranes were washed 2 × 5 min with 2X SSC/0.1% SDS at 42°C, followed by 2 × 15 min washes with 0.1X SSC/0.1% SDS at 42°C. Membranes were then exposed to a Molecular Dynamics phosphor screen (Amersham Pharmacia Biotech, Piscataway, NJ) or to x-ray film for documentation. To account for loading and transfer differences, the membranes were stripped by washing 2 × 15 min at 99°C with 0.1X SSC/0.5% SDS, and reprobed using a radiolabeled 905-bp mouse GAPDH cDNA (Ambion). Northern blots were performed in triplicate using independent RNA samples.
Western blot analysis
Protein samples were prepared from approximately 107 subconfluent proliferating HMEC, MCF-10A, MCF-7, and MDA-MB-231 cells using M-Per™ mammalian protein extraction reagent (Pierce, Rockford, IL) containing a complete™ mini protease inhibitor cocktail tablet (Roche Molecular Biochemicals, Indianapolis, IN), according to the instructions provided by the manufacturers. Protein concentrations were determined using the BCA™ protein assay kit (Pierce) and ~50 μg of each sample was diluted using 2X SDS sample buffer (0.125 M Tris-Cl/0.1% SDS, pH 6.8, 20% glycerol, 4% SDS, 0.2% 2-mercaptoehtanol, 0.001% bromphenol blue), and the samples were boiled for 5 min. Samples were electrophoresed using SDS-PAGE (10% acrylamide separating gel, 3.9% acrylamide stacking gel) according to the method of Laemmli [78]. Proteins were transferred at 4°C to a PVDF-Plus membrane (MSI, Westboro, MA) at 14 volts for 18 hrs. A synthetic peptide consisting of amino acids 378–393 (NDATIFKLELPLKQKG) of human CUL-5, containing a COOH-terminal cysteine, made as an amide, was produced and conjugated to ovalbumin (BioSource International, Hopkinton, MA). Rabbit polyclonal antiserum was generated against this synthetic peptide and was immunopurified using a thiol coupling gel. The blots were blocked for 1 hr at room temperature using Superblock® (Pierce, Rockford, IL), and incubated with affinity-purified anti-CUL-5 (378–393) for 1 hr at room temperature at a dilution of 1:1,000 (~1.14 μg/ml) in antibody dilution buffer (10% Superblock®/90% 10 mM Tris, pH 7.5, 100 mM NaCl, and 0.1% Tween® 20). The blots were then washed 6 × 5 min with wash buffer (10 mM Tris pH 7.5, 100 mM NaCl, and 0.1% Tween® 20). After washing, the blots received a secondary antibody for 1 hr at room temperature, that was either Immunopure® peroxidase conjugated goat anti-rabbit IgG (1:40,000, Pierce) or a Cruz marker compatible peroxidase conjugated goat anti-rabbit IgG (1:5,000, Santa Cruz Biotechnology, Inc., Santa Cruz, CA) prepared in antibody dilution buffer. The blots were then washed as previously described and exposed to Supersignal® west pico chemiluminescent substrate (Pierce). The Western blots were visualized by exposure to x-ray film. As a negative control, IgG was purified from preimmune serum using a NAb™ protein A spin chromatography kit (Pierce), and ~1.14 μg/ml of the IgG purified preimmune serum was used as the primary antibody. An additional control for the Western blots involved adding a 200 molar excess of the immunizing peptide to the anti-CUL-5 (378–393) antiserum for 30 min at room temperature prior to blotting. A cellular lysate from sol8 mouse myoblast cells (Santa Cruz Biotechnology, Inc.) was used as a positive control in the Western blots, since skeletal muscle was found to express the highest levels of CUL-5 mRNA (see Table 2). To control for sample loading differences and the integrity of the Western blotting procedure, the blots were stripped using 0.2 N NaOH and reprobed using a rabbit polyclonal antibody (1:1,000, Sigma) that recognizes actin (~42 kDa).
Human expression arrays
The human multiple tissue expression array was obtained from Clontech (Palo Alto, CA). This array contains poly A+ RNA from 61 adult tissues, 7 fetal tissues, and 8 cancer cell lines. The samples on the multiple tissue expression array are normalized using 8 different housekeeping genes [79-81]. The multiple tissue expression array was prehybridized for 30 min at 65°C in 10 ml of ExpressHyb (Clontech) containing 1 mg of sheared salmon testes DNA. The radiolabeled CUL-5 probe was prepared as described for Northern blot analysis, and 2 × 107 cpm of the probe was mixed with 30 μg of C0t-1 DNA, 150 μg of sheared salmon testes DNA, and 50 μl of 20X SSC, and the mixture was heated at 95°C for 5 min followed by 68°C for 30 min. The array was then hybridized with radiolabeled probe for 20 hrs at 65°C in 5 ml of ExpressHyb. The array was washed 5 × 20 min at 65°C with 2X SSC/0.1% SDS, and 2 × 20 min at 55°C with 0.1X SSC/0.5 % SDS. The array was exposed to a phosphor screen for 1, 2, and 3 days and to x-ray film for 3 and 4 days. The array was stripped using boiling 0.5% SDS and reprobed for 20 hrs using a radiolabeled human ubiquitin cDNA (1 × 107 cpm), to ensure the integrity of the samples on the array. The cancer profiling array contains SMART™ cDNA synthesized from CLONTECH's premium RNA™ from 241 cases of matched normal and tumor tissue from individual patients [82]. Information concerning the patient samples on this array can be obtained at http://bioinfo.clontech.com/dparray[83]. Information concerning the 50 cases of matched breast tissue is located in Table 3, and the location of samples on the array is shown in Figure 5. The matched samples on the cancer profiling array are normalized using four different housekeeping genes. The cancer profiling array was analyzed using a technique similar to that described for the multiple tissue expression array, except that the array was washed 4 × 30 min at 65°C with 2X SSC/0.5% SDS, followed by 1 × 30 min at 65°C with 0.2X SSC/0.5% SDS, with a final 5 min wash using 2X SSC. The cancer-profiling array was exposed to a phosphor-screen for 1, 2, and 3 days and to x-ray film for 3 and 4 days. The cancer profiling array was stripped and reprobed using radiolabeled human ubiquitin cDNA to ensure the integrity of the samples on the array. Both arrays were analyzed using a Molecular Dynamics STORM 860 PhosphorImager and ImageQuant software. Analysis of the arrays consisted of drawing individual boxes or grids around the hybridization signal for each sample and subtracting an average background value adjusted to the area of the box or grid for each individual sample. The 2-day exposure for both arrays was used for quantification of CUL-5 expression with the PhosphorImager, since the sample signals were in the linear range. Statistical analysis of the cancer-profiling array was performed using GraphPad prism 2.01 software (GraphPad Software, inc., San Diego, CA) and two-tailed paired t-tests (P ≤ 0.05).
List of abbreviations
ATCC, American Type Culture Collection; ATM, ataxia telangiectasia mutated; bp, base pairs; cpm, counts per minute; CUL, Cullins; DEPC, Diethyl pyrocarbonate; dNTPs, 2'-deoxynucleoside 5'-triphosphates; ER, estrogen receptor; GAPDH, glyceraldehyde 3-phospahte dehydrogenasae; hr, hour; HMEC, human mammary epithelial cells; LOH, loss of heterozygosity; min, minutes; MOPS, 3-(N-morpholino) propanesulfonic acid; nt, nucleotide; NT, no template; RT-PCR, reverse transcription polymerase chain reaction; ROC, regulator of Cullins; sec, seconds; SSC, saline sodium citrate; SEM, standard error of the mean; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; VHL, von Hippel-Lindau
Authors' contributions
MJF was responsible for designing and implementing all experiments and manuscript preparation. KAL assisted with DNA sequencing and manuscript editing. GAK was responsible for maintaining cell cultures and assisted with RT-PCR, Northern blot analysis, Western blot analysis and the expression arrays. DMS assisted with RT-PCR, restriction endonuclease confirmation of PCR products, and the expression arrays. CJM assisted with the expression arrays. AH, JRL, KP and AH assisted with RT-PCR, Northern blot analysis and restriction endonuclease confirmation of PCR products. All authors read and approved the final manuscript.
Acknowledgments
Acknowledgments
This research was supported in part by grant number R15 CA85279 from the NIH, awarded to Michael J. Fay. Amyna Husain and David Shoipe received summer research stipends from the Office of Research Affairs and Sponsored Programs at Midwestern University. Kenneth Pherson received a summer research stipend from the Greater Chicagoland Arthritis Foundation. The authors are thankful to Victoria Sears and Dana Fay for assistance with the preparation of figures and to Drs. Walter Prozialeck and Nalini Chandar for reviewing the manuscript prior to submission.
Contributor Information
Michael J Fay, Email: mfayxx@midwestern.edu.
Kenneth A Longo, Email: kalongo@umich.edu.
George A Karathanasis, Email: George.Karathanasis@mwumail.midwestern.edu.
David M Shope, Email: David.Shope@mwumail.midwestern.edu.
Craig J Mandernach, Email: CMandernach@mwumail.midwestern.edu.
Jason R Leong, Email: Jleong@mwumail.midwestern.edu.
Alfred Hicks, Email: Alfred.Hicks@mwumail.midwestern.edu.
Kenneth Pherson, Email: Kenneth.Pherson@mwumail.edu.
Amyna Husain, Email: AHusain@mwumail.midwestern.edu.
References
- Carter SL, Negrini M, Baffa R, Gillum DR, Rosenberg AL, Schwartz GF, Croce CM. Loss of heterozygosity at 11q22-23 in breast cancer. Cancer Res. 1994;54:6270–6274. [PubMed] [Google Scholar]
- Hamptom GM, Mannermaa A, Winquist R, Alavaikko M, Blanco G, Taskinen PJ, Kiviniemi H, Newsham I, Cavanee WK, Evans GA. Loss of heterozygosity in sporadic human breast carcinoma: A common region between 11q22 and 11q23. Cancer Res. 1994;54:4586–4589. [PubMed] [Google Scholar]
- Gudmundsson J, Barkardottir RB, Eiriksdottir G, Baldusson T, Arason A, Egilsson V, Ingvarsson S. Loss of heterozygosity at chromosome 11 in breast cancer: Association of prognostic factors with genetic alterations. Br J Cancer. 1995;72:696–701. doi: 10.1038/bjc.1995.396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Negrini M, Rasio D, Hampton GM, Sabbioni S, Rattan S, Carter SL, Rosenberg AL, Schwartz GF, Shiloh Y, Cavenee WB, Croce CM. Definition and refinement of chromosome 11 regions of loss of heterozygosity in breast cancer: Identification of a new region at 11q23-24. Cancer Res. 1995;55:3003–3007. [PubMed] [Google Scholar]
- Tomlinson IPM, Stickland JE, Lee ASG, Bromley L, Evans MF, Morton J, McGee JOD. Loss of heterozygosity on chromosome 11q in breast cancer. J Clin Pathol. 1995;48:424–428. doi: 10.1136/jcp.48.5.424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Winqvist R, Hampton GM, Mannermaa A, Blanco G, Alavaikko M, Kiviniemi H, Taskinen PJ, Evans GA, Wright FA, Newsham I, Cavenee WK. Loss of heterozygosity for chromosome 11 in primary human breast tumors is associated with poor survival after metastasis. Cancer Res. 1995;55:2660–2664. [PubMed] [Google Scholar]
- Byrd PJ, Stankovic T, McConville CM, Smith AD, Cooper PR, Taylor AMR. Identification and analysis of expression of human VACM-1, a cullin gene family member located on chromosome 11q22-23. Genome Res. 1997;7:71–75. doi: 10.1101/gr.7.1.71. [DOI] [PubMed] [Google Scholar]
- Driouch K, Briffod M, Bieche I, Champeme M-H, Lidereau R. Location of several putative genes possibly involved in human breast cancer progression. Cancer Res. 1998;58:2081–2086. [PubMed] [Google Scholar]
- Burnatowska-Hledin MA, Spielman WS, Smith WL, Shi P, Meyer JM, Dewitt DL. Expression cloning of an AVP-activated, calcium-mobilizing receptor from rabbit kidney medulla. Am J Physiol. 1995;268:F1198–F1210. doi: 10.1152/ajprenal.1995.268.6.F1198. [DOI] [PubMed] [Google Scholar]
- Kipreos ET, Lander LE, Wing JP, He W, Hedgecock EM. cul-1 is required for cell cycle exit in C. elegans and identifies a novel gene family. Cell. 1996;85:829–839. doi: 10.1016/s0092-8674(00)81267-2. [DOI] [PubMed] [Google Scholar]
- Mathias N, Johnson SL, Winey M, Adams AE, Goetsch L, Pringle JR, Byers B, Goebl MG. Cdc53p acts in concert Cdc4p and Cdc34p to control the G1-to-S-phase transition and identifies a conserved family of proteins. Mol Cell Biol. 1996;16:6634–6643. doi: 10.1128/mcb.16.12.6634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dias DC, Dolios G, Wang R, Pan Z-Q. CUL7: A DOC domain-containing cullin selectively binds Skp1Fbx29 to form an SCF-like complex. Proc Natl Acad Sci USA. 2002;26:16601–16606. doi: 10.1073/pnas.252646399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arai T, Kasper JS, Skaar JR, Ali SH, Takahashi C, DeCaprio JA. Targeted disruption of p185/Cul7 gene results in abnormal vascular morphogenesis. Proc Natl Acad Sci USA. 2003;100:9855–9860. doi: 10.1073/pnas.1733908100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patton EE, Willems AR, Sa D, Kuras L, Thomas D, Craig KL, Tyers M. Cdc53 is a scaffold for multiple Cdc34/Skp1/F-box protein complexes that regulate cell division and methionine biosynthesis in yeast. Genes Dev. 1998;12:692–705. doi: 10.1101/gad.12.5.692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hershko A, Ciechanover A. The Ubiquitin System. Annu Rev Biochem. 1998;67:425–479. doi: 10.1146/annurev.biochem.67.1.425. [DOI] [PubMed] [Google Scholar]
- Deshaies RJ. SCF and CULLIN/RING H2-based ubiquitin ligases. Annu Rev Cell Dev Biol. 1999;15:435–467. doi: 10.1146/annurev.cellbio.15.1.435. [DOI] [PubMed] [Google Scholar]
- Burnatowska-Hledin M, Lazdins IB, Listenberger L, Zhao P, Sharangpani A, Folta V, Card B. VACM-1 receptor is specifically expressed in rabbit vascular endothelium and renal collecting tubule. Am J Physiol. 1999;276:F199–F209. doi: 10.1152/ajprenal.1999.276.2.F199. [DOI] [PubMed] [Google Scholar]
- Hori T, Osaka F, Chiba T, Miyamoto C, Okabayashi K, Shimbara N, Kato S, Tanaka K. Covalent modification of all members of human cullin family proteins by NEDD8. Oncogene. 1999;18:6829–6834. doi: 10.1038/sj.onc.1203093. [DOI] [PubMed] [Google Scholar]
- Burnatowska-Hledin M, Zeneberg A, Roulo A, Grobe J, Zhao P, Lelkes PI, Clare P, Barney C. Expression of VACM-1 protein in cultured rat adrenal endothelial cells is linked to the cell cycle. Endothelium. 2001;8:49–63. doi: 10.3109/10623320109063157. [DOI] [PubMed] [Google Scholar]
- Cho RJ, Huang M, Campbell MJ, Dong H, Steinmetz L, Sapinso L, Hampton G, Elledge SJ, Davis RW, Lockhart DJ. Transcriptional regulation and function during the human cell cycle. Nat Genet. 2001;27:48–54. doi: 10.1038/83751. [DOI] [PubMed] [Google Scholar]
- Dealy J, Nguyen KVT, Lo J, Gstaiger M, Krek W, Elson D, Arbeit J, Kipreos ET, Johnson RS. Loss of Cul1 results in early embryonic lethality and dysregulation of cyclin E. Nat Genet. 1999;23:245–248. doi: 10.1038/13886. [DOI] [PubMed] [Google Scholar]
- Singer JD, Gurian-West M, Clurman B, Roberts JM. Cullin-3 targets cyclin E for ubiquitination and controls S phase in mammalian cells. Genes Dev. 1999;13:2375–2387. doi: 10.1101/gad.13.18.2375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li B, Ruiz JC, Chun KT. CUL-4A is critical for early embryonic development. Mol Cell Biol. 2002;22:4997–5005. doi: 10.1128/MCB.22.14.4997-5005.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ceremuga TE, Yao X-L, McCabe JT. Vasopressin-activated calcium mobilizing (VACM-1) receptor mRNA is present in peripheral organs and the central nervous system of the laboratory rat. Endocrine Res. 2001;27:433–445. doi: 10.1081/ERC-100107867. [DOI] [PubMed] [Google Scholar]
- Ceremuga TE, Yao X-L, McCabe JT. Cullin-5 is ubiquitous in the rat brain. Neurosci Lett. 2003;345:121–125. doi: 10.1016/S0304-3940(03)00298-2. [DOI] [PubMed] [Google Scholar]
- Li B, Yang F-C, Clapp DW, Chun KT. Enforced expression of CUL-4A interferes with granulocytic differentiation and exit from the cell cycle. Blood. 2003;101:1769–1776. doi: 10.1182/blood-2002-05-1517. [DOI] [PubMed] [Google Scholar]
- Savitsky K, Bar-Shira A, Gilad S, Rotman G, Ziv Y, Vanagaite L, Tagle DA, Smith S, Uziel T, Sfez S, Ashkenazi M, Pecker I, Frydman M, Harnik R, Patanjali SR, Simmons A, Clines GA, Sartiel A, Gatti RA, Chessa L, Sanal O, Lavin MF, Jaspers NGJ, Taylor AMR, Arlett CF, Miki T, Weissman SM, Lovett M, Collins SF, Shiloh Y. A single ataxia telangiectasia gene with a product similar to PI-3 kinase. Science. 1995;268:1749–1753. doi: 10.1126/science.7792600. [DOI] [PubMed] [Google Scholar]
- Kovalev S, Mateen A, Zaika AI, O'hea BJ, Moll UM. Lack of defective expression of the ATM gene in sporadic breast cancer tissues and cell lines. Int J Oncol. 2000;16:825–831. doi: 10.3892/ijo.16.4.825. [DOI] [PubMed] [Google Scholar]
- Vorechovsky I, Rasio D, Luo L, Monaco C, Hammarstrom L, Webster ABD, Zaloudik J, Barbanti-Brodano G, James M, Russo G, Croce CM, Negrini M. The ATM gene and susceptibility to breast cancer: Analysis of 38 breast tumors reveals no evidence for mutation. Cancer Res. 1996;56:2726–2732. [PubMed] [Google Scholar]
- Fitzgerald MG, Bean JM, Hedge SR, Unsal H, MacDonald DJ, Harkin DP, Finkelstein DM, Isselbacher KJ, Haber DA. Heterozygous ATM mutations do not contribute to early onset of breast cancer. Nat Genet. 1997;15:307–310. doi: 10.1038/ng0397-307. [DOI] [PubMed] [Google Scholar]
- Singhal S, Amin KM, Kruklitis R, Delong P, Friscia ME, Litzky LA, Putt ME, Kaiser LR, Albelda SM. Alterations in Cell Cycle Genes in Early Stage lung adenocarcinoma identified by expression profiling. Cancer Biol Ther. 2003;2:291–298. doi: 10.4161/cbt.2.3.399. [DOI] [PubMed] [Google Scholar]
- Burnatowska-Hledin M, Zhao P, Capps B, Poel A, Parmelee K, Mungall C, Sharangpani A, Listenberger L. VACM-1, a cullin gene family member, regulates cellular signaling. Am J Physiol Cell Physiol. 2000;279:C266–C273. doi: 10.1152/ajpcell.2000.279.1.C266. [DOI] [PubMed] [Google Scholar]
- Van Dort C, Zhao P, Parmelee K, Capps B, Poel A, Listenberger L, Kossoris J, Wasilevich B, Murrey D, Clare P, Burnatowska-Hledin M. VACM-1, a cul 5 gene, inhibits cellular growth by a mechanism that involves MAPK and p53 signaling pathways. Am J Physiol Cell Physiol. 2003;285:C1386–C1396. doi: 10.1152/ajpcell.00338.2002. [DOI] [PubMed] [Google Scholar]
- Conaway RC, Brower CS, Conaway JW. Emerging roles of ubiquitin in transcription regulation. Science. 2002;296:1254–1258. doi: 10.1126/science.1067466. [DOI] [PubMed] [Google Scholar]
- Skowyra D, Craig K, Tyers M, Elledge SJ, Harper KW. F-box proteins are receptors that recruit phosphorylated substrates to the SCF ubiquitin-ligase complex. Cell. 1997;91:209–219. doi: 10.1016/s0092-8674(00)80403-1. [DOI] [PubMed] [Google Scholar]
- Listwan J, Marti A, Sutterluty H, Gstaiger M, Wirbelauer C, Krek W. Association of human Cul-1 and ubiquitin-conjugating enzyme CDC34 with the F-box protein p45(SKP2): evidence for evolutionary conservation in the subunit composition of the CDC34-SCF pathway. EMBO J. 1998;17:368–383. doi: 10.1093/emboj/17.2.368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyapina SA, Correll CC, Kipreos ET, Deshaies RJ. Human CUL1 forms an evolutionarily conserved ubiquitin ligase complex (SCF) with SKP1 and an F-box protein. Proc Natl Acad Sci USA. 1998;95:7451–7456. doi: 10.1073/pnas.95.13.7451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu ZK, Gervais JL, Zhang H. Human CUL-1 associates with the SKP1/SKP2 complex and regulate p21CIP1/WAF1 and cyclin D proteins. Proc Natl Acad Sci USA. 1998;95:11324–11329. doi: 10.1073/pnas.95.19.11324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrano AC, Eytan E, Hershko A, Pagano M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat Cell Biol. 1999;1:193–199. doi: 10.1038/12013. [DOI] [PubMed] [Google Scholar]
- Hatakeyama S, Kitagawa M, Nakayama K, Shirane M, Matsumoto M, Hattori K, Higashi H, Nakano H, Okumura K, Onoe K, Good RA, Nakayama K. Ubiquitin-dependent degradation of Iκ Bα is mediated by a ubiquitin ligase Skp1/Cul1/F-box protein FWD1. Proc Natl Acad Sci USA. 1999;96:3859–3863. doi: 10.1073/pnas.96.7.3859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroll M, Margottin F, Kohl A, Renard P, Durand H, Concordet JP, Bachelerie F, Arenzana-Seisdeos F, Benarous R. Inducible degradation of IkappaBalpha by the proteasome requires interaction with the f-box protein h-betaTrCP. J Biol Chem. 1999;274:7941–7945. doi: 10.1074/jbc.274.12.7941. [DOI] [PubMed] [Google Scholar]
- Latres E, Chiaur DS, Pagano M. The human F box protein beta-Trcp associates with Cul1/Skp1 complex and regulates the stability of beta-catenin. Oncogene. 1999;18:849–854. doi: 10.1038/sj.onc.1202653. [DOI] [PubMed] [Google Scholar]
- Marti A, Wirbelauer C, Scheffner M, Krek W. Interactions between ubiquitin-protein ligase SCFSKP2 and E2F-1 underlies the regulation of E2F-1 degradation. Nat Cell Biol. 1999;1:14–19. doi: 10.1038/8984. [DOI] [PubMed] [Google Scholar]
- Ohta T, Michel JJ, Schottelius AJ, Xiong Y. ROC1, a homolog of APC11, represents a family of cullin partners with an associated ubiquitin ligase activity. Mol Cell. 1999;3:535–541. doi: 10.1016/s1097-2765(00)80482-7. [DOI] [PubMed] [Google Scholar]
- Sutterluty H, Chatelain E, Marti A, Wirbelauer C, Senften M, Muller U, Krek W. p45SKP2 promotes p27KIP1 degradation and induces S phase in quiescent cells. Nat Cell Biol. 1999;1:207–214. doi: 10.1038/12027. [DOI] [PubMed] [Google Scholar]
- Suzuki H, Chiba T, Kobayashi M, Takeuchi M, Suzuki T, Ichiyama A, Ikenoue T, Omata M, Furuichi K, Tanaka K. IkappaBalpha ubiquitination is catalyzed by an SCF-like complex containing SKP-1, cullin-1, and two F-box/WD40-repeat proteins, betaTrCP1 and betaTrCP2. Biochem Biophys Res Commun. 1999;256:127–132. doi: 10.1006/bbrc.1999.0289. [DOI] [PubMed] [Google Scholar]
- Tsvetkov LM, Yeh KH, Lee SJ, Sun H, Zhang H. P27Kip1 ubiquitination and degradation is regulated by the SCFSKP2 complex through phosphorylated Thr187 in p27. Curr Biol. 1999;9:661–664. doi: 10.1016/S0960-9822(99)80290-5. [DOI] [PubMed] [Google Scholar]
- Winston JT, Strack P, Beer-Romero P, Chu CY, Elledge SJ, Harper JW. The SCFbeta-TRCP-ubiquitin ligase complex associates specifically with phosphorylated destruction motifs in IkappaBalpha and beta-catenin and stimulates IkappaBalpha ubiquitination in vitro. Genes Dev. 1999;13:270–283. doi: 10.1101/gad.13.21.2751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeda I, Ohta T, Koizumi H, Fukuda M. In vitro ubiquitination of cyclin D1 by ROC1-CUL1 and ROC1-CUL3. FEBS Lett. 2001;494:181–185. doi: 10.1016/S0014-5793(01)02343-2. [DOI] [PubMed] [Google Scholar]
- Michel JJ, Xiong Y. Human CUL-1, but not other cullin family members, selectively interacts with SKP1 to form a complex with SKP2 and cyclin A. Cell Growth Differ. 1998;19:435–449. [PubMed] [Google Scholar]
- Kamura T, Burian D, Yan Q, Schmidt SL, Lane WS, Querido E, Branton PE, Shilatifard A, Conaway RC, Conaway JW. MUF1, a novel elongin BC-interacting leucine-rich protein that can assemble with cul5 and rbx1 to reconstitute a ubiquitin ligase. J Biol Chem. 2001;276:29748–29753. doi: 10.1074/jbc.M103093200. [DOI] [PubMed] [Google Scholar]
- Ohta T, Xiong Y. Phosphorylation- and SKP1-independent in Vitro ubiquitination of E2f1 by multiple ROC-cullin ligases. Cancer Res. 2001;61:1347–1353. [PubMed] [Google Scholar]
- Furukawa M, Ohta T, Xiong Y. Activation of UBC5 ubiquitin-conjugating enzyme by the RING finger of ROC1 and assembly of active ubiquitin ligases by all cullins. J Biol Chem. 2002;277:15758–15765. doi: 10.1074/jbc.M108565200. [DOI] [PubMed] [Google Scholar]
- Lisztwan J, Imbert G, Wirbelauer C, Gstaiger M, Krek W. The von Hipple-Lindau tumor suppressor protein is a component of an E3 ubiquitin-protein ligase activity. Genes Dev. 1999;13:1822–1833. doi: 10.1101/gad.13.14.1822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwai K, Yamanaka K, Kamura T, Minato N, Conaway RC, Conaway JW, Klausner RD, Pause A. Identification of the von-Hippel-Lindau tumor-suppressor protein as an active E3 ubiquitin ligase complex. Proc Natl Acad Sci USA. 1999;96:12436–12441. doi: 10.1073/pnas.96.22.12436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brower S, Sato S, Tomomori-Sato C, Kamura T, Pause A, Stearman S, Klausner RD, Malik S, Lane WS, Sorokina I, Roeder RG, Conaway JW, Conaway RC. Mammalian mediator subunit mMED8 is an elongin BC-interacting protein that can assemble with cul2 and Rbx1 to reconstitute a ubiquitin ligase. Proc Natl Acad Sci USA. 2002;99:10353–10358. doi: 10.1073/pnas.162424199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iliopoulos O, Levy AP, Jiang C, Kaelin WG, Goldberg MA. Negative regulation of hypoxia-inducible genes by von Hippel-Lindau protein. Proc Natl Acad Sci USA. 1996;93:10595–10599. doi: 10.1073/pnas.93.20.10595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maxwell PH, Wiesener MS, Chang GW, Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER, Ratcliffe PJ. The tumor suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature. 1999;399:271–275. doi: 10.1038/20459. [DOI] [PubMed] [Google Scholar]
- Cockman ME, Masson N, Mole DR, Jaakkola P, Chang G-W, Clifford SC, Mahert ER, Pugh CW, Ratcliffe PJ, Maxwell PH. Hypoxia inducible factor-α binding and ubiquitylation by the von Hippel-Lindau tumor suppressor protein. J Biol Chem. 2000;275:25733–25741. doi: 10.1074/jbc.M002740200. [DOI] [PubMed] [Google Scholar]
- Ohh M, Park CW, Ivan M, Hoffman MA, Kim TY, Huang LE, Pavletich N, Chau V, Kaelin WG. Ubiquitination of hypoxia-inducible factor requires direct binding to the beta-domain of the von Hippel-Lindau protein. Nat Cell Biol. 2000;2:423–427. doi: 10.1038/35017054. [DOI] [PubMed] [Google Scholar]
- Kamura T, Sato S, Iwai K, Czyzyk-Krzeska M, Conaway RC, Conaway JW. Activation of HIF1α ubiquitination by a reconstituted von Hippel-Lindau (VHL) tumor suppressor complex. Proc Natl Acad Sci USA. 2000;97:10430–10435. doi: 10.1073/pnas.190332597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanimoto K, Makino Y, Pereira T, Poellinger L. Mechanism of regulation of the hypoxia-indicible factor-1 alpha by the von Hippel-Lindau tumor suppressor protein. EMBO J. 2000;19:4298–4309. doi: 10.1093/emboj/19.16.4298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaelin WG. Molecular basis of the VHL hereditary cancer syndrome. Nat Rev Cancer. 2002;2:673–682. doi: 10.1038/nrc885. [DOI] [PubMed] [Google Scholar]
- Feng H, Zhong W, Punkosdy G, Gu S, Zhou L, Seabolt EK, Kipreos ET. CUL-2 is required for the G1-to-S phase transition and mitotic chromosome condensation in Caenorhabditis elegans. Nat Cell Biol. 1999;1:486–492. doi: 10.1038/70272. [DOI] [PubMed] [Google Scholar]
- Du M, Sansores-Garcia L, Zu Z, Wu KK. Cloning and expression analysis of a novel salicylate suppressible gene, Hs-CUL-3, a member of the Cullin/Cdc53 family. J Biol Chem. 1998;273:24289–24292. doi: 10.1074/jbc.273.38.24289. [DOI] [PubMed] [Google Scholar]
- Chen L-C, Manjeshwar S, Lu Y, Moore D, Ljung B-M, Kuo W-L, Dairkee SH, Wernick M, Collins C, Smith HS. The human homologue for the Caenorhabditis elegans cul-4 gene is amplified and overexpressed in primary breast cancers. Cancer Res. 1998;58:3677–3683. [PubMed] [Google Scholar]
- Gupta A, Yang L-X, Chen L-C. Study of the G2/M cell cycle checkpoint in irradiated mammary epithelial cells overexpressing Cul-4A gene. Int J Radiat Oncol Biol Phys. 2002;52:822–830. doi: 10.1016/S0360-3016(01)02739-0. [DOI] [PubMed] [Google Scholar]
- Nag A, Bondar T, Siv S, Raychaudhuri P. The xeroderma pigmentosum group E gene product DDB2 is a specific target of cullin 4A in mammalian cells. Mol Cell Biol. 2001;21:6738–6747. doi: 10.1128/MCB.21.20.6738-6747.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shiyanov P, Nag A, Raychaudhuri P. Cullin 4A associates with the UV-damaged DNA binding protein DDB. J Biol Chem. 1999;274:35309–35312. doi: 10.1074/jbc.274.50.35309. [DOI] [PubMed] [Google Scholar]
- Zhong W, Feng H, Santiago FE, Kipreos ET. CUL-4 ubiquitin ligase maintains genome stability by restraining DNA-replication licensing. Nature. 2003;423:885–889. doi: 10.1038/nature01747. [DOI] [PubMed] [Google Scholar]
- Querido E, Blanchette P, Yan Q, Kamura T, Morrison M, Boivin D, Kaelin WG, Conaway RC, Conaway JW, Branton PE. Degradation of p53 by adenovirus E4orf6 and E1B55K proteins occurs via a novel mechanism involving a cullin-containing complex. Genes Dev. 2001;15:3104–3117. doi: 10.1101/gad.926401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harada JN, Shevchenko A, Shevchenko A, Pallas DC, Berk AJ. Analysis of the adenovirus E1B-55K-anchored proteome reveals its link to the ubiquitination machinery. J Virol. 2002;76:9194–9206. doi: 10.1128/JVI.76.18.9194-9206.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soule HD, Maloney TM, Wolman SR, Peterson WD, Brenz R, McGrath CM, Russo J, Pauley RJ, Jones RF, Brooks SC. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990;50:6075–6086. [PubMed] [Google Scholar]
- Tait L, Soule SD, Russo J. Ultrastructural and immunocytochemical characterization of an immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990;50:6087–6094. [PubMed] [Google Scholar]
- Soule HD, Vazques J, Long A, Albert S, Brennan MA. A human cell line from a pleural effusion derived from a breast carcinoma. J Natl Cancer Inst. 1973;51:1409–1416. doi: 10.1093/jnci/51.5.1409. [DOI] [PubMed] [Google Scholar]
- Cailleau R, Young R, Olive M, Reeves WJJ. Breast tumor cell lines from pleural effusions. J Natl Cancer Inst. 1973;53:661–666. doi: 10.1093/jnci/53.3.661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zinda MJ, Johnson MA, Paul JD, Horn C, Konicek BW, Lu ZH, Sandusky G, Thomas JE, Neubauer BL, Lai MT, Graff JR. AKT-1, -2, and -3 are expressed in both normal and tumor tissues of the lung, breast, prostate, and colon. Clin Cancer Res. 2001;7:2475–2479. [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Spanakis E. Problems related to the interpretation of autoradiographic data on gene expression using common constitutive transcripts as controls. Nucleic Acids Res. 1993;21:3809–3819. doi: 10.1093/nar/21.16.3809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spanakis E, Brouty-Boye D. Evaluation of quantitative variation in gene expression. Nucleic Acids Res. 1994;22:799–806. doi: 10.1093/nar/22.5.799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liew CC, Hwang DM, Fung YW, Laurenson C, Cukerman E, Tsui S, Lee CY. A catalog of genes in the cardiovascular system as identified by expressed sequence tags. Proc Natl Acad Sci USA. 1994;91:10145–10649. doi: 10.1073/pnas.91.22.10645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhumabayeva B, Diatchenko A, Chenchik A, Siebert PD. Use of SMART™-generated cDNA for gene expression studies in multiple human tumors. Biotechniques. 2001;30:158–163. doi: 10.2144/01301pf01. [DOI] [PubMed] [Google Scholar]
- Disease Profiling Array Information http://bioinfo.clontech.com/dparray